 2024, Vol. 45
2024, Vol. 45



巢湖地处长江中下游, 是我国五大淡水湖之一. 作为安徽省重要淡水湖泊, 巢湖具有调节径流、供水灌溉和维持生态系统多样性等多种功能, 在区域社会、经济发展以及自然生态系统保护中发挥着重要作用[1, 2]. 然而, 随着人类活动的加剧以及城市空间的扩张, 巢湖水体营养盐浓度不断攀升, 水体富营养化及由此引发的蓝藻水华已成为威胁人类健康、破坏水域生态系统平衡的重要因素, 是全世界淡水湖泊面临的主要环境问题之一[3, 4]. 自20世纪80年代巢湖蓝藻水华开始暴发以来, 至今湖泊水华问题依然严峻, 巢湖生态治理面临着巨大压力[5]. 因此, 综合运用多种技术手段开展对蓝藻水华的日常监测, 探究蓝藻水华的环境影响因素, 是认识蓝藻水华形成和发展规律的基础, 对于提升巢湖蓝藻水华预报预警能力、推动区域水资源保护及可持续利用具有重要意义.
针对蓝藻水华监测, 目前主要形成了基于站点的水质采样和基于卫星影像的遥感监测两种技术手段. 实地采样是蓝藻水华最常用且最直接的监测方法, 具有试验简捷和易操作实施等优点, 但该方法成本高和耗时长, 且监测结果仅代表局部水域的蓝藻水华特征, 无法反映长时序、大范围湖泊蓝藻水华的动态变化[6, 7]. 卫星遥测则为弥补这一缺陷提供了新的技术手段, 国内外学者针对湖泊蓝藻水华的识别与提取, 开发了指数阈值、监督分类以及基于机器学习的自动识别等提取算法[8, 9], 该方法可及时、准确地提供蓝藻水华暴发面积和时空分布等信息. 两种方法在蓝藻水华的动态监测以及长时序分析研究中均发挥了积极作用. 然而, 或受限于实地采样和遥感数据的可获取性, 已有研究较少对两种监测方式进行有效整合, 一定程度上阻碍了对巢湖蓝藻水华形成和发展规律的认知. 因此, 充分发挥不同观测手段的技术优势, 实现对巢湖蓝藻水华的立体和综合监测, 是现阶段学者们广泛关注的焦点问题[10, 11].
有关蓝藻水华的成因, 已有研究普遍认为, 在富营养化水体中, 营养盐浓度已不再是影响蓝藻生长的限制因子, 其他环境条件如气象要素可能是诱发蓝藻水华暴发的主要因子[12, 13]. 因此, 现有研究多聚焦气温[14]、降水[15]、风速风向[16]和日照时数[17]对蓝藻水华的影响, 偏向气象条件不同取值对蓝藻水华发生的定性描述, 而有关各因子的重要性度量以及交互作用对蓝藻水华的影响则较少涉及. 此外, 由于受到蓝藻本身生理特性、水体营养盐浓度以及气象条件等诸多环境因素的影响, 蓝藻水华形成机制较为复杂[8], 各因子对蓝藻水华的影响通常表现为非线性的变化模式, 继续沿用传统的统计方法定量分析不同阶段蓝藻水华成因存在一定局限. 机器学习和人工智能技术的兴起则为处理高维数据和非线性问题提供了新的解决方案[13, 18], 目前已被广泛应用于生态与环境等复杂问题的求解过程中. 基于机器学习算法在特征重要性评估和预测等方面的优势, 理论上可为评估巢湖蓝藻水华影响因子贡献率、理解环境因子综合作用对蓝藻水华生消机制的影响提供新的研究思路, 值得进行探寻和深入研究.
为此, 本文拟充分利用巢湖水上综合观测平台和卫星遥感等多源数据, 探索不同阶段巢湖蓝藻水华发生和发展的基本规律;在此基础上, 选取同期温、压、湿、风等多种气象要素, 并结合巢湖水环境监测数据, 尝试采用增强回归树的机器学习算法, 评估和分析各因子对巢湖蓝藻水华影响的重要性及相互作用关系, 识别诱发蓝藻水华暴发的主导驱动因子, 以期为巢湖蓝藻水华预测、预警和防治提供决策参考.
1 材料与方法 1.1 研究区概况巢湖地处长江、淮河两大水系中间, 是我国第五大淡水湖, 属合肥市内湖(图 1). 巢湖东西长54.5 km, 南北宽21 km, 水域面积约770 km2, 平均水深2.89 m, 以忠庙-姥山-齐头嘴为界分为东、西两大湖区[19]. 巢湖流域水系发达, 四周河渠广布, 共有大小33条河流通过地表径流补给汇入巢湖, 通过裕溪河与长江连通[20]. 巢湖属北亚热带温润性季风气候, 气候温和, 雨量适中, 年平均气温在15 ℃左右, 降水量可达1 000 mm[2]. 近年来, 随着城市化进程的不断推进以及巢湖航运、水产养殖等产业的快速发展, 巢湖水体富营养化日趋严重, 蓝藻水华频发, 给流域水生环境和人类生产、生活造成了严重威胁[5].

|
图 1 巢湖地理区位示意 Fig. 1 Geographic location of Chaohu Lake |
本研究主要使用3类数据开展巢湖蓝藻水华时空分布及驱动因素分析, 分别是卫星遥感数据、水上综合观测平台数据和气象数据. 卫星遥感数据来源于美国国家宇航局(National Aeronautics and Space Administration, NASA)提供的MODIS数据, 该数据具有波段信息丰富、时间分辨率高、数据易处理、免费开源等优点, 已被广泛应用于生态环境变化监测的研究中[22, 23]. 本文使用的MOD09GQ地表反射率产品, 涵盖可见光到近红外波段范围的多波段信息, 空间分辨率为250 m, 重访周期为1 d. 水上综合观测平台位于巢湖北岸南淝河入河口处(31.69°N, 117.40°E), 该平台配备有WIZ原位营养盐观测以及水质多参数监测仪等设备, 可实现对水体生物(藻密度、叶绿素a浓度)、水质(电导、盐度、水温、浊度)和化学(总氮、总磷、硝酸盐、磷酸盐)等要素的综合监测. 为了与水上综合观测平台数据相匹配, 气象数据采用距离观测平台最近的巢湖国家气象基本观测站, 包含温、压、湿、风和降雨等多个要素. 有关各数据集的观测指标及详细信息参见表 1.
|
|
表 1 本研究使用的数据简介 Table 1 Dataset used in this study |
1.3 水上综合观测平台数据质控
为充分利用巢湖水上综合观测平台数据, 提高数据质量, 本研究首先对监测数据进行质量控制. 具体包括:异常值检测和时间一致性分析两个步骤. 异常值检测即根据仪器提供的各参数监测范围, 对不在正常值区间内的记录予以剔除. 然后, 针对每一个观测要素进行时间一致性分析, 将偏离该要素均值(MEAN)3个标准差(SD)以外的记录判定为离群值. 此外, 当日数据缺测率超过20%以上的日期也不予考虑. 最后, 共筛选从2015年3月至2020年3月2 122条记录用于下文的分析. 考虑到水上综合观测平台位置特殊, 其观测结果可能受南淝河入湖河流的影响较大. 本研究根据Yang等[24]推荐的方法, 以研究期内观测值与平均值的差值偏离标准差的大小为判定依据, 将观测记录分为区域背景值和非背景值. 为增强观测数据的区域代表性, 本研究选取区域背景值作为输入数据用于下文蓝藻水华驱动因素建模.
