 2024, Vol. 45
2024, Vol. 45

2. 江西师范大学鄱阳湖湿地与流域研究教育部重点实验室, 南昌 330022
 , DING Ming-jun1,2 , ZHANG Hua1,2
, DING Ming-jun1,2 , ZHANG Hua1,2 , ZHANG Yue-ju1 , HUANG Peng1 , WU Yu-ping1 , ZOU Tian-e1 , WANG Neng-yu1 , ZENG Huan1
, ZHANG Yue-ju1 , HUANG Peng1 , WU Yu-ping1 , ZOU Tian-e1 , WANG Neng-yu1 , ZENG Huan1
2. Key Laboratory of Poyang Lake Wetland and Watershed Research, Ministry of Education, Jiangxi Normal University, Nanchang 330022, China
青藏高原是我国天然草地分布面积最大的区域, 约占全国草地面积的1/3[1], 高寒草原是青藏高原广泛分布的主要植被类型之一, 约占青藏高原草地的30.98%, 是我国草地生态系统的重要组成部分[2, 3]. 与高寒草甸一样, 高寒草原在区域生态系统服务和生态系统功能上发挥着重要的作用[4]. 青藏高原独特的“高”“寒”自然环境, 造就了高寒草原脆弱的生态系统, 对区域气候变化和人类活动响应敏感[5], 近年来, 伴随着气候变化和人类活动的双重压力, 高寒草地的退化日趋严重, 据报道过去40年间1/5的高寒草原发生了严重的退化[6]. 高寒草原退化演替受众多因素的共同驱动, 自然因素中如长期干旱、风蚀、水蚀、沙尘暴、鼠、虫害以及种间竞争等;人为因素中如放牧、滥垦、樵采和开矿等. 高寒草原的退化会对草地生态系统服务功能的发挥造成严重威胁, 并可能威胁下游的生态安全和可持续发展[7, 8].
作为最为复杂的生物系统, 土壤可以为植物生长提供所需的水和营养物质, 也是生物地球化学循环发生的主要场所[9]. 微生物是土壤中各种生态过程的主要参与者, 不同的微生物在土壤养分循环中执行不同的功能, 如分解有机物和凋落物[10]. 土壤微生物在执行不同生态功能的过程中会受到生物和非生物因素的影响, 如土壤理化性质及植被特征的调节. 已有研究表明, 高寒草地退化导致植被特征(生物量、盖度、多样性)和土壤因子(营养状况、物理性质)发生了显著的改变[11 ~ 16]. 了解高寒草原退化引起的植被特征和土壤因子改变如何影响土壤微生物群落特征, 对于草地的管理和健康评价至关重要. 以往研究结果表明, 高寒草原退化会导致植被群落多样性和功能的改变[17 ~ 19]、植被群落特征与土壤养分的互作关系的改变[20 ~ 22]. 相对于高寒草甸, 高寒草原退化过程中土壤微生物群落的响应研究仍然有限[6], 且对于高寒草原退化过程中植被特征和土壤因子对微生物的交互影响及调节作用还需深入探讨. 此外, 微生物在土壤中存在复杂的相互作用, 参与形成生态网络并维持系统功能[23], 保护微生物的“相互作用”对于生态系统管理至关重要[24]. 土壤微生物之间的正向相互作用可以促进合作、资源共享和互利共生, 例如植物根际微生物与植物根系之间的相互作用, 植物通过根系分泌物提供碳源和营养物质, 而根际微生物可以提供植物所需的养分和保护植物免受病原体侵害[25]. 负向相互作用则可以导致竞争、资源争夺, 一些微生物可以通过产生抗生素或其他抑制物质来抑制其他微生物的生长. 相互作用的变化可能会改变土壤微生物群体结构和多样性, 而多样性的损失会影响植物养分获取, 进一步损害生态系统功能[26]. 土壤微生物多样性影响多个生态系统过程, 包括生物地球化学循环, 这些循环对植物的生长和营养吸收非常重要, 并通过食物链作用进一步影响整个生态系统的能量流动和物质循环[27]. 土壤微生物的多样性和功能对生态系统的稳定性至关重要, 土壤微生物通过相互作用和功能协同, 建立了复杂的微生物网络, 维持了生态系统的稳定性和功能稳定[28]. 目前对于退化过程中网络的复杂性和稳定性(拓扑特征)如何响应等相关研究仍稍显不足.
鉴于此, 本文以青藏高原高寒草原为研究对象, 综合植被盖度、高度及优势种重要值等植被特征, 划分不同退化等级, 比较不同退化阶段植被特征和土壤因子的差异;分析高寒草原不同退化过程中微生物群落的变化特征(组成、多样性及共线网络拓扑特征);探讨不同阶段植被特征和土壤因子对微生物群落的交互影响及调节作用, 以期为高寒草原健康评价和管理提供重要依据.
1 材料与方法 1.1 研究区概况三江源区位于青藏高原腹地, 地理位置为东经89°24′~102°41′, 北纬31°39′~36°16′, 海拔在3 450~6 621 m之间, 面积达36.6万km²[29]. 三江源区为典型的高原大陆性气候, 年平均气温在-5.38~4.14℃之间, 年平均降水量在262.2~772.8 mm之间, 干湿两季分明, 冷热两季交替, 年温差小、日温差大、日照时间长、辐射强烈、无四季区分[30]. 植被以耐寒旱生的多年生丛生禾草、根茎苔草和小半灌木为建群种, 草丛低矮、层次简单、草群稀疏, 覆盖度小, 伴生适应高寒生境的垫状植物层片, 具有生长季节短、生物产量较低等特点. 优势物种以紫花针茅(Stipa purpurea)、二裂委陵菜(Potentilla bifurca)和火绒草(Leontopodium nanum)为主[31].
1.2 实验设计与样品采集在植物生长旺盛季节(7~8月)在三江源进行样品采集, 一共选择了12个样点, 每个样点设置3个不同退化类型的样地, 每个样地随机设置3个1 m×1 m的样方, 对样方内植物物种组成、丰富度、多度及地上生物量等植被群落特征进行调查. 采集3个样方0~10 cm的根际土壤混合为一个样品用于土壤理化性质和微生物的测定. 野外采用环刀取样, 进行土壤含水量及容重的测定, 采集同样深度的根际土壤样品一份于聚氯乙烯自封袋保存, 带回实验室待测;同时现场采集过灭菌后土壤筛(2 mm)的土壤一份在液氮容器中保存, 随即送到上海美吉生物医药科技公司进行微生物测序.
基于样方调查结果, 并根据马玉寿等[32]及天然草地退化、沙化和盐渍化的分级指标[33]发现, 野外设置的部分样方不符合相应的退化标准, 故将不符合退化标准的样方剔除, 最终保留符合退化标准的样方24个, 其中未退化草地7份, 中度退化草地10份, 重度退化草地7份(图 1).

|
图 1 研究区采样点示意 Fig. 1 Sample sites of study area |
用于测试土壤理化性质的样品在室内风干, 土壤样品过10目筛, 使用电导率计通过电位法以1∶5的土水比测量电导率, 土壤pH采用电位法进行测定[28]. 土壤样品过100目筛, 用3种酸(硝酸、氢氟酸和高氯酸)微波消解后, 用钼酸铵-锑抗比色法测定土壤总磷(TP)[29], 利用火焰光度法测定土壤全钾(TK)[30], 采用凯氏定氮法(LY/T 1232-1999)测定土壤全氮含量;有机质测定采用H2SO4-K2Cr2O7油浴氧化滴定法(NY/T 1121.6-2006), 过10目筛的土壤样品用过氧化氢和盐酸除去土壤有机质后, 用激光散射粒度分布分析仪(HORIBA, LA-950, 日本)测定黏粒含量[25, 31].
