 2024, Vol. 45
2024, Vol. 45



2. 西安建筑科技大学环境与市政工程学院, 陕西省环境工程重点实验室, 西安 10055;
3. 西安水务集团黑河金盆水库管理公司, 西安 710401
2. Shaanxi Key Laboratory of Environmental Engineering, School of Environmental and Municipal Engineering, Xi'an University of Architecture and Technology, Xi'an 710055, China;
3. Xi'an Water Group Heihe Jinpen Reservoir Management Company, Xi'an 710401, China
细菌是湖库水生态环境的营养物质的主要分解者和能量储存者, 通过对水体中污染物的代谢和转化等途径来达到调节水质的作用[1 ~ 3], 对水库水体的净化有着重要作用.近年来, 随着全球气候变暖, 极端降雨指数呈增加趋势[4, 5], 暴雨径流入库不仅携带大量营养物质和外源微生物, 还会改变水体的温度和溶解氧[6, 7], 从而对水体的细菌群落结构和活性产生影响.水体中细菌群落的结构和活性决定着其在水域生态系统中生物地球化学循环的作用, 对水质恢复有着关键作用, 因此研究细菌群落在水体中的分布及变化对水资源的保护有着重要意义.已有研究对径流期间水体中微生物的变化进行了分析, Ibekwe等[8]发现暴雨径流造成的氨氮和pH的改变是影响细菌群落分布的主要环境因子;Li等[9]的研究表明降雨引起的水环境变化会对微生物异养代谢产生影响;Xin等[10]发现降雨引起的浊度、叶绿素和有机质浓度等的变化与微生物变化呈显著正相关.但上述研究对径流进入水体后细菌群落的变化、该变化对水质的影响和径流后形成的新的细菌群落结构却并未进行系统分析.本研究通过分析金盆水库暴雨径流前后细菌群落结构及活性的变化, 旨在从微生物角度为全球气候变化背景下暴雨事件对生态环境的影响提供参考.
1 材料与方法 1.1 研究区域概况黑河金盆水库(33°58′~34°03′N, 108°09′~108°12′E)作为西安市的主要水源地, 位于西安市周至县, 水库的总库容约为2亿m³, 有效库容达到1.77亿m³, 平均水深约60~95 m, 正常高水位为594 m, 汛限水位达到591 m.水库位于秦岭山中, 上游地带主要被森林覆盖, 7~10月是黑河流域的主要降水期, 降水量占全年降水量的59.5%[11], 多年平均降水量达到850.52 mm[7].
1.2 采样点设置及理化指标测定受温带大陆性气候影响, 西安降雨主要集中在夏秋季.2022年秋季降雨从10月1日持续到10月8日, 径流主要集中在10月4日入库.引水塔是金盆水库唯一的取水口, 其附近水质决定供水水质, 因此本研究选取引水塔附近的S1(图 1)作为采样点, 分别在径流前(9月29)、径流中(10月3、10月6和10月13)和径流后(11月6和11月26)这6 d采取了表层、中层和底层的水样进行污染物浓度测定, 将采取的水样用聚乙烯瓶装回实验室, 理化指标(总氮和总磷)的测定参考水和废水分析方法[12].水样预先过0.45 μm玻璃纤维滤膜, 再使用岛津TOC-L总有机碳测定仪测定溶解性有机碳(DOC), 水温、DO和pH值等水质参数, 采用多参数水质测定仪(型号:HACH Hydrolab-DS5X型)进行原位测定.

|
图 1 金盆水库采样点示意 Fig. 1 Schematic of sampling sites of Jinpen Reservoir |
三维荧光光谱的测定采用F-7000(日本-日立)荧光分光光度计.将采集的水样预先过0.45 μm的玻璃纤维滤膜, 使用1 cm石英比色皿装取水样进行荧光扫描, 以Elga-Q超纯水为空白, 发射波长(Em)的扫描范围设置为250~550 nm, 波长间隔设置为1 nm, 激发波长的(Ex)扫描范围设置为200~400 nm, 波长间隔设置为5 nm, 扫描速度为12 000 nm·min-1[13].
1.4 水体细菌群落代谢活性测定细菌群落代谢活性采用Biolog-ECO(ECO Microplate USA-MatrixTechnologies)平板法测定.将水样倒入经过高温灭菌的V型槽中, 用紫外灭菌过的八道电子移液枪吸取V型槽中的水样, 并在ECO平板中每孔接种150 μL水样.将接种过的ECO板放置在紫外灭菌过的聚乙烯盒中, 并于30℃恒温培养箱中培养240 h[14], 每间隔24 h测定一次ECO板各孔在590 nm波长处的吸光值.选取培养120 h[15]的各孔吸光值数据计算表征细菌群落代谢活性的3个参数, 包括平均颜色变化率(AWCD590nm)、McIntosh指数(U)和Shannon指数(H').
