 2023, Vol. 44
2023, Vol. 44



土壤侵蚀是地球表层物质迁移过程之一, 在全球变化和人为活动日益增强的背景下, 严重的土壤侵蚀使耕地面积减少、土壤质量退化, 引起一系列环境问题[1].据报道, 云南省坡耕地水土流失面积为453.37万hm2, 占全省水土流失面积的43.29%; 近年来, 由于特殊的降雨、地貌特征及不合理的耕作导致云南省坡耕地水土流失愈加严重[2].因此, 对云南省滇中红壤区坡耕地进行抗蚀机制的研究将有助于改善这一状况.
土壤团聚体是土壤结构的基本单元, 其数量和大小影响土壤径流和蓄水渗透的强弱, 稳定性可反映土壤抗蚀和抗冲能力的大小[3], 在维持土壤生态功能和保持土壤肥力方面发挥着重要作用[4].土壤团聚体的形成和稳定性受生物和非生物因素的影响[5].在非生物因子中, 轮作休耕不仅有利于耕地休养生息、保持水土和农业可持续发展, 而且有利于土壤团聚体组成和稳定[6, 7].如夏梓泰等[8]研究发现, 与玉米单作相比, 长期休闲和绿肥轮作玉米可提高土壤大团聚体含量和稳定性; Tiemann等[9]研究表明大豆-小麦-玉米覆盖作物轮作中的水稳性团聚体高于单作玉米.在生物因子中, 丛枝菌根真菌(AMF)是一种能够与地球上约80%的植物根系建立共生关系的真菌, 也是土壤微生物群落的关键组成部分和参与农业可持续发展的重要微生物群之一[10, 11], 对于改善土壤生物环境、减少土壤侵蚀和提高土壤质量至关重要[12].已有研究发现, 覆盖作物或休耕的引入影响了后茬经济作物的AMF定殖, 而AMF可以通过提高菌丝体对土壤养分的吸收来刺激作物的生长, 该过程可促进AMF分泌GRSP等有机化合物, 促进土壤团聚体的形成, 从而保护土壤有机碳不被分解[13, 14].此外, 也有研究表明AMF可以促进根系分泌物(胞外多糖和根系黏液)的产生, 这些分泌物可以将土壤颗粒黏结在一起, 从而有利于大团聚体的形成和稳定[15].Zhang等[16]研究发现, 与岩溶土壤系统中没有AMF定殖的土壤相比, AMF定殖显著增加了土壤大团聚体的含量(>2 mm).Zhang等[17]研究表明, 间作系统通过改变土壤养分和AMF的多样性, 影响土壤团聚体的形成和稳定性.Wright等[18]对北美大平原地区的小麦休耕、小麦-玉米-小米和小麦-玉米-小米-休耕等进行试验后发现轮作显著提高了土壤中AMF数量.尽管最新研究表明轮作休耕可增加团聚体稳定性, AMF有利于团聚体组成和稳定, 但关于在轮作休耕条件下AMF群落对团聚体形成和稳定的影响研究鲜见报道.
因此, 本试验以坡耕地红壤为研究对象, 设置苕子轮作玉米(V-C)、豌豆轮作玉米(P-C)、冬闲-玉米(F-C)和周年休耕(F-F)这4个处理, 分析土壤养分、AMF群落和团聚体在不同处理下的变化, 采用冗余分析进一步分析AMF对团聚体组成和稳定性的影响, 以期为当地推行合理的轮作休耕模式及增强耕地持续利用提供一定的理论依据.
1 材料与方法 1.1 试验地概况本试验田间试验于云南省昆明市盘龙区松华街道大摆村(25°02′29″N, 102°58′39″E)进行.试验地坡度为10°, 属亚热带高原季风气候, 海拔为2 234 m, 年降水量为900~1 000 mm, 年平均气温16℃.长期定位试验于2018年开始, 本试验于2020年10月开始.其土壤基本理化性质为:pH 5.20, ω[有机质(SOM)]30.15 g·kg-1, ω[碱解氮(AN)]115.3 mg·kg-1, ω[速效磷(AP)]6.41 mg·kg-1, ω[速效钾(AK)]92.36 mg·kg-1.
1.2 试验材料与设计本试验小区面积为4 m×5 m, 设置苕子轮作玉米(V-C)、豌豆轮作玉米(P-C)、冬闲-玉米(F-C)和周年休耕(F-F) 这4个处理, 每个处理3次重复, 共12个小区, 区组内小区随机排列.供试作物:玉米(云瑞“88”)、光叶紫花苕和白花矮茎豌豆.其中, V-C和P-C于2020年10月种植苕子、豌豆, 翌年4月份苕子和豌豆翻压后种植玉米; F-C于2020年10月玉米收获后进行休闲, 翌年4月种植玉米; 休耕地则为休闲处理, 休耕地生长的植物主要有马唐(Digitaria sanguinalis)、酢浆草(Oxalis corniculata)、风轮菜(Clinopodium chinense)、圆叶锦葵(Malva rotundifolia)、菊叶香藜(Chenopodium foetidum)、秋鼠麴草(Gnaphalium hypoleucum)和飞蓬(Erigeron acer)等.
