 2024, Vol. 45
2024, Vol. 45



2. 多特蒙德工业大学空间规划学院, 多特蒙德 44135
2. Department of Spatial Planning, Technical University of Dortmund, Dortmund 44135, Germany
恢复与提升植被生态系统的质量和稳定性是现如今各大生态问题的最优解[1 ~ 3], 基于此, 许多国内外学者通过研究植被生理指数以达到研究植被生态系统变化的目的, 其中, 植被净初级生产力(net primary productivity, NPP)便是其一.植被NPP作为植被光合作用效率和评估植被生长状况的一项重要指标, 可用于监测植被生长状况[4, 5]、评估生态环境质量[6]和探究碳循环现状[7]等.
全球范围内, Li等[8]通过研究全球植被NPP变化规律, 发现在全球变暖背景下, 全球热液平衡对植被NPP变化起到正向作用.Bejagam等[9]基于印度植被生态系统NPP, 发现降水是印度植被生态系统NPP的主导控制因素, 气候因素促进了印度生态系统植被NPP的上升, 而人类活动则存在抑制作用.在中国, 石智宇等[10]和Yu等[11]研究发现, 近20年来, 中国植被NPP在空间上呈现“东南高西北低”的分布格局, 时间上呈波动增加趋势, 气候变化主导的植被NPP上升区主要集中在东北和华北等地, 人类活动主导植被NPP上升区主要集中在华中和西北等地区, 而在华北的内蒙古地区, 植被NPP的变化在不同季节略有不同, 在生长季和夏秋季植被NPP总体呈下降趋势, 而春季略有上升趋势.现如今, 针对西南地区植被NPP变化及驱动因素的研究也逐渐深入, 外部驱动因素对西南地区植被NPP的影响也逐渐明朗, 徐勇等[12, 13]、Linger等[14]和茆杨等[15]将地形地貌、不同气候因子、气候变化的滞后效应和气候的季节变化等多因素考虑在内, 对西南地区植被NPP的时空变化规律及驱动因素进行了细致的研究.以上研究均致力于外部影响因素对植被生长的影响, 忽略了不同植被类型生长本身存在的客观差异, 且已有研究表明, 不同类型的植被对碳循环的影响存在差异, 不同植被类型对土壤生物群落和碳循环基因丰度等均有显著影响[16, 17].现如今针对植被自身客观差异的影响研究, 均以研究不同类型植被NDVI为主[18 ~ 21], 以不同类型植被NPP为主体的研究较少.鉴于此, 本文依据植被结构的不同, 同时考虑到人文因素对某些植被存在较大干预, 将植被分为乔木、灌木、草本、经济类和其他.本文假定在研究时段内土地利用类型发生改变, 但土地利用未发生变化的部分植被类型不随时间的迁移发生改变, 将存在植被衍生和消亡的区域(即土地利用类型发生改变的区域)进行删除, 增加实验的可靠性, 以研究2000 ~ 2021年西南地区不同类型植被NPP时空变化规律, 并利用稳定性分析与地理探测器, 揭示不同类型植被自身和气候影响因素对植被生长影响间的关系, 以期为更好厘定植被生长差异与气候影响因素的驱动机制提供新思路.
1 研究区概况研究区地处中国的西南部, 位于78°25′ ~ 110°11′E, 21°08′ ~ 36°53′N间, 含西藏、四川、重庆、贵州和云南这5个省级行政单元, 面积达234万km2.研究区是我国重要生态经济发展区, 植被类型丰富, 含乔木、灌木、草本和经济类等多种类型植被(如图 1), 受热带、亚热带季风气候和高山高原气候影响, 研究区平均气温维持在-2.78 ~ 23.93℃间, 年累计降雨量为24 ~ 2 638 mm, 平均相对湿度约58%, 水热条件适宜的同时, 每年2 200 h左右的日照时数为研究区植被的生长提供了充足的能量来源.然而, 研究区地势起伏较大, 海拔在62 ~ 8 405 m之间, 加上研究区为典型的喀斯特地貌区, 使得生态环境脆弱敏感的同时, 经济发展较为缓慢.

|
图 1 研究区植被类型空间分布示意 Fig. 1 Spatial distribution of vegetation types in the study area |
空间分辨率为500 m的MODIS NPP数据来源于National Aeronautics and Space Administration(NASA)发布的MOD17A3 C6产品(https://earthdata.nasa.gov/), 原始数据格式为HDF, 每年一期.原始数据需进行滤波、镶嵌、重采样和重投影等预处理, 从而获取1 km × 1 km覆盖西南地区的2000 ~ 2021年植被NPP时间序列.DEM数据、2000年和2020年土地利用数据和1990年代中国1∶100万植被类型空间分布数据均来源于中国科学院资源环境数据云平台(https://www.resdc.cn/), 以上数据空间分辨率均为1 km × 1 km.
1999 ~ 2021年气候因子(最低气温、最高气温、平均气温、降水、相对湿度、日照时数、气压和地表温度)来源于国家气象科学数据中心提供的逐日数据(https://data.cma.cn/), 共2 416个气象站点.首先, 需要剔除原始数据缺失造成的影响;然后, 使用ANUSPLIN进行空间插值以获取覆盖研究区、空间分辨率为1 km×1 km的气象数据时间序列.通过平均气温和降水时间序列, 计算获取可能蒸散率、生物干燥度指数、湿润指数、温暖指数和寒冷指数时间序列, 因此, 本文选取以上13个因子作为影响植被NPP变化的气候因子.本实验以全国2 416个气象站点经插值处理后取得全国气象栅格数据, 经裁剪后取得研究区数据.
本文假设2000 ~ 2020年土地利用类型不发生变化的部分植被类型不随时间发生改变.为降低植被类型因时间迁移发生改变的客观影响, 首先, 通过对2000年和2020年土地利用类型做叠置分析, 提取研究时段内土地利用类型不发生变化的部分;然后, 将其与植被类型进行叠加, 从而获取研究时段内西南地区不同类型植被分布区域.
