 2024, Vol. 45
2024, Vol. 45

2. 呼和浩特市生态环境局, 呼和浩特 010018;
3. 内蒙古自治区水资源保护与利用重点实验室, 呼和浩特 010018;
4. 黄河流域内蒙段水资源与水环境综合治理自治区协同创新中心, 呼和浩特 010018;
5. 内蒙古农业大学生命科学学院, 呼和浩特 010018
 , HAN Guo-qiang2 , ZHANG Sheng-wei1,3,4
, HAN Guo-qiang2 , ZHANG Sheng-wei1,3,4 , ZHAO Hong-bin5 , LIU Min-min1 , LIN Xi1 , WANG Shuai1
, ZHAO Hong-bin5 , LIU Min-min1 , LIN Xi1 , WANG Shuai1
2. Hohhot Ecological Environment Bureau, Hohhot 010018, China;
3. Inner Mongolia Autonomous Region Key Laboratory of Water Resources Protection and Utilization, Hohhot 010018, China;
4. Autonomous Region Collaborative Innovation Center for Integrated Management of Water Resources and Water Environment in the Inner Mongolia Reaches of the Yellow River, Hohhot 010018, China;
5. College of Life Sciences, Inner Mongolia Agricultural University, Hohhot 010018, China
草地生态系统是陆地生态系统中分布面积最广且最主要的组成部分之一, 其固碳潜力巨大, 对全球碳通量和气候变化具有重要影响[1].随着人口增长和经济发展, 人类活动加剧, 造成土地荒漠化严重, 草原退化等问题, 以上问题会进一步导致土壤物理结构被破环等, 使区域生态系统的“碳汇”能力下降, 温室气体释放增加, 促使全球变暖[2, 3].不同的草地管理模式对地上、地下生物量以及碳储量的影响不同, 草地上普遍存在的放牧会直接影响植物生长, 改变草地物种组成和生存环境[4, 5].而植被在减少温室气体、调节碳平衡和缓解气候变化方面发挥着重要的作用, 不仅是生态环境变化至关重要的元素, 也是维持土壤碳储量稳定乃至维持区域碳库稳定的关键[6, 7].因此, 研究植被特征和植被碳储量的变化对草地恢复及维持生态系统稳定具有重要意义.
禁牧可以显著提高草地生产力[8], 改善草地群落结构[9], 但有研究表明, 长期禁牧导致凋落物增多, 对草地植物种群的更新具有抑制作用, 导致物种丰富度和多样性下降、群落组成渐趋单一化[10].而适度放牧可改善未踩踏草地的光照、土壤水分和养分, 促进植被根系生长, 改善土壤碳氮固持及持水能力, 从而有利于植被碳累积[11 ~ 13].也有研究表明放牧会降低植被高度和群落覆盖度, 且植被碳储量随着放牧强度增加而逐渐减少[14].影响植被碳汇的非生物因子包括地理空间和气候因素等[15].其中, 气候条件是草地生物量碳库的主要影响因素, 温度是影响植被碳库的关键要素, 其通过影响土壤水分进而对地上生物量造成直接影响[16, 17].也有研究表明, 植被碳利用效率随着降水量的增加而增加[18].
以往的研究表明, 植被碳储量由于生物量空间异质性限制和研究方法的差异等方面因素, 使我国草地植被碳储量估算结果仍有很大差别. 对于草地植被碳储量的研究, 有基于模型模拟估算碳储量的方法[19, 20];还有直接测定土壤和植物样品中的碳含量的方法[21, 22].而由于野外实地采样以及获取地下生物量数据较为困难, 许多植被碳储量的研究缺乏地下根系碳储量、枯落物碳储量等数据. 因此, 国际上通常采用0.45或0.5换算系数进行碳储量换算, 导致不同植被碳储量资料之间存在显著差异[23].本研究通过大量野外实地采样测量了植被有机含碳率, 并通过结构方程模型评价气候因子、土壤因子和地理因子对植被碳储量的直接和间接影响, 以期为草地碳储量估算和草地恢复提供重要依据.
1 材料与方法 1.1 研究区概况研究区位于内蒙古草原, 沿干旱梯度自东向西依次分布着草甸草原、典型草原和荒漠草原, 自东向西布设了18处共22个观测站点作为研究对象, 其中包括3个草甸草原、6个荒漠草原和9个典型草原监测点;其中实施禁牧管理的有4个站点:草甸草原和荒漠草原各1个, 禁牧时间分别是3 a和4 a;典型草原有2个, 禁牧时间分别是2 a和23 a, 具体位置见图 1.此外, 草甸草原的乌拉盖站点属于适度放牧, 其余站点属于过度放牧.

|
图 1 研究区土地利用及站点位置示意 Fig. 1 Schematic of land use and site locations in the study area |
研究区以温带大陆性气候为主, 冬季长春季短, 夏季热秋季凉. 年平均气温在3.0~6.7℃, 由北向南, 呈递减趋势. 降水自东向西由500 mm递减至150 mm左右, 少而不匀且集中在夏秋季. 蒸发量大部分地区高于1 200 mm, 年平均风速在3 m·s-1左右. 光能资源非常充沛, 太阳辐射量从东北向西南递增. 植物种类丰富, 域内有植物种类约100多种. 本文草甸草原试验区土壤类型以栗钙土为主, 试验区常见优势种多为:大针茅(Stipa grandis)、糙隐子草(Cleistogenes squarrosa)、羊草(Leymus chinensis)、黄囊苔草(Carex korshinskii Kom.)、糙苏(Phlomoides umbrosa)和黄蒿(Artemisia scoparia Waldst.).典型草原试验区土壤类型以为栗钙土为主, 试验区常见优势种多为:冷蒿(Artemisia frigida)、克氏针茅(Stipa krylovii)、糙隐子草、冰草(Agropyron cristatum)和黄囊苔草. 荒漠草原试验区土壤类型以棕钙土为主, 试验区常见优势种多为冷蒿和小叶锦鸡儿(Caragana microphylla)[12].
