 2024, Vol. 45
2024, Vol. 45



2. 杭州市生态环境局淳安分局, 杭州 311700
2. Chun'an Branch, Hangzhou Bureau of Ecology and Environment, Hangzhou 311700, China
浮游植物作为水体中食物链的基础, 可以对环境做出迅速响应, 能在河流等生态系统中有效地指示水体状况[1].而浮游植物的异常增殖会引发藻类水华, 据报道, 近10年全球藻华暴发频率呈增长态势, 对内陆水体威胁加大[2].因此, 阐明浮游植物群落结构动态特征及影响因素在河流生态系统管理中至关重要[3].由于浮游植物种类繁多, 基于形态分类的群落结构信息还不足以反映水体环境综合信息.功能群是将生活习性和生长策略相似的浮游植物种类进一步归类[4], 既能有效减少非优势种对浮游植物群落的冗余性, 又能更直观反映浮游植物对环境的响应, 并能根据优势功能群预测水体环境因子和营养状况[5], 近年来得到快速的发展.
浮游植物功能群的组成和时空动态是综合环境的结果[6], 营养盐浓度、水动力条件(流态、水力滞留时间等)、气象条件(气温、降雨等)以及生物因素(浮游动物、鱼类等)均有可能是引起水体浮游植物功能群结构变化.如营养盐浓度和光可利用性是穆林河浮游植物功能群季节演替的主要因素[7].在乌江河流-水库体系中, 营养盐与功能群关系较差[8].水温、pH、溶解氧和浊度与阳澄西湖浮游植物功能群构成联系紧密[9].在四明湖水库中, 水温、透明度、硝态氮和浮游动物均对浮游植物功能群的季节演替产生影响[10].淀山湖的浮游植物功能群季节演替则受水位波动的影响[11].
富春江水库是位于钱塘江中游的一座低水头河流型水库, 曾多次暴发过蓝藻水华, 对沿江景观带来巨大挑战.前期研究发现水文气象过程对富春江段藻类群落结构具有影响, 但是环境条件与藻类群落结构变化之间的联系仍不清楚.为此, 本研究首次采用功能群分类的方法分析富春江水库及上下游江段浮游植物的群落演替的关键影响因子, 以期为富春江藻类水华防控提供参考.
1 材料与方法 1.1 采样点位设置本研究以富春江水库为对象, 于2020年8月(夏季)、11月(秋季)、2021年1月(冬季)和4月(春季)开展4次调查.如图 1所示, 以富春江水库为核心, 包括其上下游江段共设置11个采样点位, 其中S1~S5位于富春江水库上游, S6~S9位于水库库区, S10位于富春江大坝下游, S11位于钱塘江河口.

|
S1.衢江;S2.金华江;S3.兰江;S4.将军岩;S5.新安江;S6.梅城大桥;S7.三都大桥;S8.子胥渡;S9.严陵坞;S10.桐庐水厂;S11.闻家堰 图 1 采样点位示意 Fig. 1 Distribution of sampling sites in Qiangtang River |
现场用赛氏盘测定水体透明度(SD), 用美国YSI公司的EXO2多参数水质分析仪测定水温(WT)、电导率(Cond)、浊度(Turb)、pH及溶解氧(DO).用有机玻璃采水器采集表层(0.5 m)水样1.5 L, 立即冷藏, 带回实验室测定总氮(TN)、总磷(TP)、溶解性总氮(DTN)、溶解性总磷(DTP)、溶解性有机碳(DOC)、高锰酸盐指数、悬浮颗粒物(SS)等水质指标.其中, DOC采用multi N/C 2100总有机碳测定仪测定, 其余参数测定方法参照文献[12].
1.3 浮游生物采集与鉴定浮游植物定量样品采集时, 用有机玻璃采水器取水样1 L, 现场加入10 mL鲁戈试剂固定, 带回实验室静置、沉淀48 h后, 利用虹吸法将上清液吸除, 定容至30 mL, 存放在样品瓶中.用光学显微镜OLYMPUS-BX53进行鉴定和计数.浮游植物生物量由各种属细胞密度进行体积转换所得.
浮游动物调查时, 用有机玻璃采水器采集水样30 L, 再用25号浮游生物网过滤后, 用5%甲醛固定, 用于鉴定枝角类与桡足类等后生浮游动物.原生动物和轮虫的鉴定则使用浮游植物定量样品进行.浮游动物生物量根据各种类数量和个体大小计算所得.
1.4 浮游植物功能群划分与优势类群判定根据2002年Reynolds等[13]提出和2009年Padisák等[14]修正的浮游植物功能群(FG)划分方案, 钱塘江干流4个季度采集的浮游植物隶属于18个功能类群.
浮游植物物种和功能群在不同季节和不同点位的优势度(Y)计算方法如下[15].
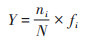
|
式中, ni为i种的浮游植物(或功能群)细胞个数, N为浮游植物总细胞个数, fi为第i种浮游植物(或功能群)出现的频率, 当Y > 0.02时, 该类即为优势属(或优势类群).以优势度大于0.5(绝优势)或者优势度大于0.1的类群(主要优势)划分C-R-S生长策略.
1.5 数据处理与分析用SPSS 23.0进行非参数检验探究水体理化指标和生物指标的季节差异.用CANOCO 5.0分析浮游植物功能群结构和环境因子间的关系.将浮游植物进行Hellinger转换, 将环境因子(除pH外)进行lg(x+1)转换, 然后进行向前选择分析(forward selection)去除共线性因子, 采用蒙特卡洛置换检验选出显著影响因子(P < 0.05)进行冗余分析(redundancy analysis, RDA), 再运用R语言“rdacca.hp”包探究环境因子的相对重要性[15].
采用非度量多维尺度分析(nonmetric multidimensional scaling, NMDS)分析浮游植物群落结构季节演替, 并使用相似性检验(analysis of similarities, ANOSIM)分析差异显著性, 用相似性百分比分析(SIMPER)找出季节间差异的原因, 即主要特征功能类群, 以上分析在PAST 2.17中完成.
