 2023, Vol. 44
2023, Vol. 44



2. 中国矿业大学矿山生态修复教育部工程研究中心, 徐州 221116;
3. 南京大学地理与海洋科学学院, 南京 210093
2. Engineering Research Center of Ministry of Education for Mine Ecological Restoration, China University of Mining and Technology, Xuzhou 221116, China;
3. School of Geography and Marine Science, Nanjing University, Nanjing 210093, China
露天采矿是人类对地表生态系统破坏最剧烈的活动, 严重干扰土壤环境和植被生长[1, 2], 继而破坏微生物的生存环境, 导致群落功能稳定性下降[3].土壤微生物是维持生物地球化学循环的重要环节[4], 在响应全球气候变化、维持生态系统功能和稳定等方面意义重大[5, 6].露采矿山进行植被恢复, 能够促进矿区土壤及微生物区系发育[7~9].近几年微生物在受损矿山生态修复中的作用已引起关注[10~12], 但多集中于矿区土壤微生物群落与环境因子之间的关联研究[13, 14], 如Guo等[13]的研究发现, 在大柳塔采煤沉陷区不同植被类型恢复模式下土壤因子对根际细菌和真菌群落组成的影响存在明显差异.Jia等[14]的研究发现, 在不同季节铜尾矿坝土壤微生物受不同的环境因子影响.然而目前学者对于矿区土壤中发挥特定功能的微生物研究相对较少, 尤其是碳循环功能基因仍缺乏深刻认知.
土壤碳循环在生物地球化学循环中具有重要地位, 微生物参与其中最重要的几个碳循环过程, 如碳降解(有机质的分解)、碳固定(CO2转化成有机物)和甲烷代谢(甲烷产生和甲烷氧化)[5, 15]. 微生物通过分泌多种酶降解动植物残体来加速碳的转化与迁移, 在土壤碳固持中发挥重要作用[16].功能基因主要通过编码特定功能蛋白参与微生物的生理生化过程并介导着元素循环, 具有指示土壤中生命活动的功能, 因此功能基因丰度可以预测土壤微生物的物质循环潜力和代谢特征[17, 18].微生物碳固定途径主要有: 卡尔文循环、还原三羧酸循环、还原乙酰辅酶A途径、3-羟基丙酸双循环、3-羟基丙酸/4-羟基丁酸循环和二羧酸/4-羟基丁酸循环这6种[15, 19].通过植物或自养微生物将CO2转化为有机化合物的生物固碳过程以及提高生态系统的碳固定能力, 是减少大气中CO2浓度的有效途径[15], 但不同植被因凋落物和根际分泌物不同, 从而导致土壤微生物多样性和功能的差异[13].前人对于功能微生物的研究主要集中于农田、森林和湿地等大型生态系统[20~23], 但是对于矿山这类人为干扰较大且功能特殊的复杂生态系统的关注相对较少, 亟待深入研究.
我国煤炭开采每年损毁土地约4.88万hm2, 平均复垦率不足30%[8].与此同时, 受损矿区多位于干旱半干旱区, 土壤贫瘠缺水且污染重[24], 许多矿山生态工程恢复效果差, 主要由于植被-土壤间恢复缺乏协同, 功能微生物的激发作用未得到充分发挥, 限制了生态系统的恢复力和自维持能力[25].因此, 厘清植被恢复引导下矿区复垦土壤功能微生物演替及驱动机制, 有助于充分挖掘功能微生物如何随着非生物和生物条件的变化而变化[26].为此, 本文选择黄土高原黑岱沟露天矿排土场不同类型的植被为研究对象, 利用实时荧光定量PCR技术检测碳循环相关功能基因的丰度, 通过探究不同植被类型对复垦土壤碳循环功能基因的影响, 揭示碳循环功能基因对不同植被类型的响应机制, 旨在为黄土高原受损矿山环境治理与生态增汇协同提供科学支撑.
1 材料与方法 1.1 研究区域概况选择内蒙古准格尔旗黑岱沟露天矿排土场复垦区(39°45′N, 111°16′E)为研究区域, 该区属典型的温带半干旱大陆性气候, 年均气温7.2℃, 年均降水量391.6 mm, 其中7~9月集中了70%.年蒸发量(1 824.7~2 896.1 mm)远高于降水量.该区为典型黄土沟壑区, 海拔为1 215~1 302 m.黑岱沟露天矿排土场占地面积为5 124.00 hm2, 从1995年起陆续开展植被恢复[27], 采用平台和边坡相间布置方式分区种植, 分3层, 每层高差为30 m.边坡为防止水土流失, 主要种植沙棘、柠条、紫穗槐等灌木和沙打旺、苜蓿、草木樨等草本.平台为培肥熟化土壤, 主要种植豆科类牧草、沙棘、油松、山杏和小叶杨等植被.