玉米采用宽窄行种植, 宽和窄行行距分别为80 cm和40 cm, 株间距为25 cm.以穴播的方式播种, 每穴播两粒, 间苗时每穴留1株, 单个小区植株共计9行, 每行16株; 苕子和豌豆采用撒播方式种植, 将种子均匀撒于已收获玉米的轮作处理小区中, 播种深度为2~3 cm.玉米进行单株精准施肥, 施N 315 kg·hm-2, P2 O5 120 kg·hm-2, K2 O 120 kg·hm-2, 其中氮肥分两次(50%做基肥, 50%做追肥), 并在喇叭口期追肥, 磷肥和钾肥全部做基肥施用; 苕子和豌豆作为绿肥作物, 不做施肥处理.玉米在播种、施肥和灌水后用薄膜覆盖, 后期根据作物的长势, 适时进行灌水、除草等作业.在轮作完豌豆及苕子后对其进行翻压处理, 苕子翻压还田生物量鲜重为10 300.13 kg·hm-2, 干重为2 631.20 kg·hm-2; 豌豆还田量鲜重为9 738.13 kg·hm-2, 干重为2 193.87 kg·hm-2, 休耕地不进行耕作等田间管理.
1.3 样品采集在2021年玉米成熟期用五点取样法采集0~20 cm土层的土样, 并将土样按其自然裂缝掰开, 拣去土样中的石块和植物根系等杂物, 为避免运输过程中土样被挤压和扰动, 将一部分土样装入塑料盒内, 以保持土壤原状结构, 并于试验条件下风干后用于土壤理化性质和团聚体的测定; 另一部分土样装入10 mL无菌离心管中并装在冰盒内, 迅速运回实验室并转移到-80℃超低温冰箱中保存, 用于提取土壤DNA.
1.4 样品测定 1.4.1 土壤基本理化性质测定土壤pH采用1∶2.5(土∶水)浸提, pH计测定; 土壤有机质测定采用重铬酸钾-外加热法; 全氮测定采用凯氏定氮法; 碱解氮采用碱解扩散法; 全磷测定采用碱熔-钼锑抗比色法; 速效磷测定采用碳酸氢钠浸提-光电比色法; 速效钾测定采用乙酸铵浸提-火焰光度法[19].
1.4.2 土壤团聚体测定及指标计算采用Elliott团聚体湿筛法[20]测定>2 mm、2~1 mm、1~0.25 mm和<0.25 mm这4个粒径的水稳性团聚体, 团聚体稳定性指标计算公式如下[21].
>0.25 mm水稳性团聚体含量(R0.25):
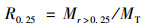
|
(1) |
式中, Mr>0.25为粒径>0.25 mm水稳性团聚体质量(g); MT为水稳性团聚体总质量(g).
平均重量直径(MWD):

|
(2) |
式中, xi为第i级水稳性团聚体平均直径(mm); ωi为第i级水稳性团聚体质量分数(%).
1.4.3 土壤DNA提取、PCR扩增及生物信息学分析按照E.Z.N.A.soil试剂盒操作规范抽提土壤DNA, 用1%的琼脂糖凝胶电泳检测DNA的提取质量.PCR采用TransGen AP221-02:TransStart Fastpfu DNA Polymerase; 全部样本按照正式试验条件进行, 每个样本3个重复, 将同一样本的PCR产物混合后用2%琼脂糖凝胶电泳检测, 使用AxyPrepDNA凝胶回收试剂盒(AXYGEN公司)切胶回收PCR产物, 用Tris_HCl缓冲液进行洗脱, 并用2%琼脂糖电泳检测; 使用NanoDrop 2000测定DNA浓度和纯度.所有样品的AMFDNA采用巢式PCR扩增, 第一对引物:AML1F和AML2R, 第二对引物:AMV4-5NF和AMDGR.参照电泳初步定量结果, 将PCR产物用QuantiFluorTM -ST蓝色荧光定量系统(Promega公司)进行检测定量, 然后按照每个样本的测序量要求, 进行相应比例的混合, 测序在MiSeq PE 300平台进行(上海美吉生物医药科技有限公司).
利用Flash 1.2.11进行pair-end双端序列拼接, 用Uparse 11进行OTU聚类, 将所有样品基于序列数41 499进行抽平, 以抽平后的OTU丰度表进行后续所有分析计算.使用Uparse 11在相似度为97%的水平上进行OTU聚类, 为了得到每个OTU对应的物种分类信息, 采用RDP classifier贝叶斯算法对97%相似水平的OTU代表序列进行分类学分析, 置信度阈值为0.7, 使用Mothur(version v.1.30.2)进行α多样性分析.
1.5 数据处理利用Microsoft Excel 2016软件进行数据的整理.采用SPSS 23.0进行统计分析; 利用单因素方差分析(One-way ANOVA)评价不同处理间各个指标的差异显著性; 用Origin 2021软件进行绘图; 采用Canoco 5.0软件进行冗余分析; 使用R语言(version 3.3.1)进行Spearman相关性分析.
2 结果与分析 2.1 轮作休耕模式下土壤团聚体组成及稳定性特征不同轮作休耕处理对各粒级土壤团聚体含量及稳定性的影响各异(表 1).V-C和F-F处理的>2 mm团聚体含量较F-C显著提高了29.71%和66.99%; V-C、P-C和F-F的R0.25较F-C处理显著提高了20.29%、16.80%和33.47%, MWD较F-C处理显著增加了23.38%、19.48%和44.16%, 而<0.25 mm团聚体含量较F-C处理显著降低了32.89%、27.23%和35.06%. 2~0.25 mm团聚体含量在各处理之间无显著差异.另外, 各处理的R0.25、MWD由大到小的顺序为:F-F>V-C>P-C>F-C, 其中F-F处理与其他处理间的差异性达显著水平(P<0.05).
|
|
表 1 轮作休耕模式下团聚体组成与稳定性1) Table 1 Aggregate composition and stability under crop rotation and fallow pattern |
2.2 轮作休耕模式下土壤养分的变化特征
不同轮作休耕处理下的土壤养分变化各异(表 2).V-C和F-C处理的pH显著高于F-F处理(P<0.05); P-C和F-F处理下的SOM含量显著高于F-C处理(P<0.05); F-F、V-C和P-C的AN含量较F-C显著提高了22.52%、9.01%和11.51%; P-C处理的AP含量较F-C处理的显著降低27.79%(P<0.05); F-F处理下的AK含量较F-C显著降低19.83%(P<0.05).
