 2019, Vol. 40
2019, Vol. 40



2. 中国科学院南京地理与湖泊研究所湖泊与环境国家重点实验室, 南京 210008
2. State Key Laboratory of Lake Science and Environment, Nanjing Institute of Geography and Limnology, Chinese Academy of Sciences, Nanjing 210008, China
我国目前66%以上的湖泊、水库处于富营养化的水平, 其中重富营养化和超富营养化的占22%, 使得富营养化成为我国湖泊目前与今后相当长一段时期内的重大水环境问题[1].太湖是我国第三大淡水湖泊, 具有复杂的河网水系, 草、藻型等不同的生态类型, 是一个大型的富营养化浅水湖泊, 面临着严重的湖泊富营养化和频繁的蓝藻水华暴发[2, 3].湖泊污染和富营养化的根源在于水体营养盐(主要是氮和磷)的积累.氮和磷是湖泊初级生产力的主要驱动因素, 是引起湖泊富营养化和蓝藻水华暴发的关键因素.相关研究表明, 浅水湖泊中的氮磷浓度具有明显的季节变化趋势, 而且这种变化趋势与湖泊的营养水平和生物活动密切相关[4].研究水体氮和磷的变化对藻类生长繁殖的影响, 对于治理富营养化水体中藻类的暴发性增长具有重要意义.
Xu等[4]通过对蓝藻水华发生过程监测和原位营养盐添加实验发现, 太湖水体溶解态氮磷浓度及浮游植物生长的限制因子随季节变化而变化, 并指出浮游植物生长的无机氮、磷阈值分别为0.30~0.80 mg ·L-1和0.014~0.214 mg ·L-1.然而, 作为溶解态营养盐再生的核心载体之一, 颗粒态营养盐是蓝藻水华再生和生长的重要营养源.有研究表明, 藻源颗粒态营养盐降解过程中释放大量溶解态营养盐, 维持蓝藻水华的暴发性生长[5].何东等[6]在藻源性颗粒态营养盐降解过程中发现, 颗粒态氮降解为氨氮后, 通过硝化作用, 先被转化为亚硝酸盐氮再被转化为硝酸盐氮, 颗粒态磷主要降解为磷酸盐.对太湖细菌的研究中也发现, 蓝藻水华的聚集、分解, 形成了以颗粒物为载体和代谢中心的营养盐循环模式[7].
关于太湖氮磷营养盐污染研究方面已取得了不少成果, 例如: Xu等[4]研究了太湖溶解性氮磷营养盐含量及其对浮游植物生长的影响; 聂泽宇等[5]基于氮磷比解析了太湖苕溪水体营养现状.但用地统计学与地理信息系统(geographic information system, GIS)相结合研究太湖氮磷营养盐时空分布及其变异特性的工作鲜见报道.
因此, 本研究基于地统计学分析方法, 以太湖2014年8月~2015年5月夏、秋、冬、春四季为研究时段, 分析了草、藻型等不同生态类型湖区颗粒态和溶解态氮、磷营养盐的来源以及赋存形态和营养盐限制类型的时空分布特征, 探寻其时空变化原因, 以期为湖泊富营养化和蓝藻水华的治理提供有益的参考.
1 材料与方法 1.1 研究区概况太湖位于长江三角洲, 湖面面积2 338.1 km, 最大水深不足3 m, 平均深度1.9 m, 是我国第三大淡水湖.太湖是长江三角洲重要的水源地, 太湖常年为周边地区人口供给饮用水及生活用水, 可满足流域内一千万以上人口的用水需求[8].过去30年间经济的高速发展被认为是太湖水质高度空间异质性最重要的原因[9].太湖是一个具有不同生态类型的浅水湖泊, 梅梁湾和东太湖分别为典型的藻型和草型湖区, 面临着富营养化水华暴发和沼泽化两类完全不同的环境问题[10].太湖流域共有172条河流或渠道与该湖相通, 水网交错, 水系复杂, 太湖多年平均水力滞留时间约为300 d[8].
1.2 样品采集与分析2014年8月(夏季)、2014年11月(秋季)、2015年2月(冬季)和2015年5月(春季)的总氮(total nitrogen, TN)、总磷(total phosphorus, TP)、溶解态总氮(total dissolved nitrogen, TDN)、溶解态总磷(total dissolved phosphorus, TDP)、磷酸盐(phosphate, PO43--P)、亚硝酸盐氮(nitrite nitrogen, NO2--N)、硝酸盐氮(nitrate nitrogen, NO3--N)、氨氮(ammonia nitrogen, NH4+-N)、悬浮物(total suspended matter, TSM)和叶绿素a(chlorophyll-a, Chla)数据均来源于中国科学院太湖湖泊生态系统研究站常规采样数据.该站在太湖的8个典型区域(竺山湾、梅梁湾、贡湖湾、胥口湾、东太湖、湖心区、西湖区和南太湖)共布设32个监测点位(图 1), 每年2月、5月、8月和11月各监测一次.南、北部湖区参照Zhou等[11]的方法进行分区, 北部湖区夏季易暴发蓝藻水华即藻型湖区, 南部湖区为沉水植物覆盖即草型湖区.

|
图 1 太湖水体采样点分布示意 Fig. 1 Distribution of the sampling sites in Lake Taihu |
参照文献[12]中的方法, 分析水体TN、TP、TDN、TDP、PO43--P、NO2--N、NO3--N、NH4+-N、TSM和Chla质量浓度. TDN、TDP、PO43--P、NO2--N、NO3--N和NH4+-N样品用GF/F玻璃纤维滤膜过滤后测定. TN、TP、TDN和TDP用过硫酸钾高压消解后用分光光度法测定, PO43--P用钼锑抗比色法测定, NO2--N采用N-(1-萘基)-乙二胺分光光度法测定, NO3--N用酚二磺酸法测定, NH4+-N用钠氏试纸比色法测定.颗粒氮(particulate nitrogen, PN)由TN和TDN之差获得, 颗粒态磷(particulate phosphorus, PP)由TP和TDP之差获得, 无机氮(inorganic nitrogen, DIN)为NO2--N、NO3--N和NH4+-N质量浓度之和, 有机氮(organic nitrogen, DON)为TN和DIN之差, 有机磷(organic phosphorus, DOP)为TP与PO43--P之差.用重量法测定TSM. Chla采用荧光分光光度法测定.