2 结果与分析 2.1 浮游植物细胞密度及优势属时空变化4次调查在富春江水库共鉴定出浮游植物80种, 隶属于6门55属, 包括绿藻门22属35种、硅藻门18属29种、蓝藻门9属9种、甲藻门3属3种、裸藻门2属2种及隐藻门1属2种.
富春江水库浮游植物细胞密度时空变化如图 2所示, 浮游植物细胞密度季节差异显著(P < 0.01), 具体大小表现为:冬季 > 夏季 > 秋季 > 春季, 平均细胞密度分别为6.23 × 106、3.98 × 106、9.50 × 105和1.04 × 105 cells·L-1, 全年均值为2.82 × 106 cells·L-1.

|
不同小写字母表示具有差异性 图 2 富春江水库浮游植物细胞密度时空变化 Fig. 2 Temporal-spatial variation in phytoplankton cell density in Fuchunjiang Reservoir |
春季细胞密度最高值2.82 × 105 cells·L-1, 位于钱塘江河口(S11);最低值位于金华江(S2), 为4.8 × 104 cells·L-1.夏季细胞密度最高值在富春江库区三都大桥(S7), 为1.32 × 107 cells·L-1, 最低值为3.6 × 104 cells·L-1, 在新安江.秋季细胞密度最高值在衢江, 为6.37 × 106 cells·L-1, 最低值在富春江段的子胥渡(S8)和桐庐水厂(S10), 为4.50 × 103 cells·L-1.冬季最高细胞密度达到了3.74 × 107 cells·L-1, 出现在金华江(S2);最低细胞密度为4.8 × 104 cells·L-1, 位于富春江库区严陵坞(S9).冬季兰江、金华江和衢江的硅藻细胞密度异常高, 其中金华江段硅藻门细胞密度占据了总细胞数的99%, 硅藻中小环藻细胞密度超过了81%.
富春江水库浮游植物由蓝藻门和硅藻门物种交替占优, 季节上, 夏季优势属为硅藻门的沟链藻(Aulacoseira)和蓝藻门的平裂藻(Merismopedia)、微囊藻(Microcystis)、长孢藻(Dolichospermum)及假鱼腥藻(Pseudoanabaena);秋季演替为沟链藻和长孢藻;冬季演替为小环藻(Cyclotella)和直链藻(Melosira);春季演替为硅藻门的沟链藻、直链藻、小环藻和针杆藻(Synedra)(图 3).空间上, 上游江段(S1~S4)小环藻优势度最高, 其中衢江伴随着沟链藻, 金华江伴随着直链藻;新安江段至富春江段(S5~S10)演替为沟链藻优势度最高;蓝藻门微囊藻、假鱼腥藻、平裂藻、长孢藻和颤藻(Oscillatoria)在兰江至富春江段(S3~S4和S6~S10)交替占优;钱塘江河口占优物种数最多(6种)且均为硅藻门, 其中小环藻优势度最高, 辐环藻(Actinocyclus)次之(图 3).

|
红色表示优势属, 绿色表示非优势属, 黄色表示未出现 图 3 富春江浮游植物优势属及优势度 Fig. 3 Dominant species and dominance index of phytoplankton in Fuchunjiang Reservoir |
根据FG功能群分类方法可将富春江水库浮游植物分为18个功能类群, 分别为C、D、N、P、MP、X1、Y、F、G、J、K、H1、L0、S1、M、X2、W1和W2.其中C、D、P、MP、H1、L0、S1、M、J和Y等10个群为优势类群.富春江水库各浮游植物功能群的代表性藻属、生境特征和生长策略见表 1.
|
|
表 1 富春江水库浮游植物功能群代表性藻属、生境特征及生长策略 Table 1 Functional groups of phytoplankton in Fuchunjiang Reservoir representative algae, habitat characteristics, and growth strategies |
富春江水库浮游植物功能群优势度时空动态如图 4所示.全年来看, 优势类群为C+P+L0, 代表性藻种分别为小环藻、沟链藻和平裂藻, 反映出富春江总体混合作用强烈、营养水平偏高的环境特征.季节上, 夏季功能群类型最丰富, 为P+L0+J+M+S1+H1+MP型, 除了全年都占优的沟链藻为代表的P类群, 还包括多个以蓝藻和绿藻为代表的类群;秋季演替为以长孢藻为代表的H1类群占优, 总体为P+H1, 反映出水环境由扰动强烈向流态稳定的低流环境变化特征;冬季小环藻的突增, 促使C类群占据优势, 总体为P+C, 反映出换水减弱、富营养化加剧的环境条件变化;而春季类群逐渐丰富, 为C+D+P+MP, 反映出交换加强、生境多样等环境变化.

|
红色表示优势类群, 绿色表示非优势类群, 黄色表示未出现 图 4 富春江水库浮游植物功能群优势度时空演替 Fig. 4 Seasonal succession of phytoplankton functional group dominance in Fuchunjiang Reservoir |
从生长策略的角度看, 季节变化为夏季(R/S)→秋季(R)→冬季(R)→春季(R).全年几乎都以R策略为主导, 耐受干扰, 并且能在短期内将资源转换为自身能量.根据欧美湖泊的演替模式来看, 缺少冬末春初C或CS策略→春季S策略的过程, 推测富春江水库浮游植物群落结构可能尚未稳定, 与富春江生境长期处于高干扰状态的环境特点吻合.
空间上, 多种生长策略浮游植物混生, 自上游至下游呈现S1、S2(C+P)→S3(C+P+H1)→S4(C+P+M+Y+L0)→S5(C+D+P+MP)→S6(P+L0)→S7、S8(P+L0+H1)→S9、S10(P+MP+L0+H1)→S11(P+MP+C+D+J)变化特征.生长策略变化特征为:上游河段S1~S5(R)→中游库区S6(R/S)→S7、S8(R)→S9(R/CR/S)→S10(R/CR)→S11(CR).兰江及以上江段(S1~S4)优势度最高的为功能群C, 功能群P次之, 反映出采样点较高的营养水平和较强的扰动;新安江段S5主要的是P型;富春江段S6~S10最主要的功能群也为P, 但其优势度沿程减小, 反映出库区水深加大、分层稳定的生态效应.MP功能群从富春江段开始沿程递增, 直至钱塘江河口S11占据绝对优势, 反映出水体高强度的扰动, 与钱塘江口感潮江段频繁发生的涌潮扰动及快速冲刷有关.
