 2023, Vol. 44
2023, Vol. 44

2. 中国环境科学研究院国家环境保护饮用水水源地保护重点实验室, 北京 100012;
3. 中国环境科学研究院国家环境保护洞庭湖科学观测研究站, 岳阳 414000;
4. 华东师范大学生态与环境科学学院, 上海 200241;
5. 湖南省洞庭湖生态环境监测中心, 岳阳 414000
 , YIN Xue-yan1,2,3 , WANG Xing1,2,3
, YIN Xue-yan1,2,3 , WANG Xing1,2,3 , HUANG Min-sheng4 , HUANG Dai-zhong5 , WANG En-rui1,2 , ZHANG Yun-yu4
, HUANG Min-sheng4 , HUANG Dai-zhong5 , WANG En-rui1,2 , ZHANG Yun-yu4
2. State Environmental Protection Key Laboratory of Drinking Water Source Protection, Chinese Research Academy of Environmental Sciences, Beijing 100012, China;
3. State Environmental Protection Scientific Observation and Research Station for Lake Dongting, Chinese Research Academy of Environmental Sciences, Yueyang 414000, China;
4. School of Ecological and Environmental Sciences, East China Normal University, Shanghai 200241, China;
5. Ecological and Environmental Monitoring Center of Dongting Lake of Hunan, Yueyang 414000, China
浮游植物是水生态系统中最重要的初级生产者, 具有细胞个体小、结构简单且对环境的敏感性高, 变化较快等特点[1], 在河流湖泊等水体中与非生物因素, 如氮磷营养盐、高锰酸盐指数、溶解氧和温度等具有相互影响, 同步变化的效应[2, 3].因此, 被作为一种有效的水体生态环境评价方法来分析水质状况[4, 5].
目前, 常用于浮游植物评价水环境的方法主要有两种:基于亲缘关系的系统分类法和基于生境特征划分功能群的方法(FG)[6, 7].如系统分类法是以浮游植物指示种、属和门来归类数据, 各指示种代表水体环境状态, 或借助多样性指数法来进行评价, 如Shannon-Wiener多样性指数可以反映浮游植物群落特征[8], 进而推断出水质情况.而功能群则是以浮游植物的生理特征和适应环境的机制, 将生境相同或相似的种类归为同一功能群, 目前已划分出39个功能类群[9, 10].功能群可以综合反映水深、水力冲刷、水体混合状态、透明度、营养程度、酸碱性和牧食作用等生境[11].在实际应用中, 通过优势功能群所适应的生境类型来判断水体理化和生物环境特征[9, 10].同时, 由于功能群与环境特征相互响应, Padisák等[12]提出了一种基于功能群的集合指数(Q指数)评价法, Q指数对物种的出现、生物量变化和栖息地类型的偏好非常敏感, 其应用不受地域限制.两种方法在水环境质量评价中都有广泛应用, 但不同的水环境类型对评价方法的适用性却有明显差异.Salmaso等[13]研究表明, 功能群对生态属性, 环境耐受性和营养状态的解释度优于浮游植物群落分类单元; 而郭芳等[11]对南亚热带8座水库的浮游植物研究表明, 群落划分法对环境识别的灵敏度高于FG功能群分类法.根据以上分析, 水环境评价中功能群分类法优势要优于群落划分法, 但这两种评价方法却得到不同的结果.因此, 对于方法的适用性及合理性需要针对具体的水环境特征进行对应分析.
有研究表明, 浮游植物的生长和分布受生物因素[14, 15](浮游动物捕食)和非生物因素[16~19](光照、温度、营养盐和水文连通性等)的影响.洞庭湖作为长江中下游距离三峡大坝最近的典型通江湖泊[20], 水系分布较为复杂, 湖区水质及水文状况受长江三口、四水及区间季节性来水携带大量营养物质影响较大[21].且洞庭湖流域以农业为主, 农业面源污染较为严重, 水位波动及降雨径流冲刷改变了湖区污染物浓度[22].这种季节性的水文水质变化对洞庭湖浮游植物生长和分布造成了一定的影响.以往对洞庭湖浮游植物的研究多集中于浮游植物群落时空分布及功能群与理化因子的响应关系[23~26], 关于水文条件对洞庭湖浮游植物影响的报道较少.此外, 对浮游植物群落和功能群仅是单独分析其与环境因子的响应关系, 并未分析两种方法的适用性.
本研究首次将浮游植物种群与FG功能群划分法对洞庭湖浮游植物进行比较分析, 并探究浮游植物群落与功能群的变化特征、优势功能群与适应生境之间的相互关系以及两种方法的适用性, 最后通过Q指数评价法和其他水质评价法进行比较, 以期为洞庭湖水生态管理及水质评价提供科学依据.
1 材料与方法 1.1 研究区域概况洞庭湖(110°40′~113°40′E, 28°30′~30°20′N)位于湖南省北部, 长江中游南岸, 流域面积2 625 km2, 容积1.78×1010m3, 平均水深6.39 m, 丰水期水位和蓄水量分别是枯水期的1.14~1.64倍和17~31倍[27].由西洞庭湖(284 km2)、南洞庭湖(897 km2)和东洞庭湖(1 217 km2)组成, 北部接长江荆江段松滋、太平和藕池三口分流, 南部接纳湘江、资水、沅江和澧水, 东部主要由新墙河、华容河和汨罗江等河流汇入, 经洞庭湖调蓄后由城陵矶汇入长江, 构成一个复杂的吞吐型通江湖泊.
1.2 监测点位布设与采样时间本研究根据洞庭湖实际水文特征、各湖区面积大小和地理位置特征, 全湖共设置11个监测点位(S1~S11).涵盖西洞庭湖区(S1~S3)、南洞庭湖区(S4~S6)和东洞庭湖区(S7~S11), 共计3个区域, 各采样点分布如图 1所示.分别于2019年3月(春季)、6月(夏季)、9月(秋季)和12月(冬季)进行调查采样.