1.3 半变异函数和普通克里金法地统计学是以区域化变量理论为基础, 以半变异函数(semi-variogram functions)为主要工具, 研究在空间分布上既有随机性又有结构性, 或空间相关和依赖性的自然现象的科学[13].区域化变量能够同时反映变量的空间分布结构性与随机性, 所以无论是用完全确定性的数学方法还是用经典统计学方法来研究、描述这类变量都是困难的或者根本不可能的[14].而地统计学中的一个基本工具——半变异函数, 则能够较好地描述区域化变量的上述特征, 从而使针对区域化变量的空间变异性分析得到实现[15].太湖由于受气温、降雨、光照、水动力条件、换水过程、湖泊底泥、围网养殖和径流注入等随机或结构因素综合影响, 形成了草、藻型等不同的生态类型; 即使在同一生态类型湖区, 不同时空位置上, 营养盐的含量也不相同; 这表明太湖是一个时空连续的变异体, 是不均一的和变化着的.利用地统计学结合GIS空间分析技术分析太湖氮磷营养盐的变异性及其时空动态变化, 将帮助人们深入认识和了解营养盐的动态变化对蓝藻水华暴发的影响机制.
利用半变异函数γ(h)分析太湖各水质参数在空间上的变异特征并区分结构因素与随机因素对各营养盐空间变异特征的影响[16], 其公式如下:
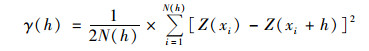
|
(1) |
式中, h为分隔两水样点的矢量, 又叫步长; N(h)为相隔h的点对数; Z(xi)和Z(xi+h)分别为营养盐在位置xi和xi+h处的实测值.
通过半变异函数可得到3个重要参数:块金值、基台值和变程值, 分别用C0、C0+C和a表示, 可一定程度上揭示某参数的空间变异和相关性. C0表示随机性变异的大小, C表示结构性变异的大小, a表示变量存在空间相关性的范围.块金值和基台值的比值C0/(C0+C)又称块金系数, 可以表示随机性变异占总变异的大小, 反映某参数的空间相关程度, 当值<25%时, 表示有强烈空间相关性, 变异性主要受结构因素(湖泊水文状况等)影响; 当值在25%~75%之间时, 表明有中等程度空间相关性, 变异性受结构和随机因素(围网养殖、径流注入、人工挖沙等人类活动)的共同作用; 当值>75%时, 表明空间相关性较弱, 变异性主要受随机因素影响[17].
普通克里金法(ordinary kriging)是建立在大量样本的基础上, 通过分析样本间的相关性, 探索其分布规律, 并进行预测[18].在赋权时, 不仅考虑距离因素, 还通过变异函数和结构分析, 考虑已知样本点的空间分布以及与未知点的空间方位关系[19].
1.4 全局空间自相关Moran´s I指数是度量空间相关性的指标, 反映营养盐在全湖范围是否存在显著空间分布模式, 计算公式如下[20]:
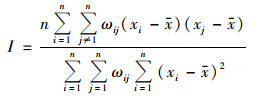
|
(2) |
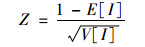
|
(3) |
式中, I为Moran指数; ωij为空间权重矩阵; xi和xj分别为第i和j个水域的实测值. I取值在[-1, 1]之间, 小于0表示负相关, 空间上呈现离散分布状态; 等于0表示不相关, 空间上呈现随机分布状态; 大于0表示正相关, 空间上呈现集聚分布状态; 对Moran´s I值采用Z检验来进行统计检验, Z为得分值, 判断全局Moran´s I的显著性; E[I]为I的期望值; V[I]为I的理论方差; P值表示所观测到的空间模式是由随机过程创建而成的概率.
当Z得分<-1.65或者>1.65, P值<0.10时, 表示结果通过了置信度为90%的显著性检验; 当Z得分<-1.96或者>1.96, P值<0.05时, 表示结果通过了置信度为95%的显著性检验; 当Z得分<-2.58或者>2.58, P值<0.01时, 表示结果通过了置信度为99%的显著性检验.
1.5 绘图及数据处理采用ArcGIS 10.2软件绘制参数空间分布图与采样点分布, 均值、标准差、正态分布以及Pearson相关系数等采用SPSS 21.0软件分析.相关分析用来检测变量之间的关系; P>0.05表示未达到显著检验水平, 0.01<P<0.05为显著水平, P<0.01为极显著水平.正态分布采用K-S检验(one sample kolmogorov-smirnov test)进行分析.
2 结果与分析 2.1 氮磷营养盐质量浓度的时空分布变化 2.1.1 季节变化太湖各形态氮磷营养盐浓度范围及均值如表 1所示, 其中, 太湖水体TN、DIN、DON、TDN和DOP总体呈现冬春高, 夏秋低的规律, TP和PP含量则呈现出夏秋高于冬春的特征(t检验, P<0.01), 与以往报道中的研究结果较为一致[21, 22]. NO3--N、PO43--P和TDP含量冬季最高, 夏季次之, 春秋最低. NO2--N、PN和Chla含量呈现为夏春高于秋冬的规律(t检验, P<0.01);而NH4+-N含量则呈现为秋冬高, 夏春低的特征, 这与朱广伟等[23]的研究结果一致.
|
|
表 1 太湖营养盐平均浓度和浓度范围/mg ·L-1 Table 1 Concentration range and mean of nutrients in Lake Taihu/mg ·L-1 |
2.1.2 空间异质性
太湖水体营养盐半变异函数模型及地统计学参数见表 2.根据空间变异分类系统(变异系数<0.1为弱变异; 0.1<变异系数<1为中等变异; 变异系数>1为强变异)[24], 各季节TN均为中等变异; 除春季TN的空间变异主要受随机因素影响外, 其余季节TN均主要受结构因素的影响.对无机氮来说, 各季节的NO2--N、秋冬季节的NO3--N以及春冬季节的NH4+-N均为强变异, 其余季节无机氮为中等变异; 夏季NO2--N、秋季NO3--N以及春季DIN的中等空间变异同时受结构因素和随机因素的影响, 其余无机氮主要受结构因素的影响.除秋季DON为强变异外, 其余季节DON均为中等变异; 夏季DON的中等空间变异主要受结构因素影响, 其余季节DON的空间变异受结构和随机因素的双重影响.对颗粒态氮来说, 春季PN为强变异, 其余季节PN均为中等变异; 春季PN强空间变异主要受随机因素的影响, 秋季PN中等程度变异同时受随机和结构因素的影响, 夏冬季节PN的空间变异主要受结构因素的影响.