2.3 浮游动物生物量时空变化本次调查共鉴定出浮游动物117种, 其中原生动物49种、轮虫34种、枝角类和桡足类各17种.浮游动物生物量的全年均值为0.42 mg·L-1, 季节变化明显, 如图 5(a)所示, 呈现夏、秋、冬、春季逐渐降低的趋势, 生物量分别为1.11、0.37、0.22和0.02 mg·L-1;从空间上来看, 兰江段生物量最高, S1~S4生物量均值达到了0.59 mg·L-1, 富春江库区段次之, S6~S9生物量均值为0.43 mg·L-1, 而新安江段最低, 生物量仅有0.03 mg·L-1 [图 5(b)].

|
不同小写字母表示具有差异性 图 5 富春江水库浮游动物生物量时空变化 Fig. 5 Temporal-spatial variation in zooplankton biomass in Fuchunjiang Reservoir |
富春江水库水体理化指标浓度的季节变化如图 6所示.总体上ρ(TN)和ρ(DTN)在季节上差异并不显著, 均值分别为2.03 mg·L-1和1.80 mg·L-1.ρ(TP)和ρ(DTP)均值分别为0.075 mg·L-1和0.048 mg·L-1.TP在夏季丰水期外源补给充足时浓度最高;春季的浓度也比较高, 但低于夏季.ρ(DOC)的季节差异较小, 均值为2.37 mg·L-1.高锰酸盐指数夏季和冬季较高, 春秋较低, 均值为1.98 mg·L-1.透明度呈现出冬季高夏季低的特征, 均值为1.06 m.ρ(Chla)季节差异显著, 冬季和夏季较高, 春秋较低, 均值为19.10 μg·L-1.Cond均值为241.38 μS·cm-1, 季节差异不显著.

|
不同小写字母表示季节间具有显著差异性 图 6 富春江水库水体理化因子季节变化 Fig. 6 Seasonal variation in physical and chemical factors in Fuchunjiang Reservoir |
空间上, 不同江段水体理化指标差异显著(图 7).ρ(TN)和ρ(DTN)分别在0.89~5.93 mg·L-1和0.84~4.53 mg·L-1之间变化, 空间差异显著, 局部城镇影响较大, 尤其在金华江, 长期处于高值.ρ(TP)变化范围为0.013~0.208 mg·L-1, 其空间变化趋势相对于DTP浓度更加剧烈, ρ(DTP)的变化范围为0.012~0.163 mg·L-1.ρ(DOC)与氮的变化情况相似, 数值在1.00~5.19 mg·L-1间波动.高锰酸盐指数在0.41~6.14 mg·L-1之间变化.透明度变化范围为0.10~4.00 m.ρ(Chla)整体在上游江段(S1~S4)最高, 富春江库区次之, 最大值在金华江S2点, 高达187.73 μg·L-1, 最低值在新安江, 为0.41 μg·L-1.Cond在105.91~660.01 μS·cm-1之间波动, 变化趋势与Chla相似, 反映出上游江段(S1~S4)水体受人类活动影响较强.

|
图 7 富春江水库水体理化因子空间变化 Fig. 7 Spatial variation in physical and chemical factors of Fuchunjiang Reservoir |
基于Bray-Curtis相似性系数的NMDS分析表明, 富春江水库浮游植物功能群的季节差异显著(P < 0.01)[图 8(a)].引起差异的主要功能群为P、C、MP、L0和H1, 共解释了83.61%的差异性, 其中P差异性贡献率高达34.32%, C功能群达到了20.9%, 其余3个最主要的功能群贡献率均在9%左右[图 8(b)].

|
图 8 富春江水库浮游植物功能群细胞密度NMDS分析及功能群差异贡献 Fig. 8 NMDS analysis and differential contribution of functional groups of phytoplankton in Fuchunjiang Reservoir |
RDA分析表明, 水温、浮游动物生物量、流量、高锰酸盐指数、TN和TP与浮游植物功能群结构关系密切(P < 0.05, 图 9).浮游动物与功能群H1、S1、L0、J、M和F呈显著正相关关系, 且这些功能群由绿藻和蓝藻物种组成;流量与功能群Y、D、X1、P和G呈显著正相关关系;功能群P与高锰酸盐指数和TP相关性最高, 功能群C与TN相关性较高.

|
(a)中红色表示解释变量环境因子, 黑色表示响应变量功能群 图 9 富春江水库浮游植物功能群与环境因子RDA分析 Fig. 9 RDA of phytoplankton functional groups and environmental factors in Fuchunjiang Reservoir |
环境因子对浮游植物功能群的相对重要性结果显示(图 10), 水温、浮游动物生物量、流量对富春江水库浮游植物最为重要, 相对重要性依次为8.28%、4.77%和4.01%(P < 0.05), 其次为TN、高锰酸盐指数和TP相对解释率较低, 且TP不显著.以上结果表明水温、浮游动物和流量是影响富春江水库浮游植物功能群的关键因子.浮游动物与富春江水库浮游植物功能群的密切联系说明良性生态系统构建的重要性.通过保护富春江渔业资源和增加滨河湿地等浮游动物庇护所, 可能有助于控制浮游植物的异常增殖.

|
蓝点无实际意义;对于每一列, 孤立红点表示各环境因子的边际效应, 多点间连线表示多个环境因子间的共同效应, 组分解释的变差百分比(来自变差分解)展示在(a)中;(b)为各环境因子的单独效应(来自层次分割), 其值等同于该环境因子的边际效应加上与其他环境因子的共同效应的平均分配值, ***表示P < 0.001, *表示P < 0.05 图 10 富春江水库环境因子对浮游植物功能群影响的相对重要性 Fig. 10 Relative importance of environmental factors on phytoplankton groups in Fuchunjiang Reservoir |
温度是富春江水库浮游植物季节演替的关键因素.本研究结果显示, 温度与夏季浮游植物细胞密度呈现显著正相关, 而与秋季、春季和冬季样点显著负相关[图 9(b)].夏季监测断面平均水温接近30℃(表 2), 此时优势功能群包含了代表蓝藻门的类群L0、S1、H1和M以及代表绿藻门的类群J, 且这5类功能群与温度呈显著正相关, 是夏季主要的优势功能群[图 10(a)].冬季监测断面平均水温为11.7℃, 此时绝对优势类群C细胞密度与温度呈现显著负相关.虽然环境因子相对重要性的结果显示温度和TN的共同影响最高(图 10), 但TN在季节上并未体现出显著的差异性, 且ρ(TN)为2.03 mg·L-1, 最高值达到5.93 mg·L-1, 体现出较高的浓营养本底, 因此浮游植物群落结构的变化不受TN的限制, 而受温度的影响更大.温度对浮游植物的影响主要体现在两个方面:浮游植物物种的温度适应性差异;浮游植物群落结构对温跃层的响应.