2.2 研究方法 2.2.1 气候变化和人类活动对植被NPP影响的定量剥离植被NPP变化受多种气候因子的调控影响, 且不同类型植被生长所需自然环境存在一定差异, 在考虑到植被生长对气候变化存在滞后效应的情况下, 本文通过联合相关分析、显著性检验和一阶偏导法[13], 为多元回归分析模型进行主导气候因子的选取.考虑到主要影响因素间可能会存在严重的多重共线性问题, 这会导致数据冗余的同时造成残差模型失真和预测结果的不稳定, 对此, 本文通过使用方差膨胀系数(variance inflation factor, VIF)检验并消除主要影响因子间的多重共线性影响.多元回归分析模型因子筛选结果如表 1所示.
|
|
表 1 不同植被类型主导气候因子筛选结果 Table 1 Result of the selection of dominant climate factors of different vegetation types |
基于上述因子选取结果, 同时结合不同类型植被NPP观测值(即植被NPP), 建立基于不同植被类型的多元回归分析模型, 生成不同气候条件影响下的不同类型植被NPP值(NPPPre, 下同), 以此探究不同类型植被NPP在气候变化下的时空变化;同时, 建立基于不同植被类型的残差分析模型, 在剥离气候因素影响的条件下, 计算出反映人类活动影响下的不同类型植被NPP值(NPPRes, 下同), 以探究不同类型植被NPP在人类活动影响下的时空变化.植被NPPPre和植被NPPRes计算公式如下[13]:
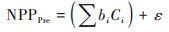
|
(1) |
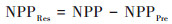
|
(2) |
式中, bi为系数;Ci为不同类型植被NPP变化对应的主导因子;ε为常量.
最后, 基于上述计算得到的气候变化影响下植被NPPPre、人类活动影响下植被NPPRes和不同类型植被NPP观测值, 利用Theil-Sen Median趋势分析法对上述不同影响条件情景下植被NPP时间序列数据的趋势进行估计[22 ~ 24], 其计算公式为:
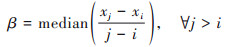
|
(3) |
式中, xi和xj分别为第i年和第j年的植被NPP值;median为取中值函数.β为变化斜率, 将β > 0、β < 0和β = 0作为植被NPP上升、下降和不变的判定条件.同时, 结合Mann-Kendall显著性检验法对西南地区植被NPP变化趋势进行显著性检验[25, 26], 当|MK| > 2.58时, 在P = 0.01水平上显著, 记为极显著变化;当1.96 < |MK| ≤ 2.58时, 在P = 0.05水平上显著, 记为显著变化;其余记为不显著变化.
2.2.2 不同类型植被NPP变化探测首先, 本文在像元尺度上计算了2000 ~ 2021年西南地区不同类型植被NPP的变异系数, 用于评估植被NPP在时间序列上的稳定性[27].计算公式如下:
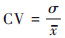
|
(4) |
式中, CV为植被NPP变异系数;σ为植被NPP标准差;x为植被NPP平均值.CV值越大, 表示数据分布越离散, 植被NPP年际变化越大;反之, 则表示数据分布紧凑, 植被NPP年际变化较为稳定.根据自然间断点法将植被NPP的稳定性分成5类:低波动变化(0.006 ~ 0.049)、较低波动变化(0.049 ~ 0.100)、中度波动变化(0.100 ~ 0.185)、较高波动变化(0.185 ~ 0.392)和高波动变化(0.392 ~ 1.000).
其次, 通过地理探测器[28, 29]探究气候要素对不同类型植被NPP空间分异的影响机制.地理探测器是探测空间分异性并揭示其背后驱动力的一组统计学方法, 若某个自变量对某个因变量有重要影响, 那么自变量和因变量的空间分布应该具有相似性是其核心思想.地理探测器包含因子探测、交互探测、生态探测和风险区探测4个模块, 本研究通过因子探测探究西南地区不同类型植被NPP空间分异与影响因子的定量关系;通过交互作用探测探究气候因子间的相互作用对不同类型植被NPP的内在联系;通过生态探测研究气候因子交互作用对不同类型植被NPP变化的显著性;最后通过风险区探测研究不同类型植被对生长气候条件需求的差异性.
因子探测是研究属性Y的空间分异性及影响因素X对属性Y的空间分异性的解释力度, 以q值度量, 计算公式如下.
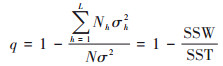
|
(5) |
式中, h为变量Y或因子X的分层, Nh和N分别为层h和全区的单元数, σh2和σ2 分别为层h和全区的Y值的方差, SSW和SST分别为层内方差之和和全区总方差.q的值域为[0, 1], q值越大, 表示X对Y的解释力越大.
为更好地研究真实情景下不同类型植被NPP与气候因子的关系, 本文在探测气候影响因素对不同类型植被NPP的驱动关系时, 将所有气候影响因素均假定为影响植被NPP的主要影响因素, 但考虑到数据冗余的问题, 对此, 基于已有的13种气象因子数据, 使用VIF检验法, 剔除因子间多重共线性问题, 避免数据的冗余.最后选取寒冷指数(X1)、地表温度(X2)、气压(X3)、相对湿度(X4)、日照时数(X5)和温暖指数(X6), 共计6个气候因子作为影响植被NPP空间分异的气候因子.本文使用2000 ~ 2021年植被NPP和6个气候因子的多年平均值作为地理探测器的输入数据.
3 结果与分析 3.1 西南地区不同植被NPP、NPPPre、NPPRes时空变化特征 3.1.1 不同植被NPP、NPPPre、NPPRes时间变化特征如图 2所示, 2000 ~ 2021年, 西南地区整体植被NPP、NPPPre和NPPRes多年平均值分别为507.77、514.96和-7.26 g·(m2·a)-1.不同影响条件下, 其时间变化均呈上升趋势, 上升速率分别为2.09、1.29和0.81 g·(m2·a)-1.在西南地区不同类型植被中, 实际情景下和仅气候变化影响下, 乔木类植被NPP和NPPPre多年平均值均最高, 均为829.06 g·(m2·a)-1, 其次为经济类植被NPP和NPPPre, 分别为708.97 g·(m2·a)-1和708.6 g·(m2·a)-1, 最低的为草本类植被, 均不足300 g·(m2·a)-1.仅人类活动影响下, 不同类型植被NPPRes多年平均值均低于-0.02 g·(m2·a)-1.从变化趋势看, 除乔木类植被NPPRes呈微弱下降趋势外, 其余类型植被NPP、NPPPre和NPPRes均呈波动上升趋势, 其中, 经济类植被NPP、NPPPre和NPPRes上升趋势最为显著, 上升速率分别为5.96、3.09和2.94 g·(m2·a)-1, 其次为灌木类植被;乔木类和草本类植被NPP、NPPPre和NPPRes上升速率较不显著, 上升速率均不足1 g·(m2·a)-1.综上可知, 2000 ~ 2021年西南地区整体植被NPP呈上升趋势, 植被NPP和NPPPre平均值最高均为乔木类植被.从上升趋势看, 除乔木类植被NPPRes出现微弱下降外, 其余类型植被NPP、NPPPre和NPPRes均为波动上升趋势, 其中经济类植被上升趋势最为显著.