|
|
表 2 太湖营养盐半变异函数模型及地统计学参数 Table 2 Oretical models and fitting parameters for anisotropic semivariograms of nutrients in Lake Taihu |
与TN的空间变异类似, 各季节TP亦为中等变异; 其中, 春秋季节TP同时受随机和结构因素的影响, 夏冬季节TP主要受结构因素影响.除秋季PO43--P为中等变异外, 其余季节无机磷均为强变异; 春季PO43--P的强变异受随机和结构因素的双重影响, 其余季节PO43--P的空间变异主要受结构因素影响.对于有机磷来说, 各季节DOP均为中等变异; 其中, 夏季DOP的空间变异受结构和随机因素的影响, 其余季节DOP空间变异主要受结构因素影响.春季PP为强变异, 其余季节PP为中等变异; 秋季PP的空间变异受结构和随机因素影响, 其余季节PP的空间变异主要受结构因素影响.
对于Chla来说, 春季Chla为强变异, 说明其含量变化幅度较大, 空间差异较大, 其余季节Chla均为中等程度变异; 冬季Chla的空间变异受结构和随机因素双重影响, 其余季节Chla的空间变异主要受结构因素影响.
2.1.3 空间插值经K-S检验发现, 秋季PN, 冬季PN、PP和Chla以及春季TN、DON和TDN符合正态分布, 其余季节营养盐均符合对数正态分布, 因此可基于最优半变异函数模型及其参数对各季节营养盐进行Kriging插值(表 2).太湖营养盐空间插值分布见图 2.从中可知, 夏季Chla、PN以及秋季PN含量的高值区出现在梅梁湾、竺山湾和大浦河等西北湖区, 呈现由西北湖区向东南湖区递减的趋势.春季PN、PP、Chla和秋季Chla含量的高值区集中在竺山湾、梅梁湾和贡湖湾等北部湖区, 由北部湖区向南部湖区递减.冬季PN、PP和Chla含量的高值区主要分布在南部湖区, 整体表现为由南部湖区向北部湖区递减的分布模式.其他季节营养盐的高值区均集中在竺山湾和大浦河附近, 呈现由竺山湾向东太湖递减的趋势.

|
图 2 太湖营养盐含量空间分布 Fig. 2 Spatial interpolation maps of nutrients in Lake Taihu |
全局空间自相关统计量(如Moran´s I)可用来测度同一属性在不同空间位置上的集中性及显著性程度[25].由表 2可知, 太湖水体春季TN、PN、PP和Chla含量的Moran´s I指数大于零, 但P值大于0.10, 没有通过90%的显著性检验, 表示结果由随机过程创建的概率较大, 即表现为随机分布.其余季节各营养盐Moran´s I指数均大于零, P值均小于0.10, Z得分均大于1.65, 均通过显著性检验, 说明其存在高度的正的全局空间自相关关系, 即表现为空间集聚性, 其空间特征为:营养盐浓度高的采样点相对趋向于与营养盐浓度高的采样点相邻, 营养盐浓度低的采样点相对趋向于与营养盐浓度低的采样点相邻.
2.2 氮磷营养盐形态及结构组成对太湖水体氮磷营养盐的形态及结构进行分析(表 3和图 3). TDN/TN比值冬春季节高于夏秋季节(t检验, P<0.01), 金颖薇等[26]也获得了类似的结论.由太湖营养盐结构组成空间分布图可知, 西北湖区由冬季以溶解态无机氮NO3--N为主, 转变为春、夏、秋季以颗粒态氮为主; 东南湖区由冬季以颗粒态氮为主, 转变为春夏季以溶解态有机氮为主, 再转变为秋季以溶解态无机氮NH4+-N为主.
|
|
表 3 太湖水体氮磷营养盐组成/% Table 3 Speciation of N and P in Lake Taihu/% |

|
图 3 太湖营养盐结构组成空间分布 Fig. 3 Spatial interpolation maps of nutrient speciation in Lake Taihu |
与氮营养盐类似, TDP/TP比值亦为冬春季节高于夏秋季节(t检验, P<0.01).从空间分布来看, 西北湖区由冬季以溶解态有机磷为主, 转变为春、夏、秋季以颗粒态磷为主; 与西北湖区恰恰相反, 东南湖区由冬季以颗粒态磷为主, 转变为春、夏、秋季以溶解态有机磷为主.
氮磷比(N/P)被广泛地作为考察营养盐结构的主要指标[27].由表 3可知, TN/TP与TDN/TDP存在着类似的季节变化规律, 均表现为春冬季节高于夏秋季节(t检验, P<0.01).西北湖区TN/TP比值由秋冬季节大于16, 降低为夏春季节的小于16;而东南湖区却由秋冬季节小于16, 升高为夏春季节大于16. TDN/TDP比值在西北湖区的空间变化规律与TN/TP比值一致, 而在东南湖区TDN/TDP比值由秋季小于16, 升高为夏、冬、春季节大于16.颗粒态氮磷比时空变化均不显著(t检验, P>0.05), 各季节西北湖区PN/PP比值均小于16, 东南湖区均大于16.
3 讨论 3.1 基于地统计分析的太湖营养盐来源解析湖泊水体中的营养盐, 依据其输入途径一般可分为外源性和内源性两类; 其中, 外源性的营养盐主要通过大气沉降、地表径流和人为排放等途径进入湖泊水体; 内源性的营养盐主要指由沉水植物、藻类、细菌及微生物等降解和分泌产生和蓄积在湖泊沉积物内可向水体中释放的营养盐[28, 29].