1.2 数据收集在研究区内各站点布设自动气象站收集包括:空气温度、相对湿度、降雨、气压和风向、风速、土壤温度和土壤水分等数据. 在每年7月植被生长旺盛时期采集各站点的土壤、植被样品, 每个研究站点随机选取3个1 m × 1 m的样方进行取样, 并记录植物种类、高度、盖度. 同时记录样方内出现的所有植物名称, 装袋标记好保存, 待结束野外工作后内业鉴定. 土壤样品则在对应样方内采集, 用直径5 cm土钻在0~50 cm各层(0~10、10~30和30~50 cm)取3个重复土样, 进行土壤理化指标测定, 同时采集环刀样.
1.3 样品测定方法 1.3.1 土壤容重土壤容重测定采用环刀法测定, 将环刀土烘干(105℃)至恒重, 烘干土质量与环刀容积的比值为土壤容重;采用电导率仪和pH计测定电导率(EC)和pH值;采用重铬酸钾氧化法[13]计算土壤有机碳(SOC);土壤有机碳累积指数按美国农业部自然资源保护局方法计算[24]:
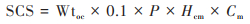
|
(1) |
式中, SCS为土壤有机碳累积量(kg·m-2);Wtoc为以 < 2 mm土壤为基数的有机碳的质量分数(%);0.1为转换因子, 常数;P为 < 2 mm的土壤容重(g·cm-3);Hcm为土层厚度(cm);Cm为 > 2 mm砾石转换系数, 如无砾石则Cm=1.
1.3.2 植被碳储量
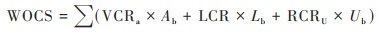
|
(2) |
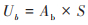
|
(3) |
式中, VCRa为地上植被含碳率, Ab为地上植被生物量(g·m-2), LCR为枯落物含碳率, Lb为枯落物生物量(g·m-2);RCRU为地下根系含碳率, Ub为地下根系生物量(g·m-2);草地地下生物量由地上生物量实测数据计算, S为根冠比
1.3.3 物种多样性指数丰富度指数(R):
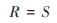
|
(4) |
香农-维纳多样性指数(H):
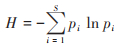
|
(5) |
均匀度指数(E):
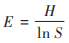
|
(6) |
优势度指数(C):
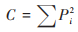
|
(7) |
式中, 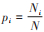
本文利用主成分分析各类型草原禁放牧条件下植物群落组成差异, 利用R 4.2.0进行Mantel-test检验来分析环境变量对植被地上生物量的影响. 使用R语言软件中的“ggalluvial”[14]程序包绘制各土层的有机碳变化图和各站点植被碳密度变化图. 最后根据“plspl”[15]程序包对气象因素、生物量、土壤水分、有机碳含量之间的关系进行路径分析, 并绘制土壤有机碳、土壤水分和植物特征的耦合关系图. 文中表格使用Excel 2016制作, 图形由Origin 2021和R语言软件进行绘制.
2 结果与分析 2.1 不同管理模式下各草原类型植物群落组成的差异如图 2所示, 禁牧条件下草甸草原禾本科植物地上生物量最高, 比典型草原禾本科植物地上生物量高10.22%, 莎草科地上生物量草甸草原略高于典型草原;典型草原菊科类、豆科类和蔷薇科植被地上生物量分别比草甸草原高3.68倍、0.99倍和0.88倍. 整体来看, 相比典型草原, 禁牧条件对草甸草原禾本科(冷蒿、黄蒿、艾蒿)的生长更有利. 其它科属生长优势相对较弱, 尤其是百合科, 草原生长的百合科植物大部分是一些茎叶矮小的葱类, 与羊草和冷蒿相比竞争优势较小. 尽管荒漠草原各科植物都不如其他两种草原生长旺盛, 但百合科植物比草甸草原多.

|
图 2 相同管理模式下不同草地类型植被地上生物量的差异 Fig. 2 Differences in above-ground biomass of vegetation in different grassland types under the same management pattern |
放牧对地上生物量有显著的影响, 图 3对比了3种草原类型中主要植被的地上生物量. 羊草、冰草、克氏针茅和中华隐子草为禾本科, 黄蒿、银灰旋花和黄囊苔草分别为菊科、旋花科和沙草科. 羊草地上生物量典型草原和草甸草原放牧比禁牧分别降低了84.55%和70.09%, 荒漠草原禾羊草地上生物量放牧比禁牧高85.08%. 放牧导致黄囊苔草和黄蒿生物量增加, 银灰旋花地上生物量减少.