|
|
表 2 富春江水库水体温度和新安江流量季节变化 Table 2 Seasonal variation of water temperature and discharge in Fuchunjiang Reservoir |
不同浮游植物物种的温度适应性具有差异, 蓝藻适宜温度为30~35℃, 绿藻适宜温度为20~35℃, 而硅藻在0~15℃也能较好生长[16, 17].夏季较高的水温有利于浮游植物的大量繁殖, 藻类的增加会导致透明度降低, 这为适合在低光照混合浑浊型水体中生存的功能群S1提供了良好的条件.夏季较高的温度不仅促进蓝藻的生长和繁殖, 也提高了蓝藻与其他藻类间的竞争力[18].冬季低温为硅藻增值提供了有利条件, 冬季监测断面平均水温约为12℃, 变化范围为9.4~13.2℃(表 2), 此时水温与汉江冬春季暴发硅藻水华的水温8.4~13.2℃接近[19], 已经达到了硅藻水华暴发的温度条件.冬季C类群(小环藻)占据绝对优势, 可见富春江水库水体富营养化程度较高, 此时金华江、兰江和衢江藻细胞密度在(1.81~2.74)× 107 cells·L-1间波动, 已经接近汉江藻类水华暴发时的峰值5.3 × 107 cells·L-1.硅藻的异常增殖在其他水体也得到了关注, 任杰等[20]对横山水库的调查发现温度处于16~26℃时针杆藻达到了最佳生长.孙祥等[21]对天目沙河水库的调查结果显示, 小环藻在13℃时开始快速增殖, 到达19℃时达到峰值, 且在31℃之后又具有增殖趋势.小环藻的适宜温度范围较广, 在温度较低范围内也能大幅增长.可见, 金华江、兰江和衢江段极有可能在冬季低温条件下暴发硅藻水华.同时温度也是影响汝溪河[22]、渭河[23]和乌江[8]等河流浮游植物群落结构演替的主要因素, 因此密切关注温度变化, 对藻类水华的预测预警和有效防范具有积极作用.
温跃层对浮游植物功能群群落结构的季节和空间变化均有影响.高温会引起水体温度分层, 影响水体垂直方向营养物质的运输, 导致浮游植物群落结构的变化[21].且温跃层会随着气温的升高, 分层概率逐渐加大[24].温跃层在季节上具有明显差异:夏季富春江水库会出现明显的温跃层;秋季、冬季和春季未出现温跃层.而夏季L0、S1、H1和M占据优势, 温跃层的出现与功能群M和H1更加喜好分层水体的生境相吻合[25].而P类群对分层十分敏感, 在夏季优势度相对较低, 增加了L0、S1、H1和M等类群的竞争优势.空间上, 温跃层主要出现在富春江库区, 库区段L0、S1、H1和M等蓝藻类群也表现出较高的优势度, 且在库区段浮游植物的生长策略也较为多变.库区段会出现季节性的蓝藻占优, 而在上下游江段硅藻占据了绝对优势.据报道, 2016年富春江部分江段在持续32d温度高于30℃的情况下, 暴发了大规模的蓝藻水华[26];2020年梅雨后持续15 d温度高于30℃的情况下浮游植物从硅藻门占优演替为蓝藻门占优, 且持续的高温晴热天气促发了富春江库区明显的水体分层[27].可见, 夏季的高温会促发富春江库区段水体分层, 增加蓝藻水华暴发的风险.
3.2 水动力扰动条件差异对钱塘江浮游植物功能群时空分布影响大水文水动力条件是富春江水库浮游植物群落结构演替的关键, 对塑造浮游植物群落结构和功能至关重要.根据RDA结果[图 9(b)], 流量是影响富春江水库浮游植物群落结构的重要因素, 其与秋、冬和春季功能群细胞密度呈显著负相关.此前在对千岛湖[28]、Nakdong河[29]和Danube河[30]的研究中也表明了流量变化对浮游植物群落结构演替的重要性.这是因为流量季节间的改变能够带来水体流速和交换速率的变化, 进而影响浮游植物群落结构的演替.流量变化能够显著影响水华强度和空间动态变化[31].田晶等[32]发现汉江叶绿素a与流量呈现显著负相关, 且当最小7 d平均流量低于780 m3·s-1时, 就有水华暴发的风险;而在香溪河流量超过100 m3·s-1时藻类生物量显著降低[33];与此同时通过流量调控来预防水华的发生也成为了一项重要措施[34], 此前吴挺峰等[35]表示当富春江水库下泄流量达到600 m3·s-1时就能防止富春江水库的水华暴发.
流量是藻类出现异常增殖的诱因.调查期间, 夏季新安江日均流量为531 m3·s-1, 流量峰值为721 m3·s-1, 此时藻细胞密度峰值(1.32 × 107 cells·L-1)出现在富春江库区.2016年水华期间新安江的流量为537 m3·s-1[26].2020年夏季出现了蓝藻异常增殖的情况, 但并未发生水华, 这可能是夏季调查时间是在梅雨期后, 新安江水库首次九孔泄洪.一方面, 可能是梅雨期降雨量增高以及暴雨冲刷抑制了藻类疯长, 且降雨量累积直接导致流量增大[27];另一方面, 泄洪对藻类的生长和繁殖也产生一定的冲击.冬季的水体流量较低, 新安江同期流量为370 m3·s-1, 此时藻密度峰值达到了2.74 × 107 cells·L-1, 这是因为流量减小使得水体流速变慢, 水体交换速率也随之减小, 使得水体趋于稳定, 有助于藻类生长.此外, 水体交换速率会影响光可获得性从而影响浮游植物生长[36].且上述3个江段水体均较浅(金华江1.2 m;衢江4.8 m;兰江3.6 m), 水柱混合程度较完全, 水体中浮游植物对光的可获得性较高.虽然冬季透明度均有大幅降低(超过50%), 但这并不影响耐受低光照与低氮碳量的功能类群C大量增长[25].此外, 兰江及以上江段一直处于高营养盐状态下, ρ(TN)和ρ(TP)最高分别达到了5.93 mg·L-1和0.21 mg·L-1, 充分满足藻类的生长条件[37].因此, 营养盐充足、温度适宜和水动力条件合适是金华江、衢江和兰江段硅藻异常增殖的关键.
