 2022, Vol. 43
2022, Vol. 43



2. 农业农村部产地环境污染防控重点实验室, 天津市农业环境与农产品安全重点实验室, 天津 300191;
3. 国信司南(北京)地理信息技术有限公司, 北京 100089
2. Key Laboratory of Original Agro-Environmental Pollution Prevention and Control, Ministry of Agriculture and Rural Affairs, Tianjin Key Laboratory of Agro-Environment and Safe-Product, Tianjin 300191, China;
3. Geo-Compass Information Technology Co., Beijing 100089, China
汞(mercury, Hg)是一种毒性强, 且可以通过大气进行长距离和跨区域传输的全球性污染物[1], 被联合国环境规划署、世界卫生组织、中国、欧盟和美国环保署等列为优先控制污染物[2].贵州铜仁市是我国涉Hg资源产业规模最大的集散地, 被称为“中国汞都”[3].然而, 由于长期金属采矿和冶炼, 向周围环境中排放了大量的含Hg“三废”[4].据报道[5], 截至2002年, 矿山活动共排放含Hg废气202.4亿m3、废水5 192万m3和废渣426万m3, 对当地农田环境造成了极其严重的污染.苟体忠等[6]对矿坑、冶炼厂及遗址和冶炼废渣堆等重点区域附近土壤进行了调查, 结果显示土壤ω(Hg)高达3.37~149 mg ·kg-1, 均值为29.00 mg ·kg-1, 是贵州省土壤背景值的263.61倍, 属于严重污染水平, 存在极强潜在生态风险.高令健等[7]的研究表明, Hg矿周边稻田土壤ω(总Hg)和ω(HgCH3)分别为0.21~207 mg ·kg-1和0.42~13 μg ·kg-1, 59%的稻田土壤处于重度Hg污染, 75%的稻田土壤存在极强的Hg潜在生态风险.Tang等[8]的研究发现, Hg矿周边农田土壤ω(总Hg)为0.12~21.7 mg ·kg-1, 均值为6.00 mg ·kg-1, 是贵州省土壤背景值的54.55倍, 达到我国农用地污染风险管控标准值(GB 15618-2018)的2.5倍.
微生物是土壤生态系统的重要组成部分, 对于重金属胁迫响应十分敏感[9].Li等[10]的研究发现, 重金属暴露期间土壤中的敏感种群会逐渐减少, 可忍受高浓度重金属的菌种逐步占主导地位, 从而限制了其他敏感性细菌的繁殖.Liu等[11]研究了长期土壤Hg污染对土壤微生物群落与功能基因的影响机制, 并基于微生物群落网络分析解释了土壤Hg污染对主要微生物生态类群的影响, 发现土壤Hg在很大程度上影响了微生物群落的分类和功能属性.Frossard等[12]研究了长期和短期Hg污染对土壤微生物组的影响, 发现长期Hg污染对土壤细菌和真菌群落结构和多样性具有明显的影响.此外, 土壤环境因素的变化也是影响土壤微生物群落结构组成的重要因素.Jiang等[13]研究了铜(Cu)矿区耐Cu植物根际土壤细菌群落结构, 发现影响细菌群落的最重要的环境因子是土壤pH而不是Cu浓度.黄健等[14]的研究表明, pH和重金属浓度是铜陵狮子山矿区土壤微生物群落结构的主要影响因子.Kenarova等[15]和Stemmer等[16]的研究表明土壤含水率(WC)、有机质(OM)、pH和速效钾(AK)等理化性质会明显影响重金属毒性, 进而影响土壤微生物群落结构.综合以上研究结果发现, 影响土壤微生物的环境因素具有多样性, 其群落结构的变化不仅与重金属浓度有关, 还取决于土壤中的理化性质等参数, 是最具潜力的敏感生物指标, 并且能及时预测土壤养分及土壤环境质量的变化[17].
微生物群落结构组成及其环境影响因子不同, 其在环境中的生态功能也不同.朱永官等[18]认为土壤健康与微生物组密切相关, 细菌和古菌等构成的土壤微生物组可以作为土壤健康的关键性评价指标, 揭示微生物群落结构及其与环境因子的关系已经成为近年来微生物生态领域的研究热点.然而, 国内外研究大多集中于矿区周边土壤重金属污染现状调查以及生态风险评估, 但对于矿区农田土壤微生物群落结构特征与环境因子关系的报道却较少, 尤其是受污染土壤健康的生物学指标确定的研究.基于此, 本文以铜仁Hg矿区周边受污染农田土壤为研究对象, 分析了土壤基本环境因子(理化性质、Hg含量、酶活性)和微生物(细菌、真菌)群落结构特征, 采用冗余分析(RDA)和相关性网络分析探究Hg矿区周边农田土壤微生物群落结构与环境因子的互作关系, 以期为Hg矿区周边农田土壤健康评价和受污染土壤修复治理提供理论基础.
1 材料与方法 1.1 研究区概况铜仁市位于贵州省东北部, 武陵山区腹地, 东临湖南省怀化市, 北与重庆市接壤, 西北高, 东南低, 全境以山地为主, 大多数地域属中亚热带季风湿润气候区.本文选择采样点涉及铜仁市境内3个区县, 分别为碧江区、石阡县和江口县, 当年种植作物均为凉粉草, 作物种植年限1 a.采样点自然概况如表 1.
|
|
表 1 研究区概况1) Table 1 Study area survey |
1.2 样地设置与土样采集
2020年10月, 在每个研究区域随机设置5个采样点, 相邻样点间距为20 m, 每个采样点设置2 m×2 m的样方, 每个样方去掉枯落物层后采用五点法采集0~20 cm的耕层土壤, 剔除土样中的植物根系、石块等杂物后, 将同一研究区域的土样充分混合均匀, 装入密封袋, 迅速带回实验室, 一部分保存于-20℃冰箱, 用于高通量测序; 一部分样品风干, 用于土壤理化指标的测定.
