 2025, Vol. 46
2025, Vol. 46

2. 河北科技大学食品与生物学院, 石家庄 050018
 , WANG Mei-qi1 , SONG Yao2 , NIU Hong-jin1 , ZHAO Jing-wei1 , HOU Rui-nan2 , CHEN Xiao-bo2
, WANG Mei-qi1 , SONG Yao2 , NIU Hong-jin1 , ZHAO Jing-wei1 , HOU Rui-nan2 , CHEN Xiao-bo2 , HUANG Ya-li1
, HUANG Ya-li1
2. College of Food Science and Biology, Hebei University of Science and Technology, Shijiazhuang 050018, China
我国是农业大国, 年均秸秆产量为8.65×108 t[1]. 秸秆作为一种数量巨大的可再生资源, 其资源化利用对保障农业可持续发展、实现我国“双碳”目标具有重要的意义[2]. 目前, 我国秸秆资源化利用量达7.3×108 t, 其中秸秆直接还田方式占61.3%[3]. 随着秸秆还田实践时间的延长, 秸秆还田效应评估是未来农田健康管理的基础.
早期研究者对秸秆还田效应的评估以理化指标为主, 小麦-玉米双季秸秆还田显著降低了土壤容重、增加了土壤孔隙度[4];增加还田深度后秸秆还田能够改变20~30 cm土层的土壤容重和孔隙度[5], 有效地改善了麦玉轮作土壤的透气、持水和导水性能[6]. 由于秸秆富含有机质, 50%和100%小麦-玉米双季秸秆还田量较对照土壤中的有机碳含量分别增加25.2%和14.2%[7];在秸秆还田配施化肥的情况下还能够显著增加土壤碱解氮的含量[8], 有效地增加了土壤的有机养分和速效养分, 为农作物生长提供了良好的理化条件.
随着研究的深入, 秸秆还田所引起的土壤生物活性指标改变越来越受到广泛的关注[9 ~ 12]. Cui等[13]研究发现小麦和玉米秸秆还田使麦玉轮作土壤的脲酶和碱性磷酸酶活性较秸秆未还田对照分别增加83.45%和53.62%. 土壤微生物作为农田生态系统中最为活跃的组分, 是反映秸秆还田土壤效应的更为直观和敏感的土壤生物活性指标[14, 15]. 与仅施化肥处理相比, 玉米秸秆还田配施化肥使向日葵土壤细菌和真菌数量分别增加了38.69%和28.04%[16]. 还有研究表明, 秸秆还田不仅能够增加土壤微生物数量[17], 还显著影响麦玉轮作土壤细菌群落结构, 从而更有效地促进秸秆分解和转化[18]. 在水稻田中的研究则发现, 水稻秸秆还田提高了参与碳循环的微生物相对丰度, 使土壤微生物群落更加趋于稳定和多样化[19].
总体来说, 秸秆还田能够明显改善当季土壤理化性质、优化土壤微生物群落[20, 21]. 多年秸秆还田对土壤性质的改变会逐年累积, 实现土壤性质的稳定提升, 存在较为明显的土壤肥效残留效应[22]. 然而, 目前对秸秆还田的研究主要集中于对比分析秸秆还田和未还田处理间的土壤效应, 对长期秸秆还田所引起的土壤效应对下茬还田秸秆的降解和微生物菌群结构的影响还未见报道. 因此, 本文以小麦-玉米轮作模式下秸秆长期还田定位试验(15 a)耕层土壤为研究对象, 采用模拟试验, 向长期秸秆还田和秸秆未还田土壤中添加秸秆后, 对比分析两种土壤中秸秆降解情况、土壤可培养微生物数量和土壤微生物群落的差异, 明确长期秸秆还田的土壤残留效应, 以期为区域秸秆安全和高效还田提供理论基础.
1 材料与方法 1.1 试验材料本试验土壤采自河北省农林科学院石家庄鹿泉区大河镇试验站(38°12′56″N, 114°38′26″E)的秸秆长期还田定位试验田. 定位试验自2007年开始, 试验田小区面积60 m2(6 m×10 m), 在小麦-玉米一年两熟轮作模式下设置秸秆还田处理(SS)和对照(NS), 其中SS为连续15 a收获时将小麦秸秆粉碎后覆盖还田、玉米秸秆粉碎后旋耕还田, NS为小麦、玉米地上部秸秆全部移出. 两处理其他管理均相同. 2021年玉米收获后, 采用“五点取样法”分别采集两处理0~20 cm土层的土壤, 作为本研究模拟试验土壤. SS土壤基本理化性质为pH 8.32、ω(有机质)24.02 g·kg-1、ω(全氮)1.44 g·kg-1、ω(有效磷)22.10 mg·kg-1和ω(速效钾)143.60 mg·kg-1. NS土壤基本理化性质为pH 8.33、ω(有机质)23.76 g·kg-1、ω(全氮)1.38 g·kg-1、ω(有效磷)21.60 mg·kg-1和ω(速效钾)150.60 mg·kg-1.
1.2 试验方法采用模拟试验评价长期秸秆还田对还田秸秆降解和土壤微生物菌群的残留效应. 将SS和NS土壤分别混匀, 分装于边长8 cm、高8 cm的花盆中, 每个花盆装土500 g, 并在每盆土壤中部埋入一个盛有5.00 g(烘干重)玉米秸秆的尼龙布袋, 每个处理9盆, 3次重复. 玉米秸秆从河北省石家庄田间采集, 剪碎至长度≤1 cm后使用. 试验过程采用称重法浇水以保持土壤湿润, 分别于处理后的0、15、30、45和60 d取出秸秆沙袋, 每个重复随机选取1盆, 测定秸秆降解相关指标. 同样, 为更好地反映秸秆还田效应, 将粉碎的玉米秸秆分别与SS和NS土壤样品混合后分装于花盆内, 于0、3、7、14、28和56 d取出土壤样品, 用于可培养微生物数量的测定, 其中秸秆还田第7 d的土壤样品用于土壤DNA提取及高通量测序.