|
图 1 洞庭湖监测点位示意 Fig. 1 Sampling sites in Dongting Lake |
用YSI-EXO多参数水质分析仪(美国维赛YSI professional plus)现场测定水温(WT)、溶解氧(DO)、电导率(Cond)和pH值等参数, 透明度(SD)用塞氏圆盘法测定.总氮(TN)、总磷(TP)、氨氮(NH4+-N)、高锰酸盐指数和叶绿素a(Chl-a)取表层0.5 m的1 L水样带回实验室测定, 各指标参照《水和废水监测分析方法(第四版)》[25]测定, 其中, TN采用碱性过硫酸钾-紫外分光光度计法; TP采用钼酸铵分光光度法; NH4+-N采用纳氏试剂光度法; 高锰酸盐指数采用酸性高锰酸钾法; Chl-a采用丙酮萃取分光光度法.浮游植物取样品取表层0.5 m的1 L, 现场加入鲁哥试剂固定, 实验室静置48 h后, 浓缩至30 mL保存; 在奥林巴斯CX31型显微镜(10×40倍)下进行计数, 参照《中国淡水藻类》[28]进行鉴定.水位和流量数据来源于湖南水文水资源勘测中心(http://slt.hunan.gov.cn/).
1.4 数据处理浮游植物的优势度Y, 采用Y>0.02作为优势种的判别标准.Shannon-Winner多样性指数(H), 可以反映水体浮游植物群落结构的复杂性, 数值越高, 其群落结构越复杂, 水质越好[8], 计算公式如下.
优势种分析(Y):
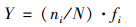
|
Shannon-Winener指数(H):

|
式中, Pi=Ni/N; Ni为第i种藻的个数, N为观察到的总个数, ni为第i种的总个体数, fi为该样品中出现的频率.H指数值分别表示:0~1重污染, 1~2 α重污染, 2~3 β中污染, >3轻或无污染.
Q指数是一种通过浮游植物功能群和其生物量对湖泊、水库营养状况进行评价的生态健康指数[12], 计算公式如下:
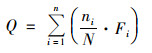
|
式中, n为浮游植物功能群的数量, N为浮游植物的总生物量, ni为第i个功能群的生物量, Fi为第i个功能群的赋值, 范围为0~5, 其中储层更原始的功能群赋值较高, 而原始条件的功能群赋值较低[29].Q指数0~5分别表示:0~1差, 1~2耐受, 2~3中等, 3~4好, 4~5极好.
综合营养状态指数(TLI)是评价湖泊富营养化程度的指标, 参照《湖泊(水库)富营养化评价方法及分级技术规定》[30]中进行计算.TLI指数0~100分别表示: 0~30贫营养, 30~50中营养, 50~60轻度富营养, 60~70中度富营养, 70~100重度富营养.
1.5 数据分析洞庭湖不同季节和湖区环境因子的显著性差异采用单因素方差分析(one-way ANOVA), 借助SPSS 21.0统计软件处理.首先, 基于DCA的分析结果, 当DCA第1轴的特征向量(DCA1)小于3时, 选择RDA分析; 当DCA1介于3~4之间时, 选择RDA和CCA均可; 当DCA1大于4时, 选择CCA分析.
基于浮游植物密度和生物量数据以及VIF(variance inflation factor)小于20的环境因子进行冗余分析(redundancy, RDA), 用来评估环境因子对浮游植物群落和浮游植物功能群的影响, RDA分析及层次分割分析使用rdacca.hp包[31]完成; 使用vegan包中的“envfit”函数对影响浮游植物群落和功能群的环境因子进行显著性检验.最后, 使用corrplot包进行环境因子与TLI指数、Shannon-Wiener指数和Q指数的相关性分析.上述分析过程及可视化均在R(v.4.0.3)中完成.
2 结果与分析 2.1 洞庭湖环境因子变化规律洞庭湖城陵矶水文站显示, 其2019年水位和流量变化趋势基本一致(图 2), 4~6月为涨水期, 7~8月为丰水期, 9~11月为退水期, 12月至次年3月为枯水期.其中, 3月、6月、9月和12月水位分别为24.60、29.57、24.52和20.17 m; 流量分别为10 566、17 641、3 885和2 158 m3·s-1.

|
图 2 洞庭湖城陵矶水文站2019年水位和流量月均值变化 Fig. 2 Monthly mean changes in water level and flow at Dongting Lake Chenglingji hydrological station in 2019 |
洞庭湖不同月份及湖区水体环境因子变化如图 3所示, 单因素方差分析结果表明, 洞庭湖不同月份的水温、DO、Cond、NH4+-N、TN和TN/TP具有显著的时间差异(P<0.05); Cond、高锰酸盐指数、TN和TP具有显著的空间差异(P<0.05).水温变化范围为9.2~30.7℃, 6月(24℃)和9月(27.8℃)明显高于3月(10.5℃)和12月(12.6℃).DO和NH4+-N与水温变化趋势相反, 6月和9月明显低于3月和12月, 变化范围分别为6.44~11.59 mg·L-1和0.02~0.36 mg·L-1.Cond变化范围为12.8~46 μS·cm-1, 12月最高, 6月最低. ρ(TN)和TN/TP变化范围分别为1.04~2.36 mg·L-1和11~57.5, 其中3月均值最高, 6月均值最低.对于空间分布, 东洞庭湖Cond、高锰酸盐指数、TN和TP明显高于西、南洞庭湖.

|
图 3 洞庭湖不同季节和湖区水体理化性质 Fig. 3 Physicochemical properties in different seasons and lakes of Dongting Lake |
调查期间, 共鉴定出浮游植物6门61属.浮游植物群落密度变化范围为(5.29~66.51)×104 ind·L-1, 其中, 9月最高(31.10×104 ind·L-1), 6月次之(31.03×104 ind·L-1), 3月为28.86×104 ind·L-1, 12月最低(21.46×104 ind·L-1)[图 4(a)].东洞庭湖显著高于西、南洞庭湖(P<0.05); 其中, 西、南洞庭湖在3月、6月和12月均以硅藻为主, 9月以蓝藻为主; 而东洞庭湖3月以硅藻为主, 6月以蓝-绿-硅藻为主, 9月以蓝藻为主.此外, 3个湖区在12月隐藻占比均有所升高.从不同时期浮游植物的优势种(表 1)看, 3月和6月直链藻优势度最高, 9月颤藻优势度最高, 12月隐藻优势度最高.