其计算方法如下:
采样孔平均颜色变化率(AWCD590nm)计算公式[16]:
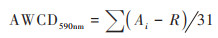
|
(1) |
式中, Ai为ECO平板中每一组的采样孔在590 nm波长下的吸光值, R为空白孔在波长590 nm下的吸光度, 若(Ai - R) ≤ 0则记为0.
McIntosh指数(U)计算公式[16]如下:
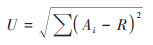
|
(2) |
式中, Ai和R的意义同上.
Shannon指数(H')计算公式[16]如下:
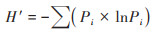
|
(3) |
式中, Pi = (Ai - R)/∑(Ai - R), 为第i孔的相对光密度与每组总的相对光密度比值.
1.5 DNA提取与高通量测序水体细菌群落结构采用高通量测序法进行分析, 样品准备:将1 L原水水样过0.22 μm的滤膜, 用灭过菌的离心管装过滤后的滤膜并于-80℃冰箱中保存.采用E.Z.N.A®soil DNA Kit基因组提取样品总DNA.采用NanoDrop2000检测提取出来的DNA的浓度和纯度[17], 以338F(ACTCCTACGGGAGGCAGCAG)和806R(GACTACHVGGGTWTCTAAT)[18, 19]为引物对16S rRNA基因的V3-V4高变区进行PCR扩增.按照97%的相似度, 采用UPARSE软件(http://drive5.com/uparse/, version 7.1)对序列进行OTU聚类, 再剔除嵌合体和单体.测序由上海美吉生物医药科技有限公司进行.
1.6 数据处理按照97%的相似度, 采用UPARSE软件(http://drive5.com/uparse/, version 7.1)对序列进行OTU聚类, 再剔除嵌合体和单体.利用RDP classifier(http://rdp.cme.msu.edu/, version 2.2)对每条序列进行物种分类注释.细菌群落多样性指数(ACE指数等)采用Mothu(version v.1.30.1)进行计算.采用冗余分析(redundancy analysis, RDA)分析细菌种类与环境因子之间的相关性.文中的柱状图和热图都采用Origin 2023进行绘制.三维荧光光谱数据采用MATLAB 2018a软件中的DOMFlour工具箱进行分析, 首先识别并修正去除样品荧光矩阵的瑞利散射和拉曼峰散射, 再进行平行因子分析, 最后采用折半分析来验证结果的可靠性[20].
2 结果与讨论 2.1 径流前后水质变化图 2为暴雨径流前后DO、水温和浊度的时空变化.暴雨径流入库后, 表层和中层水温分别降至16℃和13℃, 而底层水温则上升了约1℃.溶解氧的变化则更明显, 表层水体ρ(DO)一直保持在8 mg·L-1左右, 中层水体ρ(DO)由5.1 mg·L-1上升至9.1 mg·L-1, 底层水体ρ(DO)由0.3 mg·L-1上升至8.8 mg·L-1. Park等[21]的研究发现径流对水体分层结构的影响与径流来水浊度以及水库当时的分层状态有关.本研究中, 径流期间水体中层的浊度大幅上升, 径流来水以中部潜流的方式进入水库, 缩小了水体中层和表层的温差, 同时使中层水体DO浓度明显增加, 对水体的缺氧情况有所缓解[22].径流后, 随着气温下降, 表层水温不断降低, 水体表底温差不断减小直至完全混合, 暴雨径流在一定程度上促进了水体混合.

|
图 2 2022年10月6日暴雨径流前后采样点水温、溶解氧和浊度垂向变化 Fig. 2 Vertical variations in water temperature, dissolved oxygen, and turbidity in the sampling sites before and after the storm runoff on October 6, 2022 |
径流入库不仅使水体水温和DO浓度发生变化, 同时也携带大量污染物进入水体(图 3), 径流入库后, 中层水体ρ(DOC)、ρ(TN)和ρ(TP)分别上升至5.176、1.65和0.161 mg·L-1, 底层水体ρ(TN)和ρ(TP)浓度则分别升高至1.723 mg·L-1和0.193 mg·L-1;中层水体TN的峰值出现在10月3日, 而DOC和TP都出现在10月6日.之前对金盆水库在径流期间水质响应的研究[23]中也发现, 不同水层TN、TP和DOC会对径流有不同的响应, 且中层和底层水体变化更明显.径流后, 水体中TN、TP和DOC浓度相较径流中有所降低, 且TP削减速度显著高于DOC和TN的削减速度, 原因是TP主要以颗粒态为主, TP会随着大颗粒快速沉降而减少.径流后一个月(11月6日), 污染物浓度仍比径流前的浓度高, 说明暴雨径流对水库水质有显著影响, 且持续时间在一个月以上, 水质短期内无法恢复.此外, 大量污染物也会随着颗粒沉降到底部沉积物中, 使内源污染负荷增大.