|
|
表 2 休耕轮作模式下土壤养分的变化特征1) Table 2 Characteristics of soil nutrient changes under crop rotation and fallow pattern |
2.3 轮作休耕模式下AMF多样性的变化特征
本研究共获得629 128条原始序列, 经过筛选后获得497 988条有效序列, 占总序列数的79.16%. 测序量达到40 000时, OTU数基本处于饱和状态(图 1).测序结果基本能够反映当前样本的多样性, 各组AMF的测序数据量基本合理, 但后续研究中需要增加测序深度.本试验共获得78个OTU数目(图 2), 从门到种依次进行分类为1门、1纲、5目、7科、11属和27种.V-C、P-C、F-C和F-F中所包含的OTU数分别为55、56、56和70个; 共有的OTU数为36个; 特有的OTU数分别为1、4、1和8个; F-C与F-F共有的OTU数为53.由此可以看出, F-C与F-F之间比较接近.

|
图 1 轮作休耕模式下AMF的稀释曲线 Fig. 1 Rarefaction curve of soil AMF under crop rotation and fallow pattern |

|
图 2 轮作休耕模式下AMF OTU分布的维恩图 Fig. 2 Venn diagram of OTUs distribution of soil AMF under crop rotation and fallow pattern |
不同轮作休耕处理对AMF丰富度和多样性的影响不同(表 3).其中, F-F处理下的Chao1和ACE指数较F-C显著提高35.78%和29.56%, V-C和P-C处理的Chao1、ACE和Shannon指数较F-C有增高的趋势, F-F处理下的Shannon指数较F-C处理显著提高45.55%. 结果表明, F-F处理的AMF群落的丰富度及多样性显著高于F-C(P<0.05).
|
|
表 3 轮作休耕模式对AMF丰富度和多样性的影响 Table 3 Effects of crop rotation and fallow pattern on soil AMF richness and diversity |
2.4 轮作休耕模式对AMF属水平上群落组成的影响
在属水平上对不同处理下的AMF群落组成进行分析得出(图 3), AMF群落各属OTU数目占总OTU数目的比例大小顺序为球囊霉属(Glomus)、无梗囊霉属(Acaulospora)、类球囊霉属(Paraglomus)、巨孢囊霉属(Gigaspora)、大孢子科未分类属(Unclassified_ f_ _Gigasporaceae)、盾巨孢囊霉属(Scutellospora), 其中球囊霉属为优势属, 盾巨孢囊霉属在各处理间差异显著(P<0.05).

|
Glomus:球囊霉属; Unclassified_ f_ _Gigasporaceae:大孢子科未分类属; Acaulospora:无梗囊霉属; Paraglomus:类球囊霉属; Gigaspora:巨孢囊霉属; Scutellospora:盾巨孢囊霉属; 右侧数值为P值, *表示P<0.05 图 3 轮作休耕模式下AMF群落结构(属水平)分布特征 Fig. 3 Distribution characteristics of AMF community structure (genus level) under crop rotation and fallow pattern |
基于Bray-Curtis距离算法的主坐标分析(PCoA)表明(图 4), PC1和PC2轴分别解释了29.99%和22.40%的AMF群落组成差异, 差异贡献值为52.39%. ANOSIM检验结果表明, 4种不同种植模式下的AMF群落完全分开, 说明各处理之间差异显著(R=0.688 3, P=0.001).

|
图 4 轮作休耕模式下基于Bray-Curtis距离AMF群落PCoA图 Fig. 4 PCoA plot based on Bray-Curtis similarities of AMF community under crop rotation and fallow pattern |
为分析影响轮作休耕下AMF多样性及群落组成产生差异的关键性因子, 对其进行RDA和Spearman分析.RDA1和RDA2分别解释了69.89%和4.85%的变异量, 能够在累计变量74.74%上解释AMF多样性和丰富度指数的变化[图 5(a)].其中Chao1、ACE和Shannon指数均与AN和SOM呈正相关, 与AP、pH和AK呈负相关.AN对AMF多样性和丰富度有显著影响(P<0.05), 说明AN是轮作休耕模式下影响AMF丰富度和多样性的主要因子.Spearman相关性分析表明[图 5(b)], 球囊霉属与AN呈显著正相关(P<0.05); 大孢子科未分类属(Unclassified_ f_ _Gigasporaceae)和盾巨孢囊霉属(Scutellospora)均与AN和SOM呈显著负相关(P<0.05), 与AK呈显著正相关(P<0.05).

|
(a)中红色箭头表示土壤养分, 蓝色箭头表示AMF多样性及丰富度指数; (b)中红色表示正相关, 蓝色表示负相关; *表示P<0.05 图 5 轮作休耕模式下AMF多样性指数及属水平的相对丰度与土壤养分的关系 Fig. 5 Relationship between AMF diversity, abundance of genus levels and soil nutrients under crop rotation and fallow pattern |
冗余分析表明[图 6(a)], >2 mm及2~1 mm团聚体含量、R0.25和MWD与Shannon、ACE和Chao1之间均呈正相关关系, 其中>2 mm团聚体含量与Shannon指数相关性显著(P<0.05). Scutellospora、Unclassified_ f_ _Gigasporaceae和Paraglomus与>2 mm及2~1 mm团聚体含量、R0.25和MWD呈正相关, 其中Scutellospora与2~1 mm团聚体含量相关性显著(P<0.05); Gigaspora与>2 mm团聚体含量、R0.25和MWD正相关, 而Glomus与大团聚体含量和稳定性负相关[图 6(b)].综上, AMF多样性(Shannon指数)和Scutellospora是轮作休耕模式下促进土壤团聚体形成和稳定的主要影响因子.