1.2 样品的采集与分析2020年7月28日至8月2日开展了野外调查, 采集草地(GL, 紫花苜蓿+长茅草)、灌丛(BL, 沙棘)、针叶林(CF, 油松1.5 m×2.0 m间距种植)、阔叶林(BF, 小叶杨1.5 m×3.0 m间距种植)和针阔混交林(MF, 油松和小叶杨1.5 m×3.0 m间距隔行种植)共5种植被类型土壤样品.每个样地随机选取5个点, 每个点收集0~10 cm表土200 g, 混合为约1 000 g的混合样, 总共收集了25个混合样(图 1).土样先过2 mm筛网去除砾石和根系等杂质, 再将其分为3份, 一份新鲜土样保存于-80℃冰箱用于提取土壤总DNA等供定量PCR测试; 另一份保存于4℃冰箱内, 供理化性质和酶活性测定; 剩余一份室内自然风干, 供其他理化性质测试.

|
图 1 研究区位置和采样点分布示意 Fig. 1 Location of the study area and sampling sites |
土壤pH值(水土比为2.5∶1)采用电位法测定(PHC-3C, 上海雷磁), 土壤有机碳(SOC)采用重铬酸钾水合氧化法测定[27]; 全氮(TN)采用半微量凯氏定氮法, 铵态氮(NH4+-N)和硝态氮(NO3--N)用氯化钙浸提-紫外分光光度法进行测定[28], 土壤有效磷(AP)釆用NaHCO3浸提-钼锑抗比色法[29].过氧化氢酶(CAT)活性通过高锰酸钾滴定法进行测定, 以1 g土壤1 h内消耗0.02 mol·L-1 KMnO4的体积表示; β-葡萄糖苷酶(BG)活性通过硝基酚比色法测定, 以培养1 h后1 g土壤中的对硝基酚的微克数表示; 多酚氧化酶(PPO)活性通过邻苯三酚比色法进行测定, 以培养2 h后1 g土壤中紫色没食子素的mg数表示; 土壤脲酶(URE)用苯酚钠-次氯酸钠比色法进行测定, 土壤亮氨酸氨基肽酶(S-LAP)活性用试剂盒检测(Beijing Solarbio Science & Technology Co., Ltd.)分光光度法测定, 土壤碱性磷酸酶活性采用磷酸苯二钠比色法进行测定[30].
1.3 微生物信息测定土壤碳循环功能基因丰度采用QMEC(quantitative microbial element cycling)基因芯片进行定量化测定(美格基因, 广州), 所有的引物均得到验证[31].具体方法是:按照MagaBio土壤DNA提取试剂盒方法, 取0.25 g土壤样品, 提取土壤DNA, 用Qubit 4.0 (Thermo Fisher Scientific, Waltham, USA)仪器对提取的土壤DNA总量及纯度进行检测, 将DNA浓度稀释为20 ng·μL-1, 分别将样品板和引物板试剂添加至高通量qPCR芯片SmartChip MyDesign Chip(Takara Biomedical Technology, Clontech)微孔中, 在SmartChip实时PCR系统进行扩增, 每个引物组所有qPCR反应设3个重复, 每次运行有一个对照.初始酶在95℃活化5 min, 再进行40个循环(95℃变性30 s, 58℃退火30 s, 72℃延伸30 s).在SmartChip实时PCR系统执行qPCR反应及荧光信号检测, 自动生成扩增和溶解曲线.使用Canco软件获取样本中各基因检出值和扩增循环数(Ct), 以16S rRNA作为内参进行标准化获取样本中各基因的相对定量信息, 再利用Roche仪检测16S rRNA基因的绝对定量信息, 最终换算其他基因的绝对定量信息[31, 32].
1.4 数据统计与分析采用单因素(one-way ANOVA)方差分析检验不同植被类型间土壤化学性质、酶活性以及土壤碳循环功能基因丰度的差异显著性, 平均值多重比较统计学检验采用最小显著极差法(LSD), 通过皮尔逊(Pearson)相关性来分析参与碳循环过程的功能基因丰度与环境因子间的相关关系.利用R语言软件进行绘制土壤碳循环功能基因丰度柱形图和与环境因子的相关性热图, 在Chiplot平台(https://www.chiplot.online/)上传数据后, 运算获得土壤酶活性的小提琴图, 使用Canoco5中的冗余分析(redundancy analysis, RDA)探讨不同环境因子对碳循环功能基因的影响.利用Amos 7.0软件拟合结构方程模型(structural equation model, SEM)分析土壤化学性质、酶活性以及碳循环功能基因丰度间的关系, 其中对土壤碳循环功能基因丰度进行归一化处理[33], 划分为碳降解、碳固定和甲烷代谢这三大功能类群, 具体公式如下:
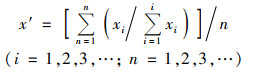
|
式中, xi表示样本单个基因的丰度, i和n分别表示所研究的样本数量和基因数量, x′表示碳降解、碳固定和甲烷代谢微生物功能类群的归一化丰度.