富春江库区与其上下游功能群组成差异显著(图 4), 流态变化对富春江水库浮游植物群落结构空间差异具有显著影响.冬季藻细胞密度峰值出现在金华江, 夏季峰值出现在三都大桥.春季在浮游植物密度整体较低的情况下, 峰值则出现在钱塘江河口.可见在河口-回水区、水流滞缓区更适宜藻类的生长和繁殖.根据张磊等[38]对三峡水库22条支流和干流断面的调查结果来看, 浮游植物细胞密度高值均在河口、回水区, 超过68%的河流集中在回水区发生水华.这是由于库区内的水动力条件更加利于河道藻类的生长和繁殖[39].
3.3 浮游动物是影响浮游植物群落结构变化的重要因子浮游动物是影响富春江水库浮游植物群落结构的重要因子.浮游动物对富春江水库浮游植物功能群的相对重要性为4.01%, 位居第二(图 10).浮游动物生物量与浮游植物群落结构在时空变化上表现出一定同步性与关联性.在夏冬两次浮游植物的异常增殖江段, 浮游植物生物量也相对较高.而充足的食物可以提高植食性浮游动物的生长速率[40], 藻类异常增殖, 为浮游动物提供了充足的食物, 促进了浮游动物生物量的同步升高.
浮游动物的群落组成、细胞密度和选择性摄食也会影响浮游植物的群落结构, 对抑制浮游植物生长具有积极作用[40, 41].结果显示浮游动物生物量与功能群H1、S1、L0、J、M和F呈显著正相关;浮游动物与夏季浮游植物功能群呈现显著正相关, 而在春季、秋季和冬季部分江段呈现一定负相关关系, 表明浮游动物对浮游植物群落结构变化显著相关.
功能群划分结果显示, X1、X2、W1和Y这4类表征对摄食作用敏感、适宜在牧食强度低的环境中增殖的功能群偶有出现在部分江段, 但优势度均不高.据此推测在富春江水库浮游动物对浮游植物具有一定的摄食作用.以沟链藻、直链藻为代表的功能群P、以微囊藻为代表的功能群M和以长孢藻为代表的功能群H1等由大型或群体藻种为代表的功能群相继占据优势, 这可能是浮游动物选择性摄食的结果.富春江水库占据绝对优势的是以沟链藻为主的功能群P, 有研究表明沟链藻可形成弯曲结构, 有效躲避摄食[42], 这也间接证明了在富春江水库浮游动物促进了浮游植物群落结构向不易被牧食的较大群体和群体种类占优转变.
调查江段浮游动物生物量为0.42 mg·L-1, 高于汉江干流(0.25 mg·L-1)[43]、湘江干流(0.24 mg·L-1)[44]、荆江(0.09 mg·L-1)[45]和渭河(0.01 mg·L-1)[22], 说明富春江水库浮游动物生物量处于较高水平.此外, 浮游动物与浮游植物生物量之比(BZP∶BPhyt)对衡量水体营养状况具有重要作用[46].调查期间, 富春江水库BZP∶BPhyt为0.17, 浮游植物生物量(2.45 mg·L-1)为浮游动物生物量6倍.BZP∶BPhyt接近汉江干流(0.16)[43], 高于荆江(0.10)[45], 低于湘江干流(1.41)[44].且在荆江段BZP∶BPhyt为0.10时, 浮游植物与浮游动物密切相关[45], 而富春江水库BZP∶BPhyt相对较高, 由此推测富春江水库浮游动物对浮游植物也具有一定捕食作用.
浮游动物通过捕食作用影响浮游植物, 同时也受到鱼类的影响.有研究表明, 上游兰江段鱼类优势种以鲫鱼和似鳊等中小型的杂食性底栖鱼类为主, 下游江段光泽黄颡鱼和增殖放流鱼种鲢和鳙是主要的优势种群, 且鱼类资源呈现小型化趋势[47, 48].这一现象增加了鱼类对大型浮游动物的捕食作用, 降低浮游动物对浮游植物的牧食效率, 不利于控制藻类的细胞密度.这与本研究中浮游动物优势种以中小型为主的结果相一致, 富春江水库偶发蓝藻水华可能也与之相关.因此, 提高富春江水库生态系统结构与功能、良性生态系统构建具有重要意义.通过优化鱼类捕捞方式和保护富春江渔业资源、调控浮游动物的群落结构和恢复大型浮游动物种类、增加滨河湿地等浮游动物庇护所, 可能有助于控制浮游植物的异常增殖.
4 结论(1)富春江水库浮游植物功能群可分为18个功能类群, 包含10个优势类群, 对水质的时空变化具有重要指示作用.根据C-R-S生长策略结果表明, 富春江水库总体具有高干扰、低胁迫的环境特征, 且水质、水文具有多变性.
(2)富春江水库浮游植物功能群具有显著的季节差异.夏季生境多变、生产力旺盛, 功能群优势类群最为丰富, 为P+L0+J+M+S1+H1+MP型;秋季水量减少、流态趋于静止演替为P+H1型;冬季换水减弱、富营养化问题加重, 演替为P+C型;春季生境多变, 演替为C+D+P+MP型.
(3)富春江水库浮游植物功能群呈现丰富多变的空间分布格局.上游江段是中富营养混合水体, 主要以小环藻为代表的C类群和以沟链藻为代表的P类群为主;中游富春江库区水深加大, 仍以P类群为主, 但沿程优势度逐渐减小, 反映出库区水深加大、分层稳定性增强的生态效应;随着河流扰动加剧, 冲刷作用的增强, 富春江大坝至下游江段MP类群逐渐占优, 且在钱塘江口占绝对优势.