|
红色箭头分别表示多样性指数和前6个属, 蓝色箭头均表示团聚体粒径及稳定性; R0.25表示>0.25 mm水稳性团聚体含量, MWD表示平均重量直径 图 6 轮作休耕模式下AMF多样性及群落与土壤团聚体的关系 Fig. 6 Relationship between AMF diversity, communities, and soil aggregate under crop rotation and fallow pattern |
土壤团聚体是土壤的重要结构单元, 其组成和稳定性通常用作土壤肥力的重要指标[7].当团聚体稳定性高时可以大幅提高土壤的水渗透性及抗侵蚀能力[22].本研究结果发现, 与冬闲-玉米(F-C)处理相比, 周年休耕(F-F)、苕子轮作玉米(V-C)和豌豆轮作玉米(P-C)处理均能显著提高土壤大团聚体(>0.25 mm)含量和团聚体稳定性(MWD), 这与夏梓泰等[8]对轮作休耕模式下土壤团聚体进行研究所得的结果相似.Seidel等[23]在季节性休耕地种植绿肥, 发现由于绿肥根系在土壤中持续活动及其被还田后的腐殖化作用, 可改良土壤团粒结构、增加大团聚体数量并提高土壤团聚体稳定性, 这与本研究的结果一致.产生本研究结果的原因可能是F-C处理冬季地表覆盖度低, 且耕作时加剧了对土壤的扰动, 使土壤团粒结构被破坏, 进而降低了土壤团聚体稳定性.而F-F、V-C和P-C处理冬季地表覆盖度高且对其扰动较少, 因此有利于减少土壤养分的流失和恢复土壤团聚体的稳定性[24], 这也与何宇等[25]通过7种不同土地利用方式对土壤团聚体进行研究所得到的结果相似.因此, 周年休耕及豆科绿肥轮作均能达到使土地休养生息、恢复地力和保持水土的效果.
3.2 轮作休耕模式对土壤养分的影响已有研究表明, 合理的绿肥轮作和休耕可以增加土壤的通透性、保水保肥、提高酸碱缓冲能力和养分含量, 从而提高作物产量[26, 27].本研究结果也表明, F-F、V-C和P-C较F-C处理能够显著提高SOM和AN的含量.其原因在于F-F处理中枯枝落叶腐解及轮作小区中绿肥被翻压后经过腐解能增加土壤中有机质和氮素的含量, 从而改善土壤环境[28].本研究中土壤pH大小表现为:F-C>V-C>P-C>F-F, 这与韩梅等[29]和钟菊新等[30]通过不同绿肥配施化肥对pH进行研究得到的结果相似.这是由于F-F处理的枯枝落叶及轮作处理的绿肥经过腐解产生有机酸所致.本研究还得出, 绿肥处理的AP和AK含量低于F-C, 这与钟菊新等[30]研究的结果一致.可能原因是绿肥增加的土壤碳氮养分激发了微生物生长活性, 使微生物对磷的需求增加, 从而导致AP优先被土壤微生物所固持[31].又由于F-F处理的植物及绿肥轮作处理的苕子和豌豆生长需要大量钾素[32], 致使土壤AK含量降低.赵秋等[33]通过对天津地区不同绿肥培肥土壤效果进行研究后发现, 冬季绿肥处理较冬闲能够显著增加AP和AK含量, 这与本研究结果相悖, 其原因可能是气候条件和土壤类型等不同.目前研究报道的结果不尽相同, 但总体上豆科绿肥与玉米轮作能够改善土壤理化性质, 提高作物产量, 对于坡耕地生态环境保护和粮食增产具有重要意义, 是一种值得推广的种植模式.
3.3 轮作休耕模式对AMF多样性及群落组成的影响AMF作为农田生态系统中的一种重要的土壤微生物, 有助于土壤健康和农业可持续发展, 其多样性及群落结构的变化受耕作措施等多种因素的影响[11, 34].在本研究中, 不同处理对AMF群落丰富度和多样性指数均产生了不同程度的影响, 这可能与绿肥的种类、休耕年限、气候条件和土壤类型等因素有关.其中F-F处理提高了AMF群落丰富度和多样性, 其原因可能是减少对土壤的扰动可以避免AMF菌丝网络遭到破坏, 从而有利于AMF生长和繁殖, 进而提高AMF群落丰富度和多样性[35].Säle等[36]将常规耕作与免耕系统进行比较, 也发现土壤频繁耕作会对AMF丰富度产生不利影响, 其原因是耕作稀释了AMF繁殖体.因此休耕可以使土地修养生息、保持质量和恢复地力, 从而有利于菌根的生长和繁殖.
本研究利用高通量测序, 共发现AMF群落1门、1纲、5目、7科、11属和27种, 基于属水平的物种丰度排在前6位的AMF属有球囊霉属、无梗囊霉属、类球囊霉属、巨孢囊霉属、大孢子科未分类属和盾巨孢囊霉属, 其中球囊霉属为优势属.已有学者对农田、森林、草地、湿地及不同免耕和有机肥施条件下的AMF多样性及群落结构进行研究, 发现球囊霉属为优势属[37], 这与本研究的结果一致.这可能是由于球囊霉属具有较高的产孢率和特定的繁殖方式, 对农业管理活动扰动的忍耐能力较强, 且能够适应不同的环境[38, 39].Higo等[40]通过对意大利黑麦草等覆盖作物或休耕地与玉米进行轮作, 表明冬季覆盖作物可影响后茬作物的AMF群落组成.本研究中苕子和豌豆作为冬季覆盖绿肥也能改变后茬玉米土壤AMF群落组成, 这与Higo等[40]研究的结果一致.