2 结果与分析 2.1 不同植被类型土壤化学性质和酶活性变化从表 1可知, 不同植被类型对土壤化学性质的影响存在显著性差异(P < 0.05).土壤pH和铵态氮含量变化趋势基本一致, 均表现为在林地(CF、BF和MF)土壤中高于GL和BL; 硝态氮含量表现出截然相反的变化趋势, 在CF和BF土壤中显著低于GL和BL(P < 0.01); 土壤有机碳、全氮含量均呈现出GL>MF>BL>BF>CF的趋势; 易氧化有机碳含量在BF土壤中显著低于其他4种植被类型(P < 0.01).速效磷含量在BL和CF土壤中较高, 在GL土壤中含量最低.总体上, GL和BL对土壤有机碳、全氮和硝态氮的积累作用优于CF.
|
|
表 1 不同植被类型土壤化学性质变化1) Table 1 Change in soil chemical properties among different vegetation types |
不同植被类型对土壤胞外酶活性的影响存在显著差异(P < 0.05, 图 2).MF土壤中过氧化氢酶(CAT)的活性比CF高出36.39%[图 2(a)]; BF和MF具有最高的β-葡萄糖苷酶(BG)活性, GL和BL次之, CF土壤的β-葡萄糖苷酶(BG)活性最低[图 2(b)]; 土壤多酚氧化酶(PPO)活性在GL和MF土壤中显著高于BL、CF和BF[图 2(c)]; 土壤脲酶(URE)和碱性磷酸酶(PHOS)活性均表现为在GL和BL土壤中显著高于CF、MF和BF土壤[图 2(d)和2(f)]; 土壤亮氨酸氨基肽酶(LAP)活性在BL和CF土壤中显著高于MF和BF土壤[图 2(e)].

|
单位中U表示1 nmol对硝基苯胺; 不同小写字母表示不同植被类型间差异显著(P < 0.05) 图 2 不同植被类型土壤酶活性变化 Fig. 2 Change in soil enzyme activities among different vegetation types |
定量PCR分析结果显示(图 3), 不同的植被类型之间淀粉(starch)、半纤维素(hemicellulose)、纤维素(cellulose)、几丁质(chitin)、果胶(pectin)和木质素(lignin)这6类天然碳化合物的降解过程及其相关基因有显著差异(P < 0.05).其中, 与淀粉降解有关的gam、Isop和amyA基因在BF土壤中最高, BL和MF次之, GL和CF土壤最低.与半纤维素降解有关的xylA、abfA和manA基因丰度在BF土壤最高, BL次之, GL和CF土壤最低.与纤维素降解有关的CDH、exg和naglu基因在BF土壤中均显著高于GL土壤.与几丁质降解有关的chiA基因和与木质素降解有关mnp、glx、pox和lig的基因均表现为BF最高, BL、CF和MF次之, GL最低.与果胶降解有关的exoPG基因在BF和BL土壤中显著高于CF.总体上, 碳降解相关功能基因丰度均表现为在BF土壤中基因丰度最高.

|
不同小写字母表示不同植被类型间差异显著(P < 0.05) 图 3 不同植被类型土壤碳降解功能基因丰度的变化 Fig. 3 Changes in abundance of functional genes of soil carbon degradation among different vegetation rehabilitation types |
不同植被类型中土壤微生物碳固定功能基因丰度如图 4所示.从中可知, 在碳固定相关功能基因中, 丰度最高的是rbcL、acsA和mct基因.参与卡尔文循环的rbcL基因丰度在BF和BL土壤较高, GL和CF土壤较低, 参与还原三羧酸循环的korA和aclB基因丰度在BF土壤最高, GL土壤中最低.参与还原乙酰辅酶A途径的acsA和acsB基因表现为:BF>BL>其它植被类型.甲烷代谢功能基因丰度(mxa、emGDH、pmoA和mmoX基因)变化趋势表现为在BF土壤最高, BL和MF次之, GL和CF土壤中最低(图 5).总体上, 碳固定和甲烷代谢相关功能基因均在BF土壤中基因丰度最高.

|
不同小写字母表示不同植被类型间差异显著(P < 0.05) 图 4 不同植被类型土壤碳固定功能基因的丰度变化 Fig. 4 Changes in abundance of functional genes of soil carbon fixation among different vegetation types |

|
图内不同的小写字母表示不同植被类型间差异显著(P < 0.05) 图 5 不同植被类型土壤甲烷代谢功能基因的丰度变化 Fig. 5 Changes in abundance of functional genes of soil methane metabolism among different vegetation types |
土壤碳循环相关功能基因与环境因子间的相关性分析如图 6所示.结果表明, 与碳降解过程相关的功能基因(gam、Isop、xylA、exg、chiA、mnp和glx)与铵态氮含量和BG酶活性呈显著正相关关系(P < 0.01), 与土壤有机碳、全氮、易氧化有机碳、硝态氮含量和脲酶活性呈显著负相关[P < 0.05, 图 6(a)].与碳固定过程相关的功能基因(korA、aclB、acsB、smtA和frdA)与铵态氮和BG酶活性呈显著正相关关系(P < 0.01), 与土壤有机碳、全氮、易氧化有机碳、硝态氮含量和脲酶活性呈显著负相关(P < 0.05); 与碳固定过程相关的功能基因(rbcL、acsA、accA和pccA)与BG酶活性显著正相关(P < 0.01), 与土壤易氧化有机碳含量呈显著负相关[P < 0.05, 图 6(b)].与甲烷代谢过程相关的功能基因(mxa、emGDH和mmoX基因)与铵态氮和BG酶活性呈显著正相关关系(P < 0.01), 与土壤有机碳、全氮、易氧化有机碳、硝态氮含量和脲酶活性呈显著负相关[P < 0.05, 图 6(b)].