物种重要值是度量群落水平反应的综合数量指标:
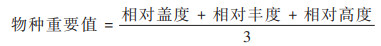
|
(1) |
丰富度指数是描述群落中所含物种丰富程度的数量指标:

|
(2) |
式中, S表示物种总数;A表示样方面积.
香农-威纳指数用来估算群落多样性的高低:
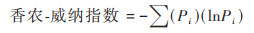
|
(3) |
式中, Pi表示第i种在全体物种中的重要性比例, 如以个体数量而言, ni表示第i种的个体数量, N表示总个体数量, 则有:
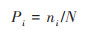
|
(4) |
辛普森多样性指数反映了群落中最常见的物种, 评估群落丰富度:

|
(5) |
式中, Pi表示物种i的相对重要值.
1.4 高通量DNA测序和生物信息学分析根据E.Z.N.A.® soil试剂盒(Omega Bio-tek, U.S.)说明书进行总DNA提取, 并使用Nanodrop检验DNA质量和浓度, 利用1%琼脂糖凝胶电泳检测DNA提取质量. 使用引物338F和806R扩增土壤细菌16S rRNA基因的V3-V4区[39], 使用引物KYO2F 1F和ITS 4R扩增土壤真菌ITS rRNA基因的ITS2区. 在20 µL反应体系中进行PCR扩增, 在95℃下变性3 min, 在95℃下30 s, 在55℃下退火30 s, 在72℃下延伸45 s的27个循环, 以及在72℃下最终延伸10 min. 将PCR产物在2%琼脂糖凝胶上提取, 然后用2%琼脂糖凝胶电泳检测, 使用AxyPrep DNA凝胶回收试剂盒(AXYGEN公司)切胶回收PCR产物, 然后加入接头引物、酶及Mix进行PCR富集, 使用NEXTFLEX Rapid DNA-Seq Kit完成文库构建, 文库在MiseqPE300平台上机测序, 使用Fastp软件对原始测序序列进行质控, 使用Flash软件进行拼接, 根据97%的相似度阈值对序列进行OTUs聚类并剔除嵌合体, 得到物种分类学注释结果. 以上过程均委托上海美吉生物医药科技有限公司完成.
1.5 数据处理与分析本研究采用R 4.0.2软件进行数据处理, 使用ggplot2包和ggpubr包来分析不同退化程度植被特征与土壤因子的差异;利用VennDiagram包分析不同退化程度高寒草原中共有和特有的OTUs, ggplot2包绘制物种丰度图, 利用线性判别分析(LDA)识别出不同退化阶段中存在显著差异的微生物;利用vegan包来比较不同退化阶段高寒草原土壤微生物群落的α多样性指数的差异;采用vegan包中的“bray”函数进行主坐标分析(PCoA)、基于Bray-Curtis进行β多样性分析, 检验不同退化阶段高寒草原土壤微生物群落的β多样性差异;基于igraph包构建无向的相关性网络, 并借助Gephi绘制网络图, 同时计算不同退化程度土壤微生物的网络拓扑特征, 分析不同退化程度高寒草原网络拓扑特征的差异;采用vegan包进行基于相似或相异度矩阵的多元回归(MRM)分析来探究植被特征和土壤因子对高寒草原土壤微生物群落的交互影响, 为了避免多重共现性的影响, 使用R语言Hmisc包的varclus()函数来评估环境变量的冗余度, 剔除冗余度很高的环境变量, 将剩余变量进行MRM分析;最后利用冗余分析(redundancy analysis, RDA)和典型相关分析(CCA)来确定影响高寒草原土壤微生物群落的关键环境因子.
2 结果与分析 2.1 高寒草原退化过程中植被特征和土壤理化性质的变化如图 2所示, 随着退化的发生, 群落植被盖度、植被高度、禾本科重要值及植物丰富度指数、地上生物量均呈现显著地下降趋势[P < 0.05, 图2(a)~2(e)], 而香农-威纳指数差异并不显著[P > 0.05, 图 2(f)];土壤有机质、全氮、全磷和粉粒含量随着退化的发生呈显著下降趋势[P < 0.05, 图2(k)、2(l)、2(p)和2(n)]. 而土壤容重、砂粒含量和全钾含量显著上升[P < 0.01, 图2(j)、2(o)和2(q)], 而土壤pH、电导率和土壤含水量和黏粒含量未发生显著变化.

|
*表示P < 0.05, **表示P < 0.01, ns表示P≥0.05 图 2 不同退化阶段高寒草原植被特征和土壤理化性质的差异比较 Fig. 2 Comparisons of vegetation characteristics and soil physical and chemical properties in the alpine steppe under different degradation levels |
将高质量测序序列基于97%的相似性进行聚类, 共得到7 714个细菌OTUs和3 966个真菌OTUs. 不同退化阶段高寒草原之间共有的细菌和真菌OTUs分别有3 339和513个, 分别占细菌和真菌总OTUs的45.47%和15.07%;细菌所有OTUs可归类39个门, 122个纲, 956个属;真菌所有OTUs可归类为14个门, 45个纲, 597个属. 随着退化程度的增加, 细菌和真菌每个阶段独有的OUTs数量和相对丰度表现为先下降后上升的趋势;相邻阶段共有的OTUs量表现为上升的趋势[图3(a)和3(b)]. 高寒草原土壤优势菌门为:放线菌门(32.14%, 相对丰度, 下同)、变形菌门(22.28%)、酸杆菌门(14.94%)、绿弯菌门(14.59%);真菌的优势菌门为:子囊菌门(68.59%)、担子菌门(13.55%)和被孢菌门(9.58%). 随着退化程度的增加, 细菌和真菌的优势菌门并未发生明显的改变, 但相对丰度发生了一定的变化, 如变形菌门相对丰度在重度退化阶段较低, 而酸杆菌门相对丰度在未退化水平最高;子囊菌门和担子菌门相对丰度在重度退化水平下较低[图3(c)和3(d)].

|
(a)和(b)分别表示不同退化阶段细菌与真菌共有与特有的OTUs, 其中括号内数值表示不同退化阶段高寒草原细菌、真菌共有和特有OTUs所占的比例;(c)和(d)分别表示不同退化阶段细菌与真菌门水平物种组成的相对丰度;(e)和(f)分别表示不同退化阶段细菌与真菌目水平物种组成的相对丰度(相对丰度 > 1%);1.未退化, 2.中度退化, 3.重度退化 图 3 高寒草原不同退化阶段土壤微生物群落组成 Fig. 3 Microbial community composition of the alpine steppe under different degradation levels |
在目水平上, 高寒草原土壤细菌平均相对丰度前10从高到低依次为Rhizobiales(8.56%)、Vicinamibacterales(8.27%)、Solirubrobacterales(5.65%)、Thermomicrobiales(5.31%)、Rubrobacterales(3.78%)、Pyrinomonadales(3.57%)、Micrococcales(3.37%)、Burkholderiales(2.99%)、Propionibacteriales(2.93%)和Gemmatimonadales(2.69%). 而土壤真菌平均相对丰度从高到低依次为Pleosporales(18.75%)、Hypocreales(9.92%)、Mortierellales(8.88%)、Helotiales(6.56%)、unclassified_k__Fungi(5.34%)、Agaricales(5.3%)、Capnodiales(4.59%)、Sordariales(3.97%)、Chaetothyriales(3.85%)和Filobasidiales(3.77%). 随着退化程度的增加, 排名前10的土壤微生物没有改变, 但其相对丰度发生了一定的变化, 如变形菌门的Rhizobiales相对丰度在未退化最高(10.09%), 而在重度退化最低(5.72%), 酸杆菌门的Vicinamibacterales相对丰度在重度退化中最高(10.4%). 子囊菌门的Pleosporales相对丰度在未退化中最低(14.78%), 在中度退化中最高(22.85%)[图3(e)和3(f)].