|
“草本类植被”和“其他”使用右侧纵坐标轴 图 2 2000 ~ 2021年西南地区各类型植被NPP时间变化趋势 Fig. 2 Temporal variation in NPP of different vegetation types in southwest China from 2000 to 2021 |
如图 3所示, 2000 ~ 2021年西南地区植被NPP多年平均值在空间上主要呈“南高北低”的分布特征, 变化趋势在-65.27 ~ 79 g·(m2·a)-1之间, 变化趋势在空间上主要呈“东高西低”的分布特征.由表 2可知, 在变化趋势面积占比上, 79.13%西南地区植被NPP呈上升趋势, 其中, 呈显著和极显著上升趋势面积占比为37.42%, 集中分布在西南地区中部、东北部和西北小部分地区.此外, 研究区东北地区是经济类植被生长的主要分布区, 西北地区则主要是以草本类植被为主.2000 ~ 2021年, 西南地区经济类植被NPP呈上升趋势面积占比为85.64%, 其中, 呈极显著上升趋势面积占比为63.04%, 集中分布在西南地区东北部;草本类植被中, 85.59%地区呈上升趋势, 其中, 呈极显著上升趋势面积占比为20.30%, 主要分布在西南地区西北部;乔木和灌木类植被NPP呈上升趋势面积占比均低于西南地区平均值, 其中, 乔木类植被NPP呈上升趋势面积占比最低, 仅58.42%.在植被NPP呈下降趋势地区中, 主要以乔木类植被为主, 乔木类植被NPP呈显著和极显著下降面积占比为10.38%, 集中分布在西藏和云南南部, 西南地区其余类型植被NPP呈显著和极显著下降面积占比均低于5%.综上可知, 空间上, 2000 ~ 2021年西南地区植被NPP变化趋势呈“东高西低”的空间分布, 草本类和经济类植被NPP上升趋势面积占比最大, 其中, 经济类植被NPP呈极显著上升趋势的面积占比最大.相较于其他类型植被, 乔木类植被NPP呈显著和极显著下降趋势面积占比最大.

|
图 3 2000 ~ 2021年西南地区植被NPP空间变化 Fig. 3 Spatial variation in vegetation NPP in southwest China from 2000 to 2021 |
|
|
表 2 西南地区不同类型植被NPP变化统计/% Table 2 Statistical results of NPP variation of different vegetation types in southwest China/% |
3.1.3 西南地区植被NPPPre空间变化
由图 4和表 3可知, 气候变化影响下, 西南地区植被NPPPre多年平均值在空间上呈“南高北低”的分布格局, 变化趋势在-54.36 ~ 26.18 g·(m2·a)-1, 其中, 西南地区76.13%植被NPPPre呈上升趋势, 显著和极显著上升趋势面积占39.76%, 植被显著上升区主要集中分布在西南地区中部的东北和西南部.气候变化影响下, 西南地区植被NPPPre同样存在显著下降现象, 超20%西南地区植被NPPPre呈下降趋势, 其中, 显著下降区主要分布在西藏和云南南部.气候变化影响下, 西南地区经济类植被NPPPre上升趋势面积占比最大, 2000 ~ 2021年, 西南地区经济类植被NPPPre呈上升趋势面积占比为85.04%, 其中, 呈显著和极显著上升趋势面积占比为58.49%, 主要分布在西南地区东北部;草本类植被NPPPre是气候变化影响下上升趋势面积占比排第二的植被类型, 超80%的草本类植被NPPPre呈上升趋势, 呈显著和极显著上升趋势面积占比为43.74%, 集中分布在西藏和四川北部;乔木和灌木类植被NPPPre在气候变化影响下, 其上升趋势面积同样大于下降趋势面积, 但乔木类植被NPPPre呈上升趋势面积占比相对较低, 呈显著和极显著上升趋势面积占比仅为23.40%.气候变化影响下, 乔木类植被NPPPre呈显著和极显著下降趋势面积占比为10.10%, 主要分布在西藏和云南南部;其余类型植被NPPPre呈显著和极显著下降面积占比均低于6.00%.综上可知, 气候变化影响下, 西南地区不同类型植被NPPPre呈上升趋势的面积均大于呈下降趋势的面积, 经济类植被NPP呈上升趋势面积占比最大, 主要分布在四川东部, 其次为草本类植被, 主要分布在西藏和四川北部.

|
图 4 2000 ~ 2021年气候变化影响下西南地区植被NPP空间变化 Fig. 4 Spatial variation in vegetation NPP under the influence of climate change in the southwest China from 2000 to 2021 |
|
|
表 3 气候变化影响下西南地区不同类型植被NPP变化统计/% Table 3 Statistical results of NPP variation of different vegetation types under the influence of climate change in southwest China/% |
3.1.4 西南地区植被NPPRes空间变化
由图 5和表 4可知, 人类活动影响下, 西南地区植被NPPRes多年平均值只在西藏地区存在相对显著的区别, 而在其余地区其植被NPPRes多年平均值均主要在0 g·(m2·a)-1上下浮动.西南地区总体植被NPPRes变化趋势在-49.77 ~ 31.03 g·(m2·a)-1, 其中, 70.68%的西南地区植被NPPRes呈上升趋势, 呈显著和极显著上升趋势面积占17.58%, 植被NPPRes显著上升区主要集中分布在四川东部.人类影响下, 西南地区部分地区植被NPPRes同样存在一定的下降现象, 其中, 呈显著和极显著下降趋势占0.41%, 主要分布在西藏和云南南部.人类活动影响下, 西南地区经济类植被NPPRes呈上升趋势面积占比最大, 83.04%地区的经济类植被呈上升趋势, 其中, 呈显著和极显著上升趋势面积占比为40.85%, 主要分布在四川东部;乔木、灌木和草本类植被NPPRes呈上升趋势面积占比均大于49%, 其中, 草本类植被NPPRes呈上升趋势面积占比为71.63%, 乔木类植被NPPRes呈上升趋势面积占比仅为49.15%, 但灌木类植被NPPRes呈显著和极显著上升趋势面积占比高于乔木和草本类植被, 为10.48%.综上可知, 人类活动影响下, 西南地区植被NPPRes以不显著上升趋势为主.植被类型中, 经济类植被NPPRes在人类活动下呈上升趋势面积占比最大, 主要分布在四川东部.乔木类植被NPPRes在人类活动影响下同样存在一定下降区域, 主要分布在西藏和四川南部.