|
(a)碳降解, (b)碳固定和甲烷代谢; 蓝色和橙色分别表示两个变量间正相关和负相关关系, 颜色越深, 关系越紧密; *、**和***分别表示P < 0.05、P < 0.01和P < 0.001 图 6 土壤碳循环微生物功能基因丰度与环境因子间的相关性热图 Fig. 6 Correlation heatmap between soil carbon cycle functional gene abundance and environmental factors |
利用冗余分析评估不同植被类型下环境因子对碳循环功能基因影响.环境因子解释了碳降解功能基因总变异的70.99%[图 7(a)], 解释了碳固定和甲烷代谢功能基因总变异的78.12%[图 7(b)].BF土壤中碳降解功能基因丰度与铵态氮和BG酶活性呈显著正相关, 与土壤易氧化有机碳显著负相关[图 7(a)].在GL土壤中, 碳固定和甲烷代谢功能基因丰度与硝态氮含量显著正相关[图 7(b)].碳固定和甲烷代谢功能基因丰度在BF土壤中与BG酶活性呈显著正相关, 与易氧化有机碳显著负相关, 而在CF土壤中恰好相反[图 7(b)].

|
NH4表示铵态氮; NO3表示硝态氮 图 7 土壤碳循环功能基因丰度与环境因子的冗余分析 Fig. 7 RDA of the soil carbon cycle functional gene abundance and environmental factors |
结构方程模型结果表明, 不同植被类型直接影响NO3--N含量, 再通过BG和URE间接影响碳降解过程功能基因丰度[图 8(a)], 此外, 不同植被类型会通过影响土壤BG酶活性来间接影响碳降解过程功能基因丰度[图 8(a)].不同植被类型同样能够通过影响土壤SOC含量和BG酶活性, 进而间接影响碳固定过程功能基因丰度[图 8(b)], 此外, 不同植被类型通过直接影响土壤中SOC、NH4+-N和NO3--N含量, 再通过BG酶活性间接影响碳固定相关功能基因丰度[图 8(b)].不同植被类型会通过影响土壤BG酶活性来间接影响甲烷代谢过程功能基因丰度[图 8(c)].不同植被类型直接影响土壤NO3--N含量, 再通过BG和URE间接影响甲烷代谢相关功能基因丰度[图 8(c)].

|
箭头上的数值表示标准通径系数, 箭头粗细表示相关性的高低, 绿色箭头表示显著正相关, 红色箭头表示显著负相关, R2表示通径解释度; *、**和***分别表示各路径显著性水平为P < 0.05、P < 0.01和P < 0.001 图 8 植被恢复中不同路径影响碳循环功能基因丰度的结构方程模型 Fig. 8 Structural equation modeling (SEM) of vegetation rehabilitation type patterns affecting soil carbon cycle functional gene abundance through different ways |
植物残体和根系分泌物是土壤有机碳的主要来源, 进行植被恢复能够有效改善土壤的养分状况[34, 35].本研究中不同植被类型的土壤化学性质差异显著, 土壤有机碳和全氮含量呈现出GL>MF>BL>BF>CF的趋势(表 1), 这可能是由于土壤有机碳主要来源于植物残体和根系分泌物, 不同植被类型的根系分泌物和植物自身生理代谢存在明显差异[36], 该结果表明草地和混交林更利于土壤有机碳和全氮的积累.土壤硝态氮含量在GL土壤高于CF和BF(表 1), 可能是因为草地种植的紫花苜蓿属于豆科植物, 能够通过自身的根瘤菌来固定大气中的氮[37], 而针叶林油松土壤相对比较贫瘠, 特别是活性氮含量[38], 这与以往笔者在黄土高原矿区植被恢复的研究结果一致, 即草地土壤中硝态氮的含量高于针叶林和阔叶林[7].综合来看, GL和BL对土壤有机碳、全氮和硝态氮等养分的积累优于CF.
植被恢复过程中, 土壤酶参与土壤碳库分解转化, 加速了物质转化与能量循环, 是评价土壤质量的重要生物活性指标[2, 39].有研究表明, 植被类型能够调节土壤的酶活性[40, 41].本研究发现不同植被类型对土壤胞外酶活性影响显著, 其中与碳降解相关的β-葡萄糖苷酶活性在GL和MF土壤显著高于CF[图 2(b)], 可能是因为针叶林(CF)植被凋落物相对较少[22], 微生物分泌碳降解酶进行催化分解时所能利用的底物也相对较少.多酚氧化酶活性在CF和MF土壤中显著高于BL[图 2(c)], 而多酚氧化酶在针叶林马尾松生境中较为活跃[30].土壤脲酶和碱性磷酸酶活性均在GL和BL土壤中显著高于CF、MF和BF[图 2(d)和2(f)], 这也与黄土高原矿区草地和灌木复垦土壤这两种酶活性高于针叶林的结果相吻合[8].