运用线性判别分析(LDA)方法来确定不同退化阶段各分类水平上具有统计学差异的生物标识, LDA的阈值设置为3.5. 不同退化阶段的高寒草原中有23个细菌分类群表现出显著差异, 由退化引起的土壤细菌分类群的显著变化主要与酸杆菌门的c__Blastocatellia、o__Blastocatellales、f__Blastocatellaceae和变形菌门的o__Burkholderiales、o__Rhizobiales、f__Xanthobacteraceae有关;而23个真菌分支在不同退化程度的高寒草原中表现出了显著差异, 分别是子囊菌门的o__Chaetothyriales、f__Lasiosphaeriaceae、f__Herpotrichiellaceae、g__Schizothecium、g__Coniochaeta和担子菌门的g__Naganishia、c__Ustilaginomycete、g__Ustilago(图 4).

|
图 4 高寒草原不同退化阶段土壤微生物的线性判别分析 Fig. 4 LDA of soil microorganisms at different degradation stages in alpine steppe |
对不同退化阶段高寒草原微生物α多样性的变化分析表明, 细菌和真菌群落的Richness指数、Chao1指数、ACE指数和Sobs指数在不同退化阶段均存在显著的差异, 但细菌和真菌α多样性指数的表现略有不同, 细菌α多样性随着退化均表现为上升后下降的趋势, 而真菌α多样性随着退化都表现显著下降的趋势(图 5), 微生物Shannon指数和Simpson指数在不同退化阶段并没有表现出显著差异(P > 0.05).

|
(a)~(f)表示土壤细菌α多样性变化, (g)~(l)表示土壤真菌α多样性变化;*表示P < 0.05, **表示P < 0.01, ns表示P ≥0.05 图 5 高寒草原不同退化阶段土壤微生物群落α多样性变化 Fig. 5 The α diversity of soil microbial community changes in different degradation stages of alpine steppe |
高寒草原不同退化阶段微生物β多样性的分析结果表明(图 6), PCoA分析显示, 主成分轴PCA1和PCA2分别解释了20.6%和16.1%细菌群落结构差异, 细菌群落β多样性整体的差异显著(P=0.013 < 0.05);PCoA分析主成分轴PCA1和PCA2分别解释了9.8%和8.7%真菌群落结构差异, 解释度较低, 表明土壤真菌群落结构是高维复杂的, 真菌群落β多样性整体的差异显著(P=0.02 < 0.05).

|
图 6 不同退化阶段高寒草原微生物群落β多样性差异性分析 Fig. 6 Analysis of β diversity of microbial communities in alpine steppe at different degradation stages |
为了进一步探究高寒草原不同退化程度土壤微生物物种的共存模式以及随着退化程度增加网络拓扑特征的变化规律, 构建了微生物共现网络(图 7). 随着退化程度的增加, 土壤细菌共现网络的节点数和聚类系数显著下降(P < 0.05), 而细菌网络的模块性没有显著变化(图 8). 土壤真菌共现网络的节点数和聚类系数随着退化显著下降(P < 0.05), 而网络模块性随着退化呈现上升的趋势(P < 0.05), 表现出了明显的模块性结构(图 8). 这表明随着退化的发生, 细菌和真菌共现网络的相互作用减弱, 网络结构更加分散, 趋向于均质化, 网络稳定性下降. 同时, 随着高寒草原退化程度的增加, 不论是细菌还是真菌, 其共现网络的正相关连接占比均大于90%, 表明高寒草原退化过程中土壤微生物相互合作的关系并未改变. 此外, 在所有细菌共现网络相互作用中, 大部分核心节点对应的是变形菌门、绿弯菌门以及放线菌门, 在所有真菌共现网络相互作用中, 大部分核心节点对应的是子囊菌门, 而它们在高寒草原微生物群落的相对丰度也是所有检测到的门水平上最高的. 因此, 变形菌门、绿弯菌门以及放线菌门作为高寒草原最重要的关键细菌类群, 子囊菌门作为高寒草原最重要的关键真菌类群, 对维持微生物群落的稳定起着至关重要的作用.

|
图 7 高寒草原不同退化阶段土壤微生物共现网络 Fig. 7 Soil microbial co-occurrence networks in alpine steppe under different degradation levels |

|
(a)~(c)表示细菌网络拓扑特征变化, (d)~(f)表示真菌网络拓扑特征变化;*表示P < 0.05, **表示P < 0.01, ns表示P > 0.05 图 8 高寒草原不同退化阶段网络拓扑特征差异分析 Fig. 8 Difference analysis of network topological characteristics in different degradation stages of alpine steppe |
植被特征和土壤理化性质与土壤微生物网络拓扑特征的相关性分析结果表明, 细菌网络拓扑特征(节点数、聚类系数和模块性)主要与植被高度、总生物量、丰富度指数、香农-威纳指数、辛普森指数和全氮含量显著相关[P < 0.01, 图 9(a)], 真菌网络拓扑特征主要与植被盖度、总生物量、禾本科重要值、土壤容重、土壤砂粒含量显著相关[P < 0.05, 图 9(b)].

|
(a)细菌, (b)真菌;a1.节点数, a2.聚类系数, a3.模块性, a4.植被盖度, a5.植被高度, a6.丰富度指数, a7.香农-威纳指数, a8.辛普森指数, a9.禾本科重要值, a10.总生物量, a11.pH, a12.有机质, a13.全氮, a14.土壤容重, a15.全磷, a16.全钾, a17粉粒含量, a18.砂粒含量;b1.节点数, b2.聚类系数, b3.模块性, b4.植被盖度, b5.植被高度, b6.丰富度指数, b7.香农-威纳指数, b8.辛普森指数, b9.禾本科重要值, b10.总生物量, b11.pH, b12.有机质, b13.全氮, b14.土壤容重, b15.全磷, b16.全钾, b17粉粒含量, b18.砂粒含量;*表示P < 0.05, **表示P < 0.01, ns表示P ≥0.05;蓝色表示正相关关系, 红色表示负相关关系, 椭圆颜色越深及越窄表示相关程度越高 图 9 共现网络拓扑特征与环境因子相关性分析 Fig. 9 Correlation analysis between topological features in the co-occurrence network and environmental factors |
为深入探究高寒草原退化过程中植被特征和土壤理化性质对微生物群落的调节作用, 本文基于相似或相异度矩阵的多元回归分析(MRM)来探究植被特征和土壤理化性质对微生物群落影响的相对贡献. 结果表明:高寒草原退化过程中植被特征和土壤理化性质对土壤细菌群落变化的解释率为53.6%, 其中植被和土壤理化性质的交互作用解释率最大, 可解释总体的23.1%;对土壤真菌群落变化的解释率为27.1%, 土壤理化性质的解释率最大, 可解释总体的13.9%, 可见土壤细菌主要受植被-土壤复合的影响更大, 而真菌主要是受土壤理化性质的影响[图 10(b)].