1.3 测定指标及方法 1.3.1 秸秆降解率测定取出沙袋并将秸秆上黏附的土壤冲洗干净, 于80 ℃烘干至恒重并记录烘干重(Wt). 秸秆降解率的计算公式如下:
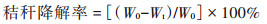
|
式中, W0为初始秸秆干质量(为5 g), Wt为降解后秸秆干质量.
秸秆降解情况的显微观察:分别取适量两处理的干燥玉米秸秆样品, 将其制成2 mm×2 mm小块通过离子溅射仪金靶溅射镀金[23], 再将镀金后的秸秆样品放置于扫描电镜下, 加速电压为3.0 kV, 放大倍数为1 500~5 000倍, 拍摄记录玉米秸秆的表面结构.
1.3.2 土壤可培养微生物数量测定采用稀释涂布平板法进行土壤中细菌、真菌、氨化菌、纤维降解菌和亚硝化细菌的计数[24], 其中细菌采用牛肉膏蛋白胨琼脂培养基, 28 ℃培养2 d;真菌采用马铃薯葡萄糖琼脂培养基, 28 ℃培养3 d;氨化菌采用牛肉膏蛋白胨培养基, 28 ℃培养3 d;纤维素降解菌采用赫奇逊培养基, 28 ℃培养14 d;亚硝化细菌采用改良斯蒂芬逊培养基, 28 ℃培养14 d.
1.3.3 DNA提取和高通量测序采用天根生化科技(北京)有限公司土壤DNA提取试剂盒(DP812, TGuide S96磁珠法)进行DNA的提取, 以质量合格的DNA为模板进行PCR扩增, 细菌16S rRNA的V3-V4高变区扩增上游引物序列为338F:5′-ACTCCTACGGGAGGCAGCA-3', 下游引物序列为806R:5′-GGACTACHVGGGTWTCTAAT-3′[25];真菌ITS基因的ITS1区域扩增上游引物序列为1737F:5′-CTTGGTCATTTAGAGGAAGTAA-3′和下游引物序列为2043R:5′-GCTGCGTTCTTCATCGA TGC-3′[26]. PCR扩增体系:95 ℃预变性3 min;95 ℃ 30 s, 50 ℃ 30 s, 72 ℃ 1 min, 共35个循环;72 ℃延伸10 min, 4 ℃结束反应. 对PCR反应产物进行检测、回收和纯化, 检测合格的样品送至北京百迈客生物科技有限公司采用Illumina NovaSeq 6000测序平台进行高通量测序.
1.3.4 数据处理高通量测序得到原始图像数据文件, 经碱基识别分析转化为原始序列后, 使用QIIME2 v2020.6中的DADA2通道去除引物、质量滤波、去噪、剪接和嵌合. 利用UCHIIME(version 8.1)软件在97%相似度下进行可操作分类单元(ASVs)聚类及各分类水平注释. 使用QIIME2软件和R语言进行α多样性分析. 使用R语言进行β多样性分析, 对获得的Bray-curtis距离矩阵(默认)进行非度量多维尺度(NMDS)分析, 并通过二维排序图展示微生物群落的组成差异. 使用QIIME2软件进行物种分类学组成分析. 利用Gephi软件绘制物种相关性网络图. 使用R语言对微生物群落组成与秸秆降解率(15 d)的关系进行冗余分析(RDA)并作图.
使用Excel 2021软件进行数据整理. 使用IBM SPSS 26.0统计分析软件对整理后的试验数据进行独立样本T检验, 使用邓肯法(Duncan)(P < 0.05和P < 0.01)进行显著性检验. 使用Origin 2021作图.
2 结果与分析 2.1 长期秸秆还田土壤残留效应对秸秆降解的影响由图 1结果可知, 两个处理的秸秆降解率均随还田时间的延长而逐渐增加, 其SS处理中秸秆降解率在不同时间段均高于NS处理. 在秸秆还田第15 d和30 d, SS处理的秸秆降解率显著高于NS处理, 增幅分别为14.16%和26.57%. 电镜观察了秸秆还田不同时间秸秆的显微结构(图 2), 结果表明随着还田时间的延长, 秸秆表面粗糙程度、碎片数量和降解孔洞均有增加. 对比分析SS和NS两处理的电镜图发现, SS处理秸秆碎片化程度更大, 且秸秆上降解孔洞也多于NS处理. 由此可知, 长期秸秆还田的土壤残留效应能够在一定时间内加快秸秆的降解.

|
*表示P < 0.05 图 1 不同处理的秸秆降解率 Fig. 1 Straw degradation rate under different treatments |

|
图 2 不同处理的秸杆扫描电镜图 Fig. 2 SEM images of straw under different treatments |
秸秆还田后的不同时间, SS和NS处理土壤中的细菌、真菌、氨化菌、亚硝化菌和纤维素降解菌的数量见表 1. 其中, 细菌数量在秸秆还田后迅速上升而后下降, 在第3 d两处理均达到最大, SS和NS处理的细菌数量分别为1.23×108 CFU·g-1和0.99×108 CFU·g-1, 但两者之间的细菌数量无显著差异. 对于真菌来说, 在秸秆还田后第3~60 d, SS处理真菌数量均显著高于NS处理(P < 0.05). 氨化菌数量在秸秆还田后呈现先上升后下降再上升的趋势, 在第7 d时氨化细菌的数量最高, NS和SS处理的氨化菌数量分别为7.99×107 CFU·g-1和5.87×107 CFU·g-1, 两个处理之间0~3 d的SS处理氨化细菌数量高于NS处理, 7~14 d SS处理低于NS处理, 但是除0 d外均未达到显著差异. 亚硝化菌数量之间的差异出现在前期, 秸秆还田后第3 d和7 d时NS处理土壤中亚硝化菌数量均显著高于SS处理(P < 0.05), 之后二者之间差异不显著. SS处理土壤中纤维素降解菌的数量在0、42和56 d较NS处理分别显著增加55.12%、85.68%和92.04%(P < 0.05), 说明在NS条件下, 土壤秸秆还田能够更加快速地促进纤维素降解菌的繁殖, 但随着易降解纤维类物质的分解, SS处理对纤维素降解菌的残留效应又显现出来.