|
图 4 洞庭湖不同季节和湖区浮游植物种群和功能群分布特征 Fig. 4 Distribution characteristics of phytoplankton population and functional groups in different seasons and lakes in Dongting Lake |
|
|
表 1 洞庭湖浮游植物优势种分布1) Table 1 Distribution of dominant species of phytoplankton in Dongting Lake |
2.2.2 浮游植物功能群划分和特征
根据功能群分类方法[10], 共划分23个功能群, 分别为B、C、D、F、G、H1、J、Lo、LM、M、MP、N、P、S1、S2、SN、T、W1、W2、X1、X2、X3和Y, 各功能群代表性种属及生境特征等见表 2.将每个样本中相对生物量>5%的功能群定义为优势功能群, 得到9组优势功能群, 分别是B、D、H1、J、Lo、MP、P、X1和Y.
|
|
表 2 洞庭湖浮游植物功能群划分 Table 2 Functional groups division of phytoplankton in Dongting Lake |
洞庭湖浮游植物功能群在时空分布上的差异主要表现在功能群生物量大小和各功能组组成比例的变化[图 4(b)], 浮游植物总生物量变化范围为0.41~7.78 mg·L-1.其中, 时间分布上, 以6月最高(3.75 mg·L-1), 9月次之(3.42 mg·L-1), 12月为2.58 mg·L-1, 3月最低(1.55 mg·L-1).空间分布上, 东洞庭湖浮游植物生物量最高, 西、南洞庭湖差异性不大(P>0.05), 且生物量均较低.3月以功能群P、MP和D占优, 占比分别为24.95%、19.92%和17.22%.6月以功能群MP、P和J占优, 占比分别为18.04%、13.05%和11.62%.9月以功能群MP和H1占优, 占比分别为48.42%和20.54%.12月以功能群Y、P和MP占优, 占比分别为20.90%、18.26%和18.20%.相对西洞庭湖和南洞庭湖, 功能群H1和Y分别在东洞庭湖的9月和12月占比较高; 功能群P和D在西、南洞庭湖3月、6月和12月占比较高.浮游植物功能群总体演替趋势为: P/MP/D(3月)→MP/P/J(6月)→MP/H1(9月)→Y/P/MP(12月).
2.3 浮游植物种群结构与功能群的驱动因子DCA结果(种群DCA1=1.92, 功能群DCA1=1.55)表明, RDA分析可以合理解释浮游植物种群和功能群驱动因子.RDA分析结果显示(图 5), 环境因子对浮游植物种群和功能群的前两轴解释率分别为71.97%和74.24%.其中, 直链藻(Mel.)和舟形藻(Nav.)与DO呈显著正相关, 与Cond、TP、高锰酸盐指数和WT呈显著负相关; 颤藻(Osc.)、伪鱼腥藻(Pse.)和栅藻(Sce.)均与Cond、TP、高锰酸盐指数和WT呈显著正相关性, 与DO呈显著负相关; 隐藻(Cry.)与WL、WT呈显著负相关.功能群MP、H1和J均与WT、高锰酸盐指数及TP呈显著正相关, 与DO显著负相关, 而功能群P、B和D与之相反; 功能群Y与WL、WT呈显著负相关; 功能群Lo与WL呈显著正相关.

|
带*的环境因子表示“envfit”函数检验P<0.05; 红色箭头表示环境因子, 蓝色箭头分别表示物种和功能群 图 5 洞庭湖不同季节和湖区浮游植物种群和功能群RDA分析 Fig. 5 RDA analysis of phytoplankton population and functional groups in different seasons and lakes in Dongting Lake |
为识别各环境因子对浮游植物种群和功能群重要性影响作用, 通过层次分割结果显示(图 6), 浮游植物种群和功能群受季节性变化的影响显著高于空间性, 浮游植物种群受空间性的影响要高于功能群.显著影响浮游植物种群的环境因子分别为:WT>高锰酸盐指数>DO>Cond>WL>TP, 显著影响浮游植物功能群的环境因子分别为:WT>DO>Cond>高锰酸盐指数>WL>TP, 主要环境因子对浮游植物种群和功能群的独立解释性排序相差不大.

|
1.季节, 2.WT, 3.高锰酸盐指数, 4.DO, 5.Cond, 6.WL, 7.TP, 8.区域, 9.SD, 10.TN, 11.pH值, 12.NH4+-N 图 6 洞庭湖不同季节和湖区环境因子对浮游植物种群和功能群的独立影响 Fig. 6 Independent effects of different seasons and environmental factors on phytoplankton population and functional groups in Dongting Lake |
根据TLI指数评价标准, 洞庭湖总体处于中营养状态[图 7(a)].各月份TLI指数差异不大:6月(46.95)>12月(46.80)>3月(46.52)>9月(46.48); 空间上, 东洞庭湖(50.55)>南洞庭湖(44.19)>西洞庭湖(42.74).此外, 东洞庭湖在6月、9月和12月达到轻度富营养状态, 而西、南洞庭湖在4个月份均处于中营养状态.

|
图 7 洞庭湖各湖区TLI指数、Shannon-Wiener指数和Q指数季节变化 Fig. 7 Seasonal variations in TLI index, Shannon-Wiener index, and Q index in Dongting Lake |
Shannon-Wiener指数显示[图 7(b)], 洞庭湖总体呈中污染状态.其中6月(3.13)>9月(2.81)>3月(2.73)>12月(2.59), 6月处于轻或无污染状态; 东洞庭湖(2.99)>西洞庭湖(2.73)>南洞庭湖(2.60), 均处于中污染状态.此外, 东洞庭湖在6月处于轻或无污染状态.