地统计中的块金系数通常用来研究和表征物质的来源[30, 31].地统计中, 影响营养盐空间分布和来源的因素一般分为随机因素(人为因素)和结构因素(湖泊水文状况等自然因素).本研究中, 太湖水体春季TN、PN、PP和Chla含量的块金系数均大于75%, 表明其空间相关性较弱, 空间异质主要受随机因素的影响; 全局空间自相关分析结果也证实, 太湖水体春季TN、PN、PP和Chla含量表现为随机分布, 即空间分布受人为陆源输入或其他随机因素的影响.春季TN、PN、PP和Chla含量由竺山湾和大浦河西北湖区向东太湖递减也证实受陆源输入的影响较大.春季竺山湾和大浦河等西北湖区氮、磷营养盐均主要以颗粒态形式存在; 且相关分析结果(图 4)显示, 春季TN、PN、PP与Chla均具有极好的正相关关系, 表明春季藻类颗粒态氮、磷是太湖水体氮磷的主要赋存形式; 因此, 春季藻类复苏应该与竺山湾、大浦河等西北部化工、印染产业密集、河水污染严重等人类因素有关; 叶宏萌等[32]研究太湖北部流域水质状况时发现, 太湖北部流域受人类影响较大, 氮磷污染严重.另外, 春季TN、PN、PP和Chla与悬浮物(TSM)均具有较好的相关性, 这说明春季TN、PN、PP、Chla与TSM具有同源性; 张润宇等[33]指出, 太湖春季东南风或偏南风引起的强烈的沉积物再悬浮, 使水体中悬浮物显著升高; 因此, 春季藻类复苏除与人类因素有关外, 还可能与风浪扰动引起的沉积物再悬浮等随机因素有关.

|
图 4 太湖营养盐相关性分析 Fig. 4 Analysis of correlation of nutrients in Lake Taihu |
夏季NO2--N和DOP, 秋季NO3--N、DON、PN、TP和PP, 冬季DON和Chla, 春季DIN、DON、TP和PO43--P含量的块金系数在25%~75%之间, 属于中等程度的空间变异, 说明同时受随机因素和结构因素的双重影响.夏季NO2--N和DOP, 秋季NO3--N、DON、PN、TP和PP, 冬季DON和Chla, 春季DIN、DON、TP和PO43--P含量的高值区均位于竺山湾和大浦河区域, 表现为由竺山湾和大浦河向东太湖递减的趋势, 暗示着人为陆源因素的影响.夏季DOP、秋季PN、TP、PP、春季TP与Chla均存在相关关系, 表明与浮游植物新陈代谢有关.秋季PN、TP和PP、冬季Chla、春季DON、TP与TSM具有相关性, 说明可能与风浪扰动引起的沉积物再悬浮等随机因素有关.夏季NO2--N与NO3--N和NH4+-N之间均存在极显著正相关关系, 说明NO2--N来源于NO3--N的反硝化作用和NH4+-N的硝化作用; 秋季NO3--N与NO2--N之间具有极好的相关性, 说明秋季NO3--N来源于NO2--N的硝化作用; 李成高等[34]亦获得类似的结论.
其余各季节营养盐含量的块金系数均小于25%, 说明系统空间相关性很强, 即主要受结构性因素的影响.全局空间自相关分析结果也证实其余营养盐表现为空间集聚性.例如, 冬季PN和PP含量的高值区集中在西南湖区, 呈现出由西南湖区向北部湖区递减的特点; 且冬季PN、PP与Chla均存在相关关系, 这说明冬季PN、PP来自于蓝藻水华的死亡分解.孔繁翔等[35]也指出, 进入冬季后, 大量蓝藻水华在南太湖以及西太湖死亡下沉.
3.2 颗粒态氮磷的转化氮磷形态的空间分布在夏、秋和春季之间差异不大, 均表现为西北湖区以颗粒态为主, 东南湖区以溶解态为主; 而冬季氮磷形态的空间分布明显异于其他季节, 西北湖区以溶解态形态存在, 东南湖区以颗粒态形态存在, 这与蔡琨等[36]的研究结果一致.
冬季西北湖区氮磷营养盐由颗粒态为主转变为由溶解态为主, 这可能与死亡藻类下沉、迁移以及降解有关.本研究中, 冬季PN、PP的块金系数均小于25%, 主要受结构因素的影响, 且均与Chla存在正相关关系, 这说明冬季PN、PP主要来自于蓝藻水华的死亡残体.艾鹰[37]通过研究微囊藻在太湖越冬及复苏的时空格局指出, 在越冬初期, 北部湖区水柱中的部分死亡微囊藻下沉至底泥表面越冬, 部分死亡微囊藻在风力作用下迁移至南部湖区.孔繁翔等[35]也指出, 进入冬季后, 大量蓝藻水华在南太湖以及西太湖死亡下沉.何东等[6]通过分析太湖藻源性颗粒物降解过程认为, 颗粒态氮被降解为NH4+-N后, 通过硝化作用, 先被转化成NO2--N再被转化成NO3--N, 颗粒态磷主要降解为PO43--P.
春季东南湖区氮磷营养盐以颗粒态形态赋存转变为以溶解态形式赋存, 这应该与春季蓝藻复苏有关.艾鹰等[37]的报道指出, 春季复苏期间, 微囊藻生物量首先从南部湖区的表层水体中开始增长, 而后随着风向的转变, 南部的微囊藻向竺山湾等北部湖区迁移, 在条件合适时在北部形成早期的水华.
颗粒态氮磷营养盐的时空变化过程与孔繁翔等[32]提出的蓝藻复苏四阶段理论有着很好的契合关系, 即藻源性颗粒物的迁移、转化和再生, 是藻类生长和水华发生重要的影响因素.
3.3 氮磷比时空变化分析氮磷比可影响藻类等浮游植物的生长, 是湖泊富营养化的一个重要的影响因子[38].有研究发现不同的营养盐比例可以控制藻类的生长、生物量以及种群结构[39]. Redfield定律认为[40], 藻类细胞组成的原子比率C :N :P=106 :16 :1, 若氮磷比超过16 :1, 磷被认为是限制性因素; 反之, 当氮磷比小于16 :1时, 氮通常被认为是限制性因素.相关研究指出, 当水体的N/P比值在10~15之间时, 藻类生长与氮磷浓度存在直线性相关关系[41].