3.4 轮作休耕模式下AMF多样性及属水平相对丰度与土壤养分间的关系AMF群落结构受到植物种类、覆盖作物、农艺措施和养分等多种因素的影响[41], 其中养分主要包括气候条件和土壤理化性质[42].本研究得出, 球囊霉属与AN显著正相关, 这与Zhang等[43]研究玉米/大豆间作系统中AMF群落组成与养分的相关关系结果一致, 可能是因为土壤高氮含量会促进AMF孢子的形成[44]; AN与AMF多样性显著正相关, 这与Xiang等[45]研究不同土地利用对农牧交错带AMF群落的影响结果相似, 但与Jansa等[46]研究的结果相反, 说明氮含量对AMF群落的影响存在一定的阈值[47].此外, 本研究中AK与AMF大孢子科未分类属和盾巨孢囊霉属存在显著正相关关系, 这与大多数学者研究认为AK与AMF群落结构无相关关系的结果不一致.但也有研究表明, AK对AMF群落结构有显著影响[44], 这与本研究结果一致.Hammer等[48]研究发现K的有效性能够有选择地刺激AMF生长, 因为它补偿了钠诱导的离子失衡.综上, 在试验条件下AN是AMF多样性的主要影响因子, AN、SOM和AK是AMF群落结构的主要影响因子.目前, 关于土壤养分对土壤AMF群落的影响研究结果不尽相同[49], 因此不同轮作休耕模式下AMF结构特征对土壤养分的影响有待深入研究.
3.5 轮作休耕模式下AMF多样性及群落与土壤团聚体之间的关系AMF对于土壤生态系统功能至关重要, 在土壤团聚体形成、稳定以及植物和土壤系统之间的联系中发挥着关键作用[50].已有研究表明, AMF通过分泌多糖、酚酸及GRSP与土壤颗粒和微团聚体(<0.25 mm)结合, 从而促进土壤大团聚体的形成和稳定[51].本研究结果表明, >2 mm团聚体含量与Shannon指数呈显著正相关关系(P<0.05), 这与Qin等[52]研究得出的大团聚体(0.25~2 mm)所占质量分数与土壤AM真菌生物量呈正相关的结果相似. Scutellospora与2~1 mm团聚体含量、>2 mm团聚体含量、R0.25及MWD均呈正相关关系, 其中与2~1 mm团聚体含量的相关性达显著水平(P<0.05).Hart等[53]研究表明, Gigasporaceae科真菌较非Gigasporaceae科真菌能够产生更长的菌丝和更密集的菌丝簇, 其土壤黏结效果较好.本研究中的Scutellospora是Gigasporaceae科的一个属, 这与前人得出的结果相吻合.但与Lu等[54]研究得出的隔球囊霉属和球囊霉属与大团聚体所占质量分数呈正相关的结果不一致, 可能是所研究的土壤性质和气候条件等的差异所致.Wilson等[50]研究表明土壤团聚体形成与AMF的群落和丰度密切相关, 这与本研究的结果一致, 周年休耕和冬绿肥覆盖有利于后茬作物AMF定殖[55], 从而促进团聚体的形成和稳定.Lehmann等[56]研究表明腐生真菌降解有机物质(植物叶子或根), 并为土壤团聚体的形成提供必要的原材料.Cao等[57]研究也发现, AMF可以通过促进土壤腐生真菌的生长, 间接促进有机物质分解和总体胞外酶丰度提高, 这可能是AMF促进土壤团聚体形成的一种途径.在本研究中, 周年休耕和绿肥轮作小区地表覆盖度高且枯枝落叶繁多, 这可能促进了腐生真菌的生长和繁殖, 加之对AMF菌丝网络破坏较少, 最终通过AMF和腐生真菌相互作用促进了土壤团聚体的形成与稳定.
4 结论(1) 周年休耕和绿肥轮作玉米处理均能够显著提高土壤大团聚体含量及团聚体稳定性, 且周年休耕处理的效果最佳, 苕子轮作玉米处理次之.
(2) 周年休耕处理的AMF多样性和丰富度显著高于冬闲-玉米处理; 球囊霉属为各处理的AMF优势属, 盾巨孢囊霉属在各处理之间差异显著.
(3) 周年休耕和绿肥轮作玉米能够显著提高SOM和AN含量.其中SOM、AN及AK对AMF群落组成有显著影响, AN是轮作休耕模式下AMF丰富度和多样性的主要影响因子, 与球囊霉属存在显著正相关关系; AMF群落对轮作休耕模式的响应受土壤养分、绿肥种类及休耕年限等因素的综合影响.
(4) AMF多样性(Shannon指数)和盾巨孢囊霉属分别与>2 mm和2~1 mm团聚体含量显著正相关, AMF多样性(Shannon指数)和盾巨孢囊霉属对轮作休耕模式下土壤团聚体形成和稳定具有重要作用.