|
(a)和(b)别表示植被特征和土壤理化性质对土壤细菌和真菌影响的MRM分析;(c)和(d)分别表示植被特征和土壤理化性质与土壤细菌和真菌的冗余分析 图 10 高寒草原退化过程中植被特征和土壤理化性质对土壤微生物群落的影响 Fig. 10 Effects of vegetation characteristics and soil physicochemical properties on soil microbial communities during the degradation of alpine steppe |
进一步通过冗余分析来探究影响微生物群落结构的环境因子, 结果表明, 对细菌影响显著的植被因子为植被高度、辛普森指数、禾本科重要值和总生物量(P < 0.05), 土壤理化性质是土壤容重、全磷含量、全钾含量和土壤粉粒含量[P < 0.05, 图 10(c)];对土壤真菌影响显著的植被因子为植被高度和总生物量(P < 0.05), 土壤理化性质影响显著的是土壤pH、全氮含量和土壤容重[P < 0.05, 图 10(d)].
3 讨论 3.1 高寒草原退化过程中土壤微生物特征发生的显著性变化与高寒草甸退化一致, 高寒草原退化导致了地上植被特征(盖度、高度、地上生物量和丰富度指数)及土壤理化性质(养分、质地及容重)的显著改变, 但与高寒草甸退化不一致的是, 高寒草原并未导致土壤含水量的显著改变[40, 41]. 与以往研究结果相似, 随着高寒草原退化强度的增加, 微生物组成和结构发生了显著的变化[6, 42]. 由于过度放牧等人类活动的干扰, 高寒草原地上生物量大量减少, 这会直接导致返回到土壤基底的营养物质急剧下降, 并引起一系列的正反馈, 最终导致土壤微生物基质营养物质的可利用性降低, 导致土壤微生物组成和结构发生改变及多样性损失[43, 44]. 尽管存在相对丰度的差异, 本研究发现了与以往研究相似的优势菌门[42]. 在门水平上, 放线菌门、变形菌门和酸杆菌门为高寒草原优势细菌, 放线菌在低温下具有很强的代谢能力和DNA修复机制[42], 变形菌可能在系统发育、生态和致病价值中起关键作用, 并参与能量代谢, 例如有机和无机化合物的氧化以及从光中获得能量[45]. 酸杆菌在有机质分解和养分循环中起着重要作用[46, 47]. 在退化过程中, 变形菌门的相对丰度减小[图 3(c)], 变形菌门在未退化阶段相对丰度最高, 可能与未退化草原有机碳含量较高有关, 有研究表明, 变形菌门与土壤有机碳呈显著正相关关系, 相对丰度随土壤有机碳的升高而增加[48, 49]. 而其它优势门在不同退化程度没有差异, 表明放线菌、酸杆菌对高寒草原生境条件(如土壤条件和植被变化)的适应性. 真菌的菌门中, 子囊菌门在各退化中均占最大比例, 是高寒草原真菌绝对优势菌门, 主要是因为子囊菌门是营腐生生活的真菌, 在将植物残体等有机质分解为可被植物直接吸收的简单无机化合物中起重要作用[50]. 本研究发现, Rhizobiales、Vicinamibacterales和Solirubrobacterales是目水平上的优势细菌分类群, Pleosporales、Hypocreales和Mortierellales(被孢)是目水平上的优势真菌分类群. Rhizobiales目主要通过利用铵盐和硝酸盐等, 或者与植被根系形成根瘤固定大气中的氮元素等方式, 对土壤中或者大气中氮素的分布产生很大影响[51, 52]. 草地退化引起的Rhizobiales目相对丰度减少, 可能是导致高寒草原土壤氮素流失的关键过程[51], 而子囊菌门的Pleosporales目在土壤中起着分解和降解有机物质的重要作用[50].
本研究发现高寒草原土壤细菌和土壤真菌的Chao1指数和Richness指数随着退化程度的增强表现为显著下降的趋势, 而微生物Shannon指数并未表现出显著的变化特征. 微生物Shannon指数包含两方面的信息:物种丰富度和物种分布的均匀度, 即物种丰富度越高且均匀分布, 则微生物Shannon指数就越高[53]. 本文结果表明, 随着退化程度的增加, 土壤微生物的丰富度(Chao1指数和Richness指数)出现显著下降的趋势, 而微生物Shannon指数没有显著变化, 这表明随着退化程度的增加, 细菌和真菌的均质化程度在增加. 高寒草甸退化过程中也发现了同样的现象, 不同的是高寒草甸退化过程中土壤微生物的均质化要更加明显[41]. 值得注意的是, 尽管高寒草原退化过程中不同阶段细菌和真菌丰富度指数均表现出显著的差异, 但变化趋势表现不同, 细菌丰富度指数随着退化程度的增加表现为先增加后降低的变化趋势, 而真菌丰富度指数表现为逐级下降的趋势(图 4), 这可能与退化过程中环境异质性及微生物不同的生存策略有关[54, 55]. PCoA分析结果显示, 土壤细菌、真菌群落结构在整个退化高寒草原表现出显著差异, 有研究发现细菌群落结构随着草地退化程度的增加而明显不同, 并认为这些差异是由于过度放牧等人类活动促进了草地退化, 植物生长、土壤结构和养分状况受到物理伤害引起的不同退化草地之间土壤性质和植被特征的差异所致[56]. 细菌在分解有机物、氮循环和土壤养分转化等方面具有更高的活性, 对环境变化敏感, 因此更容易受到草地退化的影响, 而真菌更倾向于形成菌丝网络并与植物根系形成共生关系, 这种共生关系相对稳定, 使得真菌群落在退化情况下相对不易改变[27].
土壤微生物群落的共现模式有助于从新的角度认识退化过程中微生物的相互作用[57], 这对于理解环境改变后微生物多样性和群落稳定性具有重要意义[58]. 在本研究中, 高寒草原退化的发生在很大程度上减弱了土壤细菌和真菌之间的相互作用(图 7), 同时结合网络拓扑特征的变化发现, 节点数和聚类系数都随着退化显著下降, 表明随着退化的发生, 微生物群落共现网络的聚集性下降, 相互作用减弱, 导致了土壤微生物更加分散和均质的生存策略, 而更加分散和均质的生存策略间接使得细菌与真菌共现网络的稳定性下降[28, 59]. 但在整个退化过程中, 土壤细菌与土壤真菌之间的相互作用方式并没有发生改变, 正相关连接占比均为90%以上, 都是以合作关系为主(图 7).