(4)温度和流态是富春江水库浮游植物功能群时空差异的关键因子, 浮游动物对浮游植物功能群影响显著, 而营养盐影响较小.水文气象过程对富春江水库浮游植物群落结构影响至关重要:夏季高温季节流态变化可能导致富春江库区暴发蓝藻水华;而在冬季低温低流量下, 金华江、衢江和兰江等江段则有硅藻水华暴发的风险;降低藻类水华的风险, 仍需加大对河流氮磷负荷的控制, 结合浮游动物群落调控, 并充分考虑水利调度途径的协同.
| [1] | Moorhouse H L, Read D S, Mcgowan S, et al. Characterisation of a major phytoplankton bloom in the River Thames (UK) using flow cytometry and high performance liquid chromatography[J]. Science of the Total Environment, 2018, 624: 366-376. DOI:10.1016/j.scitotenv.2017.12.128 |
| [2] | Hou X J, Feng L, Dai Y H, et al. Global mapping reveals increase in lacustrine algal blooms over the past decade[J]. Nature Geoscience, 2022, 15(2): 130-134. DOI:10.1038/s41561-021-00887-x |
| [3] | Tian Y L, Jiang Y, Liu Q, et al. The impacts of local and regional factors on the phytoplankton community dynamics in a temperate river, Northern China[J]. Ecological Indicators, 2021, 123. DOI:10.1016/j.ecolind.2021.107352 |
| [4] | Reynolds C S. What factors influence the species composition of phytoplankton in lakes of different trophic status?[J]. Hydrobiologia, 1998, 369-370: 11-26. DOI:10.1023/A:1017062213207 |
| [5] |
陈倩, 李秋华, 马欣洋, 等. FG、MFG和MBFG浮游植物功能群的比较: 以贵州三座水库为例[J]. 环境科学, 2019, 40(9): 4061-4071. Chen Q, Li Q H, Ma X Y, et al. Comparison of functional groups of phytoplankton in FG, MFG, and MBFG: taking three reservoirs as an example in Guizhou plateau[J]. Environmental Science, 2019, 40(9): 4061-4071. |
| [6] | Kim H G, Hong S, Kim D K, et al. Drivers shaping episodic and gradual changes in phytoplankton community succession: taxonomic versus functional groups[J]. Science of the Total Environment, 2020, 734. DOI:10.1016/j.scitotenv.2020.138940 |
| [7] | Sun X, Mwagona P C, Shabani I E, et al. Phytoplankton functional groups response to environmental parameters in Muling River Basin of Northeast China[J]. Annales de Limnologie - International Journal of Limnology, 2019, 55. DOI:10.1051/limn/2019014 |
| [8] |
肖晶, 王宝利, 张海涛, 等. 乌江河流-水库体系浮游植物功能群演替及其环境影响因子辨识[J]. 地球与环境, 2019, 47(6): 829-838. Xiao J, Wang B L, Zhang H T, et al. Succession of phytoplankton functional groups in the Wujiang River-reservoir system and its environmental impact factors identification[J]. Earth and Environment, 2019, 47(6): 829-838. |
| [9] |
葛优, 周彦锋, 王晨赫, 等. 阳澄西湖浮游藻类功能群演替特征及其与环境因子的关系[J]. 中国环境科学, 2019, 39(7): 3027-3039. Ge Y, Zhou Y F, Wang C H, et al. Succession patterns of phytoplankton functional groups in western area of Yangcheng Lake and their relationship with environmental factors[J]. China Environmental Science, 2019, 39(7): 3027-3039. DOI:10.3969/j.issn.1000-6923.2019.07.040 |
| [10] |
郑诚, 陆开宏, 徐镇, 等. 四明湖水库浮游植物功能类群的季节演替及其影响因子[J]. 环境科学, 2018, 39(6): 2688-2697. Zheng C, Lu K H, Xu Z, et al. Seasonal succession of phytoplankton functional groups and their driving factors in the Siminghu Reservoir[J]. Environmental Science, 2018, 39(6): 2688-2697. |
| [11] |
杨丽, 张玮, 尚光霞, 等. 淀山湖浮游植物功能群演替特征及其与环境因子的关系[J]. 环境科学, 2018, 39(7): 3158-3167. Yang L, Zhang W, Shang G X, et al. Succession characteristics of phytoplankton functional groups and their relationships with environmental factors in Dianshan Lake, Shanghai[J]. Environmental Science, 2018, 39(7): 3158-3167. |
| [12] |
史鹏程, 朱广伟, 杨文斌, 等. 新安江水库悬浮颗粒物时空分布、沉降通量及其营养盐效应[J]. 环境科学, 2020, 41(5): 2137-2148. Shi P C, Zhu G W, Yang W B, et al. spatial-temporal distribution of suspended solids and its sedimentation flux and nutrients effects in Xin'anjiang Reservoir, China[J]. Environmental Science, 2020, 41(5): 2137-2148. |
| [13] | Reynolds C S, Huszar V, Kruk C, et al. Towards a functional classification of the freshwater phytoplankton[J]. Journal of Plankton Research, 2002, 24(5): 417-428. DOI:10.1093/plankt/24.5.417 |
| [14] | Padisák J, Crossetti L O, Naselli-Flores L. Use and misuse in the application of the phytoplankton functional classification: a critical review with updates[J]. Hydrobiologia, 2009, 621(1): 1-19. DOI:10.1007/s10750-008-9645-0 |
| [15] | Lai J S, Zou Y, Zhang J L, et al. Generalizing hierarchical and variation partitioning in multiple regression and canonical analyses using the rdacca. hp R package[J]. Methods in Ecology and Evolution, 2022, 13(4): 782-788. DOI:10.1111/2041-210X.13800 |
| [16] |
朱广伟, 金颖薇, 任杰, 等. 太湖流域水库型水源地硅藻水华发生特征及对策分析[J]. 湖泊科学, 2016, 28(1): 9-21. Zhu G W, Jin Y W, Ren J, et al. Characteristics of diatom blooms in a reservoir-water supply area and the countermeasures in Taihu Basin, China[J]. Journal of Lake Sciences, 2016, 28(1): 9-21. |
