 2022, Vol. 43
2022, Vol. 43

2. 糙米品质安全控制湖南省工程实验室, 长沙 410004
 , HUO Yang1 , ZHOU Hang1,2
, HUO Yang1 , ZHOU Hang1,2 , QIU Yin-yan1 , ZENG Peng1,2 , GU Jiao-feng1,2 , LIAO Bo-han1,2
, QIU Yin-yan1 , ZENG Peng1,2 , GU Jiao-feng1,2 , LIAO Bo-han1,2
2. Hunan Engineering Laboratory for Control of Rice Quality and Safety, Changsha 410004, China
镉(cadmium, Cd)是一种具有高毒性的重金属, 是土壤重金属污染的主要元素之一[1, 2].土壤Cd污染主要来源于工矿业废渣和废水的排放, 以及农药和化肥的不当使用[3].稻米Cd含量超标一直是公众关注的焦点.为了消除或减少土壤Cd污染对水稻生产以及人类健康的负面影响, 已有学者对必需营养元素影响土壤-水稻系统中Cd的吸收、迁移、转运过程开展了研究, 如水稻Cd累积受到铁、锰、钙、锌和硅等元素的影响[4~8], 这些元素可与Cd竞争相关转运蛋白, 减少水稻中Cd的累积[9~12].铁和锰能促进水稻根表铁膜的形成, 影响根部Cd吸收[13, 14], 钙可与土壤溶液中的Cd2+在水稻根表面竞争吸收[15], 锌是Cd在水稻根系的主要竞争阳离子[16].
磷(phosphorus, P)是植物生长的必需元素[17], 是ATP、RNA、DNA和磷脂等基本生物分子的主要组成部分, 几乎影响着植物的每一个发育和生化过程[18].尽管土壤中P总含量很高, 但生物可利用的无机磷酸盐缺乏限制了农业生产, 因此农业上常施用P肥增加水稻产量, 改善稻米品质.然而, 过量施用P肥会导致P的流失和富营养化[19].有研究表明, P对水稻体内Cd的吸收、转运、分布和累积存在促进或抑制效果[20, 21]. Adhikari等[22]的研究发现施用磷酸钙[Ca3 (PO4)2]显著降低了水稻植株Cd含量, 并提高了鲜重、根重、株高和叶绿素含量. Lee等[23]在Cd污染稻田土壤中施用磷酸二氢钾(KH2PO4), 结果发现, 随着P肥施用量的增加(0~390 kg·hm-2), 水稻籽粒、茎和根中Cd含量显著降低.然而, 有研究表明, 在缺P和富P条件下, Cd在水稻中的含量没有明显变化[24], 甚至外施P导致水稻Cd累积量的增加[25].如Liu等[26]的研究表明增施外源P可显著增加水稻幼苗芽和根中Cd含量.以上研究表明, 在Cd污染耕地中P盈亏对水稻Cd吸收转运的影响还不清晰.本文通过水培试验, 研究Cd胁迫下, 缺P和富P条件下水稻Cd积累和转运的影响, 以期为Cd污染稻田科学合理施用P肥提供理论依据.
1 材料与方法 1.1 供试材料供试水稻品种为湖南省常规晚籼稻黄华占, 水稻种子由湖南亚华种业有限公司提供.试验前, 将颗粒饱满的水稻种子用10%的H2 O2溶液表面灭菌15 min, 以去离子水洗涤5次后浸泡24 h, 覆盖纱布于(30±0.5)℃的暗室内培养48 h.萌发后的幼苗移入1/4强度的木村B培养液[27], 在30℃/20℃(昼/夜)温度周期, 相对湿度80%, 14 h/10 h(昼/夜)光照周期环境下, 培养至三叶一心水稻幼苗.
1.2 试验设计水稻水培试验通过向营养液中添加氯化镉(CdCl2·2.5H2 O, 分析纯)模拟Cd胁迫环境, ρ(Cd)为0.02 mg·L-1(以Cd元素计).在水稻营养液为缺P[营养液中ρ(P)≤6.0 mg·L-1]和富P[营养液中ρ(P)≥12.0 mg·L-1]两种状态[28]中进行Cd胁迫处理, 其中缺P状态下营养液中设置3个P处理, ρ(P)分别为1.5、3.0和6.0 mg·L-1; 富P状态下营养液中设置3个P处理, ρ(P)分别为12.0、24.0和48.0 mg·L-1.营养液中P的添加均为NaH2PO4(分析纯), 以P元素计.试验共6个处理, 均重复3次, 共计18盆, 各试验处理编号分别记为P1.5、P3、P6、P12、P24和P48.