本研究中, TN/TP比值与TDN/TDP比值的时空分布规律一致, 且时空变化差异显著.在蓝藻生长和暴发的春夏季节, 北部湖区尤其竺山湾和梅梁湾区域, TN/TP比值和TDN/TDP比值均小于16, 以溶解态氮限制为主, 溶解态磷相对充足; 在蓝藻开始死亡和下沉的秋冬季节, 北部湖区TN/TP比值和TDN/TDP比值均大于16, 表现为溶解态磷限制; 而南部湖区恰恰相反, 春夏季节TN/TP比值和TDN/TDP比值均大于16, 秋冬季节TN/TP比值和TDN/TDP比值均小于16, 这与Xu等[4]在太湖的氮磷分析结果一致.与TN/TP比值和TDN/TDP比值的时空分布相比, PN/PP比值的时空变化差异并不显著, 四季PN/PP比值均表现为西北部湖区小于16, 东南部湖区大于16.相关分析结果(图 4)显示, 各季节TN/TP比值与TDN/TDP比值均具有极显著正相关关系, 说明TDN/TDP比值的时空变化可在一定程度上反映TN/TP比值的变化.
太湖北部湖区夏春季节TDN浓度降低, 而TDP浓度升高(图 2), 共同导致TDN/TDP比值降低.夏春季节Chla含量集中分布在北部湖区(图 2), 藻类生长吸收利用可能是夏春季节TDN含量大幅下降的因素之一; 周笑白等[42]指出, 太湖丰水期TDN浓度降低的主要原因可能是丰水期蓝藻迅速生长, 吸收了大量的营养盐; Xu等[4]认为藻类的吸收利用对溶解态氮浓度具有明显的影响.夏春季节高降雨的稀释作用, 有可能是造成高水位、低氮的原因之一; 赵林林等[43]报道, 总氮浓度与水位反相关说明了高水位稀释了总氮浓度.春夏季节低氮现象应该还与反硝化脱氮作用有关; Xu等指出[4], 夏春季节温度相对较高, 细菌活性旺盛, 反硝化脱氮作用强烈, 易形成低氮特征; Zhang等[44]认为夏春季节富营养化促进反硝化作用, 从而降低氮浓度.
夏春季节TDP含量的块金系数均小于25%, 说明系统空间相关性很强, 即主要受结构性因素的影响, 因此太湖北部湖区夏春季节TDP浓度升高可能与浮游植物新陈代谢释放自生磷和沉积物静态释放营养盐有关. Chao等[45]的研究表明, 浮游植物代谢产物及其残体降解过程会向水体释放生物可利用的营养盐; Hudson等[46]测定了北美20个淡水湖泊中磷的再生速率, 发现湖泊藻类所需的磷优先取自藻类群体自身的再生磷, 其次才是外源磷.文威等[47]指出, 在沉积物-水界面没有受到破坏的情况下, 溶解性的磷通过浓度梯度从沉积物间隙水中扩散进入上覆水中; 静态释放的强度主要受沉积物-水界面的温度、氧化还原环境及营养盐浓度梯度的影响.
太湖南部湖区夏春季节TDN浓度升高, 而TDP浓度变化不显著, 因此TDN浓度升高是导致TDN/TDP比值升高的主要原因.夏春季节TDN含量的块金系数均小于25%, 主要受结构性因素的影响, TDN浓度升高可能与南部湖区蓝藻水华少, 藻类生长吸收利用少以及沉积物静态释放营养盐有关.
与溶解态氮磷比不同, 颗粒态氮磷比时空变化不显著; 太湖西北湖区四季PN/PP比值均小于16, 东南湖区均大于16.春季蓝藻复苏期西北湖区PN/PP比值小于16, 应该与春季西北湖区PP含量增高有关.春季PP含量的增高主要受人为或风力扰动等随机因素的影响.张红爱等[48]指出, 春季是磷肥大量使用的季节, 太湖地区农田水稻土土壤磷素地表径流流失的主要形态是颗粒态磷.陈敏鹏等[49]发现, 太湖春季东南风或偏南风引起强烈的沉积物再悬浮, 水体中颗粒态磷显著增高, 同时也使悬浮颗粒物对溶解态活性磷的吸附作用加强.秋季西北湖区PN/PP比值小于16, PP含量增高受结构因素和随机因素的双重影响.夏、冬季节PN/PP比值小于16, PP含量增高主要与浮游植物新陈代谢有关.
西北湖区颗粒态氮磷比四季都处于利于藻类生长的范围内, 这种现象无疑对太湖水体的蓝藻暴发和富营养化具有潜在的促进作用, 因此必须重视颗粒态氮磷营养盐对湖泊富营养化的影响研究.
4 结论(1) 时空分布上, 水体中氮、磷含量整体表现为冬季高于其他季节, 呈现由西北湖区向东南湖区递减的特征; 颗粒态氮、磷与叶绿素a含量则表现为夏季高于其他季节, 冬季高值区均位于南部湖区, 其余季节高值区集中在西北湖区.
(2) 随季节变化, 太湖草、藻型湖区氮磷营养盐形态组成发生了大的变化; 藻型湖区由冬季以硝酸盐氮和有机磷为主, 转变为其余季节以颗粒态氮磷为主, 而草型湖区由冬季以颗粒态氮磷为主, 转变为其余季节以氨氮和有机氮磷为主.