1.3 土壤基本理化指标的测定土壤pH、有机质(OM)、全氮(TN)、碱解氮(AN)、有效磷(AP)和速效钾(AK)的测定参照文献[19]的测定方法; 土壤蔗糖酶(SC)、酸性磷酸酶(ACP)、过氧化氢酶(CAT)和脲酶(URE)活性的测定参照文献[20]的测定方法; 土壤总Hg(THg)和有效态Hg(HCl-Hg)分别采用王水(1+1)消解和0.1 mol ·L-1 HCl浸提, 并用原子荧光光度法进行测定[21].
1.4 土壤微生物测定 1.4.1 高通量测序土壤微生物高通量测序由上海派森诺生物科技股份有限公司完成, 方法如下.
(1) DNA提取 采用E.Z.N.ATMMag-Bind土壤试剂盒(OMEGA M5635-02, 美国)提取土壤总DNA, 同时采用Quantifluor-ST fluorometer荧光分光光度计(E6090, 美国)和1.2%的琼脂糖凝胶电泳检测DNA的浓度及质量.
(2) PCR扩增 细菌的16S rRNA基因V3-V4区域采用引物(338F: 5′-ACTCCTACGGGAGGCAGC A-3′; 806R: 5′-GGACTACHVGGGTWTCTAAT-3′)进行扩增; 真菌的ITS1基因280bp区域采用引物(ITS5F GGAAGTAAAAGTCGTAACAAGG; ITS2RGC TGCGTTCTTCATCGATGC)进行扩增.
(3) 测序 基于16S rDNA序列对PCR产物进行纯化、定量和均一化构建文库并在LabChip上进行质检, 质检合格的文库用Illumina MiSeq进行高通量测序.
1.4.2 序列处理与分析采用DADA2方法对原始测序序列进行引物去除, 质量过滤, 去噪(denoise), 拼接和去嵌合体等步骤.对前述获得的序列构建ASV表(amplicon sequence variant), 得到ASV代表序列(DADA2不以相似度聚类, 只进行去重即相当于以100%相似度聚类, 使用DADA2质控后产生的每个去重序列称为ASV).对于不同类别的序列, 分别采用各自特定的数据库作为ASV分类地位鉴定的模板序列, 如针对细菌的16S rRNA基因, 选用Silva数据库(Release132, http://www.arb-silva.de); 针对真菌ITS序列, 选用Unite数据库(Release 8.0, https://unite.ut.ee/).
1.5 数据处理采用地积累指数法(Igeo)[22]及Hakanson潜在生态风险指数法(Ei)[23]评价土壤Hg的污染状况, 具体计算公式如下:
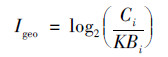
|
(1) |
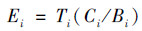
|
(2) |
式中: Ci为土壤样品中重金属i的实测含量(mg ·kg-1); Bi为贵州省土壤重金属i的背景值(mg ·kg-1); K=1.5, 为地壳运动引起的背景值变动系数; Ti为重金属i的毒性系数(THg=40.0).
采用Excel 2013、SPSS 25.0对土壤理化性质和微生物群落组成数据进行处理, 采用单因素方差分析(ANOVA)和多重比较法(Duncan)进行差异显著性分析; 采用RDA分析以及关联网络分析土壤微生物群落与环境因子间的关系; 采用派森诺云平台对相关数据进行作图.
2 结果与讨论 2.1 农田土壤理化因子与Hg污染状况 2.1.1 土壤基本理化指标土壤理化性质是土壤、植被以及周围环境共同作用的结果, 是土壤质量变化与土壤发育最基本的表征和核心研究内容[24].由表 2可知, 4个研究区域土壤pH值差异显著(P < 0.05), 其中, BJ和JK的pH值为7.10~7.54, 为弱碱性; SQ和TR的pH值为5.08~5.98, 为弱酸性.TR的OM含量分别较BJ、SQ和JK提高4.34%、5.52%和1.04%, 但各区域之间差异不显著(P < 0.05).BJ的TN含量分别较SQ和JK提高15.67%和9.93%, 且差异显著(P < 0.05), 与TR之间差异不显著, 但仍提高1.97%.JK的AN含量较SQ提高54.33%, 且差异显著(P < 0.05), 与BJ和TR之间差异不显著, 但仍提高8.70%和4.17%.TR的AP含量分别较BJ和SQ提高268.86%和100.50%, 且差异显著(P < 0.05), 与JK之间差异不显著, 但仍提高10.76%. 4个区域土壤AK含量差异均达显著水平, 其中, JK的AK含量分别较BJ、SQ和TR提高32.00%、53.33%和9.97%(P < 0.05).根据全国第2次土壤普查的养分分级标准发现[25], 研究区域土壤ω(OM)介于20~30 g ·kg-1, 达到3级标准, 为适量水平; ω(TN)介于1~2 g ·kg-1, 达到3级或2级标准, 为适量和丰富水平; ω(AN)介于30~90 mg ·kg-1, 达到5级或4级标准, 为很缺乏和缺乏水平; ω(AP)大于20 mg ·kg-1, 达到2级或1级标准, 为丰富和很丰富水平, ω(AK)大于200 mg ·kg-1, 达到1级标准, 为很丰富水平.根据土壤肥力综合评价标准(integrated fertility index, IFI)[26], BJ和JK的IFI分别为0.76和0.77, 均大于0.75, 属于Ⅰ级水平; SQ和TR的IFI分别为0.64和0.74, 介于0.50~0.75, 属于Ⅱ级水平.