|
|
表 1 不同处理的土壤可培养微生物数量1)/CFU·g-1 Table 1 Number of soil culturable microorganisms under different treatments/CFU·g-1 |
2.3 长期秸秆还田土壤残留效应对土壤微生物菌群结构的影响 2.3.1 ASVs数量的变化
图 3(a)和图 3(b)分别为两个处理的细菌和真菌ASVs的韦恩图. 如图 3(a)所示, NS特有细菌ASVs数量为7个、SS特有细菌ASVs数量为5个, 二者共有数量为1 876个, 占两处理ASVs总数的99.36%, 在细菌ASVs水平上SS对土壤细菌的残留效应非常小, 这与SS的残留效应对可培养细菌数量不存在显著差异相吻合. 由图 3(b)可知, 两个处理共有真菌ASVs数量为1 673个, 其中NS特有真菌ASVs数量为144个、SS特有真菌ASVs数量为38个, 共有ASVs占总数的90.19%, 说明SS的残留效应能够使其中独有的真菌ASVs减少.

|
图 3 不同处理的土壤细菌ASVs和真菌ASVs水平的韦恩图 Fig. 3 Venn diagram of soil bacteria and fungi at ASVs level under different treatments |
根据ACE指数、Chao1指数、Shannon指数和Simpson指数分析了两处理土壤细菌和真菌的多样性(表 2), 与NS处理相比, SS处理的细菌ACE指数、真菌ACE指数和Chao1指数显著降低(P < 0.05), 其他细菌和真菌多样性指数均不存在显著差异. 表明秸秆同时还田的情况下, SS的残留效应造成土壤细菌和真菌群落丰富度降低.
|
|
表 2 不同处理的土壤微生物α多样性指数1) Table 2 Soil microbial α diversity index under different treatments |
2.3.3 β多样性的变化
通过非度量多维尺度(NMDS)分析了两处理间土壤微生物群落的差异(图 4). NS处理的土壤细菌群落分布在第2、3和4象限, SS处理则主要分布于第1象限, β多样性应力值Stress = 0.015 8, r = 0.46, P = 0.001, 说明两处理的细菌群落区分度较为明显, 两处理间细菌菌群存在极显著差异[图 4(a)]. 对于真菌群落来说, NS和SS处理的真菌群落应力值Stress = 0.107 4, r = 0.32, P = 0.013, 两处理的真菌群落结构存在显著差异[图 4(b)]. 上述结果说明SS处理对土壤微生物菌群的定向选择和培育作用的残留效应较为明显.

|
蓝色椭圆线表示NS处理, 橙色椭圆线表示SS处理 图 4 基于Bray-curtis距离的土壤细菌群落和真菌群落的NMDS分析 Fig. 4 NMDS analysis of soil bacterial communities and fungal communities based on Bray-curtis distance |
SS和NS处理的土壤细菌群落门水平和属水平组成如图 5所示, 两处理土壤样品中细菌群落相对丰度较高的前10个菌门相同[图 5(a)], 其中变形菌门(Proteobacteria, 40.33%)和酸杆菌门(Acidobacteria, 26.32%)相对丰度较大, 占总序列的66.65%;其后依次为放线菌门(Actinobacteria)、芽单胞菌门(Gemmatimonadetes)、绿弯菌门(Chloroflexi)、拟杆菌门(Bacteroidetes)、己科河菌门(Rokubacteria)、硝化螺旋菌门(Nitrospirae)、厚壁菌门(Firmicutes)和浮霉菌门(Planctomycetes). 与NS处理相比, SS处理的酸杆菌门(Acidobacteria)、己科河菌门(Rokubacteria)和浮霉菌门(Planctomycetes)的相对丰度显著增加(P < 0.05), 增幅分别为25.92%、39.19%和45.17%;变形菌门(Proteobacteria)、放线菌门(Actinobacteria)和芽单胞菌门(Gemmatimonadetes)等相对丰度降低, 但差异不显著.

|
(a)门水平, (b)属水平 图 5 不同处理的土壤细菌门水平和属水平群落组成 Fig. 5 Bacteria community composition at phylum and genus levels under different treatments |
两处理土壤样品中细菌群落相对丰度较高的前10个菌属相同[图 5(b)], 其中Subgroup6纲未定属(uncultured_bacterium_c_Subgroup_6)相对丰度较大, 平均丰度为15.39%;其后依次为鞘氨醇单胞菌属(Sphingomonas)、芽单胞菌科未定属(uncultured_bacterium_f_Gemmatimonadaceae)、克雷伯氏菌属(Klebsiella)、罗库菌目未定属(uncultured_bacterium_o_Rokubacteriales)、微球菌科未定属(uncultured_bacterium_f_Micrococcaceae)、RB41、硝化螺旋菌属(Nitrospira)、不动杆菌属(Acinetobacter)和δ-变形菌纲未定属(uncultured_bacterium_c_Deltaproteobacteria). 与NS处理相比, SS处理中Subgroup6纲未定属(uncultured_bacterium_c_Subgroup_6)和罗库菌目未定属(uncultured_bacterium_o_Rokubacteriales)的相对丰度显著增加(P < 0.05), 增幅为31.26%和28.91%;克雷伯氏菌属(Klebsiella)、不动杆菌属(Acinetobacter)和鞘氨醇单胞菌属(Sphingomonas)等相对丰度降低, 但差异不显著. 可见, SS处理对土壤细菌群落的残留效应会造成细菌在门、属水平上均存在显著的差异.