| [17] |
程兵芬, 夏瑞, 张远, 等. 基于拐点分析的汉江水华暴发突变与归因研究[J]. 生态环境学报, 2021, 30(4): 787-797. Cheng B F, Xia R, Zhang Y, et al. The evolution and mutation attribution of water bloom in Hanjiang River based on mutation test method[J]. Ecology and Environmental Sciences, 2021, 30(4): 787-797. |
| [18] | Mcnaughton S J. Relationships among functional properties of californian grassland[J]. Nature, 1967, 216(5111): 168-169. |
| [19] |
辛小康, 王英才, 胡圣, 等. 2018年汉江下游硅藻水华成因分析[J]. 水电能源科学, 2019, 37(3): 25-28. Xin X K, Wang Y C, Hu S, et al. Cause analysis of diatom bloom of lower reaches of Hanjiang River in 2018[J]. Water Resources and Power, 2019, 37(3): 25-28. |
| [20] |
任杰, 朱广伟, 金颖薇, 等. 换水率和营养水平对太湖流域横山水库硅藻水华的影响[J]. 湖泊科学, 2017, 29(3): 604-616. Ren J, Zhu G W, Jin Y W, et al. Combined effects of water exchange rate and nutrient on diatom proliferation in Hengshan Reservoir, Taihu Basin[J]. Journal of Lake Sciences, 2017, 29(3): 604-616. |
| [21] |
孙祥, 朱广伟, 笪文怡, 等. 水文气象因素对东南山区水库硅藻异常增殖的影响[J]. 环境科学, 2018, 39(3): 1129-1140. Sun X, Zhu G W, Da W Y, et al. Effects of hydrological and meteorological conditions on diatom proliferation in reservoirs[J]. Environmental Science, 2018, 39(3): 1129-1140. |
| [22] |
向蓉, 李巧玉, 喻燚, 等. 汝溪河浮游硅藻功能群特征及其与环境因子相关性分析[J]. 环境科学, 2017, 38(8): 3290-3301. Xiang R, Li Q Y, Yu Y, et al. Functional group characteristics of planktonic diatoms and their relationship with environmental factors in the Ruxi River[J]. Environmental Science, 2017, 38(8): 3290-3301. |
| [23] |
赵耿楠, 潘保柱, 丁一桐, 等. 渭河干流和秦岭北麓典型支流浮游植物功能群特征及水质评价[J]. 生态学报, 2021, 41(8): 3226-3237. Zhao G N, Pan B Z, Ding Y T, et al. Characteristics and water quality evaluation of phytoplankton functional groups in the Weihe River mainstem and its tributaries in the northern foot of the Qinling Mountains[J]. Acta Ecologica Sinica, 2021, 41(8): 3226-3237. |
| [24] |
笪文怡, 朱广伟, 黎云祥, 等. 新安江水库河口区水质及藻类群落结构高频变化[J]. 环境科学, 2020, 41(2): 713-727. Da W Y, Zhu G W, Li Y X, et al. High-Frequency dynamics of water quality and phytoplankton community in inflowing river mouth of Xin'anjiang Reservoir, China[J]. Environmental Science, 2020, 41(2): 713-727. |
| [25] |
胡韧, 蓝于倩, 肖利娟, 等. 淡水浮游植物功能群的概念、划分方法和应用[J]. 湖泊科学, 2015, 27(1): 11-23. Hu R, Lan Y Q, Xiao L J, et al. The concepts, classification and application of freshwater phytoplankton functional groups[J]. Journal of Lake Sciences, 2015, 27(1): 11-23. |
| [26] | Guo C X, Zhu G W, Paerl H W, et al. Extreme weather event may induce Microcystis blooms in the Qiantang River, Southeast China [J]. Environmental Science and Pollution Research, 2018, 25(22): 22273-22284. DOI:10.1007/s11356-018-2216-7 |
| [27] |
张萍, 国超旋, 俞洁, 等. 钱塘江干流夏季浮游植物群落结构特征及其对水文气象的响应[J]. 湖泊科学, 2022, 34(2): 418-432. Zhang P, Guo C X, Yu J, et al. Characteristics of phytoplankton community structure and its response to hydro-meteorology in summer of Qiantang River[J]. Journal of Lake Sciences, 2022, 34(2): 418-432. |
| [28] |
笪文怡, 朱广伟, 吴志旭, 等. 2002—2017年千岛湖浮游植物群落结构变化及其影响因素[J]. 湖泊科学, 2019, 31(5): 1320-1333. Da W Y, Zhu G W, Wu Z X, et al. Long-term variation of phytoplankton community and driving factors in Qiandaohu Reservoir, Southeast China[J]. Journal of Lake Sciences, 2019, 31(5): 1320-1333. |
| [29] | Kim H G, Recknagel F, Kim H W, et al. Implications of flow regulation for habitat conditions and phytoplankton populations of the Nakdong River, South Korea[J]. Water Research, 2021, 207. DOI:10.1016/j.watres.2021.117807 |
| [30] | Rusanov A G, Bíró T, Kiss K T, et al. Relative importance of climate and spatial processes in shaping species composition, functional structure and beta diversity of phytoplankton in a large river[J]. Science of the Total Environment, 2022, 807. DOI:10.1016/j.scitotenv.2021.150891 |
| [31] | Park Y, Pyo J, Kwon Y S, et al. Evaluating physico-chemical influences on cyanobacterial blooms using hyperspectral images in inland water, Korea[J]. Water Research, 2017, 126: 319-328. DOI:10.1016/j.watres.2017.09.026 |
| [32] |
田晶, 郭生练, 王俊, 等. 汉江中下游干流水华关键环境因子识别及阈值分析[J]. 水资源保护, 2022, 38(5): 196-203. Tian J, Guo S L, Wang J, et al. Identification and threshold analysis of key factors of algal bloom in middle and lower reaches of the Hanjiang River[J]. Water Resources Protection, 2022, 38(5): 196-203. |
| [33] |