|
图 5 2000 ~ 2021年人类活动影响下西南地区植被NPP空间变化 Fig. 5 Spatial variation in vegetation NPP under the influence of human activities in southwest China from 2000 to 2021 |
|
|
表 4 人类活动影响下西南地区不同类型植被NPP变化统计/% Table 4 Statistical results of NPP variation of different vegetation types under the influence of human activities in southwest China |
3.2 不同类型植被生长稳定性分析
如图 6所示, 空间上, 2000 ~ 2021年西南地区植被NPP波动变化呈“南低北高”的空间分布格局, 各类型植被NPP变化主要表现为较低波动和中波动变化, 各类植被NPP呈较低波动变化和中波动变化的占比之和均超过85%.波动系数平均值以乔木类(0.101)、灌木类(0.105)、草本类(0.110)和经济类(0.114)的顺序依次升高.乔木类和灌木类植被NPP主要呈较低波动变化, 二者呈较低波动变化的占比分别为54.05%和52.87%, 其中, 乔木类植被NPP呈较低波动变化主要分布在西藏和云南南部, 而灌木类植被NPP呈较低波动变化主要分布在云南和贵州地区.草本类和经济类植被NPP主要为中波动变化, 其植被NPP呈中波动变化百分比分别为53.48%和44.31%, 其中草本类植被NPP呈中波动变化主要分布在西藏中部, 经济类植被NPP呈中波动变化则主要分布在四川东部, 但分布在四川东部的经济类植被NPP存在一定的较高波动变化, 占比为8.22%.综上可知, 西南地区植被NPP波动变化呈“南低北高”的空间分布格局, 整体表现为较低波动和中波动变化, 其中乔木类和灌木类植被NPP变化以较低波动变化为主, 草本类和经济类植被NPP变化以中波动变化为主.

|
图 6 西南地区植被NPP稳定性空间分布 Fig. 6 Spatial distribution of the stability of the vegetation NPP in southwest China |
如图 7所示, 地表温度和相对湿度对西南地区植被NPP空间分异的解释力最高, 其q值均为0.62, 其次为气压和温暖指数, q值分别为0.57和0.55, 最低为寒冷指数, q值仅为0.31.在不同类型植被中, 乔木类植被NPP空间分异受气候因子影响最低, 气候因子对乔木类植被NPP空间分异的解释力均值仅为0.18, 其中日照时数q值最低, 仅为0.03;相比于乔木类植被NPP, 经济类植被NPP空间分异与气候因子存在更高的相关性, 气候因子对经济类植被NPP空间分异的解释力均值达0.33, 其中, 日照时数和寒冷指数对经济类植被NPP空间分异的解释力最大, 其q值分别为0.43和0.41, 相对湿度对经济类植被NPP的解释力最低, 其q值仅为0.14;气候因子对灌木和草本类植被NPP整体空间分异的解释力相对较高, q值均值分别为0.51和0.64, 其中, 地表温度、温暖指数、气压和相对湿度对灌木类植被NPP空间分异的影响较大, 而在草本类植被中, 日照时数的影响力同样不可忽视.综上可知, 地表温度、气压、相对湿度、日照时数和温暖指数对西南地区植被NPP空间分异的影响较大, 而在不同类型植被中, 气候因子对植被NPP空间分异的解释力度存在显著差异, 其中, 对草本类植被NPP空间分异呈现较高的解释力, 而对乔木类植被NPP空间分异的解释力相对较低.

|
图 7 因子探测结果 Fig. 7 Results of factor detection |
如图 8所示, 从交互作用结果看, 西南地区气候因子的双因子作用q值均大于单因子q值, 气候因子交互作用结果均表现为双因子增强或非线性增强, 其中, 相对湿度∩地表温度和相对湿度∩日照时数的q值最高, 均为0.75.由此可知, 相对湿度和地表温度及日照时数的交互作用是影响西南地区植被生长的主要气候因素.在不同类型植被中, 乔木类植被NPP受相对湿度∩日照时数的影响最大, 其q值为0.46;灌木类植被NPP受地表温度∩相对湿度、地表温度∩日照时数和日照时数∩气压的影响最大, 其q值均为0.74;草本类植被NPP受日照时数∩温暖指数的影响最大, 交互后q值达0.86;经济类植被NPP受气压∩日照时数的影响最大, 交互后q值为0.54.由此可知, 不同类型植被NPP受气候因子交互影响存在显著的异质性, 其中, 日照时数与不同气候因子的交互作用是影响各类型植被生长的最大因素;如图 8所示, 从交互作用提升效果看, 日照时数∩温暖指数对西南地区植被NPP分异的影响力提升最大, 达30.91%.在不同类型植被中, 交互作用对乔木类植被NPP空间分异的影响力提升最为显著, 其中, 相对湿度与其他气候因子交互效果最为显著, 除相对湿度∩气压的影响力提升54.55%外, 相对湿度与其他气候因子交互影响力提升均超过90%.综上可知, 相对湿度∩地表温度和相对湿度∩日照时数是影响西南地区植被生长的主要因素, 日照时数∩温暖指数的交互作用对西南地区植被NPP影响力提升最大, 而在各类型植被中, 日照时数与不同气候因子的交互作用是影响植被生长的共同因素, 而相对湿度与其他气候影响因子的交互作用对乔木类植被NPP影响力提升最大.

|
左下角表示交互探测结果, 右上角表示提升率 图 8 交互探测结果和交互作用探测q值相较于因子探测q值提升率 Fig. 8 Results of interaction detection and growth rate of q value of interaction detection compared with factor detection |
如图 9所示, 在西南地区整体生态探测中, 除气压和相对湿度、日照时数和温度指数、寒冷指数和不同气候因子组合间对植被NPP的影响均存在显著的差异性外, 其余因子组合对植被NPP空间分异均不存在显著差异的影响, 这表明了寒冷指数对西南地区植被生长影响不同于其他气候影响因子, 这与因子探测中, 寒冷指数对植被NPP空间分异的解释力显著低于其他气候影响因子的结果存在高度相关性.在西南地区不同类型植被生态探测中, 乔木和经济类植被生态探测结果存在高度一致性, 除经济类植被中的寒冷指数与日照时数间存在显著差异性外, 在乔木和经济类植被中, 地表温度与其他气候因子间均无显著差异性.灌木和草本类植被生态探测结果存在高度一致性, 除寒冷指数与日照时数在灌木类植被中表现为存在显著差异性外, 其余情况均为存在显著差异性, 剩余气候因子组合间几乎表现为无显著差异性, 结合因子探测结果可知, 气候因子对乔木和经济类植被NPP空间分异的影响呈高度相似性, 对灌木和草本类植被NPP空间分异的影响呈高度相似性.综上可知, 寒冷指数与其他气候因子组合对西南地区植被生长的空间分异存在显著差异, 主要发生在灌木和草本类植被中, 在乔木和经济类植被中则相反.