| [1] |
李硕, 刘磊. AnnAGNPS模型在潋水河流域产水、产沙的模拟评价[J]. 环境科学, 2010, 31(1): 49-57. Li S, Liu L. Evaluation of AnnAGNPS model for simulation water and sediment yield in the lianshui river watershed[J]. Environmental Science, 2010, 31(1): 49-57. |
| [2] |
赵桂茹, 程易, 杨友琼, 等. 玉米‖马铃薯接种AMF对坡耕地水土及氮素流失的影响[J]. 云南农业大学学报(自然科学), 2022, 37(3): 523-529. Zhao G R, Cheng Y, Yang Y Q, et al. Effects of maize intercropping potato inoculation with arbuscular mycorrhizal fungi on soil erosion and nitrogen losses in slope farmland[J]. Journal of Yunnan Agricultural University (Natural Science), 2022, 37(3): 523-529. |
| [3] | Barthès B, Roose E. Aggregate stability as an indicator of soil susceptibility to runoff and erosion; validation at several levels[J]. CATENA, 2002, 47(2): 133-149. DOI:10.1016/S0341-8162(01)00180-1 |
| [4] |
罗晓虹, 王子芳, 陆畅, 等. 土地利用方式对土壤团聚体稳定性和有机碳含量的影响[J]. 环境科学, 2019, 40(8): 3816-3824. Luo X H, Wang Z F, Lu C, et al. Effects of land use type on the content and stability of organic carbon in soil aggregates[J]. Environmental Science, 2019, 40(8): 3816-3824. |
| [5] |
刘亚龙, 王萍, 汪景宽. 土壤团聚体的形成和稳定机制: 研究进展与展望[J]. 土壤学报, 2023, 60(3): 627-643. Liu Y L, Wang P, Wang J K. Formation and stability mechanism of soil aggregates: progress and prospect[J]. Acta Pedologica Sinica, 2023, 60(3): 627-643. |
| [6] |
李丽, 张安录. 轮作休耕及其补偿的法律意蕴、法理证成及入法进路[J]. 中国土地科学, 2021, 35(11): 27-35. Li L, Zhang A L. Legal implication, legal evidence and legalization path of crop rotation and fallow with compensation[J]. China Land Science, 2021, 35(11): 27-35. |
| [7] | Tian X L, Wang C B, Bao X G, et al. Crop diversity facilitates soil aggregation in relation to soil microbial community composition driven by intercropping[J]. Plant and Soil, 2019, 436(1-2): 173-192. DOI:10.1007/s11104-018-03924-8 |
| [8] |
夏梓泰, 程伟威, 李永梅, 等. 轮作休耕模式对土壤团聚体及有机碳含量的影响[J]. 水土保持研究, 2022, 29(3): 31-37. Xia Z T, Cheng W W, Li Y M, et al. Effects of crop rotation and fallow patterns on soil aggregates and organic carbon content[J]. Research of Soil and Water Conservation, 2022, 29(3): 31-37. |
| [9] | Tiemann L K, Grandy A S, Atkinson E E, et al. Crop rotational diversity enhances belowground communities and functions in an agroecosystem[J]. Ecology Letters, 2015, 18(8): 761-771. DOI:10.1111/ele.12453 |
| [10] | Marinho F, Oehl F, Da Silva I R D, et al. High diversity of arbuscular mycorrhizal fungi in natural and anthropized sites of a Brazilian tropical dry forest (Caatinga)[J]. Fungal Ecology, 2019, 40: 82-91. DOI:10.1016/j.funeco.2018.11.014 |
| [11] | Bender S F, Wagg C, Van Der Heijden M G A. An underground revolution: biodiversity and soil ecological engineering for agricultural sustainability[J]. Trends in Ecology & Evolution, 2016, 31(6): 440-452. |
| [12] | Mardhiah U, Caruso T, Gurnell A, et al. Arbuscular mycorrhizal fungal hyphae reduce soil erosion by surface water flow in a greenhouse experiment[J]. Applied Soil Ecology, 2016, 99: 137-140. DOI:10.1016/j.apsoil.2015.11.027 |
| [13] | Xiao L, Zhang Y, Li P, et al. Effects of freeze-thaw cycles on aggregate-associated organic carbon and glomalin-related soil protein in natural-succession grassland and Chinese pine forest on the Loess Plateau[J]. Geoderma, 2019, 334: 1-8. DOI:10.1016/j.geoderma.2018.07.043 |
| [14] | Higo M, Tatewaki Y, Gunji K, et al. Cover cropping can be a stronger determinant than host crop identity for arbuscular mycorrhizal fungal communities colonizing maize and soybean[J]. PeerJ, 2019, 7. DOI:10.7717/peerj.6403 |
| [15] | Rillig M C, Mummey D L. Mycorrhizas and soil structure[J]. New Phytologist, 2006, 171(1): 41-53. DOI:10.1111/j.1469-8137.2006.01750.x |
| [16] | Zhang Z F, Mallik A, Zhang J C, et al. Effects of arbuscular mycorrhizal fungi on inoculated seedling growth and rhizosphere soil aggregates[J]. Soil and Tillage Research, 2019, 194. DOI:10.1016/j.still.2019.104340 |
| [17] | Zhang S, Meng L B, Hou J, et al. Maize/soybean intercropping improves stability of soil aggregates driven by arbuscular mycorrhizal fungi in a black soil of northeast China[J]. Plant and Soil, 2022. DOI:10.1007/s11104-022-05616-w |
| [18] | Wright S F, Anderson R L. Aggregate stability and glomalin in alternative crop rotations for the central Great Plains[J]. Biology and Fertility of Soils, 2000, 31(3-4): 249-253. DOI:10.1007/s003740050653 |
| [19] | 鲍士旦. 土壤农化分析[M]. (第三版). 北京: 中国农业出版社, 2018. |
| [20] | Elliott E T. Aggregate structure and carbon, nitrogen, and phosphorus in native and cultivated soils[J]. Soil Science Society of America Journal, 1986, 50(3): 627-633. DOI:10.2136/sssaj1986.03615995005000030017x |
| [21] |