根据Q指数应用的建议, 洞庭湖水体识别的各功能群的F值权重如表 2所示, 洞庭湖总体处于好状态[图 7(c)].其中3月(3.87)>6月(3.72)>12月(3.61)>9月(3.51), 均处于好状态; 空间上, 西洞庭湖(3.91)>南洞庭湖(3.63)>东洞庭湖(3.56), 南洞庭湖在9月处于中等状态.
通过对各月份的环境因子之间及与TLI指数、Shannon-Wiener指数和Q指数之间进行相关性分析表明(图 8), WL与Flow、WT呈显著正相关, 与Cond呈显著负相关, 水温与DO、NH4+-N呈显著负相关; Cond与高锰酸盐指数、TP呈显著正相关; DO与NH4+-N、TN及TP呈显著正相关; 高锰酸盐指数与TP显著正相关.环境因子与指数之间, TLI指数与Cond、高锰酸盐指数及TP呈显著正相关, 与SD、TN/TP呈显著负相关; Shannon-Wiener指数与WL、Flow、Cond、高锰酸盐指数及TLI指数呈显著正相关; Q指数与Cond、高锰酸盐指数及TLI指数呈显著负相关.

|
1.WL, 2.Flow, 3.WT, 4.pH值, 5.Cond, 6.DO, 7.SD, 8.高锰酸盐指数, 9. NH4+-N, 10.TP, 11.TN, 12.TN/TP, 13.TLI指数, 14.Shannon-Wiener指数, 15.Q指数; *表示P<0.05, **表示P<0.01, ***表示P<0.001 图 8 洞庭湖环境因子与TLI指数、Shannon-Wiener指数和Q指数的Pearson相关性 Fig. 8 Pearson correlation of environmental factors with TLI index, Shannon-Wiener index and Q index in Dongting Lake |
从浮游植物时间分布来看, 洞庭湖浮游植物种群密度和功能群生物量变化趋势一致, 3月和12月浮游植物密度和生物量较低, 而在6月和9月整体呈上升趋势.温度的变化对浮游植物密度和生物量有着明显的影响, 是影响浮游植物季节变化的重要因子[32].此外, 东洞庭湖浮游植物密度、生物量和蓝绿藻在各月份显著高于西、南洞庭湖, 究其原因, 一方面是由于东洞庭湖位于回水区, 受西、南洞庭湖、区间来水以及岳阳市主城区城市、船舶生活污水汇集影响, 导致Cond、高锰酸盐指数、TN和TP明显高于西、南洞庭湖, 促进了浮游植物的生长繁殖; 另一方面由于西、南洞庭湖水文连通性较好, 在6月和9月水位较高, 湖区面积增大, 快速交换的水体对营养盐及浮游植物密度和生物量有一定的稀释作用[33], 水体的稀释超过了浮游植物的生长速度, 导致其密度和生物量下降[34].
从空间分布来看, 西、南洞庭湖浮游植物种群在3月、6月和12月均以硅藻为主, 9月以蓝藻为主; 而东洞庭湖3月以硅藻为主, 6月以蓝-绿-硅藻为主, 9月以蓝藻为主; 3个湖区在12月隐藻占比均有所升高.这些种群在分类学上, 生态特征各异, 与栖息的环境紧密相关.李哲等[35]研究表明, 适宜的流速有助于浮游植物生长, 高营养盐浓度不是浮游植物生长的限制因素, 但水体流速却是主要限制因子.蓝绿藻较适宜在稳定的水体中生长, 而硅藻由于个体比重大, 需要水体的扰动以增加其浮力对抗[36].洞庭湖是典型的通江湖泊, 受水位波动影响明显.6月三口、四水来水明显增多, 导致西、南洞庭湖水体流速较快[37], 虽然水温较高, 但仍以硅藻占优.反观东洞庭湖由于处在回水区, 水体扰动较小, 在6月蓝绿藻占比已明显增加.
本次调查洞庭浮游植物优势种为直链藻、舟形藻、针杆藻、小环藻、菱形藻、羽纹藻、栅藻、鱼腥藻、伪鱼腥藻、颤藻和隐藻, 这些种类在形态上的共同特征是细胞体积大或形态拉长成长条状, 在生理上这些种群对中低营养环境有较好的适应能力[10, 38].优势功能群为B、D、H1、J、Lo、MP、P、X1和Y, 所有功能群均对混合程度较高、浑浊水体或中低营养环境有较好的适应能力, 证实了洞庭湖的典型环境, 这与汪星等[25]对洞庭湖浮游植物功能群的研究一致.
3.2 洞庭湖浮游植物种群、功能群驱动因素及适用性分析自然水体中影响浮游植物的因素主要有环境选择和迁移扩散两个方面[39, 40].在水力滞留时间(大于40 d)较长的湖库中, 浮游植物主要受环境选择作用[40].与上述研究所不同的是, 虽然在三峡大坝运行后, 洞庭湖水力停留时间从18 d增加到29 d[20](小于40 d), 但浮游植物种群和功能群的季节性变化驱动大于空间扩散(图 6).可能是由于东洞庭湖与西、南洞庭湖的地理环境差异造成的, 西、南洞庭湖水文连通性较好, 更多的是空间扩散; 而东洞庭湖处于回水区, 水力停留时间的增加进一步促进了环境选择的作用.
有研究结果表明浮游植物种群和功能群演替的主要驱动因子是WT、高锰酸盐指数、DO、Cond、WL和TP.在另一通江湖泊的鄱阳湖中也发现了相似的规律[41], 水位变化、水温、透明度、电导率和氮营养盐是影响浮游植物的主要因素.略有不同的是, 磷营养盐成为本研究的主要驱动因子.前期研究表明, 洞庭湖氮磷浓度均高于富营养化阈值, 磷是浮游植物生长的潜在限制因子[42].水位波动可以直接影响浮游植物生长, 也可以通过影响理化参数间接影响浮游植物生长[43].本研究中, 水位与电导率呈显著负相关(图 8), 说明水位的上升对营养盐有一定的稀释作用, 但由于水温较高、光照较强, 营养盐浓度仍大于浮游植物生长阈值, 有利于浮游植物生长繁殖, 浮游植物密度和生物量在6月和9月整体呈上升趋势.DO也是浮游植物生长必不可少的条件, 孙来康等[44]研究表明, 由于蓝藻的光合作用, 使得丰水期DO升高, 与之呈正相关.本研究却发现蓝绿藻与DO呈负相关性, 硅藻与DO呈正相关, 这可能是由于DO与水温呈极显著负相关(图 8), 温度越低升温后DO浓度下降越明显[45].在水温和DO对浮游植物生产效率的耦合作用中, 水温的驱动性要更高.在适当的范围内, 有机物能够促进浮游植物的生长和代谢, 同时浮游植物的光合作用也会产生大量有机物, 导致水体高锰酸盐指数增加, 同时有机污染物被分解会产生大量含有氮磷等的营养物质, 可进一步促进浮游植物的生长[46].