2.3.5 对真菌群落组成的影响NS和SS处理土壤真菌群落门和属水平组成如图 6所示, 两处理土壤样品中真菌群落相对丰度前10位的菌门均相同[图 6(a)], 其中子囊菌门(Ascomycota, 66.41%)和担子菌门(Basidiomycota, 17.10%)相对丰度较大;之后依次为被孢霉门(Mortierellomycota)、壶菌门(Chytridiomycota)、罗兹菌门(Rozellomycota)、油壶菌门(Olpidiomycota)、捕虫霉亚门(Zoopagomycota)、球囊菌门(Glomeromycota)、毛霉门(Mucoromycota)和Aphelidiomycota. 与NS处理相比, SS处理显著增加了油壶菌门(Olpidiomycota)、捕虫霉亚门(Zoopagomycota)、球囊菌门(Glomeromycota)和毛霉门(Mucoromycota)的相对丰度(P < 0.05), 增幅分别为12.09%、13.35%、176.00%和94.54%;同时显著降低了子囊菌门(Ascomycota)、担子菌门(Basidiomycota)、罗兹菌门(Rozellomycota)和Aphelidiomycota的相对丰度(P < 0.05), 降幅分别为9.14%、1.62%、24.50%和88.00%.

|
(a)门水平, (b)属水平 图 6 不同处理的土壤真菌门水平和属水平群落组成 Fig. 6 Fungal community composition at phylum and genus levels under different treatments |
两处理土壤中优势真菌属也基本相同[图 6(b)], 其中镰刀菌属(Fusarium, 7.92%)和被孢霉菌属(Mortierella, 6.67%)相对丰度较大, 占总序列的14.59%;之后依次为白粉菌属(Erysiphe)、曲霉菌属(Aspergillus)、枝孢菌属(Cladosporium)、拟青霉菌属(Paecilomyces)、Papiliotrema、青霉菌属(Penicillium)、Moesziomyces和汉纳酵母菌属(Hannaella). 与NS处理相比, SS处理显著增加了拟青霉菌属(Paecilomyces)、青霉菌属(Penicillium)和Moesziomyces的相对丰度(P < 0.05), 增幅分别为5.48%、8.79%和2.98%;显著降低了镰刀菌属(Fusarium)的相对丰度(P < 0.05), 降幅为2.24%. 可以看出, SS处理对土壤真菌群落组成具有明显的残留效应.
2.3.6 分子生态网络分析基于NS处理和SS处理的测序结果构建了细菌分子生态网络图[图 7(a)和7(b)], 并确定了网络拓扑参数(表 3). 与NS处理相比, SS处理的细菌网络平均度升高、平均路径长度变短, 而网络节点数、模块性降低, 说明长期秸秆还田的残留效应使土壤细菌网络间互作程度增加、细菌间连接紧密度更高、细菌物种间关系复杂程度、菌群结构和功能分类程度降低[27]. 分析NS和SS处理的真菌网络拓扑参数(表 3)和网络图[图 8(a)和8(b)]发现, 与NS处理相比, SS处理的真菌网络节点数、模块性升高, 而平均度降低、平均路径长度升高, 说明SS处理的残留效应使土壤中真菌物种间关系更加复杂、真菌类群结构和功能分类程度更高、真菌菌群间网络连接紧密度降低. 本研究构建的4个网络均以正相关关系占优势, 表明了各处理的土壤微生物之间均以协同合作关系为主, 竞争关系较弱[28].

|
点的大小表示不同菌属丰度;点的不同颜色表示不同的模块;红线表示正相关, 绿线表示负相关 图 7 土壤细菌群落NS和SS处理的分子生态网络 Fig. 7 Bacterial communities molecular ecological networks of NS and SS treatments |
|
|
表 3 不同处理的土壤微生物群落分子生态网络拓扑性质 Table 3 Topological properties of molecular ecological network of soil microbial communities under different treatments |

|
点的大小表示不同菌属丰度;点的不同颜色表示不同的模块;红线表示正相关, 绿线表示负相关 图 8 土壤真菌群落NS和SS处理的分子生态网络 Fig. 8 Fungal communities molecular ecological networks of NS and SS treatments |
为了研究土壤微生物群落组成与秸秆降解率(第15 d)之间的关系, 进行了RDA分析. 由图 9(a)可知, 轴1和轴2分别解释细菌属水平群落结构变异的87.78%和9.96%. 细菌属水平群落组成与秸秆降解率(R2=0.98, P < 0.01, envfit函数检验)存在极显著相关关系. MND1、Steroidobacter和溶杆菌属(Lysobacter)等细菌属与秸秆降解率呈正相关关系;不动杆菌属(Acinetobacter)、硝化螺旋菌属(Nitrospira)和克雷伯氏菌属(Klebsiella)等细菌属与秸秆降解率呈负相关关系. 由图 9(b)可知, 轴1和轴2分别解释真菌属水平群落结构变异的90.39%和8.67%. 真菌属水平群落组成与秸秆降解率(R2=0.97, P < 0.01, envfit函数检验)存在极显著相关性. 青霉菌属(Penicillium)、被孢霉菌属(Mortierella)和拟青霉菌属(Paecilomyces)等真菌属与秸秆降解率呈正相关关系;曲霉菌属(Aspergillus)、镰刀菌属(Fusarium)和枝孢菌属(Cladosporium)等真菌属与秸秆降解率呈负相关关系.