|
|
表 2 土壤基本理化特性1) Table 2 Physical and chemical properties of experimental soil |
2.1.2 土壤Hg污染特征与环境风险
由表 3可知, 研究区域土壤ω(总Hg)为0.22~0.52 mg ·kg-1, 较贵州省土壤背景值(0.11 mg ·kg-1)高100% ~136.36%, 均低于土壤环境质量标准中的风险筛选值和风险管控值(GB 15618-2018).不同区域土壤THg和HCl-Hg含量均存在一定差异, 且土壤总Hg和HCl-Hg含量从高到低均为: SQ>BJ>JK>TR.可见, 土壤HCl-Hg与总Hg含量密切相关, 总Hg的污染状况亦可反映HCl-Hg的污染状况, 明确研究区域土壤Hg的污染状况, 可为当地农作物食品安全生产提供一定的理论支撑.由表 3可以看出, 不同区域土壤Hg的Igeo和Ei指数差异均达显著水平(P < 0.05).根据Igeo分级标准[22], JK和TR的Igeo指数介于0~1, 属于轻度污染; SQ和BJ的Igeo指数介于1~2, 属于偏中度污染.根据Ei分级标准[23], TR的Ei指数介于40~80, 存在中等潜在生态风险; BJ和JK的Ei指数介于80~160, 存在强潜在生态风险; SQ的Ei指数介于160~320, 存在很强潜在生态风险.Yu等[4]和Tang等[8]调查了Hg矿附近农田土壤Hg的污染状况, 结果均表明, Hg属于极强污染范畴(Igeo>5), 存在强或很强潜在生态风险(Ei>160).对比以上研究结果发现, 距离Hg矿区较远的土壤, Igeo指数虽有所降低, 但仍存在一定程度的污染.因此, 建议当地农业及环保部门制定科学合理的土地规划政策和监督机制, 因地制宜建立土壤污染监测、预测和评价系统, 以保证当地农田土壤生态结构、功能稳定性及可持续生产能力.采取必要的钝化修复技术(含硫材料、生物炭等)、叶面阻控技术(含硒叶面剂)和农艺调控(水分调节、深翻)措施等来抑制土壤Hg生物有效性或降低农产品超标风险.
|
|
表 3 土壤Hg污染特征与环境风险 Table 3 Soil Hg pollution characteristics and environmental risks |
2.1.3 土壤酶活性
土壤酶活性是土壤生物活性和土壤肥力的重要指标, 对土壤养分溶解和生物化学过程反应速率等都具有显著促进作用[27], 对重金属污染十分敏感, 常用作判断污染物对生物潜在毒性的手段[28].土壤SC活性表征了土壤有机碳累积与分解转化规律[29]; ACP活性是评价土壤磷素生物转化方向与强度的指标[27]; CAT活性与土壤呼吸作用强度和土壤微生物的生命活动相关, 它在一定程度上反映了土壤微生物学过程的强度[28]; URE活性可以用来表征土壤有机态氮向有效态氮的转化能力和土壤无机氮的供应能力[29].由表 4可知, 不同区域土壤CAT活性差异不显著, SC和ACP活性差异均达显著水平, 其中, BJ的SC和ACP活性均达最大值, SC活性分别较SQ、JK和TR提高127.01%、212.56%和389.71%(P < 0.05); ACP活性分别较SQ、JK和TR提高221.43%、688.14%和97.03%(P < 0.05).JK的URE活性分别较BJ、SQ和TR提高33.33%、88.24%和40.35%, 且差异显著(P < 0.05).结合表 3和表 4发现, 当土壤中Hg污染程度较高时, 酶活性反而有一定程度地下降.如SQ的Hg污染程度最高, 而酶活性却不是最大值, 说明高浓度的Hg会对土壤酶活性产生抑制效应.其机制可能包括: ①Hg与酶分子中的活性部位配位结合, 形成比较稳定的络合物, 产生了与底物的竞争性抑制作用[30]; ②Hg通过抑制土壤微生物的生长和繁殖, 减少其体内酶的合成和分泌, 进而导致土壤酶活性下降[30].
|
|
表 4 土壤酶活性 Table 4 Soil enzyme activity |
2.2 调查区土壤微生物群落结构特征 2.2.1 细菌群落结构特征
群落柱形图可直观反映不同区域土壤优势菌群分布比例, 以及各优势菌群在不同区域土壤中的分布比例.本研究所有供试土壤样品共检测到细菌19门、83纲、330目、714科和2 657属.从门水平的分类来看[图 1(a)], 土壤细菌主要来自变形菌门(Proteobacteria)、放线菌门(Actinobacteria)、酸杆菌门(Acidobacteria)、绿弯菌门(Chloroflexi)、芽单胞菌门(Gemmatimonadetes)、拟杆菌门(Bacteroidetes)、厚壁菌门(Firmicutes)、棒状杆菌门(Rokubacteria)、浮霉菌门(Planctomycetes)和WPS-2. 其中, Proteobacteria、Actinobacteria、Acidobacteria和Chloroflexi占所有微生物总数的84.01%, 是所测土壤的优势菌群, 这与多数重金属污染环境中观察到的细菌群落组成一致.Frossard等[12]也研究发现, Proteobacteria、浮霉菌门(Planctomycetes)、Actinobacteria和Bacteroidetes是长期Hg污染土壤中的优势菌群.Li等[10]、Ma等[31]和Zhao等[32]的调查发现在铅锌冶炼厂周边农田土壤中, 微生物群落通常以Proteobacteria、Actinobacteria、Firmicutes、Acidobacteria和Bacteroidetes为优势菌群.Zeng等[33]、Jacquiod等[34]和Schneider等[35]的研究发现重金属污染土壤中最主要的菌群为Proteobacteria、Actinobacteria、Firmicutes和Chloroflexi, 它们可以耐受高含量的重金属, 如Cd和Zn等.在金矿和铅锌矿开采区周边农田土壤中, Proteobacteria、Bacteroidetes、Acidobacteria、Firmicutes、Gemmatimonadetes和Actinobacteria的分布比例均比较高[36].在本研究中, Gemmatimonadetes、Bacteroidetes和Firmicutes的相对丰度均超过1%, 也是主要的土壤细菌门类, 与上述研究结果一致.在BJ、JK和TR的土壤中, 检测出Rokubacteria, 在SQ和TR的土壤中, 检测出WPS-2.整体而言, 在门分类水平上, 4个区域之间土壤细菌群落的相对丰度存在差异性.针对优势菌群, Proteobacteria差异不明显(P < 0.05); BJ的Actinobacteria显著高于SQ和TR, 但与JK差异不显著(P < 0.05); SQ的Acidobacteria显著高于BJ, 但与JK和TR之间差异不显著(P < 0.05); TR的Chloroflexi显著高于BJ、SQ和JK(P < 0.05)[图 1(b)].Proteobacteria的相对丰度变化为26.40% ~29.71%(SQ>JK>TR>BJ), Actinobacteria为19.25% ~37.22%(BJ>JK>SQ>TR), Acidobacteria为13.18% ~16.90%(SQ>TR>JK>SQ), Chloroflexi为9.00% ~21.16%(TR>BJ>SQ>JK).