1.4 巢湖蓝藻水华反演 1.4.1 蓝藻水华覆盖度计算鉴于水体表面悬浮藻类在近红外波段的高反射率以及水体在红光-近红外波段的强吸收特征, 基于可见光、近红外波段的卫星遥感资料为蓝藻监测提供了一种新方法[25]. 国内外学者针对从水体中提取藻类分布信息开展了大量研究, 同时也形成了较为成熟的湖泊蓝藻水华卫星遥感监测技术. 本研究依据中国气象局制定的《湖泊蓝藻水华卫星遥感监测技术导则》[26], 进行巢湖蓝藻水华信息的提取与分析. 该方法以归一化植被指数(NDVI)为输入参数, 通过计算水体蓝藻水华覆盖度指数(FCI), 实现像元尺度蓝藻水华的动态监测, 具体计算公式如下:
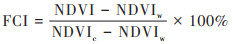
|
(1) |
式中, FCI为某一像元的蓝藻水华覆盖度, NDVIw和NDVIc分别为清洁水体和蓝藻水华的NDVI值, 根据《湖泊蓝藻水华卫星遥感监测技术导则》, 其值一般取-0.2和0.81. NDVI为可见光与近红外波段反射率差值与二者之和的比值, 计算公式如下:
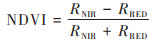
|
(2) |
式中, RNIR为近红外波段反射率, RRED为红光波段反射率;二者分别对应MODIS数据的第2和第1波段, 中心波长分别为859 nm和645 nm.
1.4.2 蓝藻水华覆盖面积与暴发频率根据单像元蓝藻水华覆盖度(FCI), 本研究以FCI指数不等于0为判定标准, 统计覆盖度不为0的蓝藻水华像元个数, 并结合单像元的实际面积, 计算整个湖区蓝藻水华发生面积S.
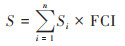
|
(3) |
式中, i为蓝藻水华影响的像元序号, n为蓝藻水华影响的像元总数, Si为第i个像元的面积, FCI为像元i的蓝藻水华覆盖度.
为对比分析蓝藻水华在不同等级和不同季节的暴发频率, 本研究依据《湖泊蓝藻水华卫星遥感监测技术导则》, 将蓝藻水华发生分为4个等级:无蓝藻水华覆盖(FCI = 0), 轻度水华覆盖(0 < FCI ≤ 30%)、中度水华覆盖(30% < FCI ≤ 60%)和重度水华覆盖(60% < FCI ≤ 100%), 然后分别在季节和年尺度上, 统计蓝藻水华的暴发频率. 蓝藻水华暴发频率定义为单像元在卫星遥感有效监测时段内, 蓝藻水华发生的相对频率[公式(4)]. 该指数的取值范围为0~100%, 数值越大, 说明该像元发生蓝藻水华的概率越高, 是蓝藻水华发生的热点区域.
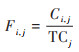
|
(4) |
式中, Fi, j为像元i在j时段内的蓝藻水华暴发频率, Ci, j为研究时段内单像元蓝藻水华发生不为0的次数, TCj为j时段内卫星遥感监测的影像总数.
1.4.3 蓝藻水华物候信息提取蓝藻水华物候是藻类生物长期适应节候所表现出的周期性变化规律, 本研究中特指蓝藻水华暴发开始时间与结束时间. 然而, 目前对蓝藻水华物候的定义尚未形成统一标准. 一般认为, 蓝藻水华暴发开始于每年的春季, 持续到第2年冬季结束. 本研究参照袁俊等[23]对巢湖蓝藻水华物候的定义, 选取自3月开始蓝藻水华面积首次超过全湖面积5%的日期为暴发开始时间, 最后一次藻华面积大于全湖5%的日期为结束时间. 本研究将上一年藻华暴发的结束时间到当年藻华首次暴发时间定义为非暴发期, 而暴发期则是当年蓝藻水华首次暴发到最后一次规模暴发的持续时间.
1.5 增强回归树模型 1.5.1 增强回归树简介增强回归树(BRT)是机器学习算法在统计学领域中的应用, 它由两种方法组合而成, 即回归树算法和增强技术, 二者的结合使其能准确描述自变量与因变量之间的相关关系, 量化各因子对因变量总体方差的解释能力, 探测多因子之间的交互作用[27]. BRT作为一种非参数的机器学习模型, 对数据分布形态无额外要求, 可适用于有序、离散、连续和分类的多种数据类型;同时, 该模型根据增强取样算法, 自动检测最佳拟合方案, 能有效减少缺失值和异常值对模型结果的影响. 根据BRT模型的运行结果, 自变量对因变量的作用关系以影响载荷的方式表达, 可轻松实现各指标因子对因变量解释能力相对贡献的量化, 并提供因子间的交互作用强度. BRT模型以其稳定和高效等优势, 已在自然地理区划[28]、物种分布预测[29]和生态变化驱动分析[30]等研究领域, 都取得了较为理想的结果. 有关BRT模型的基本原理、运行机制与应用领域详见Elith等[27]的研究.
1.5.2 模型参数设置及敏感性分析在应用BRT技术进行建模之前, 需要根据研究对象特征, 针对性设置适合该样本的相关模型参数. 总的来说, BRT共有5个关键参数需要用户预先定义, 即采样率、回归树复杂度、学习率、回归树数量和损失函数分布. 根据前人的研究经验, 本研究使用的损失函数分布类型为gaussian分布, 并设置模型的采样率为50%. 本研究未对回归树数量进行具体设置, 因为该参数是在模型运行过程中根据回归树复杂度和学习率自动确定的. 针对BRT模型中回归树复杂度和学习率两个参数, 本研究拟开展多组参数敏感性分析试验, 以探求实现最佳预测结果的参数配置. 在相关研究的基础上, 本文分别设置学习率为0.1、0.05和0.01, 树复杂度为2、4、6、8和10, 并采用交叉验证的方法对比任意参数组合下观测与拟合数据的平均绝对误差(MAE)和R2, 从而决定模型的最佳参数组合.