|
图 9 不同处理下土壤微生物群落组成与秸秆降解率的RDA分析 Fig. 9 RDA analysis of soil microbial community composition and straw degradation rate under different treatments |
土壤微生物是农田生态系统中最为活跃的组分. 还田秸秆降解后能够为土壤微生物提供丰富的碳源和氮源, 从而促进了微生物的生长繁殖, 增加了土壤微生物数量[29], 改变了土壤微生物菌群的结构[19]. 有研究发现, 小麦秸秆堆肥还田[30]、玉米秸秆旋耕还田[31]和稻麦双季秸秆还田[32]具有增加土壤真菌和纤维素降解菌数量的作用. 这些微生物菌群的改变能够进一步加快还田秸秆的降解速率.
土壤微生物残留效应是指长期受某一种特定因素的影响形成特定的微生物特性, 当这一因素的影响终止后, 其对土壤微生物特性的影响仍然可以持续存在一段时间[33]. 为了验证秸秆长期还田带来的秸秆降解效应和微生物数量是否持续, 本研究向长期秸秆还田和未还田两种土壤中添加秸秆进行模拟试验, 发现长期秸秆还田土壤的秸秆降解率在15 d和30 d时均显著高于对照, 这种加快秸秆降解的效应采用电镜成像技术也能够得到验证[34]. 为了分析长期还田土壤中秸秆降解率高于秸秆未还田土壤的原因, 本研究测定了两种土壤同时秸秆还田60 d内, 土壤可培养细菌、真菌、氨化菌、亚硝化菌和纤维素降解菌的数量, 发现在长期秸秆还田土壤中真菌和纤维素降解菌的数量高于对照, 纤维素降解菌作为秸秆降解主要驱动力在长期秸秆还田土壤的富集从而加快了SS土壤中秸秆的降解, 这种效果显示了长期秸秆还田诱导的微生物效应能够在下茬秸秆还田后仍然显示残留效应. 同样, 这种残留效应在施用有机肥和种植作物后也均存在, Li等[22]的研究表明, 小麦季施用有机肥, 能够改变玉米季土壤碳、氮循环功能, 增加玉米的产量. Hannula等[35]的研究表明, 上茬作物的残留物质储存于根茬内并残留于土壤中, 通过残留效应影响下茬植物的生长.
3.2 长期秸秆还田残留效应能够富集具有秸秆降解作用的微生物菌群分子生物学和高通量测序技术的发展使得从群落组成、丰富度和结构等多个层面分析农艺措施对土壤微生物多样性的影响成为可能. 孔培君等[36]研究发现, 玉米秸秆还田配施化肥能够提高土壤细菌丰富度指数. 陈梦妮等[37]研究发现, 小麦-玉米双季秸秆还田可提高土壤真菌丰富度指数. 本研究对比分析了向SS和NS两种土壤同时进行秸秆还田7 d时细菌和真菌α多样性, 其中NS处理土壤细菌和真菌丰富度反而高于SS还田土壤, 并未呈现出SS的残留效应. 分析其原因可能是:与秸秆长期还田土壤SS相比, NS对秸秆还田刺激的短时效应更为明显, 从而使土壤中的微生物菌群多样性迅速增加. 本研究两处理的菌群结构发现, 与NS处理相比, SS土壤中酸杆菌门(Acidobacteria)、浮霉菌门(Planctomycetes)和己科河菌门(Rokubacteria)等土壤细菌优势菌门的相对丰度显著增加. Liu等[38]研究发现, 小麦秸秆还田和水稻-小麦双季秸秆还田均可增加土壤细菌中酸杆菌门的相对丰度, 表明秸秆还田定向微生物培育的残留效应在一定时间内能够持续. 酸杆菌门是土壤生态系统中的优势菌门[39], 在养分贫瘠和退化土壤中丰度更高, 能够利用难降解有机质作为营养物质生长, 其相对丰度与土壤有机质含量呈显著负相关[40]. 然而, 秸秆还田具有提高土壤有机质的作用, 其丰度增加可能是因为增加了较难降解的木质纤维素类有机质从而实现了酸杆菌门的定向诱导. 浮霉菌门是一种公认的温和寡营养型物种(moderate oligotrophs), 对不同的环境具有广泛的适应性. 有研究指出, 水稻-小麦双季秸秆还田能够降低浮霉菌门的相对丰度[41], 玉米秸秆还田也产生类似的效应[42], 这与本研究长期秸秆还田土壤中浮霉菌门显著增加结果相悖, 可能是因为SS和NS两种土壤中的微生物菌群对秸秆还田的应激反应存在较大的差异.
3.3 长期秸秆还田对微生物生态网络具有明显的残留效应微生物不是孤立地存在, 而是处在复杂的关联网络中. 众多研究表明, 连接更紧密的网络能够促进有机碳的分解和利用[43]. 土壤微生物群落能够通过调整自身的菌群结构来适应环境和养分的不断变化, 秸秆还田释放的特定养分使土壤微生物的生态网络更加紧密和复杂[44], 并促进特定微生物网络模块内物种的生长, 形成强烈的共生关系, 从而提高微生物功能和活性, 加快秸秆的降解[14]. 其中纤维素分解微生物作为微生物网络中的关键菌群不仅在玉米秸秆降解过程中发挥作用, 而且也起到了稳定堆肥网络的作用[45]. 本研究对比分析SS和NS土壤秸秆还田后的微生物生态网络结构发现, 前者的细菌网络物种互作程度高、且网络连接更为紧密, 真菌网络更加复杂和稳定. 这与Tang等[46]研究的秸秆还田当季微生物网络的反应结果一致, 表明对于土壤微生物分子网络来说, 长期秸秆还田具有明显的残留效应. 总之, 微生物是秸秆降解的驱动力, 长期秸秆还田具有明显的土壤微生物效应, 且该效应能够在一定时间内残留.