|
图 3 相同管理模式下不同草地类型主要植被地上生物量 Fig. 3 Above-ground biomass of major vegetation in different grassland types under the same management pattern |
禁牧对不同草原类型植物多样性影响不同. 如图 4(b)和4(e)所示, 3种草原类型的地上生物量和植被覆盖度均为禁牧高于放牧. 其中禁牧和放牧地上生物量差异最大, 典型草原扎鲁特旗(北)放牧比禁牧低63.89%, 次之是草甸草原乌拉盖, 放牧比禁牧低62.65%;而同为典型草原, 由于地理位置的不同, 扎鲁特旗(北)和锡林浩特(南)地上生物量差异较大. 植被覆盖度荒漠草原召河差异最大, 放牧比禁牧低49.45%. 如图 4(c)和4(f)所示, 典型草原扎鲁特旗(北)均匀度指数和香农-维纳多样性指数差异最大, 放牧比禁牧分别低43.06%和45.47%;而草甸草原和荒漠草原均匀度指数和香农-维纳多样性指数为放牧高于禁牧, 其中草甸草原乌拉盖差异最大, 放牧比禁牧高35.66%和53.26%. 如图 4(a)所示, 除草甸草原外, 荒漠草原和典型草原优势度指数放牧高于禁牧. 物种丰富度指数[图 4(d)]荒漠草原和草甸草原差异较小, 两个典型草原因为地理位置不同, 对物种丰富度的影响不同, 锡林浩特(南)为放牧高于禁牧20%, 扎鲁特旗(北)为禁牧高于放牧12.5%.

|
图 4 禁牧和放牧下草原各站点物种多样性 Fig. 4 Species diversity of grassland sites under grazing and grazing exclusion |
各类型草原总平均植物碳储量由大到小分别为:草甸草原 > 典型草原 > 荒漠草原, 但是从图 5可以看出, 对于同一类型草原不同站点的植被碳储量差异不同, 例如:典型草原正镶白旗(东、西)站点、阿巴嘎旗(南、北)站点、扎鲁特旗(南、北)站点和东乌珠穆沁旗(东、西)站点, 它们的地下根系和地上植被碳储量有明显差异, 其中正镶白旗(西)总植被碳储量最高为412.41 g·m-2, 阿巴嘎旗(南)仅有57.20 g·m-2, 正镶白旗(西)比阿巴嘎旗(南)高6.2倍. 草甸草原东乌珠穆沁旗(东、西)站点植物碳储量相近, 而乌拉盖植被碳储量高于东乌珠穆沁旗1.39倍. 荒漠草原站点的植被碳储量普遍较低, 苏尼特右旗(北)植被碳储量最高, 为73.96 g·m-2, 比草甸草原最高植被碳储量乌拉盖低73.96%, 而四子王旗植被碳储量最低, 与草甸草原最高植被碳储量乌拉盖低92.09%.

|
a1. 正镶白旗(西), a2. 正镶白旗(东), a3. 西乌珠穆沁旗(东), a4. 锡林浩特(南), a5. 扎鲁特旗(北), a6. 扎鲁特旗(南), a7. 西乌珠穆沁旗(西), a8. 阿巴嘎旗(北), a9. 阿巴嘎旗(南), b1. 乌拉盖, b2. 东乌珠穆沁旗(东), b3. 东乌珠穆沁旗(西), c1. 苏尼特右旗(北), c2. 苏尼特右旗(南), c3. 苏尼特左旗(南), c4. 召河, c5. 苏尼特左旗(北), c6. 四子王旗 图 5 放牧条件下不同草原类型植被碳储量变化 Fig. 5 Changes in carbon stocks in vegetation of different grassland types under grazing conditions |
如图 6所示, 禁牧会促使植被碳的累积, 但禁牧对不同草原类型会有不同的影响. 典型草原扎鲁特旗(北)植被碳储量禁牧和放牧差异最大, 地上植被碳储量禁牧高于放牧3.77倍、地下植被碳储量禁牧高于放牧1.77倍, 但锡林浩特(南)的差异较小, 并且扎鲁特旗(北)的禁牧区植被碳储量远高于锡林浩特(南), 是由于扎鲁特旗(北)为短期禁牧区而锡林浩特(南)为长期禁牧区, 表明长期禁牧不利于植被碳储量的增加. 地上植被碳储量和地下植被碳储量禁牧放牧差异较小的是草甸草原, 禁牧分别高于放牧1.28倍和1.54倍;荒漠草原地上植被和地下根系碳储量禁放牧的差异最小, 禁牧分别高于放牧0.66倍和0.63倍.

|
c1. 乌拉盖, c2. 锡林浩特(南), c3. 扎鲁特旗(北), c4. 召河 图 6 禁牧和放牧条件下植被碳储量变化 Fig. 6 Changes in vegetation carbon stocks under grazing and grazing exclusion |
图 7显示, 草甸草原和典型草原生物量对植被和根系碳储量均为正向影响, 而在荒漠草原生物量对植被和根系碳储量没有间接影响;草甸草原和典型草原植被和根系受地理因子影响较大, 而荒漠草原受地理位置影响较小;除典型草原地下根系碳储量, 气象因子对3种草原的植被和根系碳储量直接影响都小于间接影响, 在荒漠草原地下根系碳储量几乎不受气候因素的直接影响.

|
(a)草甸草原, (b)典型草原, (c)荒漠草原;实线表示路径系数;蓝色和红色箭头分别表示因果关系的正向和负向 图 7 放牧条件下不同草原类型植被碳储量与环境因子的影响关系 Fig. 7 Relationship between carbon stocks of different grassland types of vegetation and environmental factors under grazing exclusion |
结构方程模型表明, 草甸草原和典型草原土壤因子对植被碳储量直接影响为抑制作用, 间接和总影响为积极作用, 而对地下根系碳储量均为正面影响;荒漠草原土壤因子对植被碳储量的直接影响为正面影响, 对植被和根系碳储量均为负面的间接和总影响. 典型草原土壤容重对地上植被碳储量负面影响最大, 通径系数为-0.61, 其次为草甸草原, 土壤容重对地上植被碳储量负面影响值为-0.46;在荒漠草原中土壤容重对地上植被碳储量为促进作用, 通径系数为0.33, 土壤有机碳对地上植被碳储量的负面影响最大, 通径系数为-0.38, 土壤水分对地下根系碳储量负面影响最大, 通径系数为-0.36.