(3) 营养结构上, 藻型湖区总氮/总磷比值由秋冬季节大于16, 降低为夏春季节的小于16;而草型湖区却由秋冬季节小于16, 升高为夏春季节大于16.溶解态氮磷比在藻型湖区的空间变化规律与总氮/总磷比值一致, 而在草型湖区溶解态氮磷比由秋季小于16, 升高为夏、冬、春季节大于16.颗粒态氮磷比时空变化均不显著(t检验, P>0.05), 各季节藻型湖区颗粒态氮磷比值均小于16, 草型湖区均大于16.
| [1] |
孔繁翔, 高光. 大型浅水富营养化湖泊中蓝藻水华形成机理的思考[J]. 生态学报, 2005, 25(3): 589-595. Kong F X, Gao G. Hypothesis on cyanobacteria bloom-forming mechanism in large shallow eutrophic lakes[J]. Acta Ecologica Sinica, 2005, 25(3): 589-595. DOI:10.3321/j.issn:1000-0933.2005.03.028 |
| [2] |
姚昕, 张运林, 朱广伟, 等. 湖泊草、藻来源溶解性有机质及其微生物降解的差异[J]. 环境科学学报, 2014, 34(3): 688-694. Yao X, Zhang Y L, Zhu G W, et al. Different degradation mechanism of dissolved organic matter derived from phytoplankton and macrophytes in Lake Taihu, China[J]. Acta Scientiae Circumstantiae, 2014, 34(3): 688-694. |
| [3] |
吕伟伟, 姚昕, 张保华, 等. 太湖颗粒态有机质的荧光特征及环境指示意义[J]. 环境科学, 2018, 39(5): 2056-2066. Lü W W, Yao X, Zhang B H, et al. Fluorescent characteristics and environmental significance of particulate organic matter in Lake Taihu, China[J]. Environmental Science, 2018, 39(5): 2056-2066. |
| [4] | Xu H, Paerl H W, Qin B Q, et al. Nitrogen and phosphorus inputs control phytoplankton growth in eutrophic Lake Taihu, China[J]. Limnology and Oceanography, 2010, 55(1): 420-432. DOI:10.4319/lo.2010.55.1.0420 |
| [5] |
聂泽宇, 梁新强, 邢波, 等. 基于氮磷比解析太湖苕溪水体营养现状及应对策略[J]. 生态学报, 2012, 32(1): 48-55. Nie Z Y, Liang X Q, Xing B, et al. The current water trophic status in Tiaoxi River of Taihu Lake watershed and corresponding coping strategy based on N/P ratio analysis[J]. Acta Ecologica Sinica, 2012, 32(1): 48-55. |
| [6] |
何东, 张毅敏, 杨飞, 等. 太湖藻源性颗粒物降解过程中营养盐转化及其生态效应[J]. 中国环境科学, 2016, 36(3): 899-907. He D, Zhang Y M, Yang F, et al. The transformation of the nutrient in the degradation process of the phytoplankton-derived particulate organic matter and it's ecological effect[J]. China Environmental Science, 2016, 36(3): 899-907. DOI:10.3969/j.issn.1000-6923.2016.03.036 |
| [7] | Tang X M, Gao G, Chao J Y, et al. Dynamics of organic-aggregate-associated bacterial communities and related environmental factors in Lake Taihu, a large eutrophic shallow lake in China[J]. Limnology and Oceanography, 2010, 55(2): 469-480. DOI:10.4319/lo.2010.55.2.0469 |
| [8] | Qin B Q, Zhu G W, Gao G, et al. A drinking water crisis in Lake Taihu, China:linkage to climatic variability and lake management[J]. Environmental Management, 2010, 45(1): 105-112. DOI:10.1007/s00267-009-9393-6 |
| [9] | Qin B Q, Xu P Z, Wu Q L, et al. Environmental issues of Lake Taihu, China[J]. Hydrobiologia, 2007, 581(1): 3-14. DOI:10.1007/s10750-006-0521-5 |
| [10] | Qin B Q, Li W, Zhu G W, et al. Cyanobacterial bloom management through integrated monitoring and forecasting in large shallow eutrophic Lake Taihu (China)[J]. Journal of Hazardous Materials, 2015, 287: 356-363. DOI:10.1016/j.jhazmat.2015.01.047 |
| [11] | Zhou Y Q, Zhang Y L, Shi K, et al. Lake Taihu, a large, shallow and eutrophic aquatic ecosystem in China serves as a sink for chromophoric dissolved organic matter[J]. Journal of Great Lakes Research, 2015, 41(2): 597-606. DOI:10.1016/j.jglr.2015.03.027 |
| [12] | 金相灿, 屠清瑛. 湖泊富营养化调查规范[M]. 第二版. 北京: 中国环境科学出版社, 1990. |
| [13] |
张峰, 杜群, 葛宏立, 等. 基于地统计学和CFI样地的浙江省森林碳空间分布研究[J]. 生态学报, 2012, 32(16): 5275-5286. Zhang F, Du Q, Ge H L, et al. Spatial distribution of forest carbon in Zhejiang Province with geostatistics based on CFI sample plots[J]. Acta Ecologica Sinica, 2012, 32(16): 5275-5286. |
| [14] | Ozcan A U, Erpul G, Basaran M, et al. Use of USLE/GIS technology integrated with geostatistics to assess soil erosion risk in different land uses of Indagi Mountain Pass-ÇankƖrƖ, Turkey[J]. Environmental Geology, 2008, 53(8): 1731-1741. DOI:10.1007/s00254-007-0779-6 |
| [15] |
曾春阳, 唐代生, 唐嘉锴. 森林立地指数的地统计学空间分析[J]. 生态学报, 2010, 30(13): 3465-3471. Zeng C Y, Tang D S, Tang J K. Spatial pattern analysis of forest ecosystem site index using geostatistical technology[J]. Acta Ecologica Sinica, 2010, 30(13): 3465-3471. |
| [16] |
车磊, 王海起, 费涛, 等. 基于多尺度最小二乘支持向量机优化的克里金插值方法[J]. 地球信息科学学报, 2017, 19(8): 1001-1010. Che L, Wang H Q, Fei T, et al. Kriging interpolation method optimized by multi-scale least squares support vector machine[J]. Journal of Geo-Information Science, 2017, 19(8): 1001-1010. |