2019年7月24日, 挑选长势一致的水稻幼苗, 以白色海绵包裹住茎部底端, 固定于黑色泡沫板(一板三穴, 一穴二株), 移栽至含有12 L木村B营养液的圆柱形黑色塑料桶中, 按照试验设计分别在营养液中添加Cd和不同浓度的P.水稻培养过程中每3 d换一次营养液, 以1.6 mol·L-1的HNO3和0.1 mol·L-1的NaOH调节营养液pH至5.5(弱酸性为水稻生长最适pH范围)[29], 同时防止营养液中生成沉淀[30], 培养至水稻成熟.本试验开展场地为中南林业科技大学露天盆栽试验区, 水稻生长光热条件为自然条件, 根据常规农艺措施对水稻进行喷施农药、除虫和追肥.
1.3 样品采集2019年11月17日, 采集成熟水稻样品, 先后用清水和去离子水多次洗涤至无营养液残留, 将水稻根、茎、叶、谷壳和糙米分离并分装, 留取一部分水稻鲜叶于4℃下冷藏保存, 待测光合色素含量.水稻各部位于通风处晾干, 在鼓风干燥箱中于105℃杀青30 min, 于70℃烘干至恒重.将烘干后的水稻各部位样品称干重, 粉碎, 置于塑料密封袋中保存备用.
1.4 样品分析与测定水稻各部位Cd含量采用干灰化法消解[31], 同时做3组空白试验, 消解液中Cd浓度以电感耦合等离子体发射光谱仪(ICP 6300, ThermoFisher)测定, 以湖南大米[GBW 10045 (GSB-23)]进行质量控制分析, Cd的回收率为94.9% ~103.8%.水稻各部位P含量采用湿式消解钼锑抗比色法[32]、以紫外可见分光光度计(UV2600, UNIC)在波长为700 nm测定, P回收率为97.0% ~99.5%.叶绿素a、叶绿素b和类胡萝卜素以紫外可见光光度计分别在665、647和470 nm波长下测定.
1.5 数据分析试验数据以Office Excel 2019和IBM SPSS 26进行分析, 所有数据均以OriginPro 9.0绘图.表格数据表示为平均值±标准偏差(n=3), 同列数据不同字母表示不同处理间差异显著(P<0.05).采用单因素ANOVA中Duncan多重比较法分析各处理差异(P<0.05). Pearson线性相关分析用于调查数据之间的关系, 以两个显著性水平(0.05和0.01)呈现结果.
水稻各部位的Cd积累量由以下公式计算:
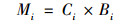
|
式中, C为各部位Cd含量, mg·kg-1; B为各部位干重, g; M为各部位Cd累积量, μg; i表示水稻各部位, 根、茎、叶、谷壳和糙米.
水稻各部位间转运能力以转运系数(translocation factor, TF)表示, 即后一部位Cd含量(C)与前一部位Cd含量的比值, 如TF根-茎=C茎/C根.
2 结果与分析 2.1 水稻缺P和富P对水稻生长和叶片光合色素的影响由表 1可知, Cd胁迫下水稻缺P和富P状况对其各部位生物量及总生物量的影响不显著(P<0.05).在水稻缺P条件下, 随ρ(P)的增加(1.5~6.0 mg·L-1), 各处理水稻各部位生物量影响不显著; 而在水稻富P条件下, 随ρ(P)增加(12.0~48.0 mg·L-1), 各处理水稻各部位生物量变化表现为先增加后减少, 且P24>P48>P12.糙米生物量是评估水稻产量的重要指标, 在水稻缺P和富P条件下, 随着营养液中ρ(P)的提高(1.5~48.0 mg·L-1), 糙米生物量的范围为13.4~17.6 g·株-1, 其中P24处理下糙米生物量最大.