|
图 3 径流前后DOC、TN和TP浓度变化 Fig. 3 Variations in DOC, TN, and TP before and after storm runoff |
通过对径流前、径流中和径流进入水库后一个月的采样点表层、中层和底层水样进行高通量测序, 共获得704 441条优化序列, 测序的结果覆盖率都大于99%.表 1列出暴雨径流前后各层水体的OTUs数量和α多样性指数变化.各层水体的OTUs数量在径流刚结束时都有明显上升, 底层上升幅度显著高于表层和中层水体. α多样性指数中[22 ~ 25], Shannon指数值越大, 则代表群落多样性越高, 而Simpson指数值则越小, 代表群落多样性越高[26];ACE指数和Chao指数数值越大, 则表示细菌群落丰富度越高[14, 27].如表 1所示, 在径流前, 中层水体的Shannon指数高于底层和表层, 说明中层水体的细菌群落多样性较高, 其稳定性也越好.径流进入水体后, 各层水体的Shannon指数都明显升高, 说明径流进入水库后, 给水体带入更多种类的菌种, 使水体的细菌群落多样性显著增加.虽然径流期间, 中层水体的DO和营养盐浓度变化程度高于其他水体, 但因为中层水体细菌群落稳定性高于其他水层, 即抗干扰能力更强, 所以径流进入水体后, 其群落多样性变化程度小于其他水层.群落丰富度指数ACE和Chao也有相同变化趋势, 且在径流一个月以后, 水体的OTUs和α多样性指数虽然有所降低, 但仍然高于径流前.根据Møller等[28]的研究, 细菌门的相对丰度和其多样性有关, 径流期间细菌多样性和丰富度的增加, 使细菌群落在水生态环境中有更强的竞争力, 对径流后污染物的降解和水质的恢复有重要作用.
|
|
表 1 暴雨径流前后水体细菌群落多样性指数变化 Table 1 Temporal and spatial variations in bacterial community diversity index in the sampling sites before and after the storm runoff |
通过高通量测序分析, 共得到3 247条OTUs, 分别属于42个门、123个纲、299个目、474个科、879个属和1 632个种.图 4展示了采样期间门水平上细菌群落结构变化, 占比前5的是变形菌门(Proteobacteria)、放线菌门(Actinobacteriota)、蓝藻门(Cyanobacteria)、酸杆菌门(Acidobacteriota)和拟杆菌门(Bacteroidota), 这和之前的研究[14]相符.径流前, 表层水体优势菌是蓝藻门, 占比70.7%, 而中层和底层水体的优势菌则是变形菌门, 分别占比32.3%和40.2%.径流进入水体后, 表层的蓝藻门占比由70.7%降至49.3%, 而变形菌门和放线菌门占比则分别上升至21.5%和11.5%.中层和底层的水体则是变形菌门和拟杆菌门占比上升较多, 中层的变形菌门和拟杆菌门占比分别上升至43.4%和20.2%, 底层则分别上升至52.8%和14.4%.变形菌对温度变化具有较高的敏感度, 且其相对丰度与温度呈负相关[29], 径流期间水温降低, 所以变形菌门显著上升.此外, 中层水体的厚壁菌门(Firmicutes)由径流前的0.5%上升至1.6%, 厚壁菌门是沉积物中的优势菌门[30], 中层水体在径流期间浊度显著提高, 河道沉积物中的微生物随着径流冲刷进入水库, 所以厚壁菌门有所上升.径流后一个月, 各层水体门水平细菌群落结构差异减小, 在径流中变化明显的菌群都没有恢复到径流前的水平, 而是形成一个新的结构.可能是因为径流后, 水体达到混合状态, 而不是恢复到分层状态, 所以细菌群落结构会受自然混合和径流的双重影响.

|
1.径流前-表层, 2.径流前-中层, 3.径流前-底层, 4.径流中-表层, 5.径流中-中层, 6.径流中-底层, 7.径流后-表层, 8.径流后-中层, 9.径流后-底层 图 4 暴雨径流前后水体细菌群落结构变化 Fig. 4 Changes in bacterial community in the sampling sites before and after the storm runoff |
进一步分析属水平细菌群落组成, 图 5为属水平占比前10%的物种分布, 占比较高的是蓝藻门的Chloroplast和Cyanobium_PCC-6307.在径流进入水体后, 表层水体的Cyanobium_PCC-6307的丰度由21 827降至3 326个OUT, Chloroplast则由8 335上升至16 226个OUT.通过共生网络分析技术进一步探究径流期间水体中相对丰度位于前40的属水平细菌群落的相互关系(图 6), 菌种的丰度由节点的大小表示, 节点之间的条带越粗代表两类细菌的相互影响关系越密[31].模块Ⅰ中的属水平菌种丰度较高, 且相互之间的影响较为密切.模块Ⅰ中丰度较高的细菌是CL500-29_marine_group、hgcI_clade和Methylobacter;CL500-29_marine_group和hgcI_clade, 广泛存在于湖库水体中, 其中CL500-29_marine_group可以在有氧的条件下降解DOC, hgcI_clade则对碳水化合物和含氮化合物具有较强的吸收能力[32], 径流期间这些菌种较高的丰度为后续污染物的生物降解提供了条件.模块Ⅰ中的菌种之间既有负相关也有正相关, 正负交互作用, 表明菌种之间竞争或互惠等共生模式对金盆水库细菌群落结构起着重要作用, 对于水库水质安全和生态稳定也有着重要作用.