| [17] |
彭景涛, 李国胜, 傅瓦利, 等. 青海三江源地区退化草地土壤全氮的时空分异特征[J]. 环境科学, 2012, 33(7): 2490-2496. Peng J T, Li G S, Fu W L, et al. Temporal-spatial variations of total nitrogen in the degraded grassland of Three-River Headwaters region in Qinghai Province[J]. Environmental Science, 2012, 33(7): 2490-2496. |
| [18] |
符刚, 曾强, 赵亮, 等. 基于GIS的天津市饮用水水质健康风险评价[J]. 环境科学, 2015, 36(12): 4553-4560. Fu G, Zeng Q, Zhao L, et al. Health risk assessment of drinking water quality in Tianjin based on GIS[J]. Environmental Science, 2015, 36(12): 4553-4560. |
| [19] |
王建民, 苏巧梅, 杜孙稳. Kriging空间插值方法在地表形变监测中的应用[J]. 中国土地科学, 2013, 27(12): 87-90. Wang J M, Su Q M, Du S W. The Application of kriging spatial interpolation method in monitoring surface deformation[J]. China Land Sciences, 2013, 27(12): 87-90. DOI:10.3969/j.issn.1001-8158.2013.12.014 |
| [20] |
赵科理, 傅伟军, 叶正钱, 等. 电子垃圾拆解区土壤重金属空间异质性及分布特征[J]. 环境科学, 2016, 37(8): 3151-3159. Zhao K L, Fu W J, Ye Z Q, et al. Spatial variation of soil heavy metals in an e-waste dismantling area and their distribution Characteristics[J]. Environmental Science, 2016, 37(8): 3151-3159. |
| [21] |
汪明, 武晓飞, 李大鹏, 等. 太湖梅梁湾不同形态磷周年变化规律及藻类响应研究[J]. 环境科学, 2015, 36(1): 80-86. Wang M, Wu X F, Li D P, et al. Annual variation of different phosphorus forms and response of algae growth in Meiliang Bay of Taihu Lake[J]. Environmental Science, 2015, 36(1): 80-86. DOI:10.3969/j.issn.1673-7148.2015.01.012 |
| [22] |
冯露露, 李正魁, 周涛. 太湖浮游植物和各形态无机氮的时空分布特征[J]. 湖泊科学, 2012, 24(5): 739-745. Feng L L, Li Z K, Zhou T. Temporal and spatial distributions of phytoplankton and various forms of inorganic nitrogen in Lake Taihu[J]. Journal of Lake Sciences, 2012, 24(5): 739-745. DOI:10.3969/j.issn.1003-5427.2012.05.015 |
| [23] |
朱广伟, 秦伯强, 张运林, 等. 2005-2017年北部太湖水体叶绿素a和营养盐变化及影响因素[J]. 湖泊科学, 2018, 30(2): 279-295. Zhu G W, Qin B Q, Zhang Y L, et al. Variation and driving factors of nutrients and chlorophyll-a concentrations in northern region of Lake Taihu, China, 2005-2017[J]. Journal of Lake Sciences, 2018, 30(2): 279-295. |
| [24] |
李传华, 赵军, 师银芳, 等. 基于变异系数的植被NPP人为影响定量研究——以石羊河流域为例[J]. 生态学报, 2016, 36(13): 4034-4044. Li C H, Zhao J, Shi Y F, et al. The impact of human activities on net primary productivity based on the coefficient of variation:A case study of the Shiyang River Basin[J]. Acta Ecologica Sinica, 2016, 36(13): 4034-4044. |
| [25] |
于文丽, 蒲英霞, 陈刚, 等. 基于空间自相关的中国省际人口迁移模式与机制分析[J]. 地理与地理信息科学, 2012, 28(2): 44-49. Yu W L, Pu Y X, Chen G, et al. Spatial analysis of the patterns and mechanism of inter-provincial migration flows in China[J]. Geography and Geo-Information Science, 2012, 28(2): 44-49. |
| [26] |
金颖薇, 朱广伟, 许海, 等. 太湖水华期营养盐空间分异特征与赋存量估算[J]. 环境科学, 2015, 36(3): 936-945. Jin Y W, Zhu G W, Xu H, et al. Spatial distribution pattern and stock estimation of nutrients during bloom season in Lake Taihu[J]. Environmental Science, 2015, 36(3): 936-945. |
| [27] | Collins S M, Oliver S K, Lapierre J F, et al. Lake nutrient stoichiometry is less predictable than nutrient concentrations at regional and sub-continental scales[J]. Ecological Applications, 2017, 27(5): 1529-1540. DOI:10.1002/eap.2017.27.issue-5 |
| [28] | Berthold M, Karsten U, Von Weber M, et al. Phytoplankton can bypass nutrient reductions in eutrophic coastal water bodies[J]. Ambio, 2018, 47(S1): 146-158. DOI:10.1007/s13280-017-0980-0 |
| [29] | Robertson B P, Savage C. Mud-entrained macroalgae utilise porewater and overlying water column nutrients to grow in a eutrophic intertidal estuary[J]. Biogeochemistry, 2018, 139(1): 53-68. DOI:10.1007/s10533-018-0454-x |
| [30] |
周楠, 赵卫红, 苗辉. 獐子岛附近海域溶解有机物的荧光特征[J]. 光谱学与光谱分析, 2012, 32(4): 1007-1011. Zhou N, Zhao W H, Miao H. Fluorescence characteristics of dissolved organic matter in the sea nearby Zhangzi Island[J]. Spectroscopy and Spectral Analysis, 2012, 32(4): 1007-1011. DOI:10.3964/j.issn.1000-0593(2012)04-1007-05 |
| [31] |
曹阳, 滕彦国, 王威, 等. 金积水源地地下水体重金属空间分布及来源[J]. 环境科学与技术, 2013, 36(11): 163-167, 198. Cao Y, Teng Y G, Wang W, et al. Investigation of heavy metals in Jinji Groundwater Source of Wuzhong City:spatial distribution and source apportionment[J]. Environmental Science & Technology, 2013, 36(11): 163-167, 198. |
| [32] |