在草甸草原, 土壤有机碳对地上植被和地下根系碳储量的积极影响最大, 通径系数分别为0.54和0.87;典型草原中土壤有机碳对地上植被和地下根系碳储量积极影响最大, 通径系数分别为0.77和0.62.而荒漠草原土壤温度对地上植被和地下根系碳储量影响最大, 通径系数分别为0.46和0.65.
如图 7显示, 草甸草原的地上植被碳储量对气候、土壤、地理和生物量响应明显, 而气候因子对地下植被碳储量间接影响大于直接影响. 荒漠草原植被碳储量受气候因子影响较大, 另外地下植被碳储量受生物量影响也比较大. 气候因子中降雨和风速所占的权重均大于0.7, 其中降水通过引起土壤水分变化进而对地上植被和地下植被碳储量产生影响. 土壤水分对典型草原地上和地下植被碳储量为正向直接影响, 路径系数为0.24和0.12, 土壤水分对草甸草原地上和地下植被碳储量为负向直接影响, 路径系数为-0.14和-0.16, 土壤水分对荒漠草原地上和地下植被碳储量为负向直接影响, 路径系数为-0.36和-0.36. 气候因子对荒漠草原地上和地下植被碳储量的负向影响最大.
地理因子正向的间接影响对地上、地下植被碳储量的影响也较大, 而土壤因子对地上碳储量负向的直接影响和对地下碳储量的正向间接影响也较大, 其中除了pH之外, 其他潜在变量土壤水分、土壤温度、土壤容重和土壤有机碳的载荷范围在0.73~0.9, 均起着重要作用. 并且土壤因子相互之间也存在关系, 土壤温度对土壤有机碳在草甸草原和典型草原为正向影响, 而在荒漠草原为负向影响;且土壤容重对土壤有机碳影响较大.
为了识别和量化植被碳储量与生物量、地理位置、气候因子变量之间的关系, 使用偏最小二乘路径模型, 分析各个变量对生物量和植被碳储量的影响. 图 8显示经度对草甸草原和荒漠草原的生物量和地上植被碳储量有显著的正向影响, 相反对典型草原的地上生物量和植被碳储量有不利影响. 而纬度对3种类型草原多为负向影响, 较少部分为正向影响且影响较小.

|
(a)草甸草原, (b)典型草原, (c)荒漠草原;实线表示路径系数;蓝色和红色箭头分别表示因果关系的正向和负向 图 8 放牧条件下不同草原类型植被碳储量、生物量与地理位置、环境因子的影响关系 Fig. 8 Relationships between carbon stocks and biomass of vegetation in different grassland types under grazing conditions and the influence of geographic location and environmental factors |
气候因子主要为降雨、饱和水汽压差、空气温度和相对湿度, 以上因素之间饱和水汽压差主要影响空气温度, 而饱和水汽压差受降雨影响较大. 气候因素对草甸草原影响较大, 且饱和水汽压差、空气温度和相对湿度为主要影响因子;典型草原和荒漠草原降雨对地上植被碳储量影响较大且为负向影响.
3 讨论 3.1 植物多样性对草地管理模式的响应本研究结果发现, 放牧对草地植被群落组成和结构产生影响, 这可能与放牧活动引起的土壤环境变化有关[25, 26].牲畜是草地生态系统的重要组成部分, 其直接影响着草地地上生物量和植被的群落组成[27].因此, 禁牧避免了牲畜的采食、践踏和其他人为干扰条件, 使草原禾本科植物、菊科、豆科和蔷薇科植株能够完成正常的生长、开花和结实等生命周期, 地上生物量明显高于放牧草地. 其次禁牧后土壤条件得到改善, 有利于退化草地的再生和发展. 而放牧区莎草科植物地上生物量大于禁牧区, 主要由于莎草科代表植物株高矮小, 生态幅度广, 耐寒耐旱, 在草原退化时地上生物量增加[28].此外, 不同草地类型下, 植被群落组成对地上生物量的贡献差异较大, 而草甸草原植被功能群落的种类组成变化对放牧最为敏感.
放牧条件下, 植被叶面积减少, 光合能力降低, 草地物种组成产生变化. 其中, 植被盖度被认为最容易受到放牧影响, 导致盖度减少. 同时, 典型草原放牧对植被盖度的影响差异较小, 而草甸草原和荒漠草原植被过于旺盛或稀疏, 放牧导致植被盖度下降较大, 表明放牧对植被盖度影响程度不仅取决于放牧强度, 还与草地类型有关. 不同草地类型的局域物种库存在显著差异也导致了放牧后群落结构的变化. 放牧显著提高草甸草原植物群落的多样性和物种均匀度, 降低了优势度指数, 主要是由于草食动物的采食显著降低了优势种的生物量和相对多度, 促进了物种多样性和均匀度[29].相比草甸草原, 典型草原放牧导致植被多样性和均匀度降低, 而优势度指数提高. 典型草原大部分位于干旱区, 属于生产力比较低的草原, 当地的物种主要是一些资源保守种, 倾向于资源的保存, 而保守种在放牧干扰下往往走向死亡[30, 31].荒漠草原在放牧情况下物种丰富度下降, 多样性、均匀度和优势度指数相对禁牧略微升高. 牲畜的选择性采食削弱了优势物种的竞争力, 为一些一二年生物物种的生长提供机会, 从而弥补因放牧压力导致群落多样性的损失[32, 33].禁牧促进植物生长, 而植物通过叶片蒸腾作用释放水蒸气, 从而促进水分循环和二氧化碳的吸收[34].