|
1.径流前-表层, 2.径流前-中层, 3.径流前-底层, 4.径流中-表层, 5.径流中-中层, 6.径流中-底层, 7.径流后-表层, 8.径流后-中层, 9.径流后-底层 图 5 属水平细菌群落结构变化 Fig. 5 Changes in bacterial community structureat genus level |

|
图 6 基于Spearman显著性分析的水体细菌共生网络分析(P < 0.01) Fig. 6 Co-occurrence network of bacterial community based on Spearman significance analysis (P < 0.01) |
表征细菌群落代谢活性的3个参数中, AWCD590 nm数值越大, 则表示微生物对碳源利用程度及微生物活性越高[33], McIntosh指数(U)和Shannon指数(H')这两个参数均表征细菌群利用碳源能力的多样性, 数值越大表明细菌群落功能多样性越高.表 2列出暴雨径流前后水体微生物的AWCD590 nm及多样性指数变化.暴雨径流进入水体后, AWCD590 nm值上升至1以上, McIntosh指数(U)上升至9以上, Shannon指数(H')上升幅度则略小, 说明径流入库使水体菌群代谢活性及功能多样性显著增加.对比不同水层发现, 中层水体的细菌群落活性提高程度显著高于表层和底层水体, 根据径流期间水体水质变化, 中层水体在径流期间DO上升了4 mg·L-1, 上升幅度高于表层和底层, 虽然DO和水温都会影响微生物活性[34], 水体温度有所下降, 但下降幅度较小.
|
|
表 2 细菌群落功能多样性指数变化1) Table 2 Variations in functional diversity indices of bacterial communities |
图 7为细菌对各类碳源的利用程度, 径流前水体细菌群落对各类碳源利用程度都较低, 各层水体的细菌群落对氨基酸类、酯类和醇类的利用程度高于其他3种碳源.径流入库后细菌群落对六大碳源的利用程度都有明显提高, 且在径流一个月后, 水体细菌活性及对碳源的利用程度仍然维持较高水平.虽然径流入库主要使中层水体DO浓度提升, 底层仍处于厌氧状态, 但中层和底层水体细菌的活性及对碳源利用程度都明显升高, 中层水体碳源利用程度提高率略高于底层水体, 主要因为中层和底层水体营养盐都显著升高, 虽然增加的营养盐如DOC主要为外源性, 不易被细菌分解利用[35], 但也有研究认为[36], 在贫营养水体中, 外源碳的输入是对水体碳源的补给, 外源碳对于贫营养水体中细菌代谢生长的促进作用不容忽视.在径流一个月后, 细菌对这些碳源的利用程度仍然较高, 说明径流入库对细菌的代谢活性的影响比较持久.细菌群落所偏好的碳源种类也有所变化, 径流前水体细菌群落对氨基酸类、酯类和醇类利用程度较高, 而径流后, 细菌群落则对糖类、氨基酸类和酯类利用程度较高.研究者普遍认为[37], 外源有机碳中芳香化程度较高, 而内源有机碳则含有更多的氨基酸, 径流前水体中有机碳主要为内源有机碳, 因此水体细菌群落偏好的碳源为氨基酸, 而径流入库后, 外源性有机物的输入促进水体细菌的呼吸和代谢, 呼吸过程对有机物如糖类的利用率则会增加, 因此径流后细菌偏好的碳源为糖类和氨基酸类.

|
1.径流前-表层, 2.径流前-中层, 3.径流前-底层, 4.径流中-表层, 5.径流中-中层, 6.径流中-底层, 7.径流后-表层, 8.径流后-中层, 9.径流后-底层 图 7 采样点水体细菌群落对六大碳源的利用程度 Fig. 7 Relative use efficiency of six carbon sources by the microbial community at sampling sites |
在暴雨径流输入水库的众多污染物中, 高含量的溶解性有机质(DOM)对水质威胁较大, 例如产生气味以及饮用水处理过程中产生消毒副产物[38], 而且DOM对于维持微生物生长具有重要作用[39], 微生物也参与了DOM转化的两条途径:无机化和腐殖化[40].因此, 分析径流前后DOM的变化对研究暴雨径流期间微生物种群结构的变化十分重要.