|
不同小写字母表示不同采样点同一物种间的显著性差异(P < 0.05) 图 1 不同采样点土壤细菌门水平上群落结构特征 Fig. 1 Soil bacterial community structure at the phylum level in different sampling soils |
本研究所有供试土壤样品共检测到真菌15门、27纲、30目、25科和65属.从门水平的分类来看[图 2(a)], 土壤真菌主要来自子囊菌门(Ascomycota)、担子菌门(Basidiomycota)和被孢霉门(Mortierellomycota), 且这3种菌群占所有微生物总数的75.60%, 是土壤中优势菌群, 这与多数重金属污染环境中观察到的真菌群落组成一致.Frossard等[12]的研究发现, Ascomycota和Basidiomycota是长期Hg污染土壤中的优势真菌菌群. Chun等[37]的研究发现, Mortierellomycota和镰刀霉(Fusarium)是废弃矿场周围土壤中的优势真菌菌群.Lin等[38]的研究发现, Ascomycota、Basidiomycota、壶菌门(Chytridiomycota)、球囊菌门(Glomeromycota)、接合菌门(Zygomycota)和红藻门(Rozellomycota)在混合重金属污染(Cu、Zn、Cd、Pb、Ni和Cr)土壤中含有较高的比例.陈建文等[39]的研究表明, Ascomycota、Basidiomycota和Mortierellomycota是铜尾矿坝及其周边土壤中的优势真菌菌群.在JK和TR的土壤中, 检测出Glomeromycota, 在SQ的土壤中, 检测出Chytridiomycota和梳霉门(Kickxellomycota).不同区域土壤真菌优势菌群相对丰度存在差异, 其中, BJ的Ascomycota显著高于SQ和JK, 但与TR差异不显著(P < 0.05); SQ的Basidiomycota显著高于BJ、JK和TR(P < 0.05); TR的Mortierellomycota显著高于BJ、SQ和JK(P < 0.05)[图 2(b)].Ascomycota的相对丰度变化为54.04% ~76.29%(BJ>TR>SQ>JK); Basidiomycota为5.04% ~17.38%(SQ>BJ>TR>JK); Mortierellomycota为1.16% ~7.00%(TR>SQ>JK>BJ).

|
不同小写字母表示不同采样点同一物种间的显著性差异(P < 0.05) 图 2 不同采样点土壤真菌门水平上群落结构特征 Fig. 2 Soil fungal community structure at the phylum level in different sampling soils |
土壤微生物的群落结构组成与其所处的土壤环境有关, 微生物对土壤环境因子的适应能力决定了其群落结构组成状况[40].土壤微生物群落结构与环境因子间的RDA分析以及关联网络可以预测土壤微生物群落结构与环境因子之间的关系.
由图 3可以看出, 环境因子对土壤细菌群落结构的解释量为93.6%, 其中, 前两个排序轴的解释变量分别为43.84%和26.41%.根据箭头长度可以看出, pH、URE、SC和CAT与细菌种类分布相关性较好.采用envfit函数检验每个环境因子的显著性, 结果显示pH(r2=0.637, Pr=0.003)、红色箭头表示环境因子, 箭头长度表示环境因子与细菌群落结构相关性的强弱; 蓝色箭头表示细菌类型, 箭头指向与环境因子箭头同向表示高度正相关SC(r2=0.609, Pr=0.014)和CAT(r2=0.518, Pr=0.047)对土壤细菌群落结构组成的影响达显著水平(P < 0.05).以上结果说明, pH是土壤细菌群落结构的主要驱动因子, 这与我国长白山[41]和英国农田[42]土壤微生物群落结构的主要驱动因子一致.其原因是pH可以直接或间接地和其它土壤变量, 如土壤含水量、养分有效性等相关作用, 从而影响土壤微生物群落结构的组成和分布状况[43].此外, 以上结果还说明SC和CAT活性是土壤细菌群落结构的主要环境因子.程济南等[44]的研究发现, 土壤URE、SC、ACP活性是甘肃典型高寒草原土壤细菌群落结构的主要环境因子, SC活性是真菌群落结构的主要环境因子.姚兰等[43]的研究发现, 土壤URE和SC活性是安徽黄山不同海拔梯度土壤细菌群落结构的主要环境因子.Xiao等[45]的研究发现, 土壤ACP活性是福建省天宝岩国家级自然保护区长苞铁杉林土壤细菌群落结构的主要环境因子.综上所述, 生态环境不同, 土壤微生物群落结构与酶活性之间的关系也不同, 其主要与气候、植被和土壤理化性质的变化有关.

|
图 3 土壤细菌群落结构与环境因子间的RDA分析 Fig. 3 RDA analysis of soil bacterial community structure and environmental factors |
利用相关性网络进一步分析不同环境因子与丰富度前10土壤细菌群落结构之间的关系, 结果发现(图 4), Proteobacteria与CAT、Acidobacteria与SC、Rokubacteria与pH和URE呈极显著正相关(P < 0.01); Bacteroidetes与AK呈显著正相关(P < 0.05); WPS-2与pH和URE呈极显著负相关(P < 0.01); Acidobacteria与AP、Actinobacteria与pH、SC和ACP、Bacteroidetes与HCl-Hg、Planctomycetes与pH呈显著负相关(P < 0.05).对相关性网络分析结果进行关系对去重和数据筛选后发现, pH是Acidobacteria、Rokubacteria、WPS-2和Planctomycetes群落结构的主要驱动因子, 这与Rousk等[42]和Guo等[46]的研究结果相符.AP和SC活性均是Acidobacteria和WPS-2群落结构的主要环境因子, 这与Shen等[41]和姚兰等[43]的研究结果一致.ACP活性是Chloroflexi群落结构的主要环境因子.CAT活性是Proteobacteria群落结构的主要环境因子, 这与刘爽等[40]的研究结果相符.HCl-Hg是Firmicutes群落结构的主要环境因子, 这与Liu等[11]的研究结果一致, 其原因主要是Firmicutes具有高繁殖能力, 从而能够在受到Hg的干扰后, 快速占据生态位.