3.2 高寒草原退化过程中植被特征和土壤理化性质对微生物群落的调节通过对网络拓扑特征与植被和土壤理化性质的相关性分析显示, 土壤细菌网络拓扑特征与辛普森指数、土壤养分以及质地显著相关(P < 0.05), 土壤真菌网络拓扑特征与植被盖度、总生物量、土壤养分及质地显著相关(P < 0.05). 地上植被盖度会影响根际微生物与植物根系之间的相互作用, 植物通过根系分泌物与土壤微生物进行交流和相互作用, 地上植被盖度会影响根际交流的强弱, 而根际微生物与植物根系存在着复杂的相互作用关系, 包括共生、拮抗和协同作用等[60 ~ 62]. 这些相互作用可以改变土壤微生物的群落结构和多样性, 进而影响微生物网络的拓扑特征和功能, 而地上生物量直接或间接影响着土壤中的有机质输入和分解速率, 土壤中碳源以及营养物质的变化可能会改变细菌之间的相互作用[63], 较高的地上植被总生物量通常与更丰富的根系分泌物相关联, 这可能促进土壤微生物的繁殖和活性, 进而影响微生物之间的相互作用[25]. 土壤质地的变化会影响微生物的空间分布和运动性, 进而影响它们之间的相互作用[64]. 例如, 粒径较大的砂质土壤可能提供更多的微生物栖息空间, 促进微生物之间的物理接触和相互作用[65]. 微生物可以改变生存策略以适应环境变化, 并可以在营养缺乏的环境中与其它微生物物种合作[66]. 在本研究中, 随着高寒草原退化程度的增加, 土壤细菌与土壤真菌之间的相互作用方式并没有发生改变, 都是以合作关系为主, 表明土壤微生物群落可以相互合作以适应环境的变化.
以往研究表明, 由于植物生长对土壤结构及土壤养分状况的影响, 微生物的组成及结构也会随着植被类型的变化而改变[67]. RDA分析结果显示, 土壤细菌主要受植被特征(植被高度、辛普森指数、禾本科重要值和总生物量)和土壤理化性质(土壤容重、全磷含量、全钾含量和土壤粉粒含量)的影响;CCA分析结果显示, 土壤真菌主要受植被特征(植被高度和总生物量)和土壤理化性质(土壤pH、全氮含量和土壤容重)的影响, 地上植被特征(植被高度、总生物量和辛普森指数)显著影响了土壤细菌和真菌群落的组成和结构, 这是因为地上植被的消失会限制一些土壤微生物群落的生长和种群, 由于减少的凋落物输入到土壤中引起微生物底物可用性降低[44];土壤容重会影响土壤的孔隙度和通气性, 而微生物需要氧气进行呼吸和代谢活动, 可能会限制某些需氧微生物的生长[68]. 禾本科重要值和粉粒含量显著影响了细菌群落的组成和结构, 可能是因为禾本科植物分泌的有机物质可以作为微生物的碳源, 促进细菌的生长和代谢活动, 例如, 有机酸可以作为细菌的碳源, 并促进细菌对其他有机化合物的降解[25, 69], 而土壤粉粒含量会影响土壤表面的粗糙度, 使得用于吸附有机物质和微生物细胞的结合位点发生改变并影响用于定殖的土壤细菌微生境[70];而土壤全磷和全钾是植物生长发育的限制因素, 进而影响了土壤细菌生长, 而它们的含量在高寒草原土壤中非常有限, 从而间接影响了细菌群落的组成与结构[6]. 本研究发现土壤pH值和全氮含量显著影响了真菌群落, 土壤pH值已被广泛认为是全球微生物多样性和丰富性的最佳预测因子[67], 有研究表示高寒草甸真菌群落退化过程中真菌群落结构受到多种环境变量的影响, 包括土壤pH值[40, 71]. 土壤中的氮也是微生物生长和代谢所必需的营养元素, 土壤中的氮含量可以影响微生物的生理活动、代谢途径和群落结构[72]. 本研究中, 地上植被因子对土壤细菌群落结构变化的解释率为10.7%, 而土壤因子和植被-土壤交互作用对细菌群落结构变化的解释率分别为19.8%和23.1%, 土壤细菌受植被-土壤耦合的影响更大, 在对高寒草甸退化对土壤细菌群落的影响的研究中也得出相似结论[73]. 而植被特征对真菌群落变化的解释率仅为5.3%, 土壤理化性质单独解释率为13.9%, 说明真菌主要受土壤理化性质的影响, 可能原因是高寒草原退化过程中地上植被稀疏, 土壤含水量较少, 植物根系的生长和活动也会受到限制, 从而导致真菌无法获得足够的水分和碳源来维持其共生关系[74], 这与赵文等的研究结果一致[75].
4 结论(1)高寒草原退化显著改变了地上植被特征与土壤理化性质, 使得植被盖度、高度、总生物量和禾本科重要值降低, 土壤养分含量如土壤有机质、全氮含量、全磷和全钾都随着退化显著下降, 此外土壤质地也发生了改变, 土壤容重增大, 土壤粉粒含量下降, 砂粒含量上升, 土壤沙化严重.
(2)高寒草原退化未改变土壤微生物的组成, 但改变了不同微生物的组成比例和微生物多样性, 土壤微生物α多样性(Richness、Chao1、ACE和Sobs指数)显著下降, 但细菌和真菌的响应存在差异, 细菌α多样性随着退化均表现为先上升后下降的趋势, 而真菌α多样性随着退化都呈现显著下降的趋势. 但微生物Shannon指数在不同退化程度高寒草原没有显著差异, 表明高寒草原退化导致了微生物分布的均匀度上升, 结合微生物群落的β多样性, 退化显著改变了微生物群落的结构.
(3)结合网络拓扑特征的变化发现, 高寒草原退化过程中土壤细菌和真菌的种间相互作用减弱, 网络结构更加分散, 更趋向于平均化, 共现网络的节点数和聚类系数显著下降, 稳定性下降, 但土壤微生物相互合作的关系并未改变. 植被高度、总生物量、土壤容重和土壤质地是影响土壤微生物相互作用的主要因素.
(4)高寒草原退化过程中, 植被特征(植被高度、辛普森指数、禾本科重要值、总生物量)和土壤理化性质(pH值、全氮含量、土壤容重、全磷含量和全钾含量)是影响土壤微生物群落的主要驱动因子, 其中土壤细菌主要受植被-土壤耦合的影响更大, 而真菌主要受土壤理化性质的影响更大.