1.5.3 驱动力模型构建本研究中, 有关输入数据准备、BRT模型构建与结果后处理等操作均在R语言环境下进行, 主要用到了dismo、raster和sp等软件包. 为了研究不同时期蓝藻水华暴发的驱动因素, 本研究针对蓝藻水华暴发期和非暴发期, 构建2组BRT模型, 分别为:①暴发期内, 蓝藻细胞已大量生长并形成优势种群, 在适宜气象条件驱使下, 藻类优势种群上浮、聚集在水体表面, 形成水华. 由此可见, 气象因素是暴发期蓝藻水华形成的诱导因素. 因此, 本研究以卫星遥感监测的水华面积为因变量, 综合选取影响蓝藻水华发生的内因(藻密度)和外因(气象要素)构建BRT模型, 探索暴发期影响巢湖蓝藻水华面积变化的驱动因素. ②非暴发期间, 在适宜生态环境条件下, 冬季下沉到湖泊底泥的蓝藻细胞开始生长、繁殖并逐渐进入到水柱中. 该阶段, 蓝藻细胞密度的变化与水质、营养盐和藻细胞的生理活性都有一定联系. 为此, 本研究基于水上综合观测平台数据, 以水体藻密度为因变量, 以涵盖物理、化学和气象等要素的综合监测数据为自变量构建BRT模型, 从而探索非暴发期驱动水体藻密度变化的主控因子.
根据BRT模型的运行结果, 本研究以各因子贡献率作为衡量解释变量重要性的评价指标, 对暴发期和非暴发期各影响因子重要性进行排序, 识别影响巢湖蓝藻水华发生的主控因子. 同时绘制因变量随主导影响因子变化的偏依赖度分布, 分析蓝藻水华随主控因子取值变化的响应模式, 确定诱发蓝藻水华暴发的因子取值区间. 此外, 本研究依据BRT模型交互作用探测结果, 评价因子间交互作用强度, 并结合R语言的effects软件包, 进一步分析各因子的不同取值水平对蓝藻水华发生的综合影响.
2 结果与分析 2.1 巢湖蓝藻水华时空变化分析基于质控后的巢湖水上综合观测平台数据和卫星遥感蓝藻水华反演结果, 本研究首先分析了水体中藻密度与水华面积随时间的变化规律. 由图 2可知, 水体中藻密度表现出明显的季节变化, 藻密度在冬、春季普遍较低(< 3 000万个·L-1), 而在夏、秋季达到较高水平, 水体中平均藻密度在5 000万个·L-1以上. 根据水体藻密度变化, 孔繁翔等[31]将蓝藻水华的形成划分为4个阶段, 本文的研究结果也表现出类似的阶段变化. 即蓝藻细胞在春季开始复苏, 藻密度在3月以后缓慢上升;进入到6月, 蓝藻细胞开始大量积聚形成水华;而随着冬季气温的不断降低, 水体中藻密度急剧下降, 蓝藻开始进入休眠期.

|
图 2 2015~2020年巢湖藻密度与藻华面积时间变化 Fig. 2 Time series of cyanobacteria density and algal bloom area in Chaohu Lake from 2015 to 2020 |
依据卫星遥感监测结果(图 2), 巢湖蓝藻水华在春季和冬季暴发规模较小, 水华发生面积在100 km2以下, 夏、秋季是蓝藻水华的高发季节, 平均水华面积为165 km2. 2015年9月蓝藻水华面积高达386 km2, 为2015~2020年藻华面积峰值(最大值), 2016年以来, 巢湖蓝藻水华暴发面积峰值基本稳定在250 km2上下. 根据相关性分析结果, 巢湖蓝藻水华面积与蓝藻密度显著相关(R = 0.6), 二者表现为较一致的变化趋势, 表明蓝藻细胞是水华暴发的生物基础, 水体中蓝藻密度越大, 当年发生大规模水华暴发的概率也越高. 但在个别年份, 蓝藻密度与藻华面积偏离较大, 这同时也说明蓝藻水华发生还可能受到气象等其他环境因子的影响.
结合卫星遥感影像, 本研究进一步分析了不同程度蓝藻水华在不同季节的空间分布特征(图 3). 监测结果表明, 巢湖蓝藻水华以轻度暴发为主, 约占全年暴发频率的30%以上, 其次是重度暴发, 暴发频率在10%左右, 中度暴发占比最小. 具体到各季节, 轻度暴发的蓝藻水华在春季发生频率较高, 主要发生在巢湖西半湖和沿岸地区;而在秋季发生频率最小, 尤其是巢湖的中、东半湖区, 轻度水华发生的频率低于40%. 中度暴发的蓝藻水华在夏、秋季最为严重, 整个湖区约有60%以上的水域水华暴发程度较高, 表明夏、秋季是蓝藻水华规模暴发的主要季节;重度暴发的蓝藻水华主要发生在夏季, 整个湖区的暴发频率在20%以上, 秋季重度蓝藻水华发生的频率稍低(约在10%左右), 冬季虽然气温较低, 但依然有相当面积湖区发生重度水华, 表明巢湖蓝藻水华暴发持续时间较长, 冬季仍有可能发生大面积的蓝藻水华.

|
色柱表示蓝藻水华暴发频率, 单位为% 图 3 不同程度蓝藻水华暴发频率 Fig. 3 Spatial distribution of the frequency of different degrees of algal blooms in Chaohu Lake |
为探究气象条件对巢湖蓝藻水华发生的影响, 本研究全面选取包含温、压、湿、风等14个气象因子作为解释变量进行BRT建模. 由于解释变量之间天然存在某种联系, 各因子间可能存在一定的多重共线性. 为了消除这种影响, 本研究首先对各因子进行相关分析, 形成解释变量相关矩阵分布. 然后, 以相关系数 > 0.7为原则, 剔除因子间相关性较大的解释变量, 同时对剩余的变量进行多重共线性检测, 结果表明经筛选后的各因子间不存在或存在较弱的共线关系, 可用于下文BRT模型的构建. 最终筛选的气象因子包括:平均气温、相对湿度、日降水量、夜间降水量、平均风速、风向和日照时数.
针对BRT模型中回归树复杂度和学习率两个参数, 本文在总结现有研究的基础上, 设计了两参数不同取值组合下的15组参数敏感性试验, 并计算该参数设置下BRT模型的MAE平均值和R2值. 根据图 4的参数敏感性分析结果, 本研究以MAE最小、R2最大为判定依据, 识别不同时期巢湖蓝藻水华驱动因素分析模型的最佳参数组合. 最终得到非暴发期BRT模型的最佳参数组合为学习率0.1、回归树复杂度8, 暴发期实现BRT模型最优拟合的两参数分别为0.1和10.