|
线条粗细表示相关性强弱 图 4 土壤细菌群落结构与环境因子间的相关性网络分析 Fig. 4 Correlation network analysis of soil bacterial community structure and environmental factors |
由图 5可以看出, 环境因子对土壤真菌群落结构的解释量为81.7%, 其中, 前两个排序轴的解释变量分别为28.24%和18.13%.根据箭头长度可以看出, AN、ACP、pH、URE、HCl-Hg、TN、AK和AP与真菌种群分布相关性较好.Envfit函数检验结果显示AN(r2=0.804, Pr=0.002)、ACP(r2=0.658, Pr=0.005)、URE(r2=0.603, Pr=0.012)、AK(r2=0.506, Pr=0.039)、pH(r2=0.491, Pr=0.044)和HCl-Hg(r2=0.486, Pr=0.046)对土壤真菌群落结构组成的影响达显著水平(P < 0.05).以上分析结果说明pH、AN、AK、HCl-Hg、ACP和URE活性是土壤真菌群落结构的主要环境因子.针对土壤pH, Högberg等[47]的研究发现, pH是瑞典北部森林土壤真菌群落结构的主要驱动因子; Zhang等[48]的研究发现, pH是北极圈群岛土壤真菌群落结构的主要驱动因子, 与本研究结果一致.其原因是pH可以通过改变土壤环境因子而影响真菌群落结构.针对土壤AN和AK, 代红翠等[49]的研究现, AN和AK是病理营养型真菌群落结构变化的主要驱动因子; 裴振等[50]的研究表明, AN是黄河三角洲盐碱地土壤真菌群落结构的主要驱动因子; Zhang等[51]的研究发现, AK是高寒地区土壤真菌群落组成的主要驱动因子.其原因可能是: ①有机肥或化肥的施用提高了土壤中AN和AK的含量, 为真菌的生存和繁殖提供了所需的营养和能源物质.②真菌在土壤N元素循环中具有重要的作用, N素的添加会影响微生物N循环过程, 如硝化作用和固氮; 同时, 土壤中N含量的变化也会通过影响地上作物的多样性和优势度而对土壤真菌群落结构产生影响[52].针对土壤HCl-Hg, 其可表征土壤Hg的生物有效性, 主要包括生物可利用性和迁移能力, 了解Hg矿周边农田土壤HCl-Hg与微生物群落结构的关系, 可为当地土壤重金属环境风险管控和土壤修复策略的制定等提供理论参考.Frossard等[12]的研究结果显示, 长期Hg污染会对土壤细菌和真菌群落结构产生明显的影响; Liu等[53]的研究也表明, 土壤Hg的积累会显著影响土壤微生物群落结构, 其原因是在长期Hg污染的情况下, 土壤微生物群落对其生存环境做出了相应的变化, 已经进化出在不利环境中生存的策略.在本研究中, 土壤真菌对Hg污染的响应优于土壤细菌.

|
红色箭头表示环境因子, 箭头长度表示环境因子与真菌群落结构相关性的强弱; 蓝色箭头表示真菌类型, 箭头指向与环境因子箭头同向表示高度正相关 图 5 土壤真菌群落结构与环境因子间的RDA分析 Fig. 5 RDA analysis of soil fungal community structure and environmental factors |
利用关联网络进一步分析不同环境因子与丰富度前10土壤真菌群落结构之间的关系. 结果发现(图 6): Ascomycota与ACP, Basidiomycota与HCl-Hg呈显著正相关(P < 0.05); Basidiomycota与pH、AK、AN和URE, Glomeromycota与ACP, Chytridiomycota与pH和AN呈极显著负相关(P < 0.01); Chytridiomycota与URE, Basidiobolomycota与HCl-Hg, Kickxellomycota与AK和AN, Mucoromycota与pH和AN呈显著负相关(P < 0.05).对相关性网络分析结果进行关系对去重和数据筛选后发现, pH是Chytridiomycota、Mucoromycota、Basidiomycota和Ascomycota群落结构的主要驱动因子, 这与Liu等[11]的研究结果相符.TN是Rozellomycota群落结构的主要驱动因子; URE活性是Kickxellomycota、Chytridiomycota、Mucoromycota和Basidiomycota群落结构的主要环境因子, 这与韩士昌[54]的研究结果相符.AP是Kickxellomycota和Mortierellomycota群落结构的主要驱动因子, 这与Liu等[55]的研究结果相符, 可能是因为P是微生物细胞结构的重要组成元素, 从而导致微生物表现出对P养分的依赖.AN、AK和SC活性均是Kickxellomycota群落结构的主要环境因子, 这与程济南等[44]和代红翠等[49]的研究结果相符.ACP是Ascomycota、Glomeromycota和Rozellomycota群落结构的主要环境因子.以上结果说明土壤真菌群落结构变化与土壤基本理化性质、酶活性和HCl-Hg含量密切相关.

|
线条粗细表示相关性强弱 图 6 土壤真菌群落结构与环境因子间的相关性网络分析 Fig. 6 Correlation network analysis of soil fungal community structure and environmental factors |
(1) 研究区农田土壤存在一定程度的Hg污染, 其中, JK和TR属于轻度污染, SQ和BJ属于偏中度污染; TR存在中等潜在生态风险, BJ和JK存在强潜在生态风险, SQ存在很强潜在生态风险.
(2) 研究区农田土壤细菌优势菌群主要包括Proteobacteria、Actinobacteria、Acidobacteria和Chloroflexi; 真菌优势菌群主要包括Ascomycota、Basidiomycota和Mortierellomycota.