|
|
表 1 各处理对水稻生物量的影响1) Table 1 Effects of different treatments on biomass of rice plants |
光合作用是植物吸收光能转化为生物能的主要机制[33], 因此光合色素的含量对水稻生长有重要影响[34].图 1为各处理水稻鲜叶片中光合色素含量.在Cd胁迫且水稻缺P和富P条件下, 随着ρ(P)的提高(1.5~48.0 mg·L-1), 光合色素(叶绿素a、叶绿素b和类胡萝卜素)的含量呈现先上升后下降的趋势, 均在P12处理下达到最高含量, 分别为1.70、0.40和0.41 mg·g-1.水稻缺P条件下(≤6.0 mg·L-1), 当ρ(P)从1.5 mg·L-1增加至6.0 mg·L-1, 叶片中光合色素含量提高, 叶绿素a、叶绿素b和类胡萝卜素的最大增幅分别为43.2%、31.0%和28.2%; 而当水稻富P条件下(≥12.0 mg·L-1), ρ(P)从12.0 mg·L-1增加到48.0 mg·L-1, 叶片中光合色素含量逐渐下降, 3种光合色素含量的最大降幅分别为65.1%、56.6%和60.4%.上述结果说明, Cd胁迫且水稻缺P条件下, 增加P可促进水稻叶片光合色素的合成, 而富P条件下, 增加P则抑制水稻叶片光合色素的合成.

|
不同字母表示P<0.05水平上差异显著, 下同 图 1 各处理水稻光合色素含量 Fig. 1 Contents of photosynthetic pigment in rice leaves with different treatments |
从图 2可知, 水稻缺P和富P条件下, 各处理水稻根、茎、叶、谷壳和糙米中的Cd含量变化显著.水稻缺P条件下(≤6.0 mg·L-1), 随各处理ρ(P)的提高(1.5~6.0 mg·L-1), 水稻根、茎、谷壳和糙米中Cd含量逐渐增加, 最大增幅分别为191.4%、86.4%、73.3%和132.1%, 而水稻叶中Cd含量在P3处理下达到最大值.在水稻富P条件下(≥12.0 mg·L-1), 随ρ(P)的提高(12.0~48.0 mg·L-1), 水稻根、茎、叶、谷壳和糙米中Cd含量增加, 最大增幅分别为217.0%、194.6%、606.5%、35.9%和191.2%.这说明水稻缺P和富P条件下P均促进了水稻各部位对Cd的吸收, 其中P1.5处理下糙米ω(Cd)最低, 为0.63 mg·kg-1, 而P48处理下糙米中ω(Cd)最高, 达2.41 mg·kg-1.

|
图 2 不同处理下水稻各部位Cd含量 Fig. 2 Content of Cd in different tissues of rice plants with different treatments |
Cd胁迫下缺P和富P处理水稻各部位中Cd的累积量见图 3.各部位Cd累积量占总累积的比例差异结果表明, 根是水稻吸收和积累Cd的主要部位(31.7% ~62.7%), 其次是茎(24.2% ~37.0%)和糙米(8.9% ~18.8%), 叶和谷壳中Cd累积量较低.在缺P(≤6.0 mg·L-1)和富P(≥12.0 mg·L-1)处理下, 水稻Cd累积量呈现出分段增加趋势.在水稻缺P条件下, 随着ρ(P)由1.5 mg·L-1提高至6.0 mg·L-1, 地上部位Cd的累积量提高了80.4% ~94.3%, 其中糙米累积量提高了115.1% ~132.0%; 水稻各部位在P6处理下均达到最大值, 其中根系和糙米Cd累积量分别为151.5 μg·株-1和22.2 μg·株-1.在水稻富P条件下, 当ρ(P)由12.0 mg·L-1提高至48.0 mg·L-1时, 地上部位Cd累积量提高了30.0% ~268.9%, 其中糙米累积量提高了39.9% ~214.4%; 水稻各部位在P48处理下均达到最大值, 其中根系和糙米Cd累积量分别为132.0 μg·株-1和37.6 μg·株-1.

|
图 3 不同处理下水稻各部位Cd累积量 Fig. 3 Cd accumulation in rice tissues under different treatments |
转运系数表示Cd在水稻不同部位间的迁移能力. P浓度水平可影响各部位之间Cd的转运系数.从图 4可知, TF根-茎的转运系数为0.071~0.139, 低于地上部位之间的转运系数.地上部位茎与各部位间转运系数大小顺序为: TF茎-叶>TF茎-糙米>TF茎-谷壳, 表明茎中Cd向叶转运的能力强于向糙米和谷壳转运能力.水稻缺P和富P条件下, P浓度增加对TF根-茎没有明显影响.TF茎-叶、TF茎-糙米和TF茎-谷壳的变化趋势基本一致, 在水稻缺P(≤6.0 mg·L-1)条件时, 随P浓度上升均呈现先增加后减少, P6下最低; 水稻富P(≥12.0 mg·L-1)条件时, 随P浓度上升, 茎向各部位间转运系数变化趋势不一, 其中TF茎-谷壳呈逐渐降低趋势, 而TF茎-叶持续增加.