|
黑色的虚线和数值表示不同参数组合下MAE值, 蓝色的实线和数值表示R2值 图 4 不同阶段蓝藻水华BRT模型参数敏感性分析 Fig. 4 Sensitivity evaluation of BRT model parameters for algal blooms at different stages |
根据藻密度与叶绿素a的定义, 二者均为反映水体中藻类数量和浓度的生物监测指标[32]. 为此, 本研究首先分析了水体中藻密度与叶绿素a浓度的相关关系. 结果表明, 巢湖水体中藻密度与叶绿素a浓度显著相关(R = 0.75), 二者表现出较一致的变化趋势. 因此, 在分析非暴发期蓝藻水华驱动因素时, 本文选取直接反映水体藻类生物浓度的藻密度作为因变量, 建立同水体中物理、化学和气象等要素的驱动分析模型. 根据观测数据对模型结果的交叉验证, 本文构建的BRT模型可解释水体藻密度变异程度的80%, 表明该模型能较好地预测非暴发期水体藻类密度变化. 本研究以影响因子贡献率作为衡量解释变量重要性的评价指标, 各因子的贡献率如图 5所示. 就单个因子而言, 非暴发期内藻密度受水体溶解氧(27.22%)、pH(11.86%)和水温(10.88%)的影响较大, 而降水、日照和风速风向等气象因子对非暴发期藻类密度变化影响较小(< 3%). 本文同时根据各因子的属性特征, 将17个解释变量划分为物理、化学和气象因素这3大类. 总体而言, 非暴发期内水体藻密度受物理因素影响最大, 因子贡献率总和可达48.59%, 其次是化学因素(贡献率之和为31.71%), 虽然气象要素对水体中藻密度的影响最小, 但在单个因子中, 气温对藻密度变化的贡献较大(7.43%). 综上, 非暴发期内, 巢湖藻类密度变化主要受物理和化学因素的影响, 针对非暴发期蓝藻水华的防治应着重从改善水体的理化环境入手.

|
图 5 非暴发期藻密度变化影响因子重要性排序 Fig. 5 Ranking the importance of factors affecting cyanobacteria density during the non-outbreak period |
驱动因子贡献率分析从整体上量化了各因子对水体藻密度变化的解释能力, 但无法反映因变量随自变量取值变化的响应模式. 为此, 本研究基于BRT模型的运行结果, 重点分析了非暴发期巢湖水体藻密度随主控影响因子的变化趋势(图 6). 依据藻密度随解释变量的变化趋势, 可将其分为逐渐增加、先增后减、逐渐减小和非线性变化这4种模式. 藻密度随水体溶解氧的变化表现为逐渐增加的模式, 当水体溶解氧较低时, 藻密度较小, 而随着溶解氧的升高, 水体藻密度不断增加, 表明水体中充足的氧气是非暴发期藻类细胞生长、繁殖的有利条件. 就pH和水温因子而言, 藻密度变化表现出随二者先增后减的变化模式, 即水体藻密度先随着pH和水温的增加而逐渐增大, 分别在pH值为7.5和水温3 ℃处达到最大, 随着pH和水温的继续增加, 藻密度开始呈现下降的变化趋势, 这一变化模式表明, pH和水温对水体藻密度的影响均存在一个最适区间, 超出这个区间外的取值均不利于蓝藻细胞的生长繁殖. 相对湿度对藻密度的影响表现为逐渐减小的变化模式, 即相对湿度较小时, 水体中藻密度大, 而高湿天气则不利于藻类细胞的繁殖. 其他因素对水体藻密度的影响表现为非线性变化模式, 即藻密度在解释变量取值范围内无明显变化规律, 表明这些因素对非暴发期藻类细胞的生长、繁殖影响较小.

|
黑色实心点表示解释变量与因变量的散点分布, 蓝色实线表示BRT模型在解释变量取值范围内对因变量的拟合, 灰色阴影表示95%置信区间 图 6 非暴发期藻密度对主导影响因子取值变化响应 Fig. 6 Response of cyanobacteria density to the value of dominant influence factors during the non-outbreak period |
根据非暴发期巢湖蓝藻驱动因素分析模型, 气温对藻类细胞的生长、繁殖起着关键作用. 为此, 本文通过引入农业气象上稳定通过界限温度的概念分析气温对藻类生物复苏的影响. 本研究分别计算了2015~2020年稳定通过3~15 ℃的初日, 并与蓝藻水华首次暴发日期对比, 旨在摸清非暴发期巢湖藻华首次发生与稳定通过界限温度的关系(表 2). 结果发现, 稳定通过7 ℃初日与蓝藻水华首次暴发日期变化趋势较为一致, 二者相关系数为0.46, 平均相隔天数为11.6 d. 即当巢湖气温达到7 ℃后, 藻类开始大量生长、繁殖, 在适宜的气象和水文条件驱使下聚集形成水华, 巢湖蓝藻水华在气温稳定通过7 ℃初日11 d前后首次出现.
|
|
表 2 蓝藻水华首次暴发与稳定通过界限温度日期的关系 Table 2 Relationship between the first outbreak date of algal blooms and the time of temperature steadily remaining above limited threshold |
2.5 暴发期巢湖蓝藻水华驱动因素分析
根据目前学者们广为接受的蓝藻水华形成理论, 当水体中蓝藻细胞达到一定密度时, 在适宜的气象条件驱动下上浮积聚形成水华, 蓝藻水华暴发是内因和外因综合作用的结果[8, 31]. 由非暴发期蓝藻水华影响驱动分析, 物理和化学等因素对水体藻密度变化的解释能力较强(方差解释率 > 80%), 为降低模型复杂度同时消除数据冗余, 本研究选取藻密度作为水华暴发的内因, 并全面选取包含温、压、湿、风和降水等14个气象因子作为藻华暴发的外力因子进行BRT建模. 在多因子共线性检测及变量筛选的基础上, 本文以遥感反演的蓝藻水华面积为因变量, 选取包含藻密度的内因以及气温、相对湿度、日降水量、夜间降水量、平均风速、风向和日照时数的外因作为解释变量构建暴发期藻华面积变化驱动模型.
根据暴发期蓝藻水华驱动分析模型, 研究发现, 气温对蓝藻水华暴发的影响最大, 占各要素总体贡献率的48%, 藻密度和日照时数的影响次之, 贡献率分别为22%和19%, 气温、藻密度、日照时数和风速这4个因子的累计贡献率高达95%, 表明暴发期蓝藻水华的发生主要与这4个因素密切相关(图 7). 为此, 本研究进一步分析了暴发期藻华面积对气温、藻密度、日照时数和风速的偏依赖度. 由图 8(a), 当气温在7 ℃以下, 偏依赖度为负且变化幅度较小, 表明低温对蓝藻水华发生具有一定抑制作用, 这种抑制作用随着气温的升高而逐渐减小, 偏依赖度在温度为15 ℃时开始转为正向促进, 并在18 ℃达到最大, 随后均保持在0以上, 表明高温是诱导蓝藻水华发生的有利因素. 就偏依赖度随藻密度变化趋势而言, 4 600万个·L-1以下藻密度均不利于蓝藻水华的发生, 高密度的藻类对藻华发生有促进作用, 但当水体藻密度超过一定阈值后, 该促进作用趋于平稳, 此时藻密度已不再是影响水华发生的主要因素[图 8(b)]. 风速对蓝藻水华的影响表现出逐渐减小的变化趋势, 2.8 m·s-1以下风速对蓝藻水华发生有促进作用, 风速过大则不利于蓝藻的上浮与积聚[图 8(d)]. 蓝藻水华随日照时数表现为先增后减的变化模式, 在自变量取值范围内, 偏依赖度首先缓慢增加, 在9.5 h日照处达到峰值后迅速减小, 当日照时数超过10.5 h, 则对蓝藻水华的发生有抑制作用[图 8(c)]. 以上结果表明, 巢湖蓝藻水华发生受藻类生物量和气象条件的综合影响, 各因子对蓝藻水华的影响均存在一个最适区间, 超出或低于这个区间的因子取值均不利于巢湖蓝藻水华的发生.