(3) pH、SC和CAT活性是土壤细菌群落结构组成的主要环境因子, 影响Proteobacteria、Acidobacteria、Actinobacteria、Chloroflexi、Firmicutes、Rokubacteria和Planctomycetes分布; pH、AN、AK、HCl-Hg、ACP和URE活性是土壤真菌群落结构组成的主要环境因子, 影响土壤Ascomycota、Basidiomycota、Mortierellomycota、Glomeromycota、Chytridiomycota、Rozellomycota、Kickxellomycota和Mucoromycota分布.
| [1] | Yuan C S, Jhang Y M, Ie I R, et al. Exploratory investigation on spatiotemporal variation and source identification of atmospheric speciated mercury surrounding the Taiwan Strait[J]. Atmospheric Pollution Research, 2021, 12(3): 54-64. DOI:10.1016/j.apr.2021.01.015 |
| [2] | Liu Y, Lin C J, Yuan W, et al. Translocation and distribution of mercury in biomasses from subtropical forest ecosystems: evidence from stable mercury isotopes[J]. Acta Geochimica, 2021, 40(1): 42-50. DOI:10.1007/s11631-020-00441-3 |
| [3] | Wu Z Y, Zhang L N, Xia T X, et al. Heavy metal pollution and human health risk assessment at mercury smelting sites in Wanshan district of Guizhou Province, China[J]. RSC Advances, 2020, 10(39): 23066-23079. DOI:10.1039/D0RA01004A |
| [4] | Yu G, Chen F, Zhang H L, et al. Pollution and health risk assessment of heavy metals in soils of Guizhou, China[J]. Ecosystem Health and Sustainability, 2021, 7(1). DOI:10.1080/20964129.2020.1859948 |
| [5] |
尹德良, 何天容, 罗光俊, 等. 汞矿区土壤汞的影响因素及修复技术进展[J]. 环境科学导刊, 2014, 33(1): 62-67. Yin D L, He T R, Luo G J, et al. The affecting factors of mercury in soil and progress of remediation technologies in mercury mines[J]. Environmental Science Survey, 2014, 33(1): 62-67. DOI:10.3969/j.issn.1673-9655.2014.01.013 |
| [6] |
苟体忠, 阮运飞. 万山汞矿区土壤重金属污染特征及来源解析[J]. 化工环保, 2020, 40(3): 336-341. Gou T Z, Ruan Y F. Characteristics and source of heavy metals in contaminated soil in Wanshan mercury mine area[J]. Environmental Protection of Chemical Industry, 2020, 40(3): 336-341. DOI:10.3969/j.issn.1006-1878.2020.03.018 |
| [7] |
高令健, 毛康, 张伟, 等. 贵州万山汞矿区稻田土壤汞的分布及污染特征[J]. 矿物岩石地球化学通报, 2021, 40(1): 148-154. Gao L J, Mao K, Zhang W, et al. Temporal and spatial distribution and pollution characteristics of mercury in paddy soils of the Wanshan mercury mining area, Guizhou Province[J]. Bulletin of Mineralogy, Petrology and Geochemistry, 2021, 40(1): 148-154. |
| [8] | Tang X C, Wu X L, Xia P H, et al. Health risk assessment of heavy metals in soils and screening of accumulating plants around the Wanshan mercury mine in Northeast Guizhou Province, China[J]. Environmental Science and Pollution Research, 2021, 28(35): 48837-48850. DOI:10.1007/s11356-021-14145-5 |
| [9] | Shuaib M, Azam N, Bahadur S, et al. Variation and succession of microbial communities under the conditions of persistent heavy metal and their survival mechanism[J]. Microbial Pathogenesis, 2021, 150. DOI:10.1016/j.micpath.2020.104713 |
| [10] | Li B Q, Xu R, Sun X X, et al. Microbiome-environment interactions in antimony-contaminated rice paddies and the correlation of core microbiome with arsenic and antimony contamination[J]. Chemosphere, 2021, 263. DOI:10.1016/j.chemosphere.2020.128227 |
| [11] | Liu Y R, Delgado-Baquerizo M, Bi L, et al. Consistent responses of soil microbial taxonomic and functional attributes to mercury pollution across China[J]. Microbiome, 2018, 6(1). DOI:10.1186/s40168-018-0572-7 |
| [12] | Frossard A, Donhauser J, Mestrot A, et al. Long-and short-term effects of mercury pollution on the soil microbiome[J]. Soil Biology and Biochemistry, 2018, 120: 191-199. DOI:10.1016/j.soilbio.2018.01.028 |
| [13] | Jiang L F, Song M K, Yang L, et al. Exploring the influence of environmental factors on bacterial communities within the rhizosphere of the Cu-tolerant plant, Elsholtzia splendens[J]. Scientific Reports, 2016, 6(1). DOI:10.1038/srep36302 |
| [14] |
黄健, 朱旭炎, 陆金, 等. 狮子山矿区不同土地利用类型对土壤微生物群落多样性的影响[J]. 环境科学, 2019, 40(12): 5550-5560. Huang J, Zhu X Y, Lu J, et al. Effects of different land use types on microbial community diversity in the Shizishan Mining area[J]. Environmental Science, 2019, 40(12): 5550-5560. |
| [15] | Kenarova A, Radeva G, Traykov I, et al. Community level physiological profiles of bacterial communities inhabiting uranium mining impacted sites[J]. Ecotoxicology and Environmental Safety, 2014, 100: 226-232. DOI:10.1016/j.ecoenv.2013.11.012 |
| [16] | Stemmer M, Watzinger A, Blochberger K, et al. Linking dynamics of soil microbial phospholipid fatty acids to carbon mineralization in a 13 C natural abundance experiment: impact of heavy metals and acid rain[J]. Soil Biology and Biochemistry, 2007, 39(12): 3177-3186. DOI:10.1016/j.soilbio.2007.07.013 |
| [17] |
尹亚丽, 王玉琴, 李世雄, 等. 三江源区退化高寒草甸土壤微生物群落季节特征研究[J]. 生态环境学报, 2018, 27(10): 1791-1800. Yin Y L, Wang Y Q, Li S X, et al. Study on seasonal various of characteristics of soil microbial community at degraded alpine meadows in the Three-River-Source area[J]. Ecology and Environmental Sciences, 2018, 27(10): 1791-1800. |