|
图 4 水稻各部位间Cd的转运系数 Fig. 4 Translocation factors of Cd among various rice tissues |
P/Cd可以反映水稻各部位P和Cd增加量之间的相对关系.表 2中水稻各部位P/Cd的大小顺序为: 糙米>谷壳>叶>茎>根.在缺P(≤6.0 mg·L-1)和富P(≥12.0 mg·L-1)两种浓度范围内, 根和茎中P/Cd具有相同的变化规律, 随P浓度提高P/Cd均先增加后减少; 叶片和谷壳P/Cd的变化规律不明显, 但P48相较P1.5处理显著减少; 而糙米P/Cd分别在缺P(≤6.0 mg·L-1)和富P(≥12.0 mg·L-1)条件下表现为分段下降趋势.这表明, 水稻在缺P和富P条件下, 提高P浓度会促进水稻根、茎、叶、谷壳和糙米中Cd的累积.
|
|
表 2 各处理下水稻各部位P/Cd Table 2 P/Cd in various rice tissues with different treatments |
进一步分析了谷壳和糙米中Cd与P/Cd的相关性.各处理下, 谷壳Cd含量与P/Cd呈显著负相关关系, 糙米Cd含量与P/Cd呈极显著负相关关系(图 5), 其相关系数分别为R谷壳2=0.835 6和R糙米2=0.919 5.这些结果表明, 水稻谷壳和糙米中Cd含量随其P含量增加而显著增加.

|
图 5 水稻谷壳和糙米中Cd含量与P/Cd的关系 Fig. 5 Relationship between Cd concentrations and P/Cd in husk and brown rice |
P是水稻生长的必需元素, 也是光合色素的重要组成元素, 影响水稻叶片光合作用, 促进水稻生长, 并增加水稻的生物量[35, 36].本研究中, 在水稻缺P(≤6.0 mg·L-1)和富P(≥12.0 mg·L-1)条件下, Cd(0.02 mg·L-1)胁迫对水稻各部位生物量变化无明显影响(表 1), 而水稻各部位Cd含量及Cd累积量均呈上升趋势(图 2和图 3).这与已有研究报道的施用P促进水稻生长而降低水稻Cd积累的“稀释效应”[37]相反.其原因在于本研究中Cd胁迫下, 提高ρ(P)(1.5~48.0 mg·L-1)促进了水稻各部位Cd积累, 从而抑制水稻叶片光合作用, 对水稻生长产生毒害作用.水稻叶片中叶绿素a、叶绿素b和类胡萝卜素的含量在ρ(P)为1.5~12.0 mg·L-1范围内上升(图 1), 水稻茎、叶、谷壳和糙米中Cd含量亦呈上升趋势(图 2), 继续提高ρ(P)(12.0~48.0 mg·L-1), 叶片中Cd含量进一步上升, 过量Cd会破坏叶绿体结构, 导致光合色素含量下降(图 1).这说明Cd胁迫下, 水稻缺P状况时适当提高P浓度有利于水稻叶片中光合色素的合成, 而水稻富P状况时, 过量P导致Cd含量提高, 反而抑制了光合色素的合成[38, 39].此外, 本研究的水培溶液中ρ(Cd)为0.02 mg·L-1, 各处理水稻各部位P含量均增加, 且水稻各部位中Cd含量均较高, 尤其是糙米中ω(Cd)为0.63~2.85 mg·kg-1, 且糙米中Cd含量与P/Cd呈极显著负相关关系(图 5), 这表明Cd胁迫下, 糙米中P与Cd存在协同作用, 糙米中P含量的增加显著提高了Cd的累积, 说明水培条件下全生育期Cd胁迫对水稻生长产生明显毒害效应明显高于中重度Cd污染稻田土壤.大量研究表明, 过量Cd会损伤水稻根尖细胞核, 影响RNA的合成, 抑制酶的活性, 减少Fe元素的转运等[40, 41].