功能群对环境的反映比个体及种群更为重要、综合性更强[41].RDA分析显示, 环境因子对功能群的解释率高于种群, 表明功能群对环境因子的响应比种群更敏感[47], 这与Liu等[48]对鄱阳湖浮游植物的研究结果一致.功能群是将某一特定生境且具有相似敏感性的浮游植物归为同一类群, 通过优势功能群对应的生境类型来说明水体情况, 有利于敏感地反映环境变化[9].功能群MP在各水期均占有优势, 主要是由于洞庭湖为频繁扰动的浑浊型浅水湖泊, 适宜其生长[25].功能群P/MP/D在3月占优, 此时水温较低, 功能群P、D与WT呈负相关, 同时为混合程度较高的中富营养浑浊浅水水体.6月(涨水期)水位抬升, 水体混合程度加剧, 功能群MP占比升高; 同时由于水温的升高, 功能群J生物量升高, 功能群P占比略有下降.9月功能群MP/H1占优, 适合富营养化、分层和低氮条件的功能群H1主要出现在东洞庭湖, 不仅由于9月较高的水温为蓝藻提供了有利条件, 同时该功能群与TN、NH4+-N呈负相关, 是由于涨水期和丰水期的稀释作用, 导致洞庭湖氮营养盐浓度降低, 其在低氮条件下的固氮优势得以显现[41].12月(枯水期)功能群Y/P/MP占优, 功能群Y与WL显著负相关, 上游来水的减小, 适宜广适性中营养静水环境的功能群Y生长.
此外, 对于水体中大量的浮游植物群落种类, 只有少数物种占主要优势, 稀有物种可能会增加浮游植物群落的冗余.在数据处理过程中, 优势种的划分依据与每个物种出现的频率及物种所占该样本的比例有关, 常常把小于0.02的种类都去除.但在6月和9月水体中, 种类多样性较高, 各个种类相对优势度并不是很高(表 1), 这种去除可能会造成信息损失的风险.采用优势功能群代替优势种, 则是将属于同一功能群的种类加和, 如果该功能群的相对丰度超过5%, 就应该考虑其反映的水生态特征, 避免了一些信息的损失[49].因此, 功能群识别洞庭湖浮游植物对环境因子的响应要优于浮游植物种群.
3.3 水质评价指标适用性分析汪星等[50]采用Shannon-Wiener指数对洞庭湖水质评价表明, 水质状况良好.熊剑等[51]运用TLI指数对洞庭湖近30年水质评价结果表明, 各湖区基本均处于中营养状态, 东洞庭湖在2008~2010年和2015年达到轻度富营养水平.但一般对湖泊水质进行单一的评价往往不能客观反映水体真实营养状态.因此, 本研究采用基于功能群的Q指数、Shannon-Wiener指数和TLI指数对洞庭湖水质进行综合评价, 结果显示TLI指数与Shannon-Wiener指数显著正相关, 与Q指数显著负相关, 即当富营养化程度高时, Shannon-Wiener指数表征的水体结果呈向好趋势, Q指数表征的水体是向差趋势.TLI指数与Q指数所表征的结果一致, 但Shannon-Wiener指数存在一定差异.例如, TLI指数显示洞庭湖水质处于中营养状态, Q指数显示洞庭湖总体处于好状态, 而Shannon-Wiener指数显示洞庭湖处于中等污染状态, 东洞庭湖Shannon-Wiener指数水质优于西、南洞庭湖.这可能是由于Shannon-Wiener指数与WL、Flow正相关, 西、南洞庭湖水文连通性较好, 水体对营养盐和浮游植物稀释作用较为明显, 导致水体营养盐较低, 但浮游植物种类和数量少, 导致得出污染程度高的结论, 而出现偏差[52].
Q指数计算结果显示, 洞庭湖为好的状态.Q指数较高的可能原因是洞庭湖功能群MP、P和D占比较高, 其他赋值低的功能群H1和J等比例虽有所增加, 但在整体比例中仍未完全占据优势.东洞庭湖TLI指数在6月、9月和12月达到轻度富营养状态, 随着6月和9月温度的升高, 功能群H1和J开始占优, 导致Q指数在该区域相应地降低, 说明东洞庭湖水体污染较严重, 该区域水流较缓的湖湾区面临蓝藻生长聚集甚至暴发水华风险[53].南洞庭湖Q指数在9月处于中等状态, 是由于功能群J生物量上升所致, 这比较接近实际水体环境特征.
整体看来, Q指数在洞庭湖水体有较好的适用性, 不仅能直接利用水体中浮游植物功能组对环境作出反应, 甚至还有预测功能, 例如在夏季的蓝绿藻水华暴发前, Q指数会发生突然降低的情况[54].
4 结论(1) 调查期间, 洞庭湖浮游植物共鉴定出6门61属, 主要以硅藻、蓝藻、绿藻和隐藻为主, 优势种群为直链藻、舟形藻、针杆藻、小环藻、菱形藻、羽纹藻、栅藻、鱼腥藻、伪鱼腥藻、颤藻和隐藻.洞庭湖浮游植物共划分23个功能群, 得到9个优势功能群, 分别是B、D、H1、J、Lo、MP、P、X1和Y, 浮游植物功能群总体演替趋势为P/MP/D(3月)→MP/P/J(6月)→MP/H1(9月)→Y/P/MP(12月).