|
图 9 因子生态探测结果 Fig. 9 Results of ecological detection |
西南地区及不同类型植被生长最佳条件如表 5所示, 地表温度和相对湿度是西南地区植被NPP空间分异影响最大的两个因子, 当地表温度在21.03 ~ 28.49℃时, 植被NPPmax可达1 162.6 g·(m2·a)-1, 而当相对湿度达最佳值域67.05% ~ 74.68%时, 植被NPPmax为892.83 g·(m2·a)-1.气压、日照时数和温暖指数对西南地区植被NPP空间分异的影响同样较大, 当气压在851.41 ~ 917.11 hPa时, 植被NPPmax可达1 033.63 g·(m2·a)-1, 当温暖指数在106.46 ~ 167.2间时, 植被NPPmax可达西南地区最高值1 226.27 g·(m2·a)-1, 由此可知, 适宜的地温与有效的积温是植被NPP上升的一大重要原因, 在此基础上, 控制植被生长环境的湿度, 可保持植被生长的持续性.
|
|
表 5 风险区探测结果1) Table 5 Results of risk detection |
在西南地区不同类型植被NPP风险区探测中, 不同类型植被对环境条件的要求存在显著的异同, 差异性表现在:寒冷指数只在草本类植被中表现出较大差异, 草本类植被中, 寒冷指数的最适区域为-97.82 ~ -84.07, 而在其他类型植被中需求度均低于-122.63;气压在乔木、灌木和草本类植被中最适值域均为851.41 ~ 917.11 hPa, 而在经济类植被中为777.49 ~ 851.41 hPa;相对湿度除在经济类植被中最适值域为41.49 % ~ 49.11%外, 其余植被类型均要求相对湿度大于60%;在日照需求方面, 大于2 000 h的日照是乔木和经济类植被的最佳条件, 而灌木和草本类植被对日照的需求相对较低.相同性表现在:在地表温度和温暖指数方面, 各类型植被的需求呈高度一致性, 均为21.03 ~ 28.49℃和106.46 ~ 167.2, 当不同类型植被在地表温度和温暖指数达到相同的最适区间时, 均会达到不同的植被NPP峰值.综上可知, 地表温度和温暖指数对西南地区植被NPP积累的影响最大, 且不同类型植被对地表温度和温暖指数的需求相同, 而寒冷指数、气压、相对湿度和日照时数则在不同类型植被中存在一定差异性.
4 讨论 4.1 西南地区不同类型植被NPP时空演变差异从时间上看, 2000 ~ 2021年, 西南地区整体植被NPP呈上升趋势, 气候变化和人类活动影响下, 各类型植被NPP均主要呈上升趋势, 这与已有研究一致[12, 13, 30].2000 ~ 2021年, 西南地区年平均温度为9.3℃, 年累计降水均值为767.49 mm, 比全国年累计降水均值662.85 mm高出104.64 mm, 大量的降水导致了西南地区常年相对湿度维持在32.21% ~ 84.98%, 加上西南地区地处我国中低纬度地区, 充足的光照为植被的生长提供了稳定的能量来源.此外, 自2000年来, 在人类有计划的干预下, 我国针对西南地区各项生态问题, 实施了退耕还林、西南地区石漠化综合治理和长江防护林工程等生态工程, 同样为西南地区的生态恢复与发展做出了巨大贡献[31, 32].
从空间上看, 乔木类植被下降区主要集中在西藏和云南南部, 西藏和云南南部虽然分别为我国藏东南高原边缘森林生态功能区和川滇森林及生物多样性功能区, 但由于成立时间均在2010年之后, 创立时间较短, 加之林业活动和农业扩张等人类活动的影响[33], 导致西南地区2001 ~ 2019年间森林损失面积达375.27万hm2, 这同样是乔木类植被NPP在人类活动影响下呈下降趋势的一大原因.经济类植被NPP作为上升趋势最显著的植被类型, 主要分布在四川盆地, 四川盆地本身得天独厚的地理气候条件, 其土壤本身富含多种有利于植被生长氮磷等矿物质[34.35], 加之我国经济的不断快速发展, 四川盆地土地资源得到不断开发, 农业基础得以不断完善, 四川盆地农业发展蒸蒸日上, 这与已有研究的结果一致[30].
4.2 西南地区植被生长波动变化与影响因素间的潜在关系西南地区不同类型植被中, 经济类植被NPP变化波动最高, 其次是草本和灌木类植被, 乔木类植被NPP变化波动最低.这与植被的演替密切相关, 随着植物群落的演替, 植物群落发展由低级到高级, 由简单到复杂, 植被群落的稳定性不断提高, 而经济类植被虽然不作为植被自然演替的结果, 但其作为人类在衣、食、住和医药等方面息息相关的植被类型, 受人为调控影响较大, 对此波动变化相较自然演替形成的群落其波动变化最大, 这与已有研究结论相似[36].在本实验中可发现, 除经济类植被外, 植被NPP的波动大小与气候因子对其空间影响存在密切联系, 波动变化越大, 气候因子对植被生长的空间分异的解释力就相对越大.乔木群落因其稳定的生态结构使其受气候影响较低, 而草本类植被因其不稳定的生态群落而受气候影响调控较大, 但经济类植被因其受人文因素的影响较大, 其植被NPP波动变化大小与气候对其影响的大小成反比.