根系是水稻吸收Cd的重要部位[42].本研究结果中, 根中Cd含量、累积量和P/Cd均表现出缺P和富P下的分段变化, 并均在P6和P48处理附近较突出(图 2、图 3和表 2).这些结果说明, 水稻缺P和富P时, P对Cd在根中的吸收均表现出较大的协同作用, 高P水平促进了Cd在水稻各部位中的累积, 同时分段变化也表明, 缺P和富P时根系可能存在不同的Cd吸收转运机制.有研究表明, P促进Cd在水稻根系中的吸收可能与根系中相关转运基因的表达有关, 其中转运基因OsNramP 1和OsNramP5参与根系中Cd的转运过程[24]. OsNramP 1是缺P状态下植物根系吸收Cd的主要转运体[43].本研究中, 水稻缺P状态下, 随ρ(P)的提高(1.5~6.0 mg·L-1), 水稻各部位Cd含量均增加, 但根部Cd增加量远高于茎、叶、谷壳和糙米部位Cd增加量, 因此水稻各部位间转运系数先上升后下降, 这可能是缺P信号促进了OsNramp 1基因的表达, 导致根中Cd含量显著增加. OsNramp 5是水稻根细胞中参与吸收Cd的主要转运体, 也负责Cd从根向地上部位的转运[44], 而P的施入会促进OsNramp 5基因的表达[45].水稻富P状态下, 持续提高ρ(P)(12.0~48.0 mg·L-1)提高了各部位间转运系数, 因此高浓度的P可能促进OsNramp 5基因的表达, 从而促进了水稻对Cd的积累, 并增加了Cd从根向地上部位的转运(图 4).同时, P会促进根系生长和植物的蒸腾作用, 进而促进水稻对Cd的吸收和从根到地上部位的转运作用[46].显然, 在Cd胁迫下, 水稻缺P和富P时, P浓度的提高促进了Cd在根系中的累积, 且增强了Cd从根向茎、糙米中的转运.各处理间水稻各部位Cd含量的变化表明, 水稻P与Cd的累积存在协同作用.本研究结果显示, 水稻各部位Cd含量在P6~P12处理间突降, P6处理下水稻对Cd的累积明显高于P12处理, 这说明Cd胁迫下ρ(P)在6.0~12.0 mg·L-1是水稻Cd吸收转运调控的关键区域. P对水稻吸收Cd的影响不是单纯的促进或抑制作用, 最适宜的P浓度添加水平可能与P对水稻对Cd吸收转运的影响相关, 需要进一步研究复杂的P-Cd-水稻相互作用, 以更清楚地了解P水平影响水稻吸收和转运Cd的机制.
4 结论(1) 水培试验Cd胁迫(0.02 mg·L-1)下, ρ(P)的提高(1.5~48.0 mg·L-1)对水稻生物量无明显影响, P对糙米产量增加的贡献较低; 在ρ(P)为12.0 mg·L-1处理下, 水稻叶片光合色素含量最高, 缺P和富P均抑制光合色素的合成.
(2) P添加水平对Cd在水稻中的吸收和累积呈现分段增加趋势, 在水稻缺P(≤6.0 mg·L-1)和富P(≥12.0 mg·L-1)状态下, 提高P浓度均会增加水稻各部位Cd的累积, 糙米中Cd含量在缺P和富P下最高增幅分别为132.1%和191.2%.
(3) 在水稻缺P和富P条件下, 增加P浓度均会促进Cd从根向地上部位转运, 且茎中Cd向叶转运的能力强于向糙米和谷壳转运的能力; 糙米的P/Cd比值在缺P和富P下呈现分段减少规律, 增加P会促进糙米对Cd的累积.
| [1] | Yang J L, Li K, Zheng W, et al. Characterization of early transcriptional responses to cadmium in the root and leaf of Cd-resistant Salix matsudana Koidz[J]. BMC Genomics, 2015, 16(1). DOI:10.1186/s12864-015-1923-4 |
| [2] | Liu X D, Liu F, Huang W H, et al. Quantitative determination of Cd in soil using laser-induced breakdown spectroscopy in air and Ar conditions[J]. Molecules, 2018, 23(10). DOI:10.3390/molecules23102492 |
| [3] | Yin A G, Yang Z Y, Ebbs S, et al. Effects of phosphorus on chemical forms of Cd in plants of four spinach (Spinacia oleracea L.) cultivars differing in Cd accumulation[J]. Environmental Science and Pollution Research, 2016, 23(6): 5753-5762. DOI:10.1007/s11356-015-5813-8 |
| [4] | Chen Z, Tang Y T, Zhou C, et al. Mechanisms of Fe biofortification and mitigation of Cd accumulation in rice (Oryza sativa L.) grown hydroponically with Fe chelate fertilization[J]. Chemosphere, 2017, 175: 275-285. DOI:10.1016/j.chemosphere.2017.02.053 |
| [5] |
周一敏, 黄雅媛, 刘晓月, 等. 叶面喷施纳米MnO2对水稻富集镉的影响机制[J]. 环境科学, 2021, 42(2): 932-940. Zhou Y M, Huang Y Y, Liu X Y, et al. Effect and mechanism of foliar application nano-MnO2 on cadmium enrichment of rice[J]. Environmental Science, 2021, 42(2): 932-940. |
| [6] | Zhang S, Li Q, Nazir M M, et al. Calcium plays a double-edged role in modulating cadmium uptake and translocation in rice[J]. International Journal of Molecular Sciences, 2020, 21(21). DOI:10.3390/ijms21218058 |
| [7] | Wang H, Xu C, Luo Z C, et al. Foliar application of Zn can reduce Cd concentrations in rice (Oryza sativa L.) under field conditions[J]. Environmental Science and Pollution Research, 2018, 25(29): 29287-29294. DOI:10.1007/s11356-018-2938-6 |
| [8] | Bari M A, Prity S A, Das U, et al. Silicon induces phytochelatin and ROS scavengers facilitating cadmium detoxification in rice[J]. Plant Biology, 2020, 22(3): 472-479. DOI:10.1111/plb.13090 |