| [18] |
朱永官, 彭静静, 韦中, 等. 土壤微生物组与土壤健康[J]. 中国科学: 生命科学, 2021, 51(1): 1-11. Zhu Y G, Peng J J, Wei Z, et al. Linking the soil microbiome to soil health[J]. Scientia Sinica Vitae, 2021, 51(1): 1-11. |
| [19] | 鲍士旦. 土壤农化分析[M]. (第三版). 北京: 中国农业出版社, 2000. |
| [20] | 关松荫. 土壤酶及其研究法[M]. 北京: 农业出版社, 1986. |
| [21] |
陈芬, 余高, 吴涵茜, 等. 中药渣生物有机肥对镉-汞复合污染土壤的钝化效果[J]. 浙江大学学报(农业与生命科学版), 2020, 46(6): 737-747. Chen F, Yu G, Wu H Q, et al. Effects of bio-organic fertilizer made from Chinese traditional herb residues on heavy metal passivation in Cd and Hg compound-contaminated soils[J]. Journal of Zhejiang University (Agriculture and Life Sciences), 2020, 46(6): 737-747. |
| [22] | Müller G. Index of geoaccumulation in sediments of the Rhine River[J]. GeoJournal, 1969, 2(3): 108-118. |
| [23] | Hakanson L. An ecological risk index for aquatic pollution control. a sedimentological approach[J]. Water Research, 1980, 14(8): 975-1001. DOI:10.1016/0043-1354(80)90143-8 |
| [24] | Nadal-Romero E, Cammeraat E, Pérez-Cardiel E, et al. Effects of secondary succession and afforestation practices on soil properties after cropland abandonment in humid Mediterranean mountain areas[J]. Agriculture, Ecosystems & Environment, 2016, 228: 91-100. |
| [25] |
侯建伟, 邢存芳, 邓晓梅, 等. 不同秸秆生物炭对黄壤理化性质及综合肥力的影响[J]. 西北农林科技大学学报(自然科学版), 2019, 47(11): 49-59. Hou J W, Xing C F, Deng X M, et al. Effect of straw biochar on yellow soil physicochemical properties and comprehensive fertility[J]. Journal of Northwest A&F University (Natural Science Edition), 2019, 47(11): 49-59. |
| [26] |
杨皓, 胡继伟, 黄先飞, 等. 喀斯特地区金刺梨种植基地土壤肥力研究[J]. 水土保持研究, 2015, 22(3): 50-55. Yang H, Hu J W, Huang X F, et al. Study on soil fertility of Rosa sterilis S.D. Shi planting bases located in karst areas[J]. Research of Soil and Water Conservation, 2015, 22(3): 50-55. |
| [27] |
杨思德, 常兴平, 潘政, 等. 蚯蚓对金霉素污染土壤酶活性和微生物群落的影响[J]. 农业环境科学学报, 2021, 40(6): 1268-1280. Yang S D, Chang X P, Pan Z, et al. Effects of earthworms on soil enzyme activities and microbial communities of chlortetracycline-contaminated soils[J]. Journal of Agro-Environment Science, 2021, 40(6): 1268-1280. |
| [28] |
郭星亮, 谷洁, 陈智学, 等. 铜川煤矿区重金属污染对土壤微生物群落代谢和酶活性的影响[J]. 应用生态学报, 2012, 23(3): 798-806. Guo X L, Gu J, Chen Z X, et al. Effects of heavy metals pollution on soil microbial communities metabolism and soil enzyme activities in coal mining area of Tongchuan, Shaanxi Province of Northwest China[J]. Chinese Journal of Applied Ecology, 2012, 23(3): 798-806. |
| [29] |
张丽琼, 郝明德, 臧逸飞, 等. 苜蓿和小麦长期连作对土壤酶活性及养分的影响[J]. 应用生态学报, 2014, 25(11): 3191-3196. Zhang L Q, Hao M D, Zang Y F, et al. Effects of continuous cropping of wheat and alfalfa on soil enzyme activities and nutrients[J]. Chinese Journal of Applied Ecology, 2014, 25(11): 3191-3196. |
| [30] |
邓霞, 崔东, 王兴磊, 等. 伊犁煤矿土壤重金属累积对土壤酶活性的影响[J]. 环境化学, 2018, 37(7): 1554-1564. Deng X, Cui D, Wang X L, et al. Effects of heavy metal accumulation on soil enzyme activities in Yili coal mines[J]. Environmental Chemistry, 2018, 37(7): 1554-1564. |
| [31] | Ma Y J, Wang Y T, Chen Q, et al. Assessment of heavy metal pollution and the effect on bacterial community in acidic and neutral soils[J]. Ecological Indicators, 2020, 117. DOI:10.1016/j.ecolind.2020.106626 |
| [32] | Zhao X Q, Huang J, Lu J, et al. Study on the influence of soil microbial community on the long-term heavy metal pollution of different land use types and depth layers in mine[J]. Ecotoxicology and Environmental Safety, 2019, 170: 218-226. DOI:10.1016/j.ecoenv.2018.11.136 |
| [33] | Zeng P, Guo Z H, Xiao X Y, et al. Effects of tree-herb co-planting on the bacterial community composition and the relationship between specific microorganisms and enzymatic activities in metal(loid)-contaminated soil[J]. Chemosphere, 2019, 220: 237-248. DOI:10.1016/j.chemosphere.2018.12.073 |
| [34] | Jacquiod S, Cyriaque V, Riber L, et al. Long-term industrial metal contamination unexpectedly shaped diversity and activity response of sediment microbiome[J]. Journal of Hazardous Materials, 2018, 344: 299-307. DOI:10.1016/j.jhazmat.2017.09.046 |
| [35] | Schneider A R, Gommeaux M, Duclercq J, et al. Response of bacterial communities to Pb smelter pollution in contrasting soils[J]. Science of the Total Environment, 2017, 605-606: 436-444. DOI:10.1016/j.scitotenv.2017.06.159 |
| [36] | Guo H H, Nasir M, Lv J L, et al. Understanding the variation of microbial community in heavy metals contaminated soil using high throughput sequencing[J]. Ecotoxicology and Environmental Safety, 2017, 144: 300-306. DOI:10.1016/j.ecoenv.2017.06.048 |
| [37] | Chun S J, Kim Y J, Cui Y S, et al. Ecological network analysis reveals distinctive microbial modules associated with heavy metal contamination of abandoned mine soils in Korea[J]. Environmental Pollution, 2021, 289. DOI:10.1016/j.envpol.2021.117851 |