4 结论(1)与秸秆未还田土壤相比, 长期秸秆还田的土壤残留效应能够显著增加秸秆还田后15 ~30 d时的秸秆降解率.
(2)长期秸秆还田的土壤残留效应增加了土壤中的真菌和纤维素降解菌的数量, 降低了土壤微生物群落丰富度, 富集了具有秸秆降解作用的微生物菌群.
(3)长期秸秆还田土壤的残留效应还表现在加强了细菌网络物种互作程度和连接紧密性以及真菌网络复杂性和稳定性.
| [1] | Hou Y P, Xu X P, Kong L L, et al. The combination of straw return and appropriate K fertilizer amounts enhances both soil health and rice yield in Northeast China[J]. Agronomy Journal, 2021, 113(6): 5424-5435. DOI:10.1002/agj2.20805 |
| [2] |
高晶霞, 高昱, 牛勇琴, 等. 不同作物秸秆腐解对连作辣椒生长及根际环境的影响[J]. 西北农业学报, 2021, 30(8): 1220-1226. Gao J X, Gao Y, Niu Y Q, et al. Effect of decomposition of different crop straws on growth and rhizosphere environment of continuous cropping pepper[J]. Acta Agriculturae Boreali-occidentalis Sinica, 2021, 30(8): 1220-1226. |
| [3] | Gao Z P. Quantity and utilization of crop straw resources in China[J]. IOP Conference Series: Earth and Environmental Science, 2020, 598. DOI:10.1088/1755-1315/598/1/012103 |
| [4] | Zhang H Y, Liu Q J, Liu S T, et al. Key soil properties influencing infiltration capacity after long-term straw incorporation in a wheat (Triticum aestivum L.)–maize (Zea mays L.) rotation system[J]. Agriculture, 2023, 344. DOI:10.1016/J.AGEE.2022.108301 |
| [5] |
岳杨, 孙盼盼, 张艳辉, 等. 秸秆还田方式对土壤物理性状及春玉米产量的影响[J]. 湖北农业科学, 2021, 60(11): 35-38. Yue Y, Sun P P, Zhang Y H, et al. Effects of returning methods of straw on soil physical property and spring maize yield[J]. Hubei Agricultural Sciences, 2021, 60(11): 35-38. |
| [6] |
马胜兰, 况福虹, 林洪羽, 等. 秸秆还田量对川中丘陵冬小麦-夏玉米轮作体系土壤物理特性的影响[J]. 中国农业科学, 2023, 56(7): 1344-1358. Ma S L, Kuang F H, Lin H Y, et al. Effects of straw incorporation quantity on soil physical characteristics of winter wheat-summer maize rotation system in the central hilly area of Sichuan Basin[J]. Scientia Agricultura Sinica, 2023, 56(7): 1344-1358. |
| [7] | Su Y, Yu M, Xi H, et al. Soil microbial community shifts with long-term of different straw return in wheat-corn rotation system[J]. Scientific Reports, 2020, 10(1). DOI:10.1038/s41598-020-63409-6 |
| [8] |
马垒, 李燕, 魏建林, 等. 长期秸秆还田对潮土真菌群落、酶活性和小麦产量的影响[J]. 环境科学, 2022, 43(10): 4755-4764. Ma L, Li Y, Wei J L, et al. Effects of long-term straw returning on fungal community, enzyme activity and wheat yield in fluvo-aquic soil[J]. Environmental Science, 2022, 43(10): 4755-4764. |
| [9] |
靳玉婷, 李先藩, 蔡影, 等. 秸秆还田配施化肥对稻-油轮作土壤酶活性及微生物群落结构的影响[J]. 环境科学, 2021, 42(8): 3985-3996. Jin Y T, Li X F, Cai Y, et al. Effects of straw returning with chemical fertilizer on soil enzyme activities and microbial community structure in rice-rape rotation[J]. Environmental Science, 2021, 42(8): 3985-3996. |
| [10] |
周永学, 陈静, 李远, 等. 棉秆还田对咸水滴灌棉田土壤酶活性和细菌群落结构多样性的影响[J]. 环境科学, 2022, 43(4): 2192-2203. Zhou Y X, Chen J, Li Y, et al. Effects of cotton stalk returning on soil enzyme activity and bacterial community structure diversity in cotton field with long-term saline water irrigation[J]. Environmental Science, 2022, 43(4): 2192-2203. |
| [11] |
王子阳, 陈婉华, 袁伟, 等. 双季稻地区不同类型水稻秸秆与还田深度对还田秸秆腐解进程的影响[J]. 中国土壤与肥料, 2022(2): 170-174. Wang Z Y, Chen W H, Yuan W, et al. Effects of different types of rice straw and returning depth on decomposition course of straw in double cropping rice region[J]. Soil and Fertilizer Sciences in China, 2022(2): 170-174. |
| [12] |
王晶, 孙松青, 李雪, 等. 秸秆不同还田方式对轮作小麦—玉米产量、土壤养分及蔗糖酶活性的影响[J]. 江苏农业科学, 2023, 51(4): 85-90. Wang J, Sun S Q, Li X, et al. Effects of different straw returning methods on yield, soil nutrients and sucrase activity of wheat-maize rotation[J]. Jiangsu Agricultural Sciences, 2023, 51(4): 85-90. |
| [13] | Cui H X, Luo Y L, Chen J, et al. Straw return strategies to improve soil properties and crop productivity in a winter wheat-summer maize cropping system[J]. European Journal of Agronomy, 2022, 133. DOI:10.1016/J.EJA.2021.126436 |
| [14] |
马垒, 李燕, 魏建林, 等. 连续施用化肥及秸秆还田对潮土酶活性、细菌群落和分子生态网络的影响[J]. 植物营养与肥料学报, 2022, 28(8): 1353-1363. Ma L, Li Y, Wei J L, et al. Effects of continuous chemical fertilizer application and straw returning on soil enzyme activity, bacterial community and co-occurrence patterns in a fluvo-aquic soil[J]. Journal of Plant Nutrition and Fertilizers, 2022, 28(8): 1353-1363. |
| [15] |