邓华, 高明, 龙翼, 等. 生物炭和秸秆还田对紫色土旱坡地土壤团聚体与有机碳的影响[J]. 环境科学, 2021, 42(11): 5481-5490. Deng H, Gao M, Long Y, et al. Effects of biochar and straw return on soil aggregate and organic carbon on purple soil dry slope land[J]. Environmental Science, 2021, 42(11): 5481-5490. |
| [22] |
王兴, 钟泽坤, 张欣怡, 等. 长期撂荒恢复土壤团聚体组成与有机碳分布关系[J]. 环境科学, 2020, 41(5): 2416-2424. Wang X, Zhong Z K, Zhang X Y, et al. Relationship between the composition of soil aggregates and the distribution of organic carbon under long-term abandoned restoration[J]. Environmental Science, 2020, 41(5): 2416-2424. |
| [23] | Seidel E P, Caetano J H S, Karpinski A S, et al. Residual dry matter, weeds and soil aggregates after winter cover crop[J]. Journal of Experimental Agriculture International, 2019, 32(2). DOI:10.9734/jeai/2019/v32i230100 |
| [24] |
夏梓泰, 赵吉霞, 李永梅, 等. 周年轮作休耕模式对土壤球囊霉素和团聚体稳定性的影响[J]. 农业环境科学学报, 2022, 41(1): 99-106. Xia Z T, Zhao J X, Li Y M, et al. Effect of annual rotation and fallow pattern on the soil glomalin and aggregate stability[J]. Journal of Agro-Environment Science, 2022, 41(1): 99-106. |
| [25] |
何宇, 盛茂银, 王轲, 等. 土地利用变化对西南喀斯特土壤团聚体组成、稳定性以及C、N、P化学计量特征的影响[J]. 环境科学, 2022, 43(7): 3752-3762. He Y, Sheng M Y, Wang K, et al. Effects of land use change on constitution, stability, and C, N, P stoichiometric characteristics of soil aggregates in Southwest China karst[J]. Environmental Science, 2022, 43(7): 3752-3762. |
| [26] |
王天乐, 王晓娟, 刘恩科, 等. 甘蓝、菜豆和玉米不同轮作组合对淡褐土细菌群落和作物产量的影响[J]. 农业环境科学学报, 2022, 41(2): 375-386. Wang T L, Wang X J, Liu E K, et al. Effects of different crop rotation combinations of cabbage, kidney bean, and maize on bacterial community and crop yield in light brown soil[J]. Journal of Agro-Environment Science, 2022, 41(2): 375-386. |
| [27] | Zhang C Z, Xue W F, Xue J R, et al. Leveraging functional traits of cover crops to coordinate crop productivity and soil health[J]. Journal of Applied Ecology, 2022, 59(10): 2627-2641. |
| [28] |
刘佳, 张杰, 秦文婧, 等. 红壤旱地毛叶苕子不同翻压量下腐解及养分释放特征[J]. 草业学报, 2016, 25(10): 66-76. Liu J, Zhang J, Qin W J, et al. Decomposition and nutrient release characteristics of different Vicia villosa green manure applications in red soil uplands of South China[J]. Acta Prataculturae Sinica, 2016, 25(10): 66-76. |
| [29] |
韩梅, 刘蕊, 李正鹏, 等. 种植绿肥对作物产量和细菌群落稳定性的影响[J]. 中国环境科学, 2021, 41(11): 5382-5390. Han M, Liu R, Li Z P, et al. Green manuring increased crop yields and the stability of bacterial community[J]. China Environmental Science, 2021, 41(11): 5382-5390. |
| [30] |
钟菊新, 唐红琴, 李忠义, 等. 绿肥配施化肥对岩溶区水稻土壤细菌群落结构的影响[J]. 植物营养与肥料学报, 2021, 27(10): 1746-1756. Zhong J X, Tang H Q, Li Z Y, et al. Effects of combining green manure with chemical fertilizer on the bacterial community structure in karst paddy soil[J]. Journal of Plant Nutrition and Fertilizers, 2021, 27(10): 1746-1756. |
| [31] | Pistocchi C, Mészáros é, Tamburini F, et al. Biological processes dominate phosphorus dynamics under low phosphorus availability in organic horizons of temperate forest soils[J]. Soil Biology and Biochemistry, 2018, 126: 64-75. |
| [32] | Gao S J, Cao W D, Gao J S, et al. Effects of long-term application of different green manures on ferric iron reduction in a red paddy soil in Southern China[J]. Journal of Integrative Agriculture, 2017, 16(4): 959-966. |
| [33] |
赵秋, 张新建, 宁晓光, 等. 天津地区不同冬绿肥培肥土壤的效果[J]. 植物营养与肥料学报, 2022, 28(7): 1228-1237. Zhao Q, Zhang X J, Ning X G, et al. Effects of different winter green manure on soil fertility in Tianjin[J]. Journal of Plant Nutrition and Fertilizers, 2022, 28(7): 1228-1237. |
| [34] | Fall A F, Nakabonge G, Ssekandi J, et al. Roles of arbuscular mycorrhizal fungi on soil fertility: contribution in the improvement of physical, chemical, and biological properties of the soil[J]. Frontiers in Fungal Biology, 2022, 3. DOI:10.3389/ffunb.2022.723892 |
| [35] | Amalia A V, Dewi N R, Heriyanti A P, et al. A review on arbuscular mycorrhizal fungal communities in response to disturbance[J]. Journal of Physics: Conference Series, 2021, 1968(1). DOI:10.1088/1742-6596/1968/1/012001 |
| [36] | Säle V, Aguilera P, Laczko E, et al. Impact of conservation tillage and organic farming on the diversity of arbuscular mycorrhizal fungi[J]. Soil Biology and Biochemistry, 2015, 84: 38-52. |
| [37] |
刘文娟, 马琨, 王兵兵, 等. 冬小麦免耕覆盖与有机肥施用对土壤丛枝菌根真菌群落多样性的影响[J]. 生态学杂志, 2022, 41(6): 1101-1108. Liu W J, Ma K, Wang B B, et al. Effects of no-tillage, mulching, and organic fertilizer application on the diversity of arbuscular mycorrhizal fungi in winter wheat field[J]. Chinese Journal of Ecology, 2022, 41(6): 1101-1108. |
| [38] |