(2) RDA分析表明, 浮游植物功能群识别洞庭湖浮游植物对环境因子的响应要优于浮游植物群落.层次分割结果表明, 洞庭湖环境因子对浮游植物的驱动大于空间驱动.影响浮游植物群落的环境因子重要性排序为:WT>高锰酸盐指数>DO>Cond>WL>TP, 影响浮游植物功能群的环境因子重要性排序为:WT>DO>Cond>高锰酸盐指数>WL>TP.
(3) 采用基于功能群的Q指数、Shannon-Wiener指数和TLI指数对洞庭湖水质进行综合评价, 对比分析发现利用Q指数进行水质评价在洞庭湖水体有较好的适用性, 洞庭湖水质总体处于好状态.
| [1] |
王爱爱, 冯佳, 谢树莲. 汾河中下游浮游藻类群落特征及水质分析[J]. 环境科学, 2014, 35(3): 915-923. Wang A A, Feng J, Xie S L. Phytoplankton community structure and assessment of water quality in the middle and lower reaches of Fenhe River[J]. Environmental Science, 2014, 35(3): 915-923. |
| [2] | Chalar G. The use of phytoplankton patterns of diversity for algal bloom management[J]. Limnologica, 2009, 39(3): 200-208. DOI:10.1016/j.limno.2008.04.001 |
| [3] | Whitton B A. Changing approaches to monitoring during the period of the 'Use of Algae for Monitoring Rivers' symposia[J]. Hydrobiologia, 2012, 695(1): 7-16. DOI:10.1007/s10750-012-1121-1 |
| [4] | Xu Y Y, Cai Q H, Ye L, et al. Asynchrony of spring phytoplankton response to temperature driver within a spatial heterogeneity bay of Three-Gorges Reservoir, China[J]. Limnologica, 2011, 41(3): 174-180. DOI:10.1016/j.limno.2010.10.004 |
| [5] |
李静, 崔凯, 卢文轩, 等. 春季和夏季巢湖浮游生物群落组成及其动态分析[J]. 水生生物学报, 2015, 39(1): 185-192. Li J, Cui K, Lu W X, et al. Community dynamics of spring-summer plankton in lake Chaohu[J]. Acta Hydrobiologica Sinica, 2015, 39(1): 185-192. |
| [6] | Komárek J. Review of the cyanobacterial genera implying planktic species after recent taxonomic revisions according to polyphasic methods: state as of 2014[J]. Hydrobiologia, 2016, 764(1): 259-270. DOI:10.1007/s10750-015-2242-0 |
| [7] | Salmaso N, Padisák J. Morpho-functional groups and phytoplankton development in two deep lakes(Lake Garda, Italy and Lake Stechlin, Germany)[J]. Hydrobiologia, 2007, 578(1): 97-112. DOI:10.1007/s10750-006-0437-0 |
| [8] | Shannon C E. The mathematical theory of communication[J]. Bell Labs Technical Journal, 1950, 3(9): 31-32. |
| [9] | Reynolds C S, Huszar V, Kruk C, et al. Towards a functional classification of the freshwater phytoplankton[J]. Journal of Plankton Research, 2002, 24(5): 417-428. DOI:10.1093/plankt/24.5.417 |
| [10] | Padisák J, Crossetti L O, Naselli-Flores L. Use and misuse in the application of the phytoplankton functional classification: a critical review with updates[J]. Hydrobiologia, 2009, 621(1): 1-19. DOI:10.1007/s10750-008-9645-0 |
| [11] |
郭芳, 顾继光, 赵剑, 等. 类群划分方法对南亚热带水库夏季浮游植物群落与环境响应关系的影响[J]. 环境科学, 2020, 41(11): 5050-5059. Guo F, Gu J G, Zhao J, et al. Effects of phytoplankton classifications on the relationship between phytoplankton community and environment in Summer subtropical reservoirs, southern China[J]. Environmental Science, 2020, 41(11): 5050-5059. |
| [12] | Padisák J, Borics G, Grigorszky I, et al. Use of phytoplankton assemblages for monitoring ecological status of lakes within the Water Framework Directive: the assemblage index[J]. Hydrobiologia, 2006, 553(1): 1-14. DOI:10.1007/s10750-005-1393-9 |
| [13] | Salmaso N, Naselli-Flores L, Padisák J. Functional classifications and their application in phytoplankton ecology[J]. Freshwater Biology, 2015, 60(4): 603-619. DOI:10.1111/fwb.12520 |
| [14] | Anderson S R, Harvey E L. Seasonal variability and drivers of microzooplankton grazing and phytoplankton growth in a subtropical estuary[J]. Frontiers in Marine Science, 2019, 6. DOI:10.3389/fmars.2019.00174 |
| [15] | Kruk C, Mazzeo N, Lacerot G, et al. Classification schemes for phytoplankton: a local validation of a functional approach to the analysis of species temporal replacement[J]. Journal of Plankton Research, 2002, 24(9): 901-912. DOI:10.1093/plankt/24.9.901 |
| [16] | Bovo-Scomparin V M, Train S. Long-term variability of the phytoplankton community in an isolated floodplain lake of the Ivinhema River State Park, Brazil[J]. Hydrobiologia, 2008, 610(1): 331-344. DOI:10.1007/s10750-008-9448-3 |
| [17] | Lv H, Yang J, Liu L M, et al. Temperature and nutrients are significant drivers of seasonal shift in phytoplankton community from a drinking water reservoir, subtropical China[J]. Environmental Science and Pollution Research, 2014, 21(9): 5917-5928. DOI:10.1007/s11356-014-2534-3 |
| [18] | Yuan Y X, Jiang M, Liu X T, et al. Environmental variables influencing phytoplankton communities in hydrologically connected aquatic habitats in the Lake Xingkai basin[J]. Ecological Indicators, 2018, 91: 1-12. DOI:10.1016/j.ecolind.2018.03.085 |