| [1] |
董世魁, 汤琳, 张相锋, 等. 高寒草地植物物种多样性与功能多样性的关系[J]. 生态学报, 2017, 37(5): 1472-1483. Dong S K, Tang L, Zhang X F, et al. Relationship between plant species diversity and functional diversity in alpine grasslands[J]. Acta Ecologica Sinica, 2017, 37(5): 1472-1483. |
| [2] |
张中华, 周华坤, 赵新全, 等. 青藏高原高寒草地生物多样性与生态系统功能的关系[J]. 生物多样性, 2018, 26(2): 111-129. Zhang Z H, Zhou H K, Zhao X Q, et al. Relationship between biodiversity and ecosystem functioning in alpine meadows of the Qinghai-Tibet Plateau[J]. Biodiversity Science, 2018, 26(2): 111-129. |
| [3] |
罗正明, 刘晋仙, 胡砚秋, 等. 五台山不同退化程度亚高山草甸土壤微生物群落分类与功能多样性特征[J]. 环境科学, 2023, 44(5): 2918-2927. Luo Z M, Liu J X, Hu Y Q, et al. Taxonomic and functional diversity of soil microbial communities in subalpine meadow with different degradation degrees in Mount Wutai[J]. Environmental Science, 2023, 44(5): 2918-2927. |
| [4] | Xu Y D, Dong S K, Gao X X, et al. Aboveground community composition and soil moisture play determining roles in restoring ecosystem multifunctionality of alpine steppe on Qinghai-Tibetan Plateau[J]. Agriculture, 2021, 305. DOI:10.1016/j.agee.2020.107163 |
| [5] |
陈宁, 张扬建, 朱军涛, 等. 高寒草甸退化过程中群落生产力和物种多样性的非线性响应机制研究[J]. 植物生态学报, 2018, 42(1): 50-65. Chen N, Zhang Y J, Zhu J T, et al. Nonlinear responses of productivity and diversity of alpine meadow communities to degradation[J]. Chinese Journal of Plant Ecology, 2018, 42(1): 50-65. |
| [6] | Zhou H, Zhang D G, Jiang Z H, et al. Changes in the soil microbial communities of alpine steppe at Qinghai-Tibetan Plateau under different degradation levels[J]. Science of the Total Environment, 2019, 651: 2281-2291. DOI:10.1016/j.scitotenv.2018.09.336 |
| [7] |
孙鸿烈, 郑度, 姚檀栋, 等. 青藏高原国家生态安全屏障保护与建设[J]. 地理学报, 2012, 67(1): 3-12. Sun H L, Zheng D, Yao T D, et al. Protection and construction of the national ecological security shelter zone on Tibetan Plateau[J]. Acta Geographica Sinica, 2012, 67(1): 3-12. |
| [8] |
璩伟卿, 张博美, 黄雪, 等. 基于16S rRNA测序技术的青藏高原河流细菌群落多样性[J]. 环境科学, 2023, 44(1): 262-271. Qu W Q, Zhang B M, Huang X, et al. Bacterial community and diversity of river ecosystems on the Qinghai-Tibet Plateau based on 16S rRNA gene sequencing[J]. Environmental Science, 2023, 44(1): 262-271. |
| [9] | Wang C T, Wang G X, Wang Y, et al. Fire alters vegetation and soil microbial community in alpine meadow[J]. Land Degradation & Development, 2016, 27(5): 1379-1390. |
| [10] | Fierer N, Leff J W, Adams B J, et al. Cross-biome metagenomic analyses of soil microbial communities and their functional attributes[J]. Proceedings of the National Academy of Sciences of the United States of America, 2012, 109(52): 21390-21395. |
| [11] |
刘育红, 杨元武, 张英. 退化高寒草甸植物功能群与土壤因子关系的冗余分析[J]. 生态与农村环境学报, 2018, 34(12): 1112-1121. Liu Y H, Yang Y W, Zhang Y. Redundancy analysis of the relationship between plant functional groups and soil factors in the degraded alpine meadow[J]. Journal of Ecology and Rural Environment, 2018, 34(12): 1112-1121. |
| [12] |
李成阳, 赖炽敏, 彭飞, 等. 青藏高原北麓河流域不同退化程度高寒草甸生产力和群落结构特征[J]. 草业科学, 2019, 36(4): 1044-1052. Li C Y, Lai Z M, Peng F, et al. Alpine meadows at different stages of degradation in the Beiluhe basin of the Qinghai-Tibet Plateau: productivity and community structure characteristics[J]. Pratacultural Science, 2019, 36(4): 1044-1052. |
| [13] |
刘玉, 马玉寿, 施建军, 等. 大通河上游高寒草甸植物群落的退化特征[J]. 草业科学, 2013, 30(7): 1082-1088. Liu Y, Ma Y S, Shi J J, et al. Degradation characteristics of alpine meadow plant communities in the upper reaches of Datong River[J]. Pratacultural Science, 2013, 30(7): 1082-1088. |
| [14] |
温军, 周华坤, 陈哲, 等. 不同退化程度高寒草甸主要植物的热值研究[J]. 草业科学, 2012, 29(9): 1451-1456. Wen J, Zhou H K, Chen Z, et al. Study on caloril values of plants in alpine meadow with different degradation degrees[J]. Pratacultural Science, 2012, 29(9): 1451-1456. |
| [15] | Wang C W, Liu Z K, Yu W Y, et al. Grassland degradation has stronger effects on soil fungal community than bacterial community across the Semi-Arid region of northern China[J]. Plants, 2022, 11(24). DOI:10.3390/plants11243488 |
| [16] |
高小源, 鲁旭阳. 休牧对西藏高寒草原和高寒草甸植被与土壤特征的影响[J]. 草业科学, 2020, 37(3): 486-496. Gao X Y, Lu X Y. Effects of rest grazing on vegetation and soil characteristics of alpine steppe and alpine meadow in Tibet[J]. Pratacultural Science, 2020, 37(3): 486-496. |
| [17] |
杨飞. 放牧、增雨和增食对内蒙古草原植被群落结构及部分土壤理化性质的影响[D] 扬州: 扬州大学, 2017. Yang F. Effects of gazing, precipitation and adding food on vegetation community and partial soil physic-chemical property in inner Mongolia grassland[D]. Yangzhou: Yangzhou University, 2017. |
| [18] |
杜有新. 退化喀斯特生态系统植被群落特征及氮磷养分有效性——以贵州省普定县为例[D]. 南京: 南京农业大学, 2011. Du Y X. Characteristics of vegetation communities and nitrogen and phosphorus availability in degraded ecosystems in Karst region in Guizhou, China-A case study with different ecosystems under vegetation decline from Puding, Guizhou[D]. Nanjing: Nanjing Agricultural University, 2011. |
| [19] |
段敏杰, 高清竹, 万运帆, 等. 放牧对藏北紫花针茅高寒草原植物群落特征的影响[J]. 生态学报, 2010, 30(14): 3892-3900. Duan M J, Gao Q Z, Wan Y F, et al. Effect of grazing on community characteristics and species diversity of Stipa purpurea alpine grassland in Northern Tibet[J]. Acta Ecologica Sinica, 2010, 30(14): 3892-3900. |
| [20] |
王长庭, 龙瑞军, 王根绪, 等. 高寒草甸群落地表植被特征与土壤理化性状、土壤微生物之间的相关性研究[J]. 草业学报, 2010, 19(6): 25-34. Wang C T, Long R J, Wang G X, et al. Relationship between plant communities, characters, soil physical and chemical properties, and soil microbiology in alpine meadows[J]. Acta Prataculturae Sinica, 2010, 19(6): 25-34. |
| [21] |
李建宏, 李雪萍, 卢虎, 等. 高寒地区不同退化草地植被特性和土壤固氮菌群特性及其相关性[J]. 生态学报, 2017, 37(11): 3647-3654. Li J H, Li X P, Lu H, et al. Characteristics of, and the correlation between, vegetation and N-fixing soil bacteria in alpine grassland showing various degrees of degradation[J]. Acta Ecologica Sinica, 2017, 37(11): 3647-3654. |
| [22] |
肖玉. 青藏高原高寒草原不同退化程度植物群落特征与土壤养分的关系[D]. 兰州: 兰州大学, 2016. Xiao Y. The relationship between plant community and soil nutrients features of alpine steppe at different degradation degrees in Qinghai-Tibetan Plateau[D]. Lanzhou: Lanzhou University, 2016. |