马琨, 宋丽丽, 王明国, 等. 玉米秸秆还田对土壤丛枝菌根真菌群落的影响[J]. 应用生态学报, 2019, 30(8): 2746-2756. Ma K, Song L L, Wang M G, et al. Effects of maize straw returning on arbuscular mycorrhizal fungal community structure in soil[J]. Chinese Journal of Applied Ecology, 2019, 30(8): 2746-2756. |
| [39] | Xue J F, Pu C, Liu S L, et al. Effects of tillage systems on soil organic carbon and total nitrogen in a double paddy cropping system in Southern China[J]. Soil and Tillage Research, 2015, 153: 161-168. |
| [40] | Higo M, Takahashi Y, Gunji K, et al. How are arbuscular mycorrhizal associations related to maize growth performance during short-term cover crop rotation?[J]. Journal of the Science of Food and Agriculture, 2018, 98(4): 1388-1396. |
| [41] | Zeng H L, Yu L L, Liu P, et al. Nitrogen fertilization has a stronger influence than cropping pattern on AMF community in maize/soybean strip intercropping systems[J]. Applied Soil Ecology, 2021, 167. DOI:10.1016/j.apsoil.2021.104034 |
| [42] | Xiang D, Veresoglou S D, Rillig M C, et al. Relative importance of individual climatic drivers shaping arbuscular mycorrhizal fungal communities[J]. Microbial Ecology, 2016, 72(2): 418-427. |
| [43] | Zhang R Z, Mu Y, Li X R, et al. Response of the arbuscular mycorrhizal fungi diversity and community in maize and soybean rhizosphere soil and roots to intercropping systems with different nitrogen application rates[J]. Science of The Total Environment, 2020, 740. DOI:10.1016/j.scitotenv.2020.139810 |
| [44] | Qin H, Lu K P, Strong P J, et al. Long-term fertilizer application effects on the soil, root arbuscular mycorrhizal fungi and community composition in rotation agriculture[J]. Applied Soil Ecology, 2015, 89: 35-43. |
| [45] | Xiang D, Verbruggen E, Hu Y J, et al. Land use influences arbuscular mycorrhizal fungal communities in the farming-pastoral ecotone of northern China[J]. New Phytologist, 2014, 204(4): 968-978. |
| [46] | Jansa J, Oberholzer H R, Egli S. Environmental determinants of the arbuscular mycorrhizal fungal infectivity of Swiss agricultural soils[J]. European Journal of Soil Biology, 2009, 45(5-6): 400-408. |
| [47] |
杨蕊毓, 邓錡璋, 田丽平, 等. 不同生境下川麦冬根围土壤丛枝菌根真菌多样性[J]. 西北植物学报, 2022, 42(1): 145-153. Yang R Y, Deng Q Z, Tian L P, et al. Diversity of arbuscular mycorrhizal fungi in the rhizosphere of Ophiopogon japonicus in different habitats[J]. Acta Botanica Boreali-Occidentalia Sinica, 2022, 42(1): 145-153. |
| [48] | Hammer E C, Nasr H, Wallander H. Effects of different organic materials and mineral nutrients on arbuscular mycorrhizal fungal growth in a Mediterranean saline dryland[J]. Soil Biology and Biochemistry, 2011, 43(11): 2332-2337. |
| [49] |
南镇武, 刘柱, 代红翠, 等. 不同轮作休耕下潮土细菌群落结构特征[J]. 环境科学, 2021, 42(10): 4977-4987. Nan Z W, Liu Z, Dai H C, et al. Characteristics of bacterial community structure in fluvo-aquic soil under different rotation fallow[J]. Environmental Science, 2021, 42(10): 4977-4987. |
| [50] | Wilson G W T, Rice C W, Rillig M C, et al. Soil aggregation and carbon sequestration are tightly correlated with the abundance of arbuscular mycorrhizal fungi: results from long-term field experiments[J]. Ecology Letters, 2009, 12(5): 452-461. |
| [51] | Gao Y Z, Zong J, Que H, et al. Inoculation with arbuscular mycorrhizal fungi increases glomalin-related soil protein content and PAH removal in soils planted with Medicago sativa L.[J]. Soil Biology and Biochemistry, 2017, 115: 148-151. |
| [52] | Qin H, Niu L M, Wu Q F, et al. Bamboo forest expansion increases soil organic carbon through its effect on soil arbuscular mycorrhizal fungal community and abundance[J]. Plant and Soil, 2017, 420(1-2): 407-421. |
| [53] | Hart M M, Reader R J. Taxonomic basis for variation in the colonization strategy of arbuscular mycorrhizal fungi[J]. New Phytologist, 2002, 153(2): 335-344. |
| [54] | Lu X L, Lu X N, Liao Y C. Effect of tillage treatment on the diversity of soil arbuscular mycorrhizal fungal and soil aggregate-associated carbon content[J]. Frontiers in Microbiology, 2018, 9. DOI:10.3389/fmicb.2018.02986 |
| [55] | Morimoto S, Uchida T, Matsunami H, et al. Effect of winter wheat cover cropping with no-till cultivation on the community structure of arbuscular mycorrhizal fungi colonizing the subsequent soybean[J]. Soil Science and Plant Nutrition, 2018, 64(5): 545-553. |
| [56] | Lehmann A, Rillig M C. Understanding mechanisms of soil biota involvement in soil aggregation: A way forward with saprobic fungi?[J]. Soil Biology and Biochemistry, 2015, 88: 298-302. |
| [57] | Cao T T, Fang Y, Chen Y R, et al. Synergy of saprotrophs with mycorrhiza for litter decomposition and hotspot formation depends on nutrient availability in the rhizosphere[J]. Geoderma, 2022, 410. DOI:10.1016/j.geoderma.2021.115662 |