| [19] | Zhang M, Shi X L, Yang Z, et al. Long-term dynamics and drivers of phytoplankton biomass in eutrophic Lake Taihu[J]. Science of the Total Environment, 2018, 645: 876-886. DOI:10.1016/j.scitotenv.2018.07.220 |
| [20] | Yu Y W, Mei X F, Dai Z J, et al. Hydromorphological processes of Dongting Lake in China between 1951 and 2014[J]. Journal of Hydrology, 2018, 562: 254-266. DOI:10.1016/j.jhydrol.2018.05.015 |
| [21] | Geng M M, Wang K L, Yang N, et al. Spatiotemporal water quality variations and their relationship with hydrological conditions in Dongting Lake after the operation of the Three Gorges Dam, China[J]. Journal of Cleaner Production, 2021, 283. DOI:10.1016/j.jclepro.2020.124644 |
| [22] | Feng Y, Zheng B H, Jia H F, et al. Influence of social and economic development on water quality in Dongting Lake[J]. Ecological Indicators, 2021, 131. DOI:10.1016/j.ecolind.2021.108220 |
| [23] | Yin X Y, Yan G H, Wang X, et al. Spatiotemporal distribution pattern of phytoplankton community and its main driving factors in Dongting Lake, China-A seasonal study from 2017 to 2019[J]. Water, 2022, 14(11). DOI:10.3390/w14111674 |
| [24] |
汪梦琪, 汪金成, 王琪, 等. 洞庭湖区平水期浮游生物群落结构特征及富营养化现状[J]. 生态学杂志, 2018, 37(8): 2418-2429. Wang M Q, Wang J C, Wang Q, et al. Characteristics of plankton community structure and eutrophication status in Dongting Lake in the season with normal water level[J]. Chinese Journal of Ecology, 2018, 37(8): 2418-2429. DOI:10.13292/j.1000-4890.201808.004 |
| [25] |
汪星, 李利强, 郑丙辉, 等. 洞庭湖浮游藻类功能群的组成特征及其影响因素研究[J]. 中国环境科学, 2016, 36(12): 3766-3776. Wang X, Li L Q, Zheng B H, et al. Composition and influential factors of algal function groups in Dongting Lake[J]. China Environmental Science, 2016, 36(12): 3766-3776. DOI:10.3969/j.issn.1000-6923.2016.12.031 |
| [26] |
王丑明, 吴可方, 张屹, 等. 洞庭湖浮游植物时空变化特征及影响因素分析[J]. 淡水渔业, 2018, 48(4): 52-57. Wang C M, Wu K F, Zhang Y, et al. The analysis of spatial and temporal variations characteristics and driving factors of phytoplankton in Dongting Lake[J]. Freshwater Fisheries, 2018, 48(4): 52-57. DOI:10.3969/j.issn.1000-6907.2018.04.008 |
| [27] | Tian Z B, Zheng B H, Wang L J, et al. Long term(1997-2014) spatial and temporal variations in nitrogen in Dongting Lake, China[J]. PLoS One, 2017, 12(2). DOI:10.1371/journal.pone.0170993 |
| [28] |
胡鸿钧, 李尧英, 魏印心, 等. 中国淡水藻类[M]. 上海: 上海科学技术出版社, 1980. Hu H J, Li Y Y, Wei Y X, et al. Freshwater algae in China[M]. Shanghai: Shanghai Scientific & Technical Publishers, 1980. |
| [29] | Becker V, Caputo L, Ordóñez J, et al. Driving factors of the phytoplankton functional groups in a deep Mediterranean reservoir[J]. Water Research, 2010, 44(11): 3345-3354. DOI:10.1016/j.watres.2010.03.018 |
| [30] |
王明翠, 刘雪芹, 张建辉. 湖泊富营养化评价方法及分级标准[J]. 中国环境监测, 2002, 18(5): 47-49. Wang M C, Liu X Q, Zhang J H. Evaluate method and classification standard on lake eutrophication[J]. Environmental Monitoring in China, 2002, 18(5): 47-49. DOI:10.3969/j.issn.1002-6002.2002.05.018 |
| [31] | Lai J S, Zou Y, Zhang J L, et al. Generalizing hierarchical and variation partitioning in multiple regression and canonical analyses using the rdacca. hp R package[J]. Methods in Ecology and Evolution, 2022, 13(4): 782-788. DOI:10.1111/2041-210X.13800 |
| [32] | Temponeras M, Kristiansen J, Moustaka-Gouni M. Seasonal variation in phytoplankton composition and physical-chemical features of the shallow Lake Doirani, Macedonia, Greece[J]. Hydrobiologia, 2000, 424(1): 109-122. DOI:10.1023/A:1003909229980 |
| [33] | Ibañez M D S R. Phytoplankton composition and abundance of a central Amazonian floodplain lake[J]. Hydrobiologia, 1997, 362(1-3): 79-83. DOI:10.1023/A:1003124905996 |
| [34] | Salmaso N, Braioni M G. Factors controlling the seasonal development and distribution of the phytoplankton community in the lowland course of a large river in Northern Italy(River Adige)[J]. Aquatic Ecology, 2008, 42(4): 533-545. DOI:10.1007/s10452-007-9135-x |
| [35] |
李哲, 张曾宇, 杨中华, 等. 三峡澎溪河回水区流速对藻类原位生长速率的影响[J]. 湖泊科学, 2015, 27(5): 880-886. Li Z, Zhang Z Y, Yang Z H, et al. Effects of flow speed on the change of in situ growth rates of algae in Pengxi River backwater zone, Three Gorges Reservoir[J]. Journal of Lake Sciences, 2015, 27(5): 880-886. |
| [36] |
郑丙辉, 张佳磊, 王丽婧, 等. 大宁河水华敏感期浮游植物与环境因子关系[J]. 环境科学, 2011, 32(3): 641-648. Zheng B H, Zhang J L, Wang L J, et al. Exploration of relationships between phytoplankton and related environmental factors in the Daning River during sensitive period of algal blooms[J]. Environmental Science, 2011, 32(3): 641-648. |