纪程, 孙玉香, 孟圆, 等. 稻麦轮作体系长期秸秆还田对土壤真菌群落结构及秸秆降解潜力的影响[J]. 农业环境科学学报, 2022, 41(4): 819-825. Ji C, Sun Y X, Meng Y, et al. Effects of long-term straw incorporation on soil fungal community structure and straw decomposition potential in a rice–wheat rotation system[J]. Journal of Agro-Environment Science, 2022, 41(4): 819-825. |
| [16] |
高日平, 王伟妮, 狄彩霞, 等. 有机肥和秸秆还田对河套灌区盐渍化土壤养分及微生物数量的影响[J]. 中国土壤与肥料, 2023(2): 43-53. Gao R P, Wang W N, Di C X, et al. Effects of organic manure and straw returning on nutrient and microbial quantity of saline soil in river-loop irrigation area[J]. Soil and Fertilizer Sciences in China, 2023(2): 43-53. |
| [17] | Korav S, Yadav D B, Yadav A, et al. Rice residue management alternatives in rice–wheat cropping system: impact on wheat productivity, soil organic carbon, water and microbial dynamics[J]. Scientific Reports, 2024, 14(1). DOI:10.1038/s41598-024-52319-6 |
| [18] |
张鑫, 周卫, 艾超, 等. 秸秆还田下氮肥运筹对夏玉米不同时期土壤酶活性及细菌群落结构的影响[J]. 植物营养与肥料学报, 2020, 26(2): 295-306. Zhang X, Zhou W, Ai C, et al. Effects of nitrogen management on soil enzyme activities and bacterial community structure in summer maize growing stagesunder straw incorporation[J]. Journal of Plant Nutrition and Fertilizers, 2020, 26(2): 295-306. |
| [19] | Yan S S, Song J M, Fan J S, et al. Changes in soil organic carbon fractions and microbial community under rice straw return in Northeast China[J]. Global Ecology and Conservation, 2020, 22. DOI:10.1016/j.gecco.2020.e00962 |
| [20] |
郭晓雯, 陈静, 鲁晓宇, 等. 生物炭和秸秆还田对微咸水滴灌棉田土壤真菌群落结构多样性的影响[J]. 环境科学, 2022, 43(9): 4625-4635. Guo X W, Chen J, Lu X Y, et al. Effects of biochar and straw returning on soil fungal community structure diversity in cotton field with long-term brackish water irrigation[J]. Environmental Science, 2022, 43(9): 4625-4635. |
| [21] |
李露露, 张宁, 张业承, 等. 不同耕作方式、秸秆还田配施肥料条件下夏玉米磷素高效利用研究[J]. 山东农业科学, 2023, 55(9): 94-100. Li L L, Zhang N, Zhang Y C, et al. Phosphorus efficient utilization of summer maize under different tillage methods, straw returning and fertilizer application[J]. Shandong Agricultural Sciences, 2023, 55(9): 94-100. |
| [22] | Li G C, Niu W Q, Ma L, et al. Legacy effects of wheat season organic fertilizer addition on microbial co-occurrence networks, soil function, and yield of the subsequent maize season in a wheat-maize rotation system[J]. Journal of Environmental Management, 2023, 347. DOI:10.1016/J.JENVMAN.2023.119160 |
| [23] |
张敏华, 吕惠生, 吕春柳, 等. 超临界CO2处理玉米秸秆残留木质素形态及脱除研究[J]. 酿酒科技, 2018(3): 87-92, 97. Zhang M H, Lü H S, Lü C L, et al. Removal of residual lignin in corn stover treated by scCO2 [J]. Liquor-Making Science & Technology, 2018(3): 87-92, 97. |
| [24] |
唐海明, 石丽红, 文丽, 等. 长期施肥对双季稻田根际土壤氮素的影响[J]. 生态环境学报, 2023, 32(3): 492-499. Tang H M, Shi L H, Wen L, et al. Effects of different long-term fertilizer managements on rhizosphere soil nitrogen in the double-cropping rice field[J]. Ecology and Environmental Sciences, 2023, 32(3): 492-499. |
| [25] | Wang Y, Sheng H F, He Y, et al. Comparison of the levels of bacterial diversity in freshwater, intertidal wetland, and marine sediments by using millions of illumina tags[J]. Applied and Environmental Microbiology, 2012, 78(23): 8264-8271. |
| [26] |
程济南, 金辉, 许忠祥, 等. 甘肃典型高寒草原退化植物瑞香狼毒对根际土壤微生物群落的影响研究[J]. 微生物学报, 2021, 61(11): 3686-3704. Cheng J N, Jin H, Xu Z X, et al. Effects of degraded plant Stellera chamaejasme L. on the rhizosphere soil microbial communities in typical alpine grassland, Gansu Province[J]. Acta Microbiologica Sinica, 2021, 61(11): 3686-3704. |
| [27] | Xu Z Y, Sun R H, He T Y, et al. Disentangling the impact of straw incorporation on soil microbial communities: enhanced network complexity and ecological stochasticity[J]. Science of the Total Environment, 2023, 863. DOI:10.1016/J.SCITOTENV.2022.160918 |
| [28] |
夏皖豫, 陈彦云. 粉垄耕作对耕地土壤细菌群落多样性及微生物网络结构的影响[J]. 环境科学, 2023, 44(2): 1095-1103. Xia W Y, Chen Y Y. Effect of deep vertical rotary tillage on soil bacterial community diversity and microbial network structure in cultivated land[J]. Environmental Science, 2023, 44(2): 1095-1103. |
| [29] | Six J, Bossuyt H, Degryze S, et al. A history of research on the link between (micro)aggregates, soil biota, and soil organic matter dynamics[J]. Soil and Tillage Research, 2004, 79(1): 7-31. |