交互作用结果显示, 双因子交互作用会加强气候影响因素对植被NPP空间分异的影响强度, 这与已有研究结果一致[37, 38], 其中, 地表温度和相对湿度的交互作用是西南地区植被生长空间分异的主要影响因素, 而气压与日照时数的交互作用对西南地区植被NPP影响提升最大.从不同类型植被与气候因子间的关系看, 乔木类植被NPP生长变化主要受相对湿度、地表温度、寒冷指数和温暖指数影响, 同时相对湿度与其他气候影响因子的交互作用对乔木类植被NPP影响提升巨大, 这与已有研究结论较一致[30, 39 ~ 41].乔木类植被作为最高等级植被, 其完善的生理结构使其受气候影响相对较低.灌木类植被NPP生长变化主要受地表温度、气压、相对湿度和温暖指数调控, 而草本类植被在此基础上, 同样受日照时数影响较大, 这与已有研究的结果相一致[30, 42 ~ 44].温度、水分和能量来源是植被生长的三大重要影响因素, 灌木和草本类植被作为中低等级的植被群落, 受其影响变化较为显著, 其中草本类植被作为最低等级的植被群落, 因其植被群落功能结构最为简单, 受光照的影响最为敏感, 对此, 光照的长短, 更是决定着草本类植被生长发育和繁殖的重要因素.同时, 日照时数也是影响经济类植被NPP最大的气候因子, 这与已有研究的结果一致[12, 13, 30, 45], 经济类植被作为为人类提供经济价值的植被, 其合成有机物效率高是其一大特点, 而充足的光照是其进行光合反应的一大重要保障, 加之经济类植被本身作为人类林业活动和农业劳动的主要对象, 受人类活动影响巨大, 对此, 气候因素的影响就相对较低.
5 结论(1)时间上, 2000 ~ 2021年西南地区植被NPP呈上升趋势, 空间上, 西南地区植被NPP呈“东高西低”的分布格局.不同类型植被中, 除人类活动影响下的乔木类植被NPP存在微弱下降的现象外, 其余类型植被NPP均为波动上升趋势, 其中不同影响条件下经济类植被NPP上升趋势均最为显著, 聚集分布在四川省东部.
(2)西南地区植被NPP受主要地表温度和相对湿度的影响, 在不同类型植被中, 不同类型植被对地表温度和温暖指数的需求一致, 当地表温度为21.03 ~ 28.49℃, 温暖指数为106.46 ~ 167.2时, 不同类型植被NPP均会达到峰值, 但对寒冷指数、气压、相对湿度和日照时数存在一定的需求差异.
(3)交互作用结果显示, 日照时数∩温暖指数对西南地区植被NPP影响提升最大, 而在不同类型植被中, 乔木类植被受交互作用影响最为显著, 其中相对湿度和其他气候因子的交互作用对乔木类植被NPP影响提升最大.
(4)植被NPP波动大小与气候因子对植被NPP空间分异的解释力间存在密切联系, 自然条件下, 植被群落演替等级越高, 波动变化幅度越低, 气候因子对植被NPP空间分异解释力也越小, 而经济类植被存在截然相反的结果.
| [1] | Liu Y, Ding Z, Chen Y N, et al. Restored vegetation is more resistant to extreme drought events than natural vegetation in Southwest China[J]. Science of the Total Environment, 2023, 866. DOI:10.1016/j.scitotenv.2022.161250 |
| [2] |
许小明, 张晓萍, 何亮, 等. 黄土丘陵区不同恢复植被类型的固碳特征[J]. 环境科学, 2022, 43(11): 5263-5273. Xu X M, Zhang X P, He L, et al. Carbon sequestration characteristics of different restored vegetation types in Loess Hilly region[J]. Environmental Science, 2022, 43(11): 5263-5273. |
| [3] |
张雄一, 邵全琴, 宁佳, 等. 三北工程区植被恢复对土壤风蚀的影响及植被恢复潜力研究[J]. 地球信息科学学报, 2022, 24(11): 2153-2170. Zhang X Y, Shao Q Q, Ning J, et al. Effect of vegetation restoration on soil wind erosion and vegetation restoration potential in the three-north afforestation program[J]. Journal of Geoinformation Science, 2022, 24(11): 2153-2170. |
| [4] | Shen J L, Zhang H Y, Zhao Y K, et al. An examination of the mitigation effect of vegetation restoration on regional water poverty: Based on panel data analysis of 9 provinces in the Yellow River basin of China from 1999 to 2019[J]. Ecological Indicators, 2023, 146. DOI:10.1016/j.ecolind.2023.109860 |
| [5] | Gao J, Ji Y H, Zhang X. Net primary productivity exhibits a stronger climatic response in planted versus natural forests[J]. Forest Ecology and Management, 2023, 529. DOI:10.1016/j.foreco.2022.120722 |
| [6] | Sun H Z, Chen Y B, Xiong J N, et al. Relationships between climate change, phenology, edaphic factors, and net primary productivity across the Tibetan Plateau[J]. International Journal of Applied Earth Observation and Geoinformation, 2022, 107. DOI:10.1016/j.jag.2022.102708 |
| [7] | Liu X, Wang P, Song H, et al. Determinants of net primary productivity: Low-carbon development from the perspective of carbon sequestration[J]. Technological Forecasting and Social Change, 2021, 172. DOI:10.1016/j.techfore.2021.121006 |
| [8] | Li C H, Zhou M, Dou T B, et al. Convergence of global hydrothermal pattern leads to an increase in vegetation net primary productivity[J]. Ecological Indicators, 2021, 132. DOI:10.1016/j.ecolind.2021.108282 |
| [9] | Bejagam V, Sharma A. Impact of climatic changes and anthropogenic activities on ecosystem net primary productivity in India during 2001-2019[J]. Ecological Informatics, 2022, 70. DOI:10.1016/j.ecoinf.2022.101732 |
| [10] |
石智宇, 王雅婷, 赵清, 等. 2001-2020年中国植被净初级生产力时空变化及其驱动机制分析[J]. 生态环境学报, 2022, 31(11): 2111-2123. Shi Z Y, Wang Y T, Zhao Q, et al. The spatiotemporal changes of NPP and its driving mechanisms in China from 2001 to 2020[J]. Ecology and Environmental Sciences, 2022, 31(11): 2111-2123. |
| [11] | Yu D S, Li Y H, Yin B L, et al. Spatiotemporal variation of net primary productivity and its response to drought in Inner Mongolian desert steppe[J]. Global Ecology and Conservation, 2022, 33. DOI:10.1016/j.gecco.2021.e01991 |
| [12] |
徐勇, 黄雯婷, 郑志威, 等. 基于空间尺度效应的西南地区植被NPP影响因子探测[J]. 环境科学, 2023, 44(2): 900-911. Xu Y, Huang W T, Zheng Z W, et al. Detecting influencing factor of vegetation NPP in southwest China based on spatial scale effect[J]. Environmental Science, 2023, 44(2): 900-911. |
| [13] |
徐勇, 郑志威, 戴强玉, 等. 顾及时滞效应的西南地区植被NPP变化归因分析[J]. 农业工程学报, 2022, 38(9): 297-305. Xu Y, Zheng Z W, Dai Q Y, et al. Attribution analysis of vegetation NPP variation in Southwest China considering time-lag effects[J]. Transactions of the Chinese Society of Agricultural Engineering, 2022, 38(9): 297-305. |