叶宏萌, 袁旭音, 葛敏霞, 等. 太湖北部流域水化学特征及其控制因素[J]. 生态环境学报, 2010, 19(1): 23-27. Ye H M, Yuan X Y, Ge M X, et al. Water chemistry characteristics and controlling factors in the northern rivers in the Taihu Basin[J]. Ecology and Environmental Sciences, 2010, 19(1): 23-27. DOI:10.3969/j.issn.1674-5906.2010.01.005 |
| [33] | 张润宇, 王立英, 吴丰昌, 等.太湖春季水体磷形态的空间分布[A].见: 第十三届世界湖泊大会论文集[C].武汉: 中国环境科学学会, 中国环境保护产业协会, 2009. |
| [34] |
李成高.环境理化因子对集约化养殖海域沉积物中亚硝态氮积累和释放的影响[D].青岛: 中国海洋大学, 2007. Li C G. Influence of physical and chemical factors on nitrite nitrogen accumulation and release in sediments of the intensive mariculture area[D]. Qingdao: Ocean University of China, 2007. |
| [35] |
孔繁翔, 马荣华, 高俊峰, 等. 太湖蓝藻水华的预防、预测和预警的理论与实践[J]. 湖泊科学, 2009, 21(3): 314-328. Kong F X, Ma R H, Gao J F, et al. The theory and practice of prevention, forecast and warning on cyanobacteria bloom in Lake Taihu[J]. Journal of Lake Sciences, 2009, 21(3): 314-328. DOI:10.3321/j.issn:1003-5427.2009.03.002 |
| [36] |
蔡琨, 秦春燕, 李继影, 等. 基于浮游植物生物完整性指数的湖泊生态系统评价——以2012年冬季太湖为例[J]. 生态学报, 2016, 36(5): 1431-1441. Cai K, Qin C Y, Li J Y, et al. Preliminary study on phytoplanktonic index of biotic integrity (P-IBI) assessment for lake ecosystem health:a case of Taihu Lake in Winter, 2012[J]. Acta Ecologica Sinica, 2016, 36(5): 1431-1441. |
| [37] | 艾鹰.微囊藻在太湖越冬及复苏的时空格局[D].北京: 中国科学院大学, 2014. |
| [38] | Tong Y D, Qiao Z, Wang X J, et al. Human activities altered water N:P ratios in the populated regions of China[J]. Chemosphere, 2018, 210: 1070-1081. DOI:10.1016/j.chemosphere.2018.07.108 |
| [39] |
张佳磊, 郑丙辉, 刘德富, 等. 三峡水库大宁河支流浮游植物演变过程及其驱动因素[J]. 环境科学, 2017, 38(2): 535-546. Zhang J L, Zheng B H, Liu D F, et al. Succession pattern of phytoplankton of Daning River in the Three Gorges Reservoir and its driving factors[J]. Environmental Science, 2017, 38(2): 535-546. |
| [40] | Riley J P, Skirrow G. Chemical oceanography (2nd ed.)[J]. Beijing:Academic Press, 1982. |
| [41] |
张云, 王圣瑞, 段昌群, 等. 滇池沉水植物生长过程对间隙水氮、磷时空变化的影响[J]. 湖泊科学, 2018, 30(2): 314-325. Zhang Y, Wang S R, Duan C Q, et al. Spatial-temporal variations of nitrogen and phosphorus forms in sediment porewater as affected by submerged plant in Lake Dianchi[J]. Journal of Lake Sciences, 2018, 30(2): 314-325. |
| [42] |
周笑白, 张宁红, 张咏, 等. 太湖蓝藻的时空变化规律及治理方法[J]. 生态环境学报, 2013, 22(12): 1930-1935. Zhou X B, Zhang N H, Zhang Y, et al. The temporal and spatial distribution pattern of cyanobacteria and its control method in Taihu Lake[J]. Ecology and Environmental Sciences, 2013, 22(12): 1930-1935. DOI:10.3969/j.issn.1674-5906.2013.12.012 |
| [43] |
赵林林, 朱广伟, 顾钊, 等. 太湖水体氮、磷赋存量的逐月变化规律研究[J]. 水文, 2013, 33(5): 28-33, 45. Zhao L L, Zhu G W, Gu Z, et al. Monthly variation of nitrogen and phosphorus volume in Taihu Lake, China[J]. Journal of China Hydrology, 2013, 33(5): 28-33, 45. DOI:10.3969/j.issn.1000-0852.2013.05.006 |
| [44] | Zhang Y, Song C L, Ji L, et al. Cause and effect of N/P ratio decline with eutrophication aggravation in shallow lakes[J]. Science of the Total Environment, 2018, 627: 1294-1302. DOI:10.1016/j.scitotenv.2018.01.327 |
| [45] | Chao J Y, Zhang Y M, Kong M, et al. Long-term moderate wind induced sediment resuspension meeting phosphorus demand of phytoplankton in the large shallow eutrophic Lake Taihu[J]. PLoS One, 2017, 12(3): e173477. |
| [46] | Hudson J J, Taylor W D, Schindler D W. Planktonic nutrient regeneration and cycling efficiency in temperate lakes[J]. Nature, 1999, 400(6745): 659-661. DOI:10.1038/23240 |
| [47] |
文威, 孙学明, 孙淑娟, 等. 海河底泥氮磷营养物静态释放模拟研究[J]. 农业环境科学学报, 2008, 27(1): 295-300. Wen W, Sun X M, Sun S J, et al. Release of phosphorus and nitrogen from Haihe River sediments[J]. Journal of Agro-Environment Science, 2008, 27(1): 295-300. DOI:10.3321/j.issn:1672-2043.2008.01.054 |
| [48] |
张红爱. 太湖地区典型水稻土土壤磷素径流流失及其吸持特征的研究[J]. 安徽农业科学, 2008, 36(18): 7788-7792. Zhang H A. Research on P runoff loss in paddy soils in the Taihu Lake region and its sorption characters[J]. Journal of Anhui Agricultural Sciences, 2008, 36(18): 7788-7792. DOI:10.3969/j.issn.0517-6611.2008.18.118 |
| [49] |
陈敏鹏, 郭宝玲, 刘昱, 等. 磷元素物质流分析研究进展[J]. 生态学报, 2015, 35(20): 6891-6900. Chen M P, Guo B L, Liu Y, et al. Research on phosphorus flow analysis:Progress and perspectives[J]. Acta Ecologica Sinica, 2015, 35(20): 6891-6900. |