通过平行因子分析算法对金盆水库径流前后水样进行三维荧光分析, 共获得4种DOM组分, 4个组分的光谱图见图 8.从中可知解析的4种组分分别为C1(Ex/Em=270/460 nm)和C2组分(Ex/Em =240/436 nm)为陆源腐殖质[41], C3(Ex/Em =300/397 nm)为微生物腐殖质[42], C4(Ex/Em =275/338 nm)为来自藻类降解的类色氨酸基团.图 9为4个组分的荧光强度变化, 径流期间陆源腐殖质C1和C2的荧光强度升高程度高于另外两种组分.金盆水库位于秦岭山中, 秋季大量枯萎植物落在土壤表面, 汛期表面土层中的有机质被径流裹挟, 大量腐殖质随着径流进入水库[41], 所以水库中陆源腐殖质占比显著升高.径流后, C1~C4组分荧光强度都有不同程度下降, C1、C2、C3和C4的削减率分别为28.6%、46.1%、42.8%和59.7%.主要是因为C1、C2和C3是大分子的腐殖质, 难以被水生生物直接利用[43], 这类大分子的有机质含有比较多的可以吸收太阳光的显色基团, 会先通过化学降解转化为小分子有机质再被微生物利用.类蛋白质相较于类腐殖质更不稳定, 微生物会优先降解类蛋白.根据2.3和2.4节的结果可知, 径流期间, 变形菌和拟杆菌门都显著上升, 有研究发现变形菌和拟杆菌是太湖水体中类蛋白组分的主要分解者[44], 且细菌代谢活性及对碳源的利用程度在径流期间也有较大幅度提升, 所以径流后一个月, C4的削减率最高.

|
图 8 金盆水库4种DOM荧光组分光谱 Fig. 8 Fluorescence spectra of four DOM components in Jinpen Reservoir |

|
图 9 4种DOM组分荧光强度变化 Fig. 9 Variations in fluorescence intensity of four DOM components |
已有研究表明[45], 水体细菌群落结构在不同时期受不同环境因子影响.陈兆进等[46]在对丹江口水库细菌群落的研究中表明不同环境因子对不同的细菌有着不同的影响.本研究将水体占比前5的门水平细菌群落与环境因子进行冗余分析(图 10), 从图 10可以看出, 不同时期不同水层的细菌群落结构受不同水质指标调控.径流前和径流中, 表层细菌群落结构主要受DO和T影响, 且DO和T对蓝藻门具有较大影响, 而中层和底层水体的细菌群落主要受氮、磷、DOC和浊度的影响, 其中浊度、DOC和TP对径流中的细菌群落结构影响较大, 且和变形菌门、拟杆菌门和放线门呈正相关.进一步将细菌群落和DOM不同组分进行冗余分析, 如图 11所示, DOM中的C1和C3组分对拟杆菌门、变形菌门和放线菌门的影响较大, 且呈显著相关.根据2.1和2.4节的分析结果, 径流期间, 营养盐浓度、DOM各组分含量和浊度都大幅升高, 上述菌群占比也提升, 与相关性图相符.径流后一个月, 水体细菌群落并没有恢复到径流前, 而是形成新的结构, 且水体各层差异减小, 从图中可以明显看出, 径流后不同水层的菌群受环境因子的影响程度相似.

|
虚线表示环境因子, 实线表示细菌群落 图 10 水体细菌群落结构与水质参数之间的冗余分析 Fig. 10 Redundancy analysis of bacterial community structure and water quality parameters |

|
虚线表示机质组分, 实线表示细菌群落 图 11 水体细菌群落结构与4种DOM组分的冗余分析 Fig. 11 Redundancy analysis of bacterial community structure and four DOM components |
(1)暴雨径流入库缩小了水体表底温差, 提高了水体溶解氧和污染物浓度.径流后一个月, 污染物浓度和浊度都有所下降, 但污染物浓度仍高于径流前, 暴雨径流造成水库污染的周期较长.
(2)径流期间细菌群落丰度指数ACE及多样性指数Shannon都比径流前高, 对碳源的利用程度也显著提高, 细菌偏好的碳源转变为糖类和氨基酸类.径流后一个月, 细菌的多样性和丰富度及代谢活性有所下降, 但仍远高于径流前.
(3)细菌群落结构在径流期间发生较大改变, 蓝藻门占比下降, 变形菌门和拟杆菌门占比上升, 这两者的丰度之和由径流前的46.7%上升至67.1%.径流后一个月, 水体细菌群落结构并没有恢复到径流前, 变形菌门成为优势菌群, 且水体各层细菌群落结构差异缩小.