黄宇波, 范向军, 杨霞, 等. 水文气象因素对香溪河藻类动态的影响[J]. 中国农村水利水电, 2023(1): 1-7. Huang Y B, Fan X J, Yang X, et al. The effects of hydro-meteorological factors on the algae dynamics in Xiangxi River[J]. China Rural Water and Hydropower, 2023(1): 1-7. |
| [34] |
闫雪燕, 张鋆, 李玉英, 等. 动态调水过程水文和理化因子共同驱动丹江口水库库湾浮游植物季节变化[J]. 湖泊科学, 2021, 33(5): 1350-1363. Yan X Y, Zhang Y, Li Y Y, et al. Hydrologic and physicochemical factors co-drive seasonal changes of phytoplankton during dynamic water diversion processes in the Danjiangkou Reservoir[J]. Journal of Lake Sciences, 2021, 33(5): 1350-1363. |
| [35] |
吴挺峰, 高光, 晁建颖, 等. 基于流域富营养化模型的水库水华主要诱发因素及防治对策[J]. 水利学报, 2009, 40(4): 391-397. Wu T F, Gao G, Chao J Y, et al. Analysis on factors inducing algal bloom in reservoir based on basin eutrophication model[J]. Journal of Hydraulic Engineering, 2009, 40(4): 391-397. |
| [36] |
郭芳, 顾继光, 赵剑, 等. 类群划分方法对南亚热带水库夏季浮游植物群落与环境响应关系的影响[J]. 环境科学, 2020, 41(11): 5050-5059. Guo F, Gu J G, Zhao J, et al. Effects of phytoplankton classifications on the relationship between phytoplankton community and environment in summer subtropical reservoirs, Southern China[J]. Environmental Science, 2020, 41(11): 5050-5059. |
| [37] | Xu H, Paerl H W, Qin B, et al. Determining critical nutrient thresholds needed to control harmful cyanobacterial blooms in eutrophic Lake Taihu, China[J]. Environmental Science & Technology, 2015, 49(2): 1051-1059. |
| [38] |
张磊, 夏志强, 周伟, 等. 三峡水库春季营养盐和浮游植物空间分布及其影响机制[J]. 环境科学研究, 2015, 28(7): 1069-1077. Zhang L, Xia Z Q, Zhou W, et al. Spatial distribution of nutrients and phytoplankton and causes for their differences in Three Gorges Reservoir in spring[J]. Research of Environmental Sciences, 2015, 28(7): 1069-1077. |
| [39] |
张耀文, 李洪, 李嘉, 等. 西南山区典型河道型水库藻类功能群时空演替特征及其影响因素: 以紫坪铺水库为例[J]. 环境科学, 2018, 39(6): 2680-2687. Zhang Y W, Li H, Li J, et al. Spatiotemporal succession characteristics of algal functional groups and its impact factors for a typical channel-type reservoir in a southwest mountainous area[J]. Environmental Science, 2018, 39(6): 2680-2687. |
| [40] |
陈辉煌, 王文平, 薛媛媛, 等. 食物与桡足类营养级对亚热带分层水库浮游动物群落结构的影响[J]. 湖泊科学, 2022, 34(1): 174-183. Chen H H, Wang W P, Xue Y Y, et al. Effect of food and copepod trophic level on zooplankton community structure in a subtropical stratified reservoir[J]. Journal of Lake Sciences, 2022, 34(1): 174-183. |
| [41] | Dos Santos Severiano J, Dos Santos Almeida-Melo V L, De Melo-Magalhães E M, et al. Effects of zooplankton and nutrients on phytoplankton: an experimental analysis in a eutrophic tropical reservoir[J]. Marine and Freshwater Research, 2017, 68(6): 1061-1069. |
| [42] |
葛大艳, 刘乾甫, 赖子尼, 等. 珠三角河网链状硅藻物种组成及生态特征[J]. 生态学报, 2021, 41(6): 2482-2495. Ge D Y, Liu Q F, Lai Z N, et al. Species composition and ecological characteristics of filamentous diatoms in the Pearl River Delta[J]. Acta Ecologica Sinica, 2021, 41(6): 2482-2495. |
| [43] |
杜红春. 汉江干流浮游生物群落结构和功能群特征及水质评价[D]. 武汉: 华中农业大学, 2020. DuH C. Community structure and functional groups of plankton and water quality evaluation in Hanjiang River[D]. Wuhan: Huazhong Agricultural University, 2020. |
| [44] |
王晓清, 曾亚英, 吴含含, 等. 湘江干流浮游生物群落结构及水质状况分析[J]. 水生生物学报, 2013, 37(3): 488-494. Wang X Q, Zeng Y Y, Wu H H, et al. Investigation on plankton community composition and water quality state in the mainstream of Xiangjiang River[J]. Acta Hydrobiologica Sinica, 2013, 37(3): 488-494. |
| [45] |
郭杰, 王珂, 段辛斌, 等. 长江荆江段浮游生物群落结构特征及其与环境因子的关系[J]. 长江流域资源与环境, 2020, 29(9): 1954-1964. Guo J, Wang K, Duan X B, et al. Characteristics of plankton community and its relationships with environmental factors in the Jingjiang reach of the Yangtze River[J]. Resources and Environment in the Yangtze Basin, 2020, 29(9): 1954-1964. |
| [46] | García-Chicote J, Armengol X, Rojo C. Zooplankton abundance: a neglected key element in the evaluation of reservoir water quality[J]. Limnologica, 2018, 69: 46-54. |
| [47] |
郝雅宾, 刘金殿, 张爱菊, 等. 钱塘江水系兰溪段鱼类资源现状[J]. 水产科学, 2019, 38(4): 555-562. Hao Y B, Liu J D, Zhang A J, et al. Current status of fishery resources in Lanxi section of Qiantang River drainage[J]. Fisheries Science, 2019, 38(4): 555-562. |
| [48] |
郝雅宾, 刘金殿, 张爱菊, 等. 钱塘江下游江段鱼类资源现状[J]. 浙江农业学报, 2017, 29(10): 1620-1629. Hao Y B, Liu J D, Zhang A J, et al. Current status of fishery resources in downstream section of Qiantang River[J]. Acta Agriculturae Zhejiangensis, 2017, 29(10): 1620-1629. |