| [9] | Zhao Z Q, Zhu Y G, Cai Y L. Effects of zinc on cadmium uptake by spring wheat (Triticum aestivum, L.): long-time hydroponic study and short-time 109Cd tracing study[J]. Journal of Zhejiang University-Science A, 2005, 6(7): 643-648. DOI:10.1631/jzus.2005.A0643 |
| [10] | Shao G S, Chen M X, Wang W X, et al. Iron nutrition affects cadmium accumulation and toxicity in rice plants[J]. Plant Growth Regulation, 2007, 53(1): 33-42. DOI:10.1007/s10725-007-9201-3 |
| [11] | Tian S K, Lu L L, Zhang J, et al. Calcium protects roots of Sedum alfredii H. against cadmium-induced oxidative stress[J]. Chemosphere, 2011, 84(1): 63-69. DOI:10.1016/j.chemosphere.2011.02.054 |
| [12] | Wu D Z, Yamaji N, Yamane M, et al. The HvNramp5 transporter mediates uptake of cadmium and manganese, but not iron[J]. Plant Physiology, 2016, 172(3): 1899-1910. DOI:10.1104/pp.16.01189 |
| [13] | Fu Y Q, Yang X J, Shen H. Root iron plaque alleviates cadmium toxicity to rice (Oryza sativa) seedlings[J]. Ecotoxicology and Environmental Safety, 2018, 161: 534-541. DOI:10.1016/j.ecoenv.2018.06.015 |
| [14] | Dong M F, Feng R W, Wang R G, et al. Inoculation of Fe/Mn-oxidizing bacteria enhances Fe/Mn plaque formation and reduces Cd and As accumulation in rice plant tissues[J]. Plant and Soil, 2016, 404(1-2): 75-83. DOI:10.1007/s11104-016-2829-x |
| [15] |
王小蒙, 丁永祯, 郑向群, 等. 根施钙镁磷肥与叶喷硅/硒联合调控水稻镉吸收[J]. 环境工程学报, 2016, 10(11): 6383-6391. Wang X M, Ding Y Z, Zheng X Q, et al. Cadmium uptake in paddy rice under combined regulation of soil application of calcium magnesium phosphate and leaf spraying of Si/Se[J]. Chinese Journal of Environmental Engineering, 2016, 10(11): 6383-6391. DOI:10.12030/j.cjee.201604158 |
| [16] | Huang G X, Ding C F, Zhou Z G, et al. A tillering application of zinc fertilizer based on basal stabilization reduces Cd accumulation in rice (Oryza sativa L.)[J]. Ecotoxicology and Environmental Safety, 2019, 167: 338-344. DOI:10.1016/j.ecoenv.2018.10.044 |
| [17] | Liu Y, Xie Y R, Wang H, et al. Light and ethylene coordinately regulate the phosphate starvation response through transcriptional regulation of PHOSPHATE STARVATION RESPONSE1[J]. The Plant Cell, 2017, 29(9): 2269-2284. DOI:10.1105/tpc.17.00268 |
| [18] | Oropeza-Aburto A, Cruz-Ramírez A, Acevedo-Hernández G J, et al. Functional analysis of the Arabidopsis PLDZ2 promoter reveals an evolutionarily conserved low-Pi-responsive transcriptional enhancer element[J]. Journal of Experimental Botany, 2012, 63(5): 2189-2202. DOI:10.1093/jxb/err446 |
| [19] | Akram M S, Shahid M, Tariq M, et al. Deciphering Staphylococcus sciuri SAT-17 mediated anti-oxidative defense mechanisms and growth modulations in salt stressed maize (Zea mays L.)[J]. Frontiers in Microbiology, 2016, 7. DOI:10.3389/fmicb.2016.00867 |
| [20] | Jung H I, Chae M J, Kong M S, et al. Effect of phosphorus on the cadmium transfer and ROS-scavenging capacity of rice seedlings[J]. Korean Journal of Soil Science and Fertilizer, 2017, 50(4): 203-214. |