3.2 不同植被类型对矿区复垦土壤碳循环关键功能基因的影响在植被恢复过程中, 大量的外源碳进入土壤中后会被土壤碳降解酶分解, 释放出低分子的糖, 从而为土壤微生物生长代谢提供碳源和能量[42].淀粉、半纤维素、纤维素和木质素等有机物通过微生物的降解作用参与碳循环过程[15].淀粉是土壤中典型的易分解碳, 淀粉酶基因编码合成的淀粉酶可以特异性地分解土壤中的淀粉[22].本研究发现与淀粉降解有关的基因在BF土壤中最高, GL和CF土壤最低, 表明淀粉酶的分解潜力在阔叶林土壤中较强, 而在草地和针叶林土壤中较弱(图 3).纤维素是一种难分解的碳, 纤维素酶基因丰度能够反映群落对难分解碳的分解潜力[22].本研究发现与纤维素降解有关的基因在BF土壤中较高, 表明纤维素酶在阔叶林土壤中对难分解碳的分解潜力相对较强(图 3).总体来看, 与碳降解相关的功能基因均在BF土壤中丰度最高, 表明阔叶林土壤中碳降解酶对易分解和难分解的碳都有较强的分解潜力.
卡尔文循环是CO2固定的最主要途径, 在调节大气CO2浓度方面发挥重要作用[19].RubisCO酶催化卡尔文循环中的第一步CO2固定反应, RubisCO的功能基因rbcL作为分子标记可用来反映微生物的固碳能力[15].本研究结果发现在碳固定过程中rbcL基因丰度在BF和BL土壤较高, 而在GL和CF土壤较低, 表明固碳微生物在阔叶林和灌木土壤中固碳潜力较强, 对土壤碳汇的贡献较大(图 4).在碳固定相关功能基因中, acsA和mct基因丰度较高, 这与宁岳伟等[43]关于矿区植被恢复的碳固定基因结果相一致.此外, 参与还原三羧酸循环的korA和aclB基因和还原乙酰辅酶A途径的acsA和acsB基因一致表现为BF土壤丰度最高, 表明BF土壤微生物有较强的固碳能力(图 4).土壤甲烷代谢包括甲烷产生和甲烷氧化两个过程, 主要由产甲烷菌和甲烷氧化菌共同控制[15].本研究中甲烷代谢功能基因丰度均在BF土壤最高, GL和CF土壤中最低, 表明BF土壤微生物具有较高的甲烷产生和氧化速率(图 5).
3.3 碳循环关键功能基因与矿区土壤环境因子之间的关系微生物群落基因丰度通常伴随着植被类型和土壤特性的改变而变化[26, 44, 45].本研究发现与碳降解过程相关的功能基因与铵态氮和BG酶活性显著正相关, 与土壤有机碳、全氮、易氧化有机碳、硝态氮含量和脲酶活性显著负相关[图 6(a)], Zhang等[46]通过对两种森林类型微生物碳代谢功能基因研究发现, 土壤易氧化有机碳和全氮是影响微生物碳代谢功能的主要因素.本研究发现所有固碳基因均与易氧化有机碳含量和BG酶活性显著相关[图 6(b)], 结构方程模型结果显示, 植被恢复类型通过影响土壤有机碳含量和BG酶活性, 或通过影响土壤有机碳、铵态氮和硝态氮含量间接影响BG酶活性, 进而影响碳固定基因丰度[图 8(a)].马静等[47]在东部矿区复垦土壤的研究也同样发现固碳功能基因丰度与土壤BG酶活性显著相关.刘茗等[48]通过对亚热带不同森林植被类型土壤固碳微生物的研究, 发现土壤有机碳和全氮是影响土壤固碳细菌群落形成的主要因素.本研究中甲烷代谢功能基因与土壤易氧化有机碳、铵态氮、硝态氮、全氮含量以及BG酶和脲酶活性显著相关[图 6(c)], 宁岳伟等[43]通过对矿区复垦植被微生物功能基因的研究, 其中也发现甲烷代谢功能基因与脲酶活性显著相关.本研究BF土壤中碳循环功能基因丰度较高, 主要与铵态氮和BG酶活性较高有关, 与易氧化有机碳和脲酶活性较低有关[图 7(a)].上述结果表明驱动土壤碳循环的功能微生物与土壤肥力状况和养分循环密切相关[22], 即土壤养分状况会影响土壤中功能微生物的活性和功能发挥.
4 结论(1) 不同植被类型对土壤化学性质、酶活性和碳循环基因丰度均有显著影响, 草地和灌丛对土壤有机碳、全氮和硝态氮的积累作用优于针叶林.在所有碳固定相关功能基因中, rbcL、acsA和mct基因丰度最高.碳循环功能基因丰度在阔叶林土壤最高, 这与阔叶林土壤中铵态氮含量和BG酶活性较高以及易氧化有机碳和脲酶活性较低密切相关.
(2) 碳降解和甲烷代谢功能基因与土壤铵态氮含量和BG酶活性呈显著正相关, 与土壤有机碳、全氮、易氧化有机碳、硝态氮含量和脲酶活性呈显著负相关, 碳固定功能基因与土壤易氧化有机碳含量和BG酶活性密切相关.