3.2 植被碳储量对草地管理模式的响应放牧是影响植物生产力、植被盖度以及凋落物盖度等指标的因素之一, 从而改变植被碳储量[35].本研究发现短期禁牧[扎鲁特旗(北)]植被含碳率高于放牧区, 而长期禁牧[锡林浩特(南)]低于放牧区. 因为植被碳储量与生物量显著正相关[36], 而随着禁牧年限的增加地上生物量呈先增加后减少的趋势[11], 植被碳储量也随着生物量呈先增加后减少. 并且土壤肥力是影响植被碳储量的主导因素[37], 放牧会导致一年生植物和杂类植物物种的丰富度显著增加, 一年生植物通过呼吸作用土壤碳损失高于多年生植物[38, 39], 并且放牧也会降低土壤养分, 因此限制了植被碳的恢复[40, 41].土壤水分与植被生产力相关, 禁牧改善土壤水分, 可能更有利于植被碳储量的增加;降雨也是增加土壤水分的途径, 气象数据显示乌拉盖草原降雨量是东乌珠穆沁旗的两倍, 乌拉盖草原植被碳储量为东乌珠穆沁旗草原的2倍(图 7)[42].长期禁牧不仅抑制植物的再生和幼苗的形成, 还降低植被生产力的转化[43, 44], 导致长期禁牧后植被碳储量下降. 适度放牧会降低竞争强度, 有利于多物种共生, 为群落中物种的恢复创造空间, 加快土壤养分循环[45].因此短期禁牧后进行适度的放牧更有利于草原生态系统的恢复.
此外, 不同草地类型植物群落组成不同, 典型草原禾本科植物多于荒漠草原, 草甸草原禾本科又大于典型草原, 研究表明杂类植物比禾本科植物的根系浅, 限制它们的生物量生产, 从而禾本科植物的有机碳高于杂类植物[46, 47].长期禁牧减少了禾草科、沙草科和豆科的分配(在高寒草甸), 豆科减少是表层土壤根系减少的主要原因, 进而降低植物碳储量[21].在湿润条件下植物固碳更多[48], 植物光合作用的增强促进生物量碳库的增加[49], 因此草甸草原固碳能力强于典型草原, 典型草原又强于荒漠草原. 而植被恢复群落盖度增加保护表层土壤结构, 而大量的根系生物量和枯落物输入, 在微生物的分解作用下, 可以改善土壤理化性质, 提高土壤有机碳含量, 保护生态环境[50, 51].
3.3 不同草地环境因子对植被有机碳的影响放牧不仅改变植物群落特征, 还会降低植被生产力. 放牧通过采食以及尿液的排泄会改变土壤的理化性质. 研究发现气候因子和地理因素是对植被碳的主要影响, 尤其是降雨因素, 降雨多的草甸草原和降雨少的荒漠草原的土壤水分均产生负向影响, 典型草原的土壤水分对植被碳储量为正向影响. 产生这一现象的原因是不同植被群落的水分利用机制差异造成的, 荒漠草原为干旱区, 草原植被的水分利用机制相对保守, 其耐受水分胁迫的能力较强;而草甸草原为湿润地区, 湿润区植被的生物多样性较好, 不同物种之间的“补偿效应”能充分利用好土壤水分[52].土壤水分受降雨的影响对不同草原类型的植被碳的影响不同, 荒漠草原旱季较长会加剧水土流失, 阻碍植被的生长;草甸草原土壤水分过高也会阻碍植被的生长, 进而阻碍植被碳[53].土壤有机碳增加导致活性生物和孔隙度增加, 降低土壤容重, 有利于植被生长, 因此草甸草原和典型草原的土壤有机碳有利于植被的生长. 并且土壤有机碳作为土壤养分供植被生长, 将更多的碳储存在生物体内. 如图 6所示土壤有机碳与植被碳出现负相关, 是由于植被生长较快, 土壤有机碳作为土壤养分消耗而凋落物返回土壤较少[34].
4 结论不同管理模式下草地植被组成及其多样性差异显著. 放牧显著提高了草甸草原植被的香农-维纳多样性及物种均匀度, 降低了优势度指数. 但放牧降低荒漠草原物种丰富度, 而香农-维纳多样性、物种均匀度和优势度指数相对禁牧略微升高. 由Mantel-test分析可知典型草原的地上生物量和降雨呈显著正相关, 但草甸草原和荒漠草原的地上生物量分别与空气相对湿度呈显著正相关和显著负相关. 禁牧区地上植被和地下根系碳储量远高于放牧区, 禁牧分别高于放牧的1.5倍和1.2倍, 而长期禁牧的站点的植被碳储量明显低于短期禁牧. 结构方程模型显示:长期禁牧土壤水分和总生物量对土壤有机碳储量负向的直接影响大于短期禁牧站点. 根据研究结果可知, 气候因子尤其是降雨对于荒漠草原植被群落和碳储量影响尤为重要, 同时长期禁牧不能维持植物多样性, 且不利于草地生态系统碳累积, 应当适当放牧, 但也需要注意对土壤水分, 植被碳储量等的不利影响.