| [21] | Gao X P, Flaten D N, Tenuta M, et al. Soil solution dynamics and plant uptake of cadmium and zinc by durum wheat following phosphate fertilization[J]. Plant and Soil, 2011, 338(1-2): 423-434. DOI:10.1007/s11104-010-0556-2 |
| [22] | Adhikari A, Lee K E, Khan M A, et al. Effect of silicate and phosphate solubilizing rhizobacterium Enterobacter ludwigii GAK2 on Oryza sativa L. under cadmium stress[J]. Journal of Microbiology and Biotechnology, 2020, 30(1): 118-126. DOI:10.4014/jmb.1906.06010 |
| [23] | Lee H H, Kim K K, Lee Y B, et al. Effect of phosphate application on cadmium extractability and its uptake by rice cultivated in contaminated paddy soil[J]. Korean Journal of Environmental Agriculture, 2016, 35(4): 235-240. DOI:10.5338/KJEA.2016.35.4.35 |
| [24] | Wang X Q, Ruan W Y, Yi K K. Internal phosphate starvation signaling and external phosphate availability have no obvious effect on the accumulation of cadmium in rice[J]. Journal of Integrative Agriculture, 2019, 18(9): 2153-2161. DOI:10.1016/S2095-3119(18)62100-8 |
| [25] | Seshadri B, Bolan N S, Wijesekara H, et al. Phosphorus-cadmium interactions in paddy soils[J]. Geoderma, 2016, 270: 43-59. DOI:10.1016/j.geoderma.2015.11.029 |
| [26] | Liu H J, Zhang J L, Christie P, et al. Influence of external zinc and phosphorus supply on Cd uptake by rice (Oryza sativa L.) seedlings with root surface iron plaque[J]. Plant and Soil, 2007, 300(1-2): 105-115. DOI:10.1007/s11104-007-9393-3 |
| [27] | Li Y Y, Yu S H, Zhou X B. Effects of phosphorus on absorption and transport of selenium in rice seedlings[J]. Environmental Science and Pollution Research, 2019, 26(14): 13755-13761. DOI:10.1007/s11356-018-2690-y |
| [28] |
黄文方, 陈晓阳, 邢承华, 等. 磷对水稻耐铝性及根尖细胞壁组分的影响[J]. 中国水稻科学, 2013, 27(2): 161-167. Huang W F, Chen X Y, Xing C H, et al. Effects of phosphorous on aluminum tolerance and cell wall polysaccharide components in rice root tips[J]. Chinese Journal of Rice Science, 2013, 27(2): 161-167. |
| [29] |
朱春权, 徐青山, 曹小闯, 等. 不同属性特征基质对早稻秧苗耐低温的影响[J]. 中国水稻科学, 2021, 35(5): 503-512. Zhu C Q, Xu Q S, Cao X C, et al. Effects of substrates with different properties on chilling tolerance of early rice seedlings[J]. Chinese Journal of Rice Science, 2021, 35(5): 503-512. |
| [30] | Joy J, Silvestri A P, Franke R, et al. Calcium and phosphate compatibility in low-osmolarity parenteral nutrition admixtures intended for peripheral vein administration[J]. Journal of Parenteral and Enteral Nutrition, 2010, 34(1): 46-54. DOI:10.1177/0148607109338216 |
| [31] |
王倩倩, 贾润语, 李虹呈, 等. Cd胁迫水培试验下水稻糙米Cd累积的关键生育时期[J]. 中国农业科学, 2018, 51(23): 4424-4433. Wang Q Q, Jia R Y, Li H C, et al. Key growth stage of Cd accumulation in brown rice through a hydroponic experiment with Cd stress[J]. Scientia Agricultura Sinica, 2018, 51(23): 4424-4433. DOI:10.3864/j.issn.0578-1752.2018.23.003 |
| [32] | Sayantan D, Shardendu. Phosphorus amendment competitively prevents chromium uptake and mitigates its toxicity in Spinacea oleracea L.[J]. Indian Journal of Experimental Biology, 2015, 53(6): 395-405. |
| [33] |
李娜, 张峰举, 许兴, 等. 增温对宁夏北部春小麦叶片光合作用的影响[J]. 生态学报, 2019, 39(24): 9101-9110. Li N, Zhang F J, Xu X, et al. Effects of elevated temperature on photosynthesis of spring wheat in northern Ningxia[J]. Acta Ecologica Sinica, 2019, 39(24): 9101-9110. |