|
图 7 暴发期蓝藻水华影响因子贡献率 Fig. 7 Contribution rate of factors affecting algal blooms during the outbreak period |

|
彩色实线表示根据BRT模型计算的因变量随解释变量变化的偏依赖度分布, 绿色阴影表示解释变量的概率密度分布 图 8 暴发期蓝藻水华主导驱动因子偏依赖度分布 Fig. 8 Partial dependence plots for dominant driving factors during the outbreak period |
蓝藻水华暴发是多种因素共同作用的结果, 各驱动因子间相互制约和相互影响产生的综合效应可能会加强或削弱单个因子的作用模式, 即存在一定的交互作用. 为此, 本文根据BRT模型交互作用探测结果, 进一步分析了暴发期各因子交互作用对蓝藻水华的影响. 结果表明, 气温和风速、气温和藻密度之间存在较强的交互作用. 就气温和风速的交互作用而言, 蓝藻水华在10~20 ℃及2.6 m·s-1以下风速发生面积较大, 表明适温、小风的气象条件最有利于巢湖蓝藻水华发生[图 9(a)]. 在气温稳定条件下, 风速增大一定程度上能抑制藻华暴发. 此外, 本研究还分析了不同藻密度条件下, 藻华面积对气温的线性响应关系. 由图 9(b), 水华面积对气温的回归系数随藻密度的增加不断增大, 表明气温一定时, 水体藻密度越大, 越有利于巢湖蓝藻水华的暴发.

|
图 9 驱动因子交互作用分析 Fig. 9 Interactive detection of driving factors |
利用遥感影像准确提取蓝藻水华面积, 是监测蓝藻水华时空动态及驱动分析的前提. 基于藻类生物在不同光谱波段的变化特征, 浮游藻类指数(FAI)与蓝藻水华覆盖度(FCI)被广泛应用于水体蓝藻水华的识别与提取[2, 5]. 为评价不同指数对巢湖蓝藻水华反演的不确定性, 本研究进一步对比分析了基于FAI与FCI指数反演蓝藻水华的面积(图 10). 结果表明, 基于FAI和FCI指数提取的藻华面积一致性较高, 相关系数可达0.75. 二者表现为相同的时间变化趋势, 尤其是对大面积暴发的蓝藻水华, 二者监测结果较为一致. 然而, 基于FAI指数提取的藻华面积总体上大于FCI指数提取的藻华面积, 约平均偏大28 km2. 究其原因, 可能是两种指数所采用光谱波段的差异, 导致对“异物同谱”像元误分产生的偏差. 根据袁俊等[23]的研究, 由于水陆边界的临近效应, FAI指数可能会将靠近陆地的像元判定为蓝藻水华, 从而导致对藻华面积的高估[9]. 虽然两种方法对巢湖蓝藻水华的反演存在一定差异, 但二者总体变化趋势较为一致, 对下文蓝藻水华的驱动因素分析影响较小. 即便如此, 加强人工智能和深度学习等新方法在蓝藻水华提取中的应用, 减小蓝藻水华的提取误差, 实现蓝藻水华的智能化和自动化提取, 仍然是当前亟需开展深入研究的关键技术问题之一[10, 25].

|
图 10 基于FCI和FAI指数提取的藻华面积对比 Fig. 10 Comparison of algal bloom area extracted from FCI and FAI index |
BRT模型作为一种非参数的机器学习算法, 能有效度量各影响因子的重要性及其对因变量的变化响应规律, 尤为适用于非线性变化和发生机制复杂的蓝藻水华驱动因素分析[27]. 基于机器学习算法的特征重要性评估和预测在相关领域中显示出广阔的应用前景. 然而, 作为一个数据驱动型算法, BRT模型的输出结果可能会受到输入数据微小变化的影响而发生显著改变, 即模型表现出对输入数据较强的敏感性, 从而导致模型出现过拟合或欠拟合等问题, 影响对观测现象的解释和预测[29, 30]. 为提高模型对输入数据的鲁棒性和稳定性, 本文通过开展多组参数敏感性分析试验, 探求实现最佳预测结果的模型参数配置, 从而确保模型能够正确处理不同类型和大小的输入数据, 提取有价值的特征变量和属性信息. 尽管本文的研究结果可能受到蓝藻水华监测样本数量的影响, 但BRT模型仍显示出对因子重要性评估和交互作用探测的有效性, 模型结果对于识别诱导蓝藻水华发生的主控因子以及预测蓝藻水华暴发风险具有重要的现实意义.
3.2 巢湖蓝藻水华驱动因素科学评估蓝藻水华驱动因素是蓝藻水华预测、预警和防治工作的基础, 本研究根据国内外流行的蓝藻水华形成四阶段理论, 重点探讨了暴发期和非暴发期影响巢湖蓝藻水华发生的驱动因素, 部分成果证实了前人的假设, 同时也得出了针对巢湖蓝藻水华驱动因素的独有结论. 本研究认为, 非暴发期蓝藻细胞密度变化主要与水体溶解氧和水温等物理因素有关, 化学因素对蓝藻的影响较小. 水体中氮、磷等营养盐为蓝藻复苏提供了充足的物质来源, 富营养化是蓝藻水华暴发的先决条件[1, 7]. 近年来, 围绕巢湖水环境治理开展了大量工作, 巢湖水质有一定程度改善[20], 但2021年巢湖湖区水质仍为Ⅳ类, 呈中度富营养状态. 已有研究普遍认为, 在高浓度富营养化水体中, 营养盐浓度长期维持较高水平, 已不再是影响蓝藻生长的限制因子[12, 33]. 本文的研究结果也证实了这一结论, 水体藻密度表现出对总氮、总磷非线性的变化模式, 水体中氮、磷等营养盐浓度对非暴发期蓝藻变化的解释程度仅占总体贡献率的15.69%. 因此, 继续推进巢湖生态治理、改善巢湖水质环境, 仍然是当前巢湖蓝藻水华防治的根本工作. 此外, 本研究还发现, 气温是影响非暴发期蓝藻密度变化的主要气象因子, 这一结论与罗晓春等[13]和吴晓东等[34]的研究结果较为一致, 他们认为气温是影响复苏期蓝藻细胞生长的关键因素, 谢小萍等[14]进一步分析了太湖蓝藻复苏与稳定通过界限温度的关系, 发现蓝藻水华首次暴发一般在气温稳定通过9 ℃后1个月出现, 略高于本文稳定通过7 ℃初日的研究结果. 综上, 气温是影响淡水湖泊藻类复苏的关键气象因子.