(3) 不同植被类型会通过影响土壤BG酶活性, 或通过影响土壤NO3--N含量间接影响BG和URE酶活性, 进而影响碳降解和甲烷代谢基因丰度.不同植被类型通过影响土壤有机碳和BG酶活性, 或通过影响土壤有机碳、铵态氮和硝态氮含量间接影响BG酶活性, 进而影响碳固定功能基因丰度.
| [1] | Wang J M, Wang H D, Cao Y G, et al. Effects of soil and topographic factors on vegetation restoration in opencast coal mine dumps located in a loess area[J]. Scientific Reports, 2016, 6. DOI:10.1038/srep22058 |
| [2] |
张振佳, 曹银贵, 王舒菲, 等. 平朔黄土露天矿区复垦地表层土壤微生物与酶活性分析[J]. 生态学报, 2021, 41(1): 110-123. Zhang Z J, Cao Y G, Wang S F, et al. Characteristics and differences of surface soil microbial population and enzyme activities in opencast mining area of Pingshuo[J]. Acta Ecologica Sinica, 2021, 41(1): 110-123. |
| [3] | Li Y Y, Wen H Y, Chen L Q, et al. Succession of bacterial community structure and diversity in soil along a chronosequence of reclamation and re-vegetation on coal mine spoils in China[J]. PLoS One, 2014, 9(12). DOI:10.1371/journal.pone.0115024 |
| [4] | Bardgett R D, Van Der Putten W H. Belowground biodiversity and ecosystem functioning[J]. Nature, 2014, 515(7528): 505-511. DOI:10.1038/nature13855 |
| [5] | Zhou J Z, Xue K, Xie J P, et al. Microbial mediation of carbon-cycle feedbacks to climate warming[J]. Nature Climate Change, 2012, 2(2): 106-110. DOI:10.1038/nclimate1331 |
| [6] | Delgado-Baquerizo M, Maestre F T, Reich P B, et al. Microbial diversity drives multifunctionality in terrestrial ecosystems[J]. Nature Communications, 2016, 7. DOI:10.1038/ncomms10541 |
| [7] |
马静, 董文雪, 朱燕峰, 等. 东部平原矿区复垦土壤微生物群落特征及其组装过程[J]. 环境科学, 2022, 43(7): 3844-3853. Ma J, Dong W X, Zhu Y F, et al. Characteristics and assembly process of reclaimed soil microbial communities in eastern plain mining areas[J]. Environmental Science, 2022, 43(7): 3844-3853. |
| [8] | Zhao J, Ma J, Yang Y J, et al. Response of soil microbial community to vegetation reconstruction modes in mining areas of the Loess Plateau, China[J]. Frontiers in Microbiology, 2021, 12. DOI:10.3389/fmicb.2021.714967 |
| [9] |
马静, 卢永强, 张琦, 等. 黄土高原采煤沉陷对土壤微生物群落的影响[J]. 土壤学报, 2021, 58(5): 1278-1288. Ma J, Lu Y Q, Zhang Q, et al. Effects of coal mining subsidence on soil microbial community in the Loess Plateau[J]. Acta Pedologica Sinica, 2021, 58(5): 1278-1288. |
| [10] | Liu Y, Chen X T, Liu J X, et al. Temporal and spatial succession and dynamics of soil fungal communities in restored grassland on the Loess Plateau in China[J]. Land Degradation & Development, 2019, 30(11): 1273-1287. |
| [11] |
贺龙, 李艳琴, 李彬春, 等. 矿区不同植被复垦模式对土壤细菌群落结构的影响[J]. 环境科学, 2017, 38(2): 752-759. He L, Li Y Q, Li B C, et al. Effects of different vegetation types and reclamation years on soil bacterial community structure in reclaimed mine areas[J]. Environmental Science, 2017, 38(2): 752-759. |
| [12] | Yang Y, Cheng H, Liu L X, et al. Comparison of soil microbial community between planted woodland and natural grass vegetation on the Loess Plateau[J]. Forest Ecology and Management, 2020, 460. DOI:10.1016/j.foreco.2019.117817 |
| [13] | Guo Y N, Liu X H, Tsolmon B, et al. The influence of transplanted trees on soil microbial diversity in coal mine subsidence areas in the Loess Plateau of China[J]. Global Ecology and Conservation, 2020, 21. DOI:10.1016/j.gecco.2019.e00877 |
| [14] | Jia T, Guo T Y, Yao Y S, et al. Seasonal microbial community characteristic and its driving factors in a copper tailings dam in the Chinese Loess Plateau[J]. Frontiers in Microbiology, 2020, 11. DOI:10.3389/fmicb.2020.01574 |
| [15] |
刘洋荧, 王尚, 厉舒祯, 等. 基于功能基因的微生物碳循环分子生态学研究进展[J]. 微生物学通报, 2017, 44(7): 1676-1689. Liu Y Y, Wang S, Li S Z, et al. Advances in molecular ecology on microbial functional genes of carbon cycle[J]. Microbiology China, 2017, 44(7): 1676-1689. |
| [16] | Liang C, Schimel J P, Jastrow J D. The importance of anabolism in microbial control over soil carbon storage[J]. Nature Microbiology, 2017, 2(8). DOI:10.1038/nmicrobiol.2017.105 |