| [1] |
赵威, 李琳, 王艳杰, 等. 河南典型暖(热)性草地固碳特征及区域差异[J]. 应用生态学报, 2018, 29(6): 1867-1875. Zhao W, Li L, Wang Y J, et al. Carbon sequestration characteristics and regional differences of typical warm and tropical grasslands in Henan Province, China[J]. Chinese Journal of Applied Ecology, 2018, 29(6): 1867-1875. |
| [2] |
刘嘉琳. 内蒙古典型草原植被退化对土壤有机碳组分及其稳定性的影响[D]. 呼和浩特: 内蒙古大学, 2022. Liu J L. Effects of vegetation degradation on soil organic carbon components and their stability in typical steppe of inner Mongolia[D]. Hohhot: Inner Mongolia University, 2022. |
| [3] |
李健明, 康雨欣, 蒋福祯, 等. 基于Meta分析的煤矿区植被恢复对土壤有机碳储量的影响[J]. 环境科学, 2024, 45(3): 1629-1643. Li J M, Kang Y X, Jiang F Z, et al. Effect of vegetation restoration on soil organic carbon storage in coal mining areas based on Meta-analysis[J]. Environmental Science, 2024, 45(3): 1629-1643. |
| [4] |
郑慧, 薛江博, 桂建华, 等. 放牧强度对华北农牧交错带典型草地土壤化学计量特征的短期影响[J]. 应用生态学报, 2021, 32(7): 2433-2439. Zheng H, Xue J B, Gui J H, et al. Short-term effects of grazing intensity on soil stoichiometric characteristics of typical grassland in the agro-pastoral ecotone of northern China[J]. Chinese Journal of Applied Ecology, 2021, 32(7): 2433-2439. |
| [5] | Hewins D B, Lyseng M P, Schoderbek D F, et al. Grazing and climate effects on soil organic carbon concentration and particle-size association in northern grasslands[J]. Scientific Reports, 2018, 8(1). DOI:10.1038/s41598-018-19785-1 |
| [6] | Gong L, Liu G H, Wang M, et al. Effects of vegetation restoration on soil organic carbon in China: a meta-analysis[J]. Chinese Geographical Science, 2017, 27(2): 188-200. DOI:10.1007/s11769-017-0858-x |
| [7] |
谢绮丽, 杨鑫, 郝利娜. 2001—2020年三江源区植被覆盖时空变化特征及其影响因素[J]. 水土保持通报, 2022, 42(5): 202-212. Xie Q L, Yang X, Hao L N. Spatio-temporal variation of vegetation cover and its driving factors in Three-River Headwaters Region during 2001—2020[J]. Bulletin of Soil and Water Conservation, 2022, 42(5): 202-212. |
| [8] | Chai Q L, Ma Z Y, Chang X F, et al. Optimizing management to conserve plant diversity and soil carbon stock of semi-arid grasslands on the Loess Plateau[J]. CATENA, 2019, 172: 781-788. DOI:10.1016/j.catena.2018.09.034 |
| [9] | Liu J H, Wu J J, Su H B, et al. Effects of grazing exclusion in Xilin Gol grassland differ between regions[J]. Ecological Engineering, 2017, 99: 271-281. DOI:10.1016/j.ecoleng.2016.11.041 |
| [10] |
杨鹏年, 李希来, 李成一, 等. 黄河源区斑块化退化高寒草甸土壤微生物多样性对长期封育的响应[J]. 环境科学, 2023, 44(4): 2293-2303. Yang P N, Li X L, Li C Y, et al. Response of soil microbial diversity to long-term enclosure in degraded patches of alpine meadow in the source zone of the Yellow River[J]. Environmental Science, 2023, 44(4): 2293-2303. |
| [11] |
宋成刚, 张铭洋, 何琦, 等. 禁牧封育对祁连山南麓高寒草甸植被群落结构及土壤水分特征的影响[J]. 中国草地学报, 2023, 45(4): 22-32. Song C G, Zhang M Y, He Q, et al. Effects of Grazing prohibition on vegetation community structure and soil moisture characteristics of alpine meadows on the southern slope of the Qilian Mountains[J]. Chinese Journal of Grassland, 2023, 45(4): 22-32. |
| [12] |
萨茹拉. 不同利用方式和放牧强度对典型草原植被—土壤系统碳储量的影响[D]. 北京: 中国农业科学院, 2013. Sarula. Effects of different utilization and grazing intensity on carbon storage of vegetation-soil system in typical steppe[D]. Beijing: Chinese Academy of Agricultural Sciences, 2013. |
| [13] | Žuna Pfeiffer T, Špoljarić Maronić D, Zahirović V, et al. Effectiveness of mowing for the flora diversity preservation: a case study of steppe-like grassland in Croatia (NATURA 2000 site)[J]. Biodiversity and Conservation, 2017, 26(2): 439-460. DOI:10.1007/s10531-016-1252-1 |
| [14] |
萨茹拉, 侯向阳, 李金祥, 等. 不同放牧退化程度典型草原植被—土壤系统的有机碳储量[J]. 草业学报, 2013, 22(5): 18-26. Sarula, Hou X Y, Li J X, et al. Organic carbon storage in vegetation-soil systems of typical grazing degraded steppes[J]. Acta Prataculturae Sinica, 2013, 22(5): 18-26. |
| [15] | Fotis A T, Murphy S J, Ricart R D, et al. Above‐ground biomass is driven by mass‐ratio effects and stand structural attributes in a temperate deciduous forest[J]. Journal of Ecology, 2018, 106(2): 561-570. DOI:10.1111/1365-2745.12847 |
| [16] |
辛晓平, 丁蕾, 程伟, 等. 北方草地及农牧交错区草地植被碳储量及其影响因素[J]. 中国农业科学, 2020, 53(13): 2757-2768. Xin X P, Ding L, Cheng W, et al. Biomass carbon storage and its effect factors in steppe and agro-pastoral ecotones in northern China[J]. Scientia Agricultura Sinica, 2020, 53(13): 2757-2768. DOI:10.3864/j.issn.0578-1752.2020.13.022 |