| [23] | Freilich S, Kreimer A, Meilijson I, et al. The large-scale organization of the bacterial network of ecological co-occurrence interactions[J]. Nucleic Acids Research, 2010, 38(12): 3857-3868. |
| [24] | Yuan M M, Guo X, Wu L W, et al. Climate warming enhances microbial network complexity and stability[J]. Nature Climate Change, 2021, 11(4): 343-348. |
| [25] |
吴林坤, 林向民, 林文雄. 根系分泌物介导下植物-土壤-微生物互作关系研究进展与展望[J]. 植物生态学报, 2014, 38(3): 298-310. Wu L K, Lin X M, Lin W X. Advances and perspective in research on plant-soil-microbe interactions mediated by root exudates[J]. Chinese Journal of Plant Ecology, 2014, 38(3): 298-310. |
| [26] | Wagg C, Bender S F, Widmer F, et al. Soil biodiversity and soil community composition determine ecosystem multifunctionality[J]. Proceedings of the National Academy of Sciences of the United States of America, 2014, 111(14): 5266-5270. |
| [27] | Bardgett R D, van der Putten W H. Belowground biodiversity and ecosystem functioning[J]. Nature, 2014, 515(7528): 505-511. |
| [28] | Coyte K Z, Schluter J, Foster K R. The ecology of the microbiome: networks, competition, and stability[J]. Science, 2015, 350(6261): 663-666. |
| [29] |
胡雷, 王长庭, 王根绪, 等. 三江源区不同退化演替阶段高寒草甸土壤酶活性和微生物群落结构的变化[J]. 草业学报, 2014, 23(3): 8-19. Hu L, Wang C T, Wang G X, et al. Changes in the activities of soil enzymes and microbial community structure at different degradation successional stages of alpine meadows in the headwater region of Three Rivers, China[J]. Acta Prataculturae Sinica, 2014, 23(3): 8-19. |
| [30] | Xu X D, Lu C G, Shi X H, et al. World water tower: an atmospheric perspective[J]. Geophysical Research Letters, 2008, 35(20). DOI:10.1029/2008GL035867 |
| [31] | Zhou Z W, Liu Y, Zhu Q, et al. Comparing the variations and controlling factors of soil N2O emissions and NO3--N leaching on tea and bamboo hillslopes[J]. CATENA, 2020, 188. DOI:10.1016/j.catena.2020.104463 |
| [32] |
马玉寿, 郎百宁, 李青云, 等. 江河源区高寒草甸退化草地恢复与重建技术研究[J]. 草业科学, 2002, 19(9): 1-5. Ma Y S, Lang B N, Li Q Y, et al. Research on restoration and reconstruction techniques for degraded alpine meadow in the source region of rivers[J]. Pratacultural Science, 2002, 19(9): 1-5. |
| [33] | GB 19377-2003, 天然草地退化、沙化、盐渍化的分级指标[S]. |
| [34] | 鲍士旦. 土壤农化分析[M]. 北京: 中国农业出版社, 2000, (第三版). |
| [35] | Fu B J, Liu S L, Chen L D, et al. Soil quality regime in relation to land cover and slope position across a highly modified slope landscape[J]. Ecological Research, 2004, 19(1): 111-118. |
| [36] | Tian K, Huang B, Xing Z, et al. Geochemical baseline establishment and ecological risk evaluation of heavy metals in greenhouse soils from Dongtai, China[J]. Ecological Indicators, 2017, 72: 510-520. |
| [37] |
何兴东, 高玉葆, 刘惠芬. 重要值的改进及其在羊草群落分类中的应用[J]. 植物研究, 2004, 24(4): 466-472. He X D, Gao Y B, Liu H F. Amending of importance value and its application on classification of Leymus chinensis communities[J]. Bulletin of Botanical Research, 2004, 24(4): 466-472. |
| [38] |
马克平, 黄建辉, 于顺利, 等. 北京东灵山地区植物群落多样性的研究Ⅱ丰富度、均匀度和物种多样性指数[J]. 生态学报, 1995, 15(3): 268-277. Ma K P, Huang J H, Yu S L, et al. Plant community diversity in Dongling mountain, Beijing, China: Ⅱ. Species richness, evenness and species diversities[J]. Acta Ecologica Sinica, 1995, 15(3): 268-277. |
| [39] | Chu Z R, Wang K, Li X K, et al. Microbial characterization of aggregates within a one-stage nitritation–anammox system using high-throughput amplicon sequencing[J]. Chemical Engineering Journal, 2015, 262: 41-48. |
| [40] | Li Y M, Wang S P, Jiang L L, et al. Changes of soil microbial community under different degraded gradients of alpine meadow[J]. Agriculture, 2016, 222: 213-222. |
| [41] | Zhang Y J, Ding M J, Zhang H, et al. Degradation-driven bacterial homogenization closely associated with the loss of soil multifunctionality in alpine meadows[J]. Agriculture, 2023, 344. DOI:10.1016/j.agee.2022.108284 |
| [42] | Luo S, Png G K, Ostle N J, et al. Grassland degradation-induced declines in soil fungal complexity reduce fungal community stability and ecosystem multifunctionality[J]. Soil Biology and Biochemistry, 2023, 176. DOI:10.1016/j.soilbio.2022.108865 |
| [43] | Wu G L, Ren G H, Dong Q M, et al. Above- and belowground response along degradation gradient in an alpine grassland of the Qinghai- Tibetan Plateau[J]. CLEAN-Soil Air Water, 2014, 42(3): 319-323. |
| [44] | Zhang Y, Cao C Y, Peng M, et al. Diversity of nitrogen-fixing, ammonia-oxidizing, and denitrifying bacteria in biological soil crusts of a revegetation area in Horqin Sandy Land, Northeast China[J]. Ecological Engineering, 2014, 71: 71-79. |
| [45] | Mukhopadhya I, Hansen R, El-Omar E M, et al. IBD-what role do Proteobacteria play?[J]. Nature Reviews Gastroenterology & Hepatology, 2012, 9(4): 219-230. |
| [46] | Fang D X, Zhao G, Xu X Y, et al. Microbial community structures and functions of wastewater treatment systems in plateau and cold regions[J]. Bioresource Technology, 2018, 249: 684-693. |
| [47] |
高日平, 段玉, 张君, 等. 长期施肥对农牧交错带旱地土壤微生物多样性及群落结构的影响[J]. 环境科学, 2023, 44(2): 1063-1073. Gao R P, Duan Y, Zhang J, et al. Effects of long-term fertilization on soil microbial diversity and community structure in the Agro-pastoral ecotone[J]. Environmental Science, 2023, 44(2): 1063-1073. |
| [48] | Navarrete I A, Tsutsuki K. Land-use impact on soil carbon, nitrogen, neutral sugar composition and related chemical properties in a degraded Ultisol in Leyte, Philippines[J]. Soil Science and Plant Nutrition, 2008, 54(3): 321-331. |
| [49] | Fierer N, Bradford M A, Jackson R B. Toward an ecological classification of soil bacteria[J]. Ecology, 2007, 88(6): 1354-1364. |
| [50] |
刘金艳, 周攀, 周小路, 等. 子囊菌Sordaria tomento⁃alba的筛选及其木质素降解特性[J]. 应用与环境生物学报, 2022, 28(3): 693-698. Liu J Y, Zhou P, Zhou X L, et al. Screening of ascomycete Sordaria tomento⁃alba and its lignin degradation characteristics[J]. Chinese Journal of Applied and Environmental Biology, 2022, 28(3): 693-698. |
| [51] |