| [17] | Trivedi P, Delgado-Baquerizo M, Trivedi C, et al. Microbial regulation of the soil carbon cycle: evidence from gene-enzyme relationships[J]. The ISME Journal, 2016, 10(11): 2593-2604. DOI:10.1038/ismej.2016.65 |
| [18] | Chen J, Sinsabaugh R L. Linking microbial functional gene abundance and soil extracellular enzyme activity: Implications for soil carbon dynamics[J]. Global Change Biology, 2021, 27(7): 1322-1325. DOI:10.1111/gcb.15506 |
| [19] |
袁红朝, 秦红灵, 刘守龙, 等. 固碳微生物分子生态学研究[J]. 中国农业科学, 2011, 44(14): 2951-2958. Yuan H Z, Qin H L, Liu S L, et al. Advances in research of molecular ecology of carbon fixation microorganism[J]. Scientia Agricultura Sinica, 2011, 44(14): 2951-2958. |
| [20] |
刘琼, 魏晓梦, 吴小红, 等. 稻田土壤固碳功能微生物群落结构和数量特征[J]. 环境科学, 2017, 38(2): 760-768. Liu Q, Wei X M, Wu X H, et al. Characteristic of abundances and diversity of carbon dioxide fixation microbes in paddy soils[J]. Environmental Science, 2017, 38(2): 760-768. |
| [21] |
沈李东, 金靖昊, 刘心. 内陆湿地与水体甲烷厌氧氧化功能微生物研究进展[J]. 生态学报, 2022, 42(9): 3842-3855. Shen L D, Jin J H, Liu X. Research progress on anaerobic methanotrophs in inland wetlands and freshwater aquatic systems[J]. Acta Ecologica Sinica, 2022, 42(9): 3842-3855. |
| [22] |
李毳, 乔沙沙, 刘晋仙, 等. 温带亚高山针叶林土壤碳循环微生物群落的时空动态[J]. 中国环境科学, 2020, 40(10): 4540-4548. Li C, Qiao S S, Liu J X, et al. Spatiotemporal dynamics of microbial communities involved in soil carbon cycling of subalpine temperate coniferous forest[J]. China Environmental Science, 2020, 40(10): 4540-4548. |
| [23] |
邓玲玲, 王如海, 吴电明. 增温和互花米草入侵对崇明东滩湿地土壤碳循环功能基因的影响[J]. 南京信息工程大学学报(自然科学版), 2022, 14(1): 62-76. Deng L L, Wang R H, Wu D M. Effects of experimental warming and Spartina alterniflora invasion on soil carbon cycle functional genes in Chongming Dongtan wetland[J]. Journal of Nanjing University of Information Science & Technology (Natural Science Edition), 2022, 14(1): 62-76. |
| [24] |
周艳, 陈樯, 邓绍坡, 等. 西南某铅锌矿区农田土壤重金属空间主成分分析及生态风险评价[J]. 环境科学, 2018, 39(6): 2884-2892. Zhou Y, Chen Q, Deng S P, et al. Principal component analysis and ecological risk assessment of heavy metals in farmland soils around a Pb-Zn mine in Southwestern China[J]. Environmental Science, 2018, 39(6): 2884-2892. |
| [25] | Ayala-Orozco B, Gavito M E, Mora F, et al. Resilience of soil properties to land-use change in a tropical dry forest ecosystem[J]. Land Degradation & Development, 2018, 29(2): 315-325. |
| [26] | Yang Y F, Gao Y, Wang S P, et al. The microbial gene diversity along an elevation gradient of the Tibetan grassland[J]. The ISME Journal, 2014, 8(2): 430-440. |
| [27] | Lei H, Peng Z, Yigang H, et al. Vegetation and soil restoration in refuse dumps from open pit coal mines[J]. Ecological Engineering, 2016, 94: 638-646. |
| [28] | Barahona E, Iriarte A. An overview of the present state of standardization of soil sampling in Spain[J]. Science of the Total Environment, 2001, 264(1-2): 169-174. |
| [29] | 鲍士旦. 土壤农化分析[M]. ((第三版)). 北京: 中国农业出版社, 2000. |
| [30] | Yin R, Deng H, Wang H L, et al. Vegetation type affects soil enzyme activities and microbial functional diversity following re-vegetation of a severely eroded red soil in sub-tropical China[J]. CATENA, 2014, 115: 96-103. |
| [31] | Zheng B X, Zhu Y G, Sardans J, et al. QMEC: a tool for high-throughput quantitative assessment of microbial functional potential in C, N, P, and S biogeochemical cycling[J]. Science China Life Sciences, 2018, 61(12): 1451-1462. |
| [32] | Chen Q L, Ding J, Li C Y, et al. Microbial functional attributes, rather than taxonomic attributes, drive top soil respiration, nitrification and denitrification processes[J]. Science of the Total Environment, 2020, 734. DOI:10.1016/j.scitotenv.2020.139479 |
| [33] | Luo G W, Xue C, Jiang Q H, et al. Soil carbon, nitrogen, and phosphorus cycling microbial populations and their resistance to global change depend on soil C: N: P stoichiometry[J]. mSystems, 2020, 5(3). DOI:10.1128/mSystems.00162-20 |