| [37] |
王婷, 王坤, 王丽婧, 等. 三峡工程运行对洞庭湖水环境及富营养化风险影响评述[J]. 环境科学研究, 2018, 31(1): 15-24. Wang T, Wang K, Wang L J, et al. Impacts of the Three Gorges Dam operation on water environment and eutrophication of Dongting Lake: a review[J]. Research of Environmental Sciences, 2018, 31(1): 15-24. |
| [38] |
林秋奇, 胡韧, 段舜山, 等. 广东省大中型供水水库营养现状及浮游生物的响应[J]. 生态学报, 2003, 23(6): 1101-1108. Lin Q Q, Hu R, Duan S S, et al. Reservoir trophic states and the response of plankton in Guangdong Province[J]. Acta Ecologica Sinica, 2003, 23(6): 1101-1108. DOI:10.3321/j.issn:1000-0933.2003.06.010 |
| [39] | Xiao L J, Hu R, Peng L, et al. Dissimilarity of phytoplankton assemblages in two connected tropical reservoirs: effects of water transportation and environmental filtering[J]. Hydrobiologia, 2016, 764(1): 127-138. |
| [40] | Padisák J, Vasas G, Borics G. Phycogeography of freshwater phytoplankton: traditional knowledge and new molecular tools[J]. Hydrobiologia, 2016, 764(1): 3-27. |
| [41] |
钱奎梅, 刘宝贵, 陈宇炜. 鄱阳湖浮游植物功能群的长期变化特征(2009-2016年)[J]. 湖泊科学, 2019, 31(4): 1035-1044. Qian K M, Liu B G, Chen Y Y. Long term dynamics of phytoplankton functional groups in Lake Poyang during 2009-2016[J]. Journal of Lake Sciences, 2019, 31(4): 1035-1044. |
| [42] |
严广寒, 殷雪妍, 汪星, 等. 基于GAM模型的洞庭湖叶绿素a浓度与环境因子相关性分析[J]. 中国环境科学, 2022, 42(1): 313-322. Yan G H, Yin X Y, Wang X, et al. Analysis based on GAM model for the relationship between chlorophyll a concentration and environmental factors in Dongting Lake[J]. China Environmental Science, 2022, 42(1): 313-322. |
| [43] | Wu Z S, Liu J T, Huang J C, et al. Do the key factors determining phytoplankton growth change with water level in China's largest freshwater lake?[J]. Ecological Indicators, 2019, 107. DOI:10.1016/j.ecolind.2019.105675 |
| [44] |
孙来康, 杨涛, 万旭昊, 等. 西安城市河流浮游植物群落结构及其与环境因子的关系[J]. 水生生物学报, 2023, 47(4): 543-555. Sun L K, Yang T, Wan X H, et al. Phytoplankton community structure and its relationship with environmental factors in Xi'an urban rivers system[J]. Acta Hydrobiologica Sinica, 2023, 47(4): 543-555. |
| [45] | Kalff J. Limnology. Inland water ecosystems[M]. New Jersey: Prentice Hall, 2002. |
| [46] |
毕京博, 郑俊, 沈玉凤, 等. 南太湖入湖口叶绿素a时空变化及其与环境因子的关系[J]. 水生态学杂志, 2012, 33(6): 7-13. Bi J B, Zheng J, Shen Y F, et al. Spatial-temporal characteristics of chlorophyll-a concentration and its relationship with environmental factors in the inlets of South Taihu Lake[J]. Journal of Hydroecology, 2012, 33(6): 7-13. |
| [47] | Abonyi A, Leitão M, Lançon A M, et al. Phytoplankton functional groups as indicators of human impacts along the River Loire(France)[J]. Hydrobiologia, 2012, 698(1): 233-249. |
| [48] | Liu J F, Chen Y W, Li M J, et al. Water-level fluctuations are key for phytoplankton taxonomic communities and functional groups in Poyang Lake[J]. Ecological Indicators, 2019, 104: 470-478. |
| [49] |
胡韧, 蓝于倩, 肖利娟, 等. 淡水浮游植物功能群的概念、划分方法和应用[J]. 湖泊科学, 2015, 27(1): 11-23. Hu R, Lan Y Q, Xiao L J, et al. The concepts, classification and application of freshwater phytoplankton functional groups[J]. Journal of Lake Sciences, 2015, 27(1): 11-23. |
| [50] |
汪星, 郑丙辉, 刘录三, 等. 洞庭湖典型断面藻类组成及其与环境因子典范对应分析[J]. 农业环境科学学报, 2012, 31(5): 995-1002. Wang X, Zheng B H, Liu L S, et al. Canonical correspondence analysis of algae composition and its environmental factors of typical sections in Dongting Lake, China[J]. Journal of Agro-Environment Science, 2012, 31(5): 995-1002. |
| [51] |
熊剑, 喻方琴, 田琪, 等. 近30年来洞庭湖水质营养状况演变特征分析[J]. 湖泊科学, 2016, 28(6): 1217-1225. Xiong J, Yu F Q, Tian Q, et al. The evolution of water quality and nutrient condition in Lake Dongting in recent 30 years[J]. Journal of Lake Sciences, 2016, 28(6): 1217-1225. |
| [52] |
杜红春, 王晓宁, 吴虎, 等. 汉江中下游浮游植物群落结构、功能群特征及水质评价[J]. 长江流域资源与环境, 2021, 30(8): 1839-1847. Du H C, Wang X N, Wu H, et al. Community structure and functional groups of phytoplankton and water quality evaluation in the middle and lower Hanjiang River[J]. Resources and Environment in the Yangtze Basin, 2021, 30(8): 1839-1847. |
| [53] |
王琦, 欧伏平, 张雷, 等. 三峡工程运行后洞庭湖水环境变化及影响分析[J]. 长江流域资源与环境, 2015, 24(11): 1843-1849. Wang Q, Ou F P, Zhang L, et al. Changes of water environment in Dongting Lake and its impact analysis after the Three Gorges Project operation[J]. Resources and Environment in the Yangtze Basin, 2015, 24(11): 1843-1849. |
| [54] | Istvánovics V, Clement A, Somlyódy L, et al. Updating water quality targets for shallow Lake Balaton(Hungary), recovering from eutrophication[J]. Hydrobiologia, 2007, 581(1): 305-318. |