| [14] | Linger E, Hogan J A, Cao M, et al. Precipitation influences on the net primary productivity of a tropical seasonal rainforest in Southwest China: A 9-year case study[J]. Forest Ecology and Management, 2020, 467. DOI:10.1016/j.foreco.2020.118153 |
| [15] |
茆杨, 蒋勇军, 张彩云, 等. 近20年来西南地区植被净初级生产力时空变化与影响因素及其对生态工程响应[J]. 生态学报, 2022, 42(7): 2878-2890. Mao Y, Jiang Y J, Zhang C Y, et al. Spatio-temporal changes and influencing factors of vegetation net primary productivity in southwest China in the past 20 years and its response to ecological engineering[J]. Acta Ecologica Sinica, 2022, 42(7): 2878-2890. |
| [16] | Yang F, Zhong Y J, Han G Z, et al. Effect of different vegetation restoration on soil organic carbon dynamics and fractions in the Rainy Zone of Western China[J]. Journal of Environmental Management, 2023, 331. DOI:10.1016/j.jenvman.2023.117296 |
| [17] |
赵姣, 马静, 朱燕峰, 等. 植被类型对黄土高原露采矿山复垦土壤碳循环功能基因的影响[J]. 环境科学, 2023, 44(6): 3386-3395. Zhao J, Ma J, Zhu Y F, et al. Effects of vegetation types on carbon cycle functional genes in reclaimed soil from open pit mines in the loess plateau[J]. Environmental Science, 2023, 44(6): 3386-3395. |
| [18] |
耿庆玲, 陈晓青, 赫晓慧, 等. 中国不同植被类型归一化植被指数对气候变化和人类活动的响应[J]. 生态学报, 2022, 42(9): 3557-3568. Geng Q L, Chen X Q, He X H, et al. Vegetation dynamics and its response to climate change and human activities based on different vegetation types in China[J]. Acta Ecologica Sinica, 2022, 42(9): 3557-3568. |
| [19] |
卢乔倩, 江涛, 柳丹丽, 等. 中国不同植被覆盖类型NDVI对气温和降水的响应特征[J]. 生态环境学报, 2020, 29(1): 23-34. Lu Q Q, Jiang T, Liu D L, et al. The response characteristics of NDVI with different vegetation cover types to temperature and precipitation in China[J]. Ecology and Environmental Sciences, 2020, 29(1): 23-34. |
| [20] |
陈琪, 杨九艳, 闫瑞玲, 等. 荒漠区不同植被类型NDVI变化特征分析——以内蒙古阿拉善盟为例[J]. 中国草地学报, 2022, 44(3): 17-28. Chen Q, Yang J Y, Yan R L, et al. Analysis of NDVI variation characteristics of different vegetation types in desert areas——a case study of Alxa League, Inner Mongolia[J]. Chinese Journal of Grassland, 2022, 44(3): 17-28. |
| [21] |
缪利, 陆晴, 刘根林, 等. 1999-2019年青藏高原不同植被类型NDVI时空演变特征及其对气候因子的响应[J]. 水土保持研究, 2023, 30(1): 97-105. Miao L, Lu Q, Liu G L, et al. Spatiotemporal evolution characteristics of NDVI of different vegetation types and its response to climate factors on the Qinghai-Tibet plateau from 1999 to 2019[J]. Research of Soil and Water Conservation, 2023, 30(1): 97-105. |
| [22] | Sen P K. Estimates of the regression coefficient based on Kendall's Tau[J]. Journal of the American Statistical Association, 1968, 63(324): 1379-1389. DOI:10.1080/01621459.1968.10480934 |
| [23] |
杨靖, 戴君虎, 姚华荣, 等. 1992—2020年横断山区植被分布与植被活动变化[J]. 地理学报, 2022, 77(11): 2787-2802. Yang J, Dai J H, Yao H R, et al. Vegetation distribution and vegetation activity changes in the Hengduan Mountains from 1992 to 2020[J]. Acta Geographica Sinica, 2022, 77(11): 2787-2802. |
| [24] |
徐勇, 李欣怡, 黄雯婷, 等. 2000—2020年中国典型经济区PM2.5时空变化及其与植被景观格局的关系[J]. 环境科学, 2023, 44(4): 1852-1864. Xu Y, Li X Y, Huang W T, et al. Spatio-temporal variation in PM2.5 concentration and its relationship with vegetation landscape patterns in typical economic zones in China from 2000 to 2020[J]. Environmental Science, 2023, 44(4): 1852-1864. |
| [25] |
吴秦豫, 姚喜军, 梁洁, 等. 鄂尔多斯市煤矿区植被覆盖改善和退化效应的时空强度[J]. 干旱区资源与环境, 2022, 36(8): 101-109. Wu Q Y, Yao X J, Liang J, et al. Spatial and temporal intensity of vegetation cover improvement and degradation in coal mining areas of Erdos city[J]. Journal of Arid Land Resources and Environment, 2022, 36(8): 101-109. |
| [26] |
李婷婷, 郭增长, 马超. 中国第二、三级阶梯地形过渡带山前植被物候时空变化探析[J]. 地理研究, 2022, 41(11): 3000-3020. Li T T, Guo Z Z, Ma C, et al. Spatiotemporal changes of piedmont phenology in the transitional zone between the second and third steps, China[J]. Geographical Research, 2022, 41(11): 3000-3020. |
| [27] |
徐勇, 郑志威, 郭振东, 等. 2000 ~ 2020年长江流域植被NDVI动态变化及影响因素探测[J]. 环境科学, 2022, 43(7): 3730-3740. Xu Y, Zheng Z W, Guo Z D, et al. Dynamic variation in vegetation cover and its influencing factor detection in the Yangtze River basin from 2000 to 2020[J]. Environmental Science, 2022, 43(7): 3730-3740. |
| [28] |
王劲峰, 徐成东. 地理探测器: 原理与展望[J]. 地理学报, 2017, 72(1): 116-134. Wang J F, Xu C D. Geodetector: Principle and prospective[J]. Acta Geographica Sinica, 2017, 72(1): 116-134. |
| [29] |
徐勇, 郭振东, 郑志威, 等. 运用地理探测器研究京津冀城市群PM2.5浓度变化及影响因素[J]. 环境科学研究, 2023, 36(4): 649-659. Xu Y, Guo Z D, Zheng Z W, et al. Study of the PM2.5 concentration variation and its influencing factors in the BeijingTianjin-Hebei urban agglomeration using geo-detector[J]. Research of Environmental Sciences, 2023, 36(4): 649-659. |