| [17] | Boerner R E J. Unraveling the Gordian Knot: interactions among vegetation, topography, and soil properties in the central and southern Appalachians[J]. The Journal of the Torrey Botanical Society, 2006, 133(2): 321-361. DOI:10.3159/1095-5674(2006)133[321:UTGKIA]2.0.CO;2 |
| [18] | Da Costa A C L, Metcalfe D B, Doughty C E, et al. Ecosystem respiration and net primary productivity after 8-10 years of experimental through-fall reduction in an eastern Amazon forest[J]. Plant Ecology & Diversity, 2014, 7(1-2): 7-24. |
| [19] | Li H Y, Qu Y, Zeng X Y, et al. Dynamic response of the vegetation carbon storage in the Sanjiang plain to changes in land use/cover and climate[J]. Heritage Science, 2021, 9(1). DOI:10.1186/40494-021-00605-1 |
| [20] | Wang L X, Gao J X, Shen W M, et al. Carbon storage in vegetation and soil in Chinese ecosystems estimated by carbon transfer rate method[J]. Ecosphere, 2021, 12(1). DOI:10.1002/ecs2.3341 |
| [21] |
宋珊珊, 张建胜, 郑天立, 等. 围栏封育对青海海北高寒草甸植被碳储量的影响[J]. 草业科学, 2020, 37(12): 2414-2421. Song S S, Zhang J S, Zheng T L, et al. Effect of fencing on vegetation carbon storage in the Qinghai Haibei alpine meadows[J]. Pratacultural Science, 2020, 37(12): 2414-2421. |
| [22] | Wang S, Zhang S W, Lin X, et al. Response of soil water and carbon storage to short-term grazing prohibition in arid and semi-arid grasslands of China[J]. Journal of Arid Environments, 2022, 202. DOI:10.1016/j.jaridenv.2022.104754 |
| [23] |
季波, 何建龙, 王占军, 等. 宁夏天然草地植被碳储量特征及构成[J]. 应用生态学报, 2021, 32(4): 1259-1268. Ji B, He J L, Wang Z J, et al. Characteristics and composition of vegetation carbon storage in natural grassland in Ningxia, China[J]. Chinese Journal of Applied Ecology, 2021, 32(4): 1259-1268. |
| [24] | Wu G L, Liu Z H, Zhang L, et al. Long-term fencing improved soil properties and soil organic carbon storage in an alpine swamp meadow of western China[J]. Plant and Soil, 2010, 332(1): 331-337. |
| [25] | Lin X, Zhao H B, Zhang S W, et al. Impact of precipitation and grazing on recovery of plant vegetation in temperate grasslands[J]. Land Degradation & Development, 2023. DOI:10.1002/ldr.4980 |
| [26] | Zhao X Y, Zhao H B, Zhang S W, et al. Impact of years of grazing exclusion on surface soil temperature and hydrothermal relations in arid and semi-arid grasslands[J]. Ecohydrology, 2023. DOI:10.1002/eco.2603 |
| [27] |
李娜, 唐士明, 郭建英, 等. 放牧对内蒙古草地植物群落特征影响的meta分析[J]. 植物生态学报, 2023, 47(9): 1256-1269. Li N, Tang S M, Guo J Y, et al. Meta-analysis of effects of grazing on plant community properties in Nei Mongol grassland[J]. Chinese Journal of Plant Ecology, 2023, 47(9): 1256-1269. |
| [28] | Yan Y, Lu X Y. Is grazing exclusion effective in restoring vegetation in degraded alpine grasslands in Tibet, China?[J]. PeerJ, 2015, 3. DOI:10.7717/peerj.1020 |
| [29] |
龚心语, 黄宝荣, 邓冉, 等. 自然保护区退牧还草生态补偿标准——以向海国家级自然保护区为例[J]. 生态学报, 2021, 41(12): 4694-4706. Gong X Y, Huang B R, Deng R, et al. Ecological compensation standards for returning grazing land to grassland in nature reserves: a case study of Xianghai National Nature reserve in Jilin[J]. Acta Ecologica Sinica, 2021, 41(12): 4694-4706. |
| [30] | Larson J E, Funk J L. Seedling root responses to soil moisture and the identification of a belowground trait spectrum across three growth forms[J]. New Phytologist, 2016, 210(3): 827-838. DOI:10.1111/nph.13829 |
| [31] | Reich P B. The world-wide 'fast-slow' plant economics spectrum: a traits manifesto[J]. Journal of Ecology, 2014, 102(2): 275-301. DOI:10.1111/1365-2745.12211 |
| [32] |
周瑶. 季节性放牧对荒漠草原植物功能性状和功能多样性的影响[D]. 银川: 宁夏大学, 2021. Zhou Y. Effects of seasonal grazing on plant functional traits and functional diversity of the desert grasslands[D]. Yinchuan: Ningxia University, 2021. |
| [33] |
宿婷婷. 季节性轮牧对荒漠草原植物功能性状和多样性的影响[D]. 银川: 宁夏大学, 2020. Su T T. Effects of seasonal rotational grazing on plant functional traits and diversity of desert steppe[D]. Yinchuan: Ningxia University, 2020. |
| [34] |
宋致华. 子午岭桥山林区不同林分碳库储量及其影响因素研究[D]. 西安: 西安理工大学, 2023. Song Z H. Study on carbon storage and its influencing factors of different stand structures in Ziwuling forest region[D]. Xi'an: Xi'an University of Technology, 2023. |