(4)根据冗余分析可知, 不同时期不同水层的细菌群落结构受不同环境因子影响, 径流前和径流中, 水体表层细菌群落主要受DO和T影响, 中层和底层的细菌则受浊度、DOM、TN和TP影响较大.
| [1] |
朱亮, 赵林多, 刘钢, 等. 大运河沉积微生物群落结构特征分析[J]. 中国矿业大学学报, 2010, 39(2): 295-301. Zhu L, Zhao L D, Liu G, et al. Research on seasonal and spatial shifts in river sediments microbial community structure[J]. Journal of China University of Mining & Technology, 2010, 39(2): 295-301. |
| [2] | Yang N, Li Y, Zhang W L, et al. Cascade dam impoundments restrain the trophic transfer efficiencies in benthic microbial food web[J]. Water Research, 2020, 170. DOI:10.1016/j.watres.2019.115351 |
| [3] | Shade A, Read J S, Youngblut N D, et al. Lake microbial communities are resilient after a whole-ecosystem disturbance[J]. The ISME Journal, 2012, 6(12): 2153-2167. DOI:10.1038/ismej.2012.56 |
| [4] |
龙妍妍, 范广洲, 段炼, 等. 中国近54年来夏季极端降水事件特征研究[J]. 气候与环境研究, 2016, 21(4): 429-438. Long Y Y, Fan G Z, Duan L, et al. A study on the characteristics of summertime extreme precipitation events over China in recent 54 years[J]. Climatic and Environmental Research, 2016, 21(4): 429-438. |
| [5] | Semenza J C. Cascading risks of waterborne diseases from climate change[J]. Nature Immunology, 2020, 21(5): 484-487. DOI:10.1038/s41590-020-0631-7 |
| [6] | Fan C W, Kao S J. Effects of climate events driven hydrodynamics on dissolved oxygen in a subtropical deep reservoir in Taiwan[J]. Science of the Total Environment, 2008, 393(2-3): 326-332. DOI:10.1016/j.scitotenv.2007.12.018 |
| [7] |
黄诚, 黄廷林, 李扬, 等. 金盆水库暴雨径流时空演变过程及水质评价[J]. 环境科学, 2021, 42(3): 1380-1390. Huang C, Huang T L, Li Y, et al. Temporal and spatial evolution of storm runoff and water quality assessment in Jinpen Reservoir[J]. Environmental Science, 2021, 42(3): 1380-1390. |
| [8] | Ibekwe A M, Ma J C, Murinda S E. Bacterial community composition and structure in an Urban River impacted by different pollutant sources[J]. Science of the Total Environment, 2016, 566-567: 1176-1185. DOI:10.1016/j.scitotenv.2016.05.168 |
| [9] | Li Y X, Deng K K, Lin G J, et al. Effects of physiologic activities of plankton on CO2 flux in the Three Gorges Reservoir after rainfall during algal blooms[J]. Environmental Research, 2023, 216. DOI:10.1016/j.envres.2022.114649 |
| [10] | Xin Y, Zhang J Y, Lu T D, et al. Response of prokaryotic, eukaryotic and algal communities to heavy rainfall in a reservoir supplied with reclaimed water[J]. Journal of Environmental Management, 2023, 334. DOI:10.1016/j.jenvman.2023.117394 |
| [11] |
马卫星. 暴雨径流潜流对峡谷分层型水源水库水质影响与水质原位改善[D]. 西安: 西安建筑科技大学, 2015. Ma W X. The effects of rainstorm flow undercurrent on water quality and the in-situ water quality improvement of a canyon-shaped, stratified, source water reservoir[D]. Xi'an: Xi'an University of Architecture and Technology, 2015. |
| [12] | 国家环境保护总局. 水和废水监测分析方法[M]. (第四版). 北京: 中国环境科学出版社, 2002. |
| [13] |
林子深, 黄廷林, 李凯, 等. 李家河水库上游水体溶解性有机物组分与来源[J]. 中国环境科学, 2020, 40(3): 1275-1283. Lin Z S, Huang T L, LI K, et al. Dissolved organic matter components and sources in upstream of Lijiahe Reservoir[J]. China Environmental Science, 2020, 40(3): 1275-1283. DOI:10.3969/j.issn.1000-6923.2020.03.039 |
| [14] |
高悦, 黄廷林, 李楠, 等. 人工诱导水库分层水体提前混合对细菌群落结构和代谢活性的影响[J]. 环境科学, 2022, 43(5): 2624-2632. Gao Y, Huang T L, Li N, et al. Effect of induced mixing on bacterial community structure and metabolic activity in reservoir[J]. Environmental Science, 2022, 43(5): 2624-2632. |
| [15] | Bracken C L, Hendricks C W, Harding A K. Apparent bias in river water inoculum following centrifugation[J]. Journal of Microbiological Methods, 2006, 67(2): 304-309. DOI:10.1016/j.mimet.2006.04.003 |
| [16] |
闫苗苗, 陈胜男, 黄廷林, 等. 分层型水库藻类垂向演替的水质与细菌种群调控[J]. 环境科学, 2020, 41(7): 3285-3296. Yan M M, Chen S N, Huang T L, et al. Water quality and bacterial population driving mechanism of algae vertical succession in stratified reservoir[J]. Environmental Science, 2020, 41(7): 3285-3296. |
| [17] |