| [38] | Lin Y B, Ye Y M, Hu Y M, et al. The variation in microbial community structure under different heavy metal contamination levels in paddy soils[J]. Ecotoxicology and Environmental Safety, 2019, 180: 557-564. DOI:10.1016/j.ecoenv.2019.05.057 |
| [39] |
陈建文, 张红, 李君剑, 等. 铜尾矿坝及其周边土壤真菌群落结构与功能多样性[J]. 环境科学, 2021, 42(4): 2056-2065. Chen J W, Zhang H, Li J J, et al. Soil fungal community structure and functional diversity in a copper tailing dam and its surrounding areas[J]. Environmental Science, 2021, 42(4): 2056-2065. |
| [40] |
刘爽, 王雅, 刘兵兵, 等. 晋西北不同土地管理方式对土壤碳氮、酶活性及微生物的影响[J]. 生态学报, 2019, 39(12): 4376-4389. Liu S, Wang Y, Liu B B, et al. Effects of different land management practices on soil carbon and nitrogen, enzyme activities, and microbial diversities northwest of Shanxi[J]. Acta Ecologica Sinica, 2019, 39(12): 4376-4389. |
| [41] | Shen C C, Xiong J B, Zhang H Y, et al. Soil pH drives the spatial distribution of bacterial communities along elevation on Changbai Mountain[J]. Soil Biology and Biochemistry, 2013, 57: 204-211. DOI:10.1016/j.soilbio.2012.07.013 |
| [42] | Rousk J, Brookes P C, Bååth E. Contrasting soil pH effects on fungal and bacterial growth suggest functional redundancy in carbon mineralization[J]. Applied and Environmental Microbiology, 2009, 75(6): 1589-1596. DOI:10.1128/AEM.02775-08 |
| [43] |
姚兰, 胡立煌, 张焕朝, 等. 黄山土壤细菌群落和酶活性海拔分布特征[J]. 环境科学, 2019, 40(2): 859-868. Yao L, Hu L H, Zhang H C, et al. Elevational distribution characteristics of soil bacterial community and enzyme activities in Mount Huangshan[J]. Environmental Science, 2019, 40(2): 859-868. DOI:10.3969/j.issn.1000-6923.2019.02.051 |
| [44] |
程济南, 金辉, 许忠祥, 等. 甘肃典型高寒草原退化植物瑞香狼毒对根际土壤微生物群落的影响研究[J]. 微生物学报, 2021, 61(11): 3686-3704. Cheng J N, Jin H, Xu Z X, et al. Effects of degraded plant Stellera chamaejasme L. on the rhizosphere soil microbial communities in typical alpine grassland, Gansu[J]. Acta Microbiologica Sinica, 2021, 61(11): 3686-3704. |
| [45] | Xiao S H, You H M, You W B, et al. Rhizosphere and bulk soil enzyme activities in a Nothotsuga longibracteata forest in the Tianbaoyan National Nature Reserve, Fujian Province, China[J]. Journal of Forestry Research, 2017, 28(3): 521-528. DOI:10.1007/s11676-016-0334-y |
| [46] | Guo Q X, Yan L J, Korpelainen H, et al. Plant-plant interactions and N fertilization shape soil bacterial and fungal communities[J]. Soil Biology and Biochemistry, 2019, 128: 127-138. DOI:10.1016/j.soilbio.2018.10.018 |
| [47] | Högberg M N, Yarwood S A, Myrold D D. Fungal but not bacterial soil communities recover after termination of decadal nitrogen additions to boreal forest[J]. Soil Biology and Biochemistry, 2014, 72: 35-43. DOI:10.1016/j.soilbio.2014.01.014 |
| [48] | Zhang T, Wang N F, Liu H Y, et al. Soil pH is a key determinant of soil fungal community composition in the Ny-Ålesund region, Svalbard (High Arctic)[J]. Frontiers in Microbiology, 2016, 7. DOI:10.3389/fmicb.2016.00227 |
| [49] |
代红翠, 张慧, 薛艳芳, 等. 不同耕作和秸秆还田下褐土真菌群落变化特征[J]. 中国农业科学, 2019, 52(13): 2280-2294. Dai H C, Zhang H, Xue Y F, et al. Response of fungal community and function to different tillage and straw returning methods[J]. Scientia Agricultura Sinica, 2019, 52(13): 2280-2294. DOI:10.3864/j.issn.0578-1752.2019.13.008 |
| [50] |
裴振, 孔强, 郭笃发. 盐生植被演替对土壤微生物碳源代谢活性的影响[J]. 中国环境科学, 2017, 37(1): 373-380. Pei Z, Kong Q, Guo D F. Effect of succession of halophytic vegetation on soil microbial carbon metabolic activity[J]. China Environmental Science, 2017, 37(1): 373-380. |
| [51] | Zhang X F, Xu S J, Li C M, et al. The soil carbon/nitrogen ratio and moisture affect microbial community structures in alkaline permafrost-affected soils with different vegetation types on the Tibetan plateau[J]. Research in Microbiology, 2014, 165(2): 128-139. DOI:10.1016/j.resmic.2014.01.002 |
| [52] |
乔沙沙, 周永娜, 柴宝峰, 等. 关帝山森林土壤真菌群落结构与遗传多样性特征[J]. 环境科学, 2017, 38(6): 2502-2512. Qiao S S, Zhou Y N, Chai B F, et al. Characteristics of fungi community structure and genetic diversity of forests in Guandi Mountains[J]. Environmental Science, 2017, 38(6): 2502-2512. |
| [53] | Liu Y R, Johs A, Bi L, et al. Unraveling microbial communities associated with methylmercury production in paddy soils[J]. Environmental Science & Technology, 2018, 52(22): 13110-13118. |
| [54] |
韩士昌. 外源碳氮添加对土壤生物学活性的影响及外生菌根真菌在氮转化中的作用[D]. 济南: 济南大学, 2017. Han S C. Effects of exogenous carbon and nitrogen addition on soil biological activity and the role of ectomycorrhizal fungi in nitrogen transformation[D]. Ji'nan: University of Jinan, 2017. |
| [55] | Liu Z F, Fu B J, Zheng X X, et al. Plant biomass, soil water content and soil N: P ratio regulating soil microbial functional diversity in a temperate steppe: a regional scale study[J]. Soil Biology and Biochemistry, 2010, 42(3): 445-450. DOI:10.1016/j.soilbio.2009.11.027 |