| [30] |
徐勇, 黄海艳, 戴强玉, 等. 西南地区陆地植被生态系统NPP时空演变及驱动力分析[J]. 环境科学, 2023, 44(5): 2704-2714. Xu Y, Huang H Y, Dai Q Y, et al. Spatial-temporal variation of net primary productivity in terrestrial vegetation ecosystems and its driving forces in southwest China[J]. Environmental Science, 2023, 44(5): 2704-2714. |
| [31] |
张杰. 基于时序遥感数据的西南喀斯特地区石漠化信息提取研究[D]. 北京: 中国地质大学(北京), 2021. Zhang J. Extraction of rocky desertification information in karst area of southwest China based on time-series remote sensing data[D]. Beijing: China University of Geosciences(Beijing), 2021. |
| [32] |
叶鑫, 顾羊羊, 张琨, 等. 西南喀斯特地区石漠化治理现状分析与对策研究——以贵州省黔西南州为例[J]. 环境保护, 2020, 48(22): 30-34. Ye X, Gu Y Y, Zhang K, et al. Current Situation analysis and countermeasures of rocky desertification control in southwest karst region: a case Study of Qianxinan prefecture in Guizhou province[J]. Environmental Protection, 2020, 48(22): 30-34. |
| [33] |
王淑静, 赖佩玉, 郝斌飞, 等. 西南地区2001 ~ 2019年森林损失特征遥感监测与时空分析[J]. 遥感技术与应用, 2021, 36(3): 552-563. Wang S J, Lai P Y, Hao B F, et al. Remote sensing monitoring and spatio-temporal pattern of deforestation in southwest China from 2001 to 2019[J]. Remote Sensing Technology and Application, 2021, 36(3): 552-563. |
| [34] |
家伟, 聂富育, 杨万勤, 等. 四川盆地西缘4种人工林土壤团聚体碳氮磷生态化学计量学特征[J]. 应用与环境生物学报, 2018, 24(1): 112-118. Jia W, Nie F Y, Yang W Q, et al. Ecological stoichiometry of soil carbon, nitrogen, and phosphorus within soil aggregates of four plantations in the western edge of Sichuan basin[J]. Chinese Journal of Applied and Environmental Biology, 2018, 24(1): 112-118. |
| [35] |
袁余洋, 刘属灵, 雒昆利, 等. 四川盆地典型农业区土壤中铜、钴、钼和锌的空间分异及其影响因素[J]. 高校地质学报, 2022, 28(4): 506-515. Yuan Y Y, Liu S L, Luo K L, et al. Study on the spatial variation and driving factors of copper, cobalt, molybdenum and zinc in Top soil of typical agricultural region in Sichuan Basin[J]. Geological Journal of China Universities, 2022, 28(4): 506-515. |
| [36] |
王婧姝, 毕如田, 贺鹏, 等. 气候变化下黄土高原植被生长期NDVI动态变化特征[J]. 生态学杂志, 2023, 42(1): 67-76. Wang J S, Bi R T, He P, et al. Variations of NDVI during main growing season in the Loess Plateau under climate change[J]. Chinese Journal of Ecology, 2023, 42(1): 67-76. |
| [37] | Shi S H, Wang X L, Hu Z R, et al. Geographic detector-based quantitative assessment enhances attribution analysis of climate and topography factors to vegetation variation for spatial heterogeneity and coupling[J]. Global Ecology and Conservation, 2023, 42. DOI:10.1016/j.gecco.2023.e02398 |
| [38] | Peng W F, Kuang T T, Tao S. Quantifying influences of natural factors on vegetation NDVI changes based on geographical detector in Sichuan, western China[J]. Journal of Cleaner Production, 2019, 233: 353-367. DOI:10.1016/j.jclepro.2019.05.355 |
| [39] |
余舒含. 中国森林、草地生态系统生物量分配格局及影响因素分析[D]. 拉萨: 西藏大学, 2021. Yu S H. Biomass and allocation of Chinese forest and grassland ecosystem[D]. Lasa: Tibet University, 2021. |
| [40] |
李树强, 高丽, 李桦, 等. 陕西省国有林场森林植被固碳量时空演变及其驱动因素研究[J]. 林业经济, 2022, 44(11): 58-75. Li S Q, Gao L, Li H, et al. Study on spatial-temporal evolution and driving factors of forest vegetation carbon sequestration in state forest farms in Shaanxi province[J]. Forestry Economics, 2022, 44(11): 58-75. |
| [41] | Wang C L, Jiang Q O, Engel B, et al. Analysis on net primary productivity change of forests and its multi-level driving mechanism -A case study in Changbai Mountains in Northeast China[J]. Technological Forecasting and Social Change, 2020, 153. DOI:10.1016/j.techfore.2020.119939 |
| [42] |
刘杰, 汲玉河, 周广胜, 等. 2000—2020年青藏高原植被净初级生产力时空变化及其气候驱动作用[J]. 应用生态学报, 2022, 33(6): 1533-1538. Liu J, Ji Y H, Zhou G S, et al. Temporal and spatial variations of net primary productivity(NPP)and its climate driving effect in the Qinghai-Tibet Plateau, China from 2000 to 2020[J]. Chinese Journal of Applied Ecology, 2022, 33(6): 1533-1538. |
| [43] |
胡志强, 宋孝玉, 覃琳, 等. 2001—2020年内陆干旱草原县域尺度植被NDVI时空变化特征及其驱动因素[J]. 水土保持通报, 2022, 42(5): 213-221. Hu Z Q, Song X Y, Qin L, et al. Spatio-temporal variation characteristics and its driving factors of NDVI at county scale for an inland arid grassland during 2001-2020[J]. Bulletin of Soil and Water Conservation, 2022, 42(5): 213-221. |
| [44] | Ma R, Shen X J, Zhang J Q, et al. Variation of vegetation autumn phenology and its climatic drivers in temperate grasslands of China[J]. International Journal of Applied Earth Observation and Geoinformation, 2022, 114, doi: 1016/j.jag.2022.103064. |
| [45] | Yang S J, Song S, Li F D, et al. Evaluating farmland ecosystem resilience and its obstacle factors in Ethiopia[J]. Ecological Indicators, 2023, 146. DOI:10.1016/j.ecolind.2023.109900 |