| [30] | Yang Y F, Zhang S J, Li N, et al. Metagenomic insights into effects of wheat straw compost fertiliser application on microbial community composition and function in tobacco rhizosphere soil[J]. Scientific Reports, 2019, 9(1). DOI:10.1038/s41598-019-42667-z |
| [31] | Yang Y, Long Y, Li S W, et al. Straw return decomposition characteristics and effects on soil nutrients and maize yield[J]. Agriculture, 2023, 13(8). DOI:10.3390/agriculture13081570 |
| [32] | Guan Y P, Wu M K, Che S H, et al. Effects of continuous straw returning on soil functional microorganisms and microbial communities[J]. Journal of Microbiology, 2023, 61(1): 49-62. |
| [33] |
郎明, 张春燕, 张俊伶. 长期不同供磷水平驯化的土壤微生物的遗留效应[J]. 土壤学报, 2022, 59(6): 1650-1659. Lang M, Zhang C Y, Zhang J L. The soil microbial legacy effects of long-term gradient P fertilization based on the analysis of plant growth, nutrient absorption, soil enzyme activity and mycorrhizal characteristics[J]. Acta Pedologica Sinica, 2022, 59(6): 1650-1659. |
| [34] |
黄亚丽, 黄媛媛, 马慧媛, 等. 低温秸秆降解真菌的筛选及在秸秆还田中的应用[J]. 中国农学通报, 2020, 36(21): 53-60. Huang Y L, Huang Y Y, Ma H Y, et al. Fungi with corn straw decomposing characterization at low-temperature: screening and application in straw returning field[J]. Chinese Agricultural Science Bulletin, 2020, 36(21): 53-60. |
| [35] | Hannula S E, Heinen R, Huberty M, et al. Persistence of plant-mediated microbial soil legacy effects in soil and inside roots[J]. Nature Communications, 2021, 12(1). DOI:10.1038/s41467-021-25971-z |
| [36] |
孔培君, 郑洁, 栾璐, 等. 不同秸秆还田方式对旱地红壤细菌群落、有机碳矿化及玉米产量的影响[J]. 环境科学, 2021, 42(12): 6047-6057. Kong P J, Zheng J, Luan L, et al. Effects of different types of straw returning on the bacterial community, organic carbon mineralization and maize yield in upland red soil[J]. Environmental Science, 2021, 42(12): 6047-6057. |
| [37] |
陈梦妮, 李永山, 王慧, 等. 麦田土壤真菌多样性对麦玉轮作长期秸秆还田和配施有机肥的响应[J]. 微生物学通报, 2023, 50(6): 2481-2496. Chen M N, Li Y S, Wang H, et al. Responses of soil fungal diversity to long-term straw incorporation and manure application in the field with winter wheat-summer maize rotation[J]. Microbiology China, 2023, 50(6): 2481-2496. |
| [38] | Liu X Q, Liu H R, Zhang Y S, et al. Straw return drives soil microbial community assemblage to change metabolic processes for soil quality amendment in a rice-wheat rotation system[J]. Soil Biology and Biochemistry, 2023, 185. DOI:10.1016/J.SOILBIO.2023.109131 |
| [39] |
傅敏, 郝敏敏, 胡恒宇, 等. 土壤有机碳和微生物群落结构对多年不同耕作方式与秸秆还田的响应[J]. 应用生态学报, 2019, 30(9): 3183-3194. Fu M, Hao M M, Hu H Y, et al. Responses of soil organic carbon and microbial community structure to different tillage patterns and straw returning for multiple years[J]. Chinese Journal of Applied Ecology, 2019, 30(9): 3183-3194. |
| [40] | Fierer N, Bradford M A, Jackson R B. Toward an ecological classification of soil bacteria[J]. Ecology, 2007, 88(6): 1354-1364. |
| [41] | 张世洁. 秸秆还田对稻田硝态氮异化还原的影响及微生物学机制研究[D]. 北京: 中国科学院南京土壤研究所, 2022. |
| [42] |
徐忠山, 刘景辉, 逯晓萍, 等. 秸秆颗粒还田对黑土土壤酶活性及细菌群落的影响[J]. 生态学报, 2019, 39(12): 4347-4355. Xu Z S, Liu J H, Lu X P, et al. Effects of returning granulated corn stover on soil enzyme activities and bacterial community in black soil[J]. Acta Ecologica Sinica, 2019, 39(12): 4347-4355. |
| [43] | Lang D M, Zhou R J, Hao F G, et al. Effect of wheat straw addition on organic carbon mineralisation and bacterial community in orchard soil[J]. Journal of Soil Science and Plant Nutrition, 2023, 23(3): 4328-4341. |
| [44] | Schimel J P, Schaeffer S M. Microbial control over carbon cycling in soil[J]. Frontiers in Microbiology, 2012, 3. DOI:10.3389/fmicb.2012.00348 |
| [45] | Han Y, Liu W Y, Chang N, et al. Exploration of β-glucosidase-producing microorganisms community structure and key communities driving cellulose degradation during composting of pure corn straw by multi-interaction analysis[J]. Journal of Environmental Management, 2023, 325. DOI:10.1016/j.jenvman.2022.116694 |
| [46] | Tang L L, Xia Y, Fan C, et al. Control of Fusarium wilt by wheat straw is associated with microbial network changes in watermelon rhizosphere[J]. Scientific Reports, 2020, 10(1). DOI:10.1038/s41598-020-69623-6 |