根据前人的研究成果, 气象条件是蓝藻水华暴发的充分而非必要条件, 是推动蓝藻水华形成的外力因子[12, 31]. 本研究结果发现, 巢湖蓝藻水华暴发主要受气温、日照时数和风速的影响. 其中, 气温对蓝藻水华发生起主导作用, 占各要素总体贡献率的48%. 罗晓春等[13]的研究同时也证实了气温对蓝藻水华的重要影响, 在其构建的多组随机森林驱动分析模型中, 气温作为首要影响因子的频次高达80%, 在蓝藻生长和水华形成的不同阶段均起着关键作用. 随着气候变暖的不断加剧, 气温攀升将会导致蓝藻水华物候和暴发强度的改变, 进一步增大蓝藻水华防治难度[3, 25]. 因此, 未来需重点关注气候变化对巢湖蓝藻水华的影响. 日照是藻类进行光合作用的必要条件, 是蓝藻细胞通过伪空胞调节自身浮力的催化剂[31]. 李阔宇等[35]的研究表明, 日照条件能影响蓝藻细胞光系统Ⅱ的活性, 是影响蓝藻细胞生长、繁殖的主要限制因子. 杭鑫等[36]的研究结果也表明, 充足的日照是蓝藻水华形成的必要条件之一, 而日照时数过长能在一定程度上降低蓝藻细胞活性, 抑制蓝藻水华发生. 这与本研究的结论较一致, 即日照对蓝藻水华的影响存在一个最适区间, 超出或低于这个区间均不利于蓝藻水华的形成. 有关风速对蓝藻水华的影响, 已有研究的结论趋于一致, 一般认为, 3~4 m·s-1或更小的风速最有利于蓝藻水华的形成, 风速过大则会导致蓝藻细胞在水柱中充分混合, 不利于蓝藻的上浮积聚[16, 25]. 风向对蓝藻水华的影响主要体现在其能使湖泊水体发生流动, 从而带动蓝藻集聚形成水华. 然而, 由于巢湖湖区风向高度不稳定且复杂多变, 针对风向对巢湖蓝藻水华影响的研究, 结论分歧较大[8]. 如胡旻琪等[5]认为巢湖北岸蓝藻堆积是春夏季南风持续作用的结果, 而南岸蓝藻水华发生频率较高则与冬季盛行的西北风有关[2]. 总之, 风速和风向作为蓝藻水华暴发的动力因子, 共同决定蓝藻水华的聚集程度和分布位置.
3.3 不确定性分析及对未来研究的启示虽然本研究系统分析了暴发期和非暴发期巢湖蓝藻水华形成的驱动因素, 但由于受到多源数据时空不匹配和统计分析手段的限制, 本文还存在较大的不确定性. 首先, 基于卫星遥感的蓝藻水华监测大都发生在无云、晴朗的天气, 这在一定程度上掩盖了降水对蓝藻水华生消的影响. 根据Haakonsson等[37]和Zhou等[38]的研究, 由大规模降水导致的湖泊水位上升, 一方面能够稀释水体中营养盐物质浓度, 另一方面降水带来的水体扰动能够抑制藻类的上浮聚集, 从而不利于蓝藻水华的大面积暴发. 降水被认为是影响巢湖蓝藻水华面积月际变化的主要驱动因素之一[39, 40]. 然而, 本研究结果表明, 降水对解释暴发期蓝藻水华发生的贡献率仅占气象因子的1%, 严重低估了降水对巢湖蓝藻水华的影响. 因此, 在实地蓝藻水华监测数据可获取的情况下, 着重分析不同强度和持续时长雨量等级对巢湖蓝藻水华的影响, 将是下一步研究的重点. 其次, 在暴发期巢湖蓝藻水华驱动分析建模时, 本研究使用的气象资料为巢湖国家一般气象站点监测数据, 该站点仅代表观测位置10 km范围内气象要素的平均状态. 而因地形差异, 整个湖区气象要素变化较大, 表现为典型的局地气候特征[8]. 未来研究有必要结合中尺度气象模式, 融合精细化气象要素场, 实现气象要素对不同湖区蓝藻水华暴发影响的精准分析. 最后, 蓝藻水华变化具有高时空动态性, 湖区藻华发生面积和位置可在一天内发生多次变化[23, 25]. 受限于遥感数据的时间分辨率, 基于Landsat和MODIS数据难以实现对藻华日内变化的动态分析. 因此, 尝试利用雷达和高分遥感数据(如GOCI和高分三号)进行多源数据的联合反演, 可进一步提升蓝藻水华的动态监测能力.
4 结论(1)巢湖水体藻密度表现出较大的季节变化特征, 蓝藻细胞在春季开始复苏, 藻密度在夏、秋季达到最大, 冬季藻密度最低. 卫星遥感监测结果表明, 蓝藻水华轻度暴发在春季发生频率较高, 且主要发生在巢湖西半湖和沿岸地区, 中等程度以上的蓝藻水华暴发在夏、秋季较为严重.
(2)非暴发期间, 巢湖藻密度变化受物理和化学因素影响较大, 二者对解释藻密度方差变化的贡献率可达80.3%. 根据藻密度对各因子取值的变化响应图, 水体中高浓度的溶解氧为藻类复苏提供了有利条件, pH和水温对藻密度的影响均存在一个最适区间, 弱碱性水体和3 ℃水温最有利于藻类细胞的生长繁殖. 蓝藻水华首次暴发日期与稳定通过7 ℃初日密切相关, 一般在气温稳定通过7 ℃初日11 d前后首次出现.
(3)暴发期内, 巢湖蓝藻水华发生主要受藻类生物量和气象条件的综合影响, 气温、藻密度、日照时数和风速的累计贡献率高达95%, 且各因子均存在一个有利于蓝藻水华发生的最适区间. 根据多因子交互作用分析结果, 巢湖蓝藻水华在水体藻密度较大、气温适宜和微风等环境条件下发生概率最大.