| [34] | Crouzeilles R, Curran M, Ferreira M S, et al. A global meta-analysis on the ecological drivers of forest restoration success[J]. Nature Communications, 2016, 7. DOI:10.1038/ncomms11666 |
| [35] |
王金满, 郭凌俐, 白中科, 等. 黄土区露天煤矿排土场复垦后土壤与植被的演变规律[J]. 农业工程学报, 2013, 29(21): 223-232. Wang J M, Guo L L, Bai Z K, et al. Succession law of reclaimed soil and vegetation on opencast coal mine dump of loess area[J]. Transactions of the Chinese Society of Agricultural Engineering, 2013, 29(21): 223-232. |
| [36] | Zechmeister-Boltenstern S, Keiblinger K M, Mooshammer M, et al. The application of ecological stoichiometry to plant-microbial-soil organic matter transformations[J]. Ecological Monographs, 2015, 85(2): 133-155. |
| [37] | Yang Y, Liu B R, An S S. Ecological stoichiometry in leaves, roots, litters and soil among different plant communities in a desertified region of Northern China[J]. CATENA, 2018, 166: 328-338. |
| [38] | Zeng Q C, Jia P L, Wang Y, et al. The local environment regulates biogeographic patterns of soil fungal communities on the Loess Plateau[J]. CATENA, 2019, 183. DOI:10.1016/j.catena.2019.104220 |
| [39] |
李文杰, 张祯皎, 赵雅萍, 等. 刺槐林恢复过程中土壤微生物碳降解酶的变化及与碳库组分的关系[J]. 环境科学, 2022, 43(2): 1050-1058. Li W J, Zhang Z J, Zhao Y P, et al. Changes in soil microbial carbon-degrading enzymes and their relationships with carbon pool components during the restoration process of Robinia pseudoacacia[J]. Environmental Science, 2022, 43(2): 1050-1058. |
| [40] | He Q Q, Wu Y H, Bing H J, et al. Vegetation type rather than climate modulates the variation in soil enzyme activities and stoichiometry in subalpine forests in the eastern Tibetan Plateau[J]. Geoderma, 2020, 374. DOI:10.1016/j.geoderma.2020.114424 |
| [41] | Xiao L, Liu G B, Li P, et al. Ecoenzymatic stoichiometry and microbial nutrient limitation during secondary succession of natural grassland on the Loess Plateau, China[J]. Soil and Tillage Research, 2020, 200. DOI:10.1016/j.still.2020.104605 |
| [42] | Davidson E A, Ishida F Y, Nepstad D C. Effects of an experimental drought on soil emissions of carbon dioxide, methane, nitrous oxide, and nitric oxide in a moist tropical forest[J]. Global Change Biology, 2004, 10(5): 718-730. |
| [43] |
宁岳伟, 刘勇, 张红, 等. 煤矿矿区复垦植被类型对土壤微生物功能基因和酶活的影响[J]. 环境科学, 2022, 43(9): 4647-4654. Ning Y W, Liu Y, Zhang H, et al. Effects of different vegetation types on soil microbial functional genes and enzyme activities in reclaimed coal mine[J]. Environmental Science, 2022, 43(9): 4647-4654. |
| [44] | Yue H W, Wang M M, Wang S P, et al. The microbe-mediated mechanisms affecting topsoil carbon stock in Tibetan grasslands[J]. The ISME Journal, 2015, 9(9): 2012-2020. |
| [45] | Hu Y G, Zhang Z S, Huang L, et al. Shifts in soil microbial community functional gene structure across a 61-year desert revegetation chronosequence[J]. Geoderma, 2019, 347: 126-134. |
| [46] | Zhang Y G, Cong J, Lu H, et al. An integrated study to analyze soil microbial community structure and metabolic potential in two forest types[J]. PLoS One, 2014, 9(4). DOI:10.1371/journal.pone.0093773 |
| [47] |
马静, 董文雪, 朱燕峰, 等. 东部平原矿区复垦对土壤微生物固碳潜力的影响[J]. 煤炭学报, 2022, 47(3): 1306-1317. Ma J, Dong W X, Zhu Y F, et al. Impact of land reclamation on the carbon sequestration potential of soil microorganisms in the disturbed mining area of eastern plain[J]. Journal of China Coal Society, 2022, 47(3): 1306-1317. |
| [48] |
刘茗, 曹林桦, 刘彩霞, 等. 亚热带4种典型森林植被土壤固碳细菌群落结构及数量特征[J]. 土壤学报, 2021, 58(4): 1028-1039. Liu M, Cao L H, Liu C X, et al. Characterization of population and community structure of carbon-sequestration bacteria in soils under four types of forest vegetations typical of subtropical zone[J]. Acta Pedologica Sinica, 2021, 58(4): 1028-1039. |