| [35] |
董乙强, 孙宗玖, 安沙舟. 放牧和禁牧影响草地物种多样性和有机碳库的途径[J]. 中国草地学报, 2018, 40(1): 105-114. Dong Y Q, Sun Z J, An S Z. Effects of grazing and grazing exclusion on species diversity in grassland vegetation and organic carbon[J]. Chinese Journal of Grassland, 2018, 40(1): 105-114. |
| [36] |
董晓玉, 傅华, 李旭东, 等. 放牧与围封对黄土高原典型草原植物生物量及其碳氮磷贮量的影响[J]. 草业学报, 2010, 19(2): 175-182. Dong X Y, Fu H, Li X D, et al. Effects on plant biomass and CNP contents of plants in grazed and fenced steppe grasslands of the Loess Plateau[J]. Acta Prataculturae Sinica, 2010, 19(2): 175-182. |
| [37] | Yu L F, Sun W J, Huang Y. Grazing exclusion enhances plant and topsoil carbon stocks in arid and semiarid grasslands[J]. Agriculture, 2021, 320. DOI:10.1016/j.agee.2021.107605 |
| [38] | Zhou G Y, Zhou X H, He Y H, et al. Grazing intensity significantly affects belowground carbon and nitrogen cycling in grassland ecosystems: a meta‐analysis[J]. Global Change Biology, 2017, 23(3): 1167-1179. |
| [39] | Zhao L, Chen D D, Zhao N, et al. Responses of carbon transfer, partitioning, and residence time to land use in the plant–soil system of an alpine meadow on the Qinghai-Tibetan Plateau[J]. Biology and Fertility of Soils, 2015, 51(7): 781-790. |
| [40] | Gass T M, Binkley D. Soil nutrient losses in an altered ecosystem are associated with native ungulate grazing[J]. Journal of Applied Ecology, 2011, 48(4): 952-960. |
| [41] | Tessema Z K, De Boer W F, Baars R M T, et al. Changes in soil nutrients, vegetation structure and herbaceous biomass in response to grazing in a semi-arid savanna of Ethiopia[J]. Journal of Arid Environments, 2011, 75(7): 662-670. |
| [42] | Dang C Y, Shao Z F, Huang X, et al. Climate warming-induced phenology changes dominate vegetation productivity in Northern Hemisphere ecosystems[J]. Ecological Indicators, 2023, 151. DOI:10.1016/j.ecolind.2023.110326 |
| [43] | Risser P G. Making ecological information practical for resource managers[J]. Ecological Applications, 1993, 3(1): 37-38. |
| [44] | Altesor A, Oesterheld M, Leoni E, et al. Effect of grazing on community structure and productivity of a Uruguayan grassland[J]. Plant Ecology, 2005, 179(1): 83-91. |
| [45] | Bernhardt-Römermann M, Römermann C, Sperlich S, et al. Explaining grassland biomass - the contribution of climate, species and functional diversity depends on fertilization and mowing frequency[J]. Journal of Applied Ecology, 2011, 48(5): 1088-1097. |
| [46] | Li W B, Huang G Z, Zhang H X. Enclosure increases nutrient resorption from senescing leaves in a subalpine pasture[J]. Plant and Soil, 2020, 457(1-2): 269-278. |
| [47] | Wei Y J, Liu M Y, Wang J, et al. The effects of vegetation communities on soil organic carbon stock in an enclosed desert-steppe region of northern China[J]. Soil Science and Plant Nutrition, 2022, 68(2): 284-294. |
| [48] | Deng L, Shangguan Z P, Wu G L, et al. Effects of grazing exclusion on carbon sequestration in China's grassland[J]. Earth-Science Reviews, 2017, 173: 84-95. |
| [49] | Hu Z M, Li S G, Guo Q, et al. A synthesis of the effect of grazing exclusion on carbon dynamics in grasslands in China[J]. Global Change Biology, 2016, 22(4): 1385-1393. |
| [50] |
许小明, 张晓萍, 王浩嘉, 等. 黄土高原子午岭植被自然恢复下的固碳特征[J]. 环境科学, 2023, 44(5): 2756-2766. Xu X M, Zhang X P, Wang H J, et al. Carbon sequestration characteristics under natural vegetation restoration in Ziwuling area of the Loess Plateau[J]. Environmental Science, 2023, 44(5): 2756-2766. |
| [51] |
许小明, 张晓萍, 何亮, 等. 黄土丘陵区不同恢复植被类型的固碳特征[J]. 环境科学, 2022, 43(11): 5263-5273. Xu X M, Zhang X P, He L, et al. Carbon sequestration characteristics of different restored vegetation types in loess hilly region[J]. Environmental Science, 2022, 43(11): 5263-5273. |
| [52] |
高添. 内蒙古草地植被碳储量的时空分布及水热影响分析[D]. 北京: 中国农业科学院, 2013. Gao T. Spatio-temporal pattern of biomass carbon storage and its relationships to hydrothermal factors in Inner Mongolia's grassland[D]. Beijing: Chinese Academy of Agricultural Sciences, 2013. |
| [53] | Yang X B, Li D H, McGrouther K, et al. Effect of Eucalyptus forests on understory vegetation and soil quality[J]. Journal of Soils and Sediments, 2017, 17(9): 2383-2389. |