| [34] | Ali Q, Ali S, El-Esawi M A, et al. Foliar spray of Fe-Asp confers better drought tolerance in sunflower as compared with FeSO4: Yield traits, osmotic adjustment, and antioxidative defense mechanisms[J]. Biomolecules, 2020, 10(9). DOI:10.3390/biom10091217 |
| [35] | Siebers N, Siangliw M, Tongcumpou C. Cadmium uptake and subcellular distribution in rice plants as affected by phosphorus: soil and hydroponic experiments[J]. Journal of Soil Science and Plant Nutrition, 2013, 13(4): 833-844. |
| [36] | Zhu Z G, Li G R, Yan C H, et al. DRL1, encoding a NAC transcription factor, is involved in leaf senescence in grapevine[J]. International Journal of Molecular Sciences, 2019, 20(11). DOI:10.3390/ijms20112678 |
| [37] |
黄洋, 郭晓, 胡学玉. 生物质炭对磷镉富集土壤中两种元素生物有效性及作物镉积累的影响[J]. 环境科学, 2020, 41(6): 2861-2868. Huang Y, Guo X, Hu X Y. Effects of biochar on bioavailability of two elements in phosphorus and cadmium-enriched soil and accumulation of cadmium in crops[J]. Environmental Science, 2020, 41(6): 2861-2868. |
| [38] | Fu Y, Mason A S, Zhang Y F, et al. MicroRNA-mRNA expression profiles and their potential role in cadmium stress response in Brassica napus[J]. BMC Plant Biology, 2019, 19(1). DOI:10.1186/s12870-019-2189-9 |
| [39] | Hattab S, Dridi B, Chouba L, et al. Photosynthesis and growth responses of pea Pisum sativum L. under heavy metals stress[J]. Journal of Environmental Sciences, 2009, 21(11): 1552-1556. DOI:10.1016/S1001-0742(08)62454-7 |
| [40] | Huang Y M, Chen H Q, Reinfelder J R, et al. A transcriptomic (RNA-seq) analysis of genes responsive to both cadmium and arsenic stress in rice root[J]. Science of the Total Environment, 2019, 666: 445-460. DOI:10.1016/j.scitotenv.2019.02.281 |
| [41] | Di Toppi L S, Gabbrielli R. Response to cadmium in higher plants[J]. Environmental and Experimental Botany, 1999, 41(2): 105-130. DOI:10.1016/S0098-8472(98)00058-6 |
| [42] |
方波, 肖腾伟, 苏娜娜, 等. 水稻镉吸收及其在各器官间转运积累的研究进展[J]. 中国水稻科学, 2021, 35(3): 225-237. Fang B, Xiao T W, Su N N, et al. Research progress on cadmium uptake and its transport and accumulation among organs in rice[J]. Chinese Journal of Rice Science, 2021, 35(3): 225-237. |
| [43] | Wang H O, Wang T, Ahmad I. Involvement of phosphate supplies in different transcriptional regulation pathway of Oryza sativa L.'s antioxidative system in response to arsenite and cadmium stress[J]. Ecotoxicology, 2015, 24(6): 1259-1268. DOI:10.1007/s10646-015-1496-7 |
| [44] | Wang T K, Li Y X, Fu Y F, et al. Mutation at different sites of metal transporter gene OsNramp5 affects Cd accumulation and related agronomic traits in rice (Oryza sativa L.)[J]. Frontiers in Plant Science, 2019, 10. DOI:10.3389/fpls.2019.01081 |
| [45] |
曹庭悦, 刘鸣达, 沃惜慧, 等. 硅、磷配施对水稻镉吸收转运的影响及其机制[J]. 农业环境科学学报, 2020, 39(1): 37-44. Cao T Y, Liu M D, Wo X H, et al. Effects of combined application of silicon and phosphorus on cadmium uptake and transport in rice and its mechanisms[J]. Journal of Agro-Environment Science, 2020, 39(1): 37-44. |
| [46] | Gutsch A, Hendrix S, Guerriero G, et al. Long-term Cd exposure alters the metabolite profile in stem tissue of Medicago sativa[J]. Cells, 2020, 9(12). DOI:10.3390/cells9122707 |