孙华方, 李希来, 金立群, 等. 黄河源区斑块化退化高寒草甸土壤细菌群落多样性变化[J]. 环境科学, 2022, 43(9): 4662-4673. Sun H F, Li X L, Jin L Q, et al. Changes in soil bacterial community diversity in degraded patches of alpine meadow in the source area of the Yellow River[J]. Environmental Science, 2022, 43(9): 4662-4673. |
| [52] | Sprent J I, Ardley J, James E K. Biogeography of nodulated legumes and their nitrogen‐fixing symbionts[J]. New Phytologist, 2017, 215(1): 40-56. |
| [53] | Jing X, Sanders N J, Shi Y, et al. The links between ecosystem multifunctionality and above- and belowground biodiversity are mediated by climate[J]. Nature Communications, 2015, 6(1). DOI:10.1038/ncomms9159 |
| [54] | Delgado-Baquerizo M, Eldridge D J. Cross-biome drivers of soil bacterial alpha diversity on a worldwide scale[J]. Ecosystems, 2019, 22(6): 1220-1231. |
| [55] | Li J, Delgado-Baquerizo M, Wang J T, et al. Fungal richness contributes to multifunctionality in boreal forest soil[J]. Soil Biology and Biochemistry, 2019, 136. DOI:10.1016/j.soilbio.2019.107526 |
| [56] | Luo Z M, Liu J X, Jia T, et al. Soil bacterial community response and nitrogen cycling variations associated with subalpine meadow degradation on the Loess Plateau, China[J]. Applied and Environmental Microbiology, 2020, 86(9). DOI:10.1128/AEM.00180-20 |
| [57] | Li Y, Sun L L, Sun M L, et al. Vertical and horizontal biogeographic patterns and major factors affecting bacterial communities in the open South China Sea[J]. Scientific Reports, 2018, 8(1). DOI:10.1038/s41598-018-27191-w |
| [58] | Zhang Z J, Deng Y, Feng K, et al. Deterministic assembly and diversity gradient altered the biofilm community performances of bioreactors[J]. Environmental Science & Technology, 2019, 53(3): 1315-1324. |
| [59] | Chen B B, Jiao S, Luo S W, et al. High soil pH enhances the network interactions among bacterial and archaeal microbiota in alpine grasslands of the Tibetan Plateau[J]. Environmental Microbiology, 2021, 23(1): 464-477. |
| [60] |
樊丹丹, 刘艳娇, 曹慧丽, 等. 围栏工程对退化草地土壤理化性质和微生物群落的影响[J]. 科技导报, 2022, 40(3): 41-51. Fan D D, Liu Y J, Cao H L, et al. The impact of enclosure engineering on the soil physicochemical properties and microbial community of degraded grassland[J]. Science & Technology Review, 2022, 40(3): 41-51. |
| [61] | Xiong Q Q, Hu J L, Wei H Y, et al. Relationship between plant roots, rhizosphere microorganisms, and nitrogen and its special focus on rice: agriculture[J]. Agriculture, 2021, 11(3). DOI:10.3390/agriculture11030234 |
| [62] | Philippot L, Raaijmakers J M, Lemanceau P, et al. Going back to the roots: the microbial ecology of the rhizosphere[J]. Nature Reviews Microbiology, 2013, 11(11): 789-799. |
| [63] |
周莉, 李保国, 周广胜. 土壤有机碳的主导影响因子及其研究进展[J]. 地球科学进展, 2005, 20(1): 99-105. Zhou L, Li B G, Zhou G S. Advances in controlling factors of soil organic carbon[J]. Advances in Earth Science, 2005, 20(1): 99-105. |
| [64] |
侯芳彬, 王蕊, Ali S, 等. 黄土区沟道泥沙微生物群落变化特征及其影响因素[J]. 中国环境科学, 2019, 39(10): 4350-4359. Hou F B, Wang R, Ali S, et al. Variations of soil microbial communities along a valley bottom of the loess plateau and the influencing factors[J]. China Environmental Science, 2019, 39(10): 4350-4359. |
| [65] |
张力斌, 何明珠, 张克存, 等. 干旱风沙区植被重建初期对土壤微生物群落结构的影响[J]. 干旱区地理, 2022, 45(6): 1916-1926. Zhang L B, He M Z, Zhang K C, et al. Effect of preliminary vegetation reconstruction on soil microorganism community structure in arid desert area[J]. Arid Land Geography, 2022, 45(6): 1916-1926. |
| [66] | Zhang B G, Zhang J, Liu Y, et al. Co-occurrence patterns of soybean rhizosphere microbiome at a continental scale[J]. Soil Biology and Biochemistry, 2018, 118: 178-186. |
| [67] | Lauber C L, Hamady M, Knight R, et al. Pyrosequencing-based assessment of Soil pH as a predictor of soil bacterial community structure at the continental scale[J]. Applied and Environmental Microbiology, 2009, 75(15): 5111-5120. |
| [68] |
王芳芳, 徐欢, 李婷, 等. 放牧对草地土壤氮素循环关键过程的影响与机制研究进展[J]. 应用生态学报, 2019, 30(10): 3277-3284. Wang F F, Xu H, Li T, et al. Effects and mechanisms of grazing on key processes of soil nitrogen cycling in grassland: a review[J]. Chinese Journal of Applied Ecology, 2019, 30(10): 3277-3284. |
| [69] |
杨德春, 胡雷, 宋小艳, 等. 降雨变化对高寒草甸不同植物功能群凋落物质量及其分解的影响[J]. 植物生态学报, 2021, 45(12): 1314-1328. Yang D C, Hu L, Song X Y, et al. Effects of changing precipitation on litter quality and decomposition of different plant functional groups in an alpine meadow[J]. Chinese Journal of Plant Ecology, 2021, 45(12): 1314-1328. |
| [70] | Chau J F, Bagtzoglou A C, Willig M R. The effect of soil texture on richness and diversity of bacterial communities[J]. Environmental Forensics, 2011, 12(4): 333-341. |
| [71] |
杨鹏年, 李希来, 李成一, 等. 黄河源区斑块化退化高寒草甸土壤微生物多样性对长期封育的响应[J]. 环境科学, 2023, 44(4): 2293-2303. Yang P N, Li X L, Li C Y, et al. Response of soil microbial diversity to long-term enclosure in degraded patches of alpine meadow in the source zone of the Yellow River[J]. Environmental Science, 2023, 44(4): 2293-2303. |
| [72] |
杨雅丽, 马雪松, 解宏图, 等. 保护性耕作对土壤微生物群落及其介导的碳循环功能的影响[J]. 应用生态学报, 2021, 32(8): 2675-2684. Yang Y L, Ma X S, Xie H T, et al. Effects of conservation tillage on soil microbial community and the function of soil carbon cycling[J]. Chinese Journal of Applied Ecology, 2021, 32(8): 2675-2684. |
| [73] |
李世雄, 王彦龙, 王玉琴, 等. 土壤细菌群落特征对高寒草甸退化的响应[J]. 生物多样性, 2021, 29(1): 53-64. Li S X, Wang Y L, Wang Y Q, et al. Response of soil bacterial community characteristics to alpine meadow degradation[J]. Biodiversity Science, 2021, 29(1): 53-64. |
| [74] |
曹亚静, 赵美丞, 郑春燕, 等. 根际微生物介导的植物响应干旱胁迫机制研究进展[J]. 中国生态农业学报(中英文), 2023, 31(8): 1330-1342. Cao Y J, Zhao M C, Zheng C Y, et al. Rhizosphere microorganisms-mediated plant responses to drought stress[J]. Chinese Journal of Eco-Agriculture, 2023, 31(8): 1330-1342. |
| [75] |
赵文, 尹亚丽, 李世雄, 等. 三江源区退化高寒草甸土壤真菌群落特征[J]. 应用生态学报, 2021, 32(3): 869-877. Zhao W, Yin Y L, Li S X, et al. The characteristics of soil fungal community in degraded alpine meadow in the Three Rivers Source Region, China[J]. Chinese Journal of Applied Ecology, 2021, 32(3): 869-877. |