程豹, 望雪, 徐雅倩, 等. 澜沧江流域浮游细菌群落结构特征及驱动因子分析[J]. 环境科学, 2018, 39(8): 3649-3659. Cheng B, Wang X, Xu Y Q, et al. Bacterioplankton community structure in the Lancang River basin and the analysis of its driving environmental factors[J]. Environmental Science, 2018, 39(8): 3649-3659. |
| [18] |
王鹏, 陈波, 李传琼, 等. 赣江南昌段丰水期细菌群落特征[J]. 中国环境科学, 2016, 36(8): 2453-2462. Wang P, Chen B, Li C Q, et al. Bacterial communities in Nanchang section of the Ganjiang River in wet seaon[J]. China Environmental Science, 2016, 36(8): 2453-2462. DOI:10.3969/j.issn.1000-6923.2016.08.027 |
| [19] |
钱玮, 张济凡, 张铭连, 等. 太湖湖滨湿地浮游细菌群落结构及时间动态[J]. 基因组学与应用生物学, 2018, 37(12): 5325-5331. Qian W, Zhang J F, Zhang M L, et al. Community structure and temporal dynamics of Bacterioplankton in lakeside wetland of Tai Lake[J]. Genomics and Applied Biology, 2018, 37(12): 5325-5331. |
| [20] | Stedmon C A, Bro R. Characterizing dissolved organic matter fluorescence with parallel factor analysis: a tutorial[J]. Limnology and Oceanography: Methods, 2008, 6(11): 572-579. DOI:10.4319/lom.2008.6.572 |
| [21] | Park H, Chung S, Cho E, et al. Impact of climate change on the persistent turbidity issue of a large dam reservoir in the temperate monsoon region[J]. Climatic Change, 2018, 151(3): 365-378. |
| [22] |
吕晓龙, 黄廷林, 李楠, 等. 暴雨径流潜流过程及其对分层水库水质的影响[J]. 中国环境科学, 2019, 39(7): 3064-3072. Lü X L, Huang T L, Li N, et al. Impact of stormwater inflow on water quality of stratified source reservoir[J]. China Environmental Science, 2019, 39(7): 3064-3072. DOI:10.3969/j.issn.1000-6923.2019.07.044 |
| [23] |
曾康, 黄廷林, 马卫星, 等. 金盆水库汛期高浊水径流的潜入及热分层水体水质响应[J]. 中国环境科学, 2015, 35(9): 2778-2786. Zeng K, Huang T L, Ma W X, et al. Water-quality responses of the intrusion of high-turbidity runoff to the thermal stratified Jin-pen Reservoir during flood season[J]. China Environmental Science, 2015, 35(9): 2778-2786. DOI:10.3969/j.issn.1000-6923.2015.09.033 |
| [24] | Zhang H H, Chen S N, Huang T L, et al. Vertical distribution of bacterial community diversity and water quality during the reservoir thermal stratification[J]. International Journal of Environmental Research and Public Health, 2015, 12(6): 6933-6945. DOI:10.3390/ijerph120606933 |
| [25] | Kim O S, Imhoff J F, Witzel K P, et al. Distribution of denitrifying bacterial communities in the stratified water column and sediment-water interface in two freshwater lakes and the Baltic Sea[J]. Aquatic Ecology, 2011, 45(1): 99-112. DOI:10.1007/s10452-010-9335-7 |
| [26] |
商潘路, 陈胜男, 黄廷林, 等. 深水型水库热分层诱导水质及真菌种群结构垂向演替[J]. 环境科学, 2018, 39(3): 1141-1150. Shang P L, Chen S N, Huang T L, et al. Vertical distribution of fungal community composition and water quality during the deep reservoir thermal stratification[J]. Environmental Science, 2018, 39(3): 1141-1150. |
| [27] |
杜宛璘, 孙金辉, 麦永湛, 等. 珠江下游浮游细菌群落结构的时空分布特征[J]. 湖泊科学, 2020, 32(2): 380-394. Du W L, Sun J H, Mai J Z, et al. Spatial and temporal distribution characteristics of bacterioplankton community structure in the downstream of Pearl River[J]. Journal of Lake Sciences, 2020, 32(2): 380-394. |
| [28] | Møller A K, Søborg D A, Abu Al-Soud W, et al. Bacterial community structure in High-Arctic snow and freshwater as revealed by pyrosequencing of 16S rRNA genes and cultivation[J]. Polar Research, 2013, 32(1). DOI:10.3402/polar.v32i0.17390 |
| [29] | Eronen-Rasimus E, Lyra C, Rintala J M, et al. Ice formation and growth shape bacterial community structure in Baltic Sea drift ice[J]. FEMS Microbiology Ecology, 2015, 91(2): 1-13. |
| [30] |
王跃. 湖库沉积物酶活性和功能菌群结构研究[D]. 西安: 西安建筑科技大学, 2019. Wang Y. Enzyme activity and functional microbial community structure in sediment from lakes and reservoirs[D]. Xi'an: Xi'an University of Architecture and Technology, 2019. |
| [31] | Zheng B H, Chen Z J, Li Y Y, et al. Structural characteristics and driving factors of the planktonic eukaryotic community in the Danjiangkou Reservoir, China[J]. Water, 2020, 12(12). DOI:10.3390/W12123499 |