致谢: 感谢中国科学院南京地理与湖泊研究所段洪涛研究员及其团队成员提供的遥感影像数据支持.
| [1] |
朱广伟, 许海, 朱梦圆, 等. 三十年来长江中下游湖泊富营养化状况变迁及其影响因素[J]. 湖泊科学, 2019, 31(6): 1510-1524. Zhu G W, Xu H, Zhu M Y, et al. Changing characteristics and driving factors of trophic state of lakes in the middle and lower reaches of Yangtze River in the past 30 years[J]. Journal of Lake Sciences, 2019, 31(6): 1510-1524. |
| [2] |
祁国华, 马晓双, 何诗瑜, 等. 基于多源遥感数据的巢湖水华长时序时空变化(2009-2018年)分析与发生概率预测[J]. 湖泊科学, 2021, 33(2): 414-427. Qi G H, Ma X S, He S Y, et al. Long-term spatiotemporal variation analysis and probability prediction of algal blooms in Lake Chaohu (2009-2018) based on multi-source remote sensing data[J]. Journal of Lake Sciences, 2021, 33(2): 414-427. |
| [3] | Yan X C, Xu X G, Wang M Y, et al. Climate warming and cyanobacteria blooms: Looks at their relationships from a new perspective[J]. Water Research, 2017, 125: 449-457. DOI:10.1016/j.watres.2017.09.008 |
| [4] | Liu J G, Yang W. Water sustainability for China and beyond[J]. Science, 2012, 337(6095): 649-650. DOI:10.1126/science.1219471 |
| [5] |
胡旻琪, 张玉超, 马荣华, 等. 巢湖2016年蓝藻水华时空分布及环境驱动力分析[J]. 环境科学, 2018, 39(11): 4925-4937. Hu M Q, Zhang Y C, Ma R H, et al. Spatial and temporal dynamics of floating algal blooms in Lake Chaohu in 2016 and their environmental drivers[J]. Environmental Science, 2018, 39(11): 4925-4937. |
| [6] |
欧阳添, 赵璐, 纪璐璐, 等. 蓝藻水华过程中优势种群演替模式、效应及驱动因子分析[J]. 环境科学, 2022, 43(10): 4480-4488. Ouyang T, Zhao L, Ji L L, et al. Succession pattern and consequences of the dominant species during cyanobacterial bloom and its influencing factors[J]. Environmental Science, 2022, 43(10): 4480-4488. |
| [7] |
温成成, 黄廷林, 孔昌昊, 等. 北方富营养分层型水库藻类季节性暴发机制及其阈值分析[J]. 环境科学, 2023, 44(3): 1452-1464. Wen C C, Huang T L, Kong C H, et al. Analysis of mechanism and start-up thresholds of seasonal algal blooms in a northern eutrophic stratified reservoir[J]. Environmental Science, 2023, 44(3): 1452-1464. |
| [8] |
秦伯强, 杨桂军, 马健荣, 等. 太湖蓝藻水华"暴发"的动态特征及其机制[J]. 科学通报, 2016, 61(7): 759-770. Qin B Q, Yang G J, Ma J R, et al. Dynamics of variability and mechanism of harmful cyanobacteria bloom in Lake Taihu, China[J]. Chinese Science Bulletin, 2016, 61(7): 759-770. |
| [9] | Hu C M. A novel ocean color index to detect floating algae in the global oceans[J]. Remote Sensing of Environment, 2009, 113(10): 2118-2129. DOI:10.1016/j.rse.2009.05.012 |
| [10] | Ma J Y, Jin S G, Li J, et al. Spatio-temporal variations and driving forces of harmful algal blooms in Chaohu Lake: a multi-source remote sensing approach[J]. Remote Sensing, 2021, 13(3). DOI:10.3390/rs13030427 |
| [11] |
唐晓先, 沈明, 段洪涛. 巢湖蓝藻水华时空分布(2000-2015)[J]. 湖泊科学, 2017, 29(2): 276-284. Tang X X, Shen M, Duan H T. Temporal and spatial distribution of algal blooms in Lake Chaohu, 2000-2015[J]. Journal of Lake Sciences, 2017, 29(2): 276-284. |
| [12] | Zhang M, Duan H T, Shi X L, et al. Contributions of meteorology to the phenology of cyanobacterial blooms: implications for future climate change[J]. Water Research, 2012, 46(2): 442-452. DOI:10.1016/j.watres.2011.11.013 |
| [13] |
罗晓春, 杭鑫, 曹云, 等. 太湖富营养化条件下影响蓝藻水华的主导气象因子[J]. 湖泊科学, 2019, 31(5): 1248-1258. Luo X C, Hang X, Cao Y, et al. Dominant meteorological factors affecting cyanobacterial blooms under eutrophication in Lake Taihu[J]. Journal of Lake Sciences, 2019, 31(5): 1248-1258. |
| [14] |
谢小萍, 李亚春, 杭鑫, 等. 气温对太湖蓝藻复苏和休眠进程的影响[J]. 湖泊科学, 2016, 28(4): 818-824. Xie X P, Li Y C, Hang X, et al. The effect of air temperature on the process of cyanobacteria recruitment and dormancy in Lake Taihu[J]. Journal of Lake Sciences, 2016, 28(4): 818-824. |
| [15] |
刘心愿, 宋林旭, 纪道斌, 等. 降雨对蓝藻水华消退影响及其机制分析[J]. 环境科学, 2018, 39(2): 774-782. Liu X Y, Song L X, Ji D B, et al. Effect of the rainfall on extinction of cyanobacteria bloom and its mechanism analysis[J]. Environmental Science, 2018, 39(2): 774-782. |
| [16] |
李亚春, 谢小萍, 杭鑫, 等. 结合卫星遥感技术的太湖蓝藻水华形成风场特征[J]. 中国环境科学, 2016, 36(2): 525-533. Li Y C, Xie X P, Hang X, et al. Analysis of wind field features causing cyanobacteria bloom in Taihu Lake combined with remote sensing methods[J]. China Environmental Science, 2016, 36(2): 525-533. DOI:10.3969/j.issn.1000-6923.2016.02.032 |
| [17] |
鲁韦坤, 余凌翔, 欧晓昆, 等. 滇池蓝藻水华发生频率与气象因子的关系[J]. 湖泊科学, 2017, 29(3): 534-545. Lu W K, Yu L X, Ou X K, et al. Relationship between occurrence frequency of cyanobacteria bloom and meteorological factors in Lake Dianchi[J]. Journal of Lake Sciences, 2017, 29(3): 534-545. |
| [18] | Yu P X, Gao R, Zhang D Z, et al. Predicting coastal algal blooms with environmental factors by machine learning methods[J]. Ecological Indicators, 2021, 123. DOI:10.1016/j.ecolind.2020.107334 |
| [19] | 王苏民, 窦鸿身. 中国湖泊志[M]. 北京: 科学出版社, 1998. |
| [20] |
张民, 孔繁翔. 巢湖富营养化的历程、空间分布与治理策略(1984-2013)[J]. 湖泊科学, 2015, 27(5): 791-798. Zhang M, Kong F X. The process, spatial and temporal distributions and mitigation strategies of the eutrophication of Lake Chaohu (1984-2013)[J]. Journal of Lake Sciences, 2015, 27(5): 791-798. |