| [32] | dos Reis M C, Bagatini I L, de Oliveira Vidal L, et al. Spatial heterogeneity and hydrological fluctuations drive bacterioplankton community composition in an Amazon floodplain system[J]. PLoS One, 2020, 15(1). DOI:10.1371/journal.pone.0228721 |
| [33] | Zhang H H, Huang T L, Chen S N, et al. Microbial community functional diversity and enzymatic activity in the sediments of drinking water reservoirs, Northwest China[J]. Desalination and Water Treatment, 2014, 52(7-9): 1608-1614. DOI:10.1080/19443994.2013.827383 |
| [34] | Wang J T, Fan H B, He X J, et al. Response of bacterial communities to variation in water quality and physicochemical conditions in a river-reservoir system[J]. Global Ecology and Conservation, 2021, 27. DOI:10.1016/j.gecco.2021.e01541 |
| [35] | Moran M A, Hodson R E. Bacterial production on humic and nonhumic components of dissolved organic carbon[J]. Limnology and Oceanography, 1990, 35(8): 1744-1756. DOI:10.4319/lo.1990.35.8.1744 |
| [36] |
吴庆龙, 邢鹏, 李化炳, 等. 草藻型稳态转换对湖泊微生物结构及其碳循环功能的影响[J]. 微生物学通报, 2013, 40(1): 87-97. Wu Q L, Xing P, Li H B, et al. Impacts of regime shift between phytoplankton and macrophyte on the microbial community structure and its carbon cycling in lakes[J]. Microbiology China, 2013, 40(1): 87-97. |
| [37] | Shen Y, Fichot C G, Liang S K, et al. Biological hot spots and the accumulation of marine dissolved organic matter in a highly productive ocean margin[J]. Limnology and Oceanography, 2016, 61(4): 1287-1300. DOI:10.1002/lno.10290 |
| [38] | Tomlinson A, Drikas M, Brookes J D, et al. The role of phytoplankton as pre-cursors for disinfection by-product formation upon chlorination[J]. Water Research, 2016, 102: 229-240. DOI:10.1016/j.watres.2016.06.024 |
| [39] | Natchimuthu S, Sundgren I, Gålfalk M, et al. Spatiotemporal variability of lake pCO2 and CO2 fluxes in a hemiboreal catchment[J]. Journal of Geophysical Research: Biogeosciences, 2017, 122(1): 30-49. DOI:10.1002/2016JG003449 |
| [40] |
赵凌云. 金盆水库DOM变化规律及混合充氧削减效果研究[D]. 西安: 西安建筑科技大学, 2021. Zhao L Y. Study on the variation of DOM characteristics and the in situ reduction effect by mixing and oxygenation in Jinpen Reservoir[D]. Xi'an: Xi'an University of Architecture and Technology, 2021. |
| [41] | Shi Y, Zhang L Q, Li Y P, et al. Influence of land use and rainfall on the optical properties of dissolved organic matter in a key drinking water reservoir in China[J]. Science of the Total Environment, 2020, 699. DOI:10.1016/j.scitotenv.2019.134301 |
| [42] | Sankar M S, Dash P, Singh S, et al. Effect of photo-biodegradation and biodegradation on the biogeochemical cycling of dissolved organic matter across diverse surface water bodies[J]. Journal of Environmental Sciences, 2019, 77: 130-147. DOI:10.1016/j.jes.2018.06.021 |
| [43] | Skoog A, Wedborg M, Fogelqvist E. Decoupling of total organic carbon concentrations and humic substance fluorescence in a an extended temperate estuary[J]. Marine Chemistry, 2011, 124(1-4): 68-77. DOI:10.1016/j.marchem.2010.12.003 |
| [44] | Zhang W, Zhou Y Q, Jeppesen E, et al. Linking heterotrophic bacterioplankton community composition to the optical dynamics of dissolved organic matter in a large eutrophic Chinese lake[J]. Science of the Total Environment, 2019, 679: 136-147. DOI:10.1016/j.scitotenv.2019.05.055 |
| [45] |
刘睿, 吴巍, 周孝德, 等. 渭河浮游细菌群落结构特征及其关键驱动因子[J]. 环境科学学报, 2017, 37(3): 934-944. Liu R, Wu W, Zhou X D, et al. Bacterioplankton community structure in Weihe River and its relationship with environmental factors[J]. Acta Scientiae Circumstantiae, 2017, 37(3): 934-944. |
| [46] |
陈兆进, 丁传雨, 朱静亚, 等. 丹江口水库枯水期浮游细菌群落组成及影响因素研究[J]. 中国环境科学, 2017, 37(1): 336-344. Chen Z J, Ding C Y, Zhu J Y, et al. Community structure and influencing factors of bacterioplankton during low water periods in Danjiangkou Reservoir[J]. China Environmental Science, 2017, 37(1): 336-344. |