| [21] |
邓莎, 周键. 蓝藻水华的危害及主要控制技术研究进展[J]. 安徽农学通报, 2020, 26(18): 150-151, 198. Deng S, Zhou J. Research advances in the harm of cyanobacteria bloom and its main control technology[J]. Anhui Agricultural Science Bulletin, 2020, 26(18): 150-151, 198. DOI:10.3969/j.issn.1007-7731.2020.18.058 |
| [22] |
曹畅, 王胜蕾, 李俊生, 等. 基于MODIS数据的全国144个重点湖库营养状态监测: 以2018年夏季为例[J]. 湖泊科学, 2021, 33(2): 405-413. Cao C, Wang S L, Li J S, et al. MODIS-based monitoring of spatial distribution of trophic status in 144 key lakes and reservoirs of China in summer of 2018[J]. Journal of Lake Sciences, 2021, 33(2): 405-413. |
| [23] |
袁俊, 曹志刚, 马金戈, 等. 1980s以来巢湖藻华物候时空变化遥感分析[J]. 湖泊科学, 2023, 35(1): 57-72. Yuan J, Cao Z G, Ma J G, et al. Remote sensed analysis of spatial and temporal variation in phenology of algal blooms in Lake Chaohu since 1980s[J]. Journal of Lake Sciences, 2023, 35(1): 57-72. |
| [24] | Yang Y, Wang T, Wang P C, et al. In-situ measurement of CO2 at the Xinglong regional background station over North China[J]. Atmospheric and Oceanic Science Letters, 2019, 12(6): 385-391. DOI:10.1080/16742834.2019.1644949 |
| [25] | Le C F, Hu C M, English D, et al. Climate-driven chlorophyll-a changes in a turbid estuary: Observations from satellites and implications for management[J]. Remote Sensing of Environment, 2013, 130: 11-24. DOI:10.1016/j.rse.2012.11.011 |
| [26] | QX/T 207-2013, 湖泊蓝藻水华卫星遥感监测技术导则[S]. |
| [27] | Elith J, Leathwick J R, Hastie T. A working guide to boosted regression trees[J]. Journal of Animal Ecology, 2008, 77(4): 802-813. DOI:10.1111/j.1365-2656.2008.01390.x |
| [28] | Martin V, Pfeiffer D U, Zhou X Y, et al. Spatial distribution and risk factors of highly pathogenic avian influenza (HPAI) H5N1 in China[J]. PLoS pathogens, 2011, 7(3). DOI:10.1371/journal.ppat.1001308 |
| [29] | Gu H L, Wang J, Ma L J, et al. Insights into the BRT (boosted regression trees) method in the study of the climate-growth relationship of Masson pine in subtropical China[J]. Forests, 2019, 10(3). DOI:10.3390/f10030228 |
| [30] | Lamb S E, Haacker E M K, Smidt S J. Influence of irrigation drivers using boosted regression trees: Kansas high plains[J]. Water Resources Research, 2021, 57(5). DOI:10.1029/2020WR028867 |
| [31] |
孔繁翔, 马荣华, 高俊峰, 等. 太湖蓝藻水华的预防、预测和预警的理论与实践[J]. 湖泊科学, 2009, 21(3): 314-328. Kong F X, Ma R H, Gao J F, et al. The theory and practice of prevention, forecast and warning on cyanobacteria bloom in Lake Taihu[J]. Journal of Lake Sciences, 2009, 21(3): 314-328. DOI:10.3321/j.issn:1003-5427.2009.03.002 |
| [32] |
荀尚培, 杨元建, 何彬方, 等. 春季巢湖水温和水体叶绿素a浓度的变化关系[J]. 湖泊科学, 2011, 23(5): 767-772. Xun S P, Yang Y J, He B F, et al. Analysis of relationship between spring water temperature and chlorophyll-a in Lake Chaohu[J]. Journal of Lake Sciences, 2011, 23(5): 767-772. |
| [33] |
荣洁, 王腊春. 指数平滑法-马尔科夫模型在巢湖水质预测中的应用[J]. 水资源与水工程学报, 2013, 24(4): 98-102. Rong H, Wang L C. Application of the exponential smoothing law-Markov model in prediction of water quality of Chaohu lake[J]. Journal of Water Resources & Water Engineering, 2013, 24(4): 98-102. |
| [34] |
吴晓东, 孔繁翔, 张晓峰, 等. 太湖与巢湖水华蓝藻越冬和春季复苏的比较研究[J]. 环境科学, 2008, 29(5): 1313-1318. Wu X D, Kong F X, Zhang X F, et al. Comparison of overwintering and recruitment of cyanobacteria in Taihu Lake and Chaohu Lake[J]. Environmental Science, 2008, 29(5): 1313-1318. DOI:10.3321/j.issn:0250-3301.2008.05.028 |
| [35] |
李阔宇, 宋立荣, 万能. 底泥中微囊藻复苏和生长特性的研究[J]. 水生生物学报, 2004, 28(2): 113-118. Li K Y, Song L R, Wan N. Studies on recruitment and growth characteristic of microcystis in sediment[J]. Acta Hydrobiologica Sinica, 2004, 28(2): 113-118. DOI:10.3321/j.issn:1000-3207.2004.02.001 |
| [36] |
杭鑫, 罗晓春, 谢小萍, 等. 太湖蓝藻水华形成的适宜气象指标[J]. 气象科技, 2019, 47(1): 171-178. Hang X, Luo X C, Xie X P, et al. Suitable meteorological indicators for formation of cyanobacteria blooms in Taihu Lake[J]. Meteorological Science and Technology, 2019, 47(1): 171-178. |
| [37] | Haakonsson S, Rodríguez-Gallego L, Somma A, et al. Temperature and precipitation shape the distribution of harmful cyanobacteria in subtropical lotic and lentic ecosystems[J]. Science of the Total Environment, 2017, 609: 1132-1139. DOI:10.1016/j.scitotenv.2017.07.067 |
| [38] | Zhou Y T, Yan W J, Wei W Y. Effect of sea surface temperature and precipitation on annual frequency of harmful algal blooms in the East China Sea over the past decades[J]. Environmental Pollution, 2021, 270. DOI:10.1016/j.envpol.2020.116224 |
| [39] | 范裕祥, 金社军, 周培, 等. 巢湖蓝藻水华暴发与气象条件的关系[A]. 见: 第四届中国湖泊论坛论文集[C]. 合肥: 中国科学技术协会, 2014. 492-496. |
| [40] |
李晟铭, 刘吉平, 宋开山, 等. 基于Landsat影像巢湖蓝藻水华暴发时空变化特征及其驱动因素分析[J]. 长江流域资源与环境, 2019, 28(5): 1205-1213. Li S M, Liu J P, Song K S, et al. Analysis on spatial and temporal character of algae bloom in Lake Chaohu and its driving factors based on Landsat imagery[J]. Resources and Environment in the Yangtze Basin, 2019, 28(5): 1205-1213. |