 2022, Vol. 43
2022, Vol. 43

2. 中国科学院大学, 北京 100049;
3. 湖南师范大学资源与环境科学学院, 长沙 410006;
4. 湖南艾布鲁环保科技股份有限公司, 长沙 410000
 , XIA Meng-hua1,2 , JIANG Lei1,2 , PENG Jian1,2 , CHEN Kun1,3 , ZHAO Cong-fang1,3 , LI Xi1
, XIA Meng-hua1,2 , JIANG Lei1,2 , PENG Jian1,2 , CHEN Kun1,3 , ZHAO Cong-fang1,3 , LI Xi1 , MENG Cen1 , ZENG Rui4 , WANG Dong4 , LI Yu-yuan1 , WU Jin-shui1
, MENG Cen1 , ZENG Rui4 , WANG Dong4 , LI Yu-yuan1 , WU Jin-shui1
2. University of Chinese Academy of Sciences, Beijing 100049, China;
3. College of Resources and Environmental Sciences, Hunan Normal University, Changsha 410006, China;
4. Hunan Airbluer Environmental Technology Co., Ltd., Changsha 410000, China
根据文献[1], 全国水污染物排放量:化学需氧量(COD)2 143.98万t、氨氮(NH4+-N)96.34万t和总氮(TN)304.14万t.其中畜禽养殖业排放量:COD 1 000.53万t、NH4+-N 11.09万t和TN 59.63万t, 分别占全国排放总量的53.33%、88.49%和80.39%.因此, 随着养殖业发展水环境将面临污染物治理困难, 水体的严重富营养化等问题.
目前, 人工湿地治理养殖废水受到国内外推广和应用[2, 3].人工湿地治理模式中将基质填料与湿地植物合理搭配, 可提高污染物的净化效果.传统多孔材料不仅为植物和微生物的生长提供载体且一般带负电荷, 通过物理吸附和离子交换作用可直接吸附N、P等营养物[4].然而, 湿地系统中由于碳源不足, 造成去除效率降低, 基质填料由传统无机材料逐渐向有机碳源的新材料演化升级.由于向湿地内投加液态碳源不易于控制剂量且成本昂贵.因此, 选用作物秸秆作为基质材料既满足对污染物的拦截又大幅降低了材料成本, 是一种对农业副产品进行再利用的新途径.
有研究表明, 在硝化细菌与反硝化细菌的参与下, 污水中氮的去除主要通过硝化和反硝化作用[5, 6].其中NH4+-N的转化(硝化作用为主)是一个好氧过程, 而通过前期试验发现, 添加秸秆材料的生物基质系统处于厌氧条件下(DO<0.2 mg ·L-1), 反硝化细菌在厌氧环境下将NO3-还原为N2(NO3-→NO2-→NO→N2O→N2)脱离水体.厌氧氨氧化作用是指厌氧氨氧化菌在厌氧环境下, 以NH4+为电子供体, NO2-为电子受体将氨氮直接转化为氮气的过程(NH4++NO2-→N2+H2O)[7, 8].因此, 为进一步阐释生物基质系统内参与氮循环过程的微生物作用, 本文对参与氮循环的主要微生物功能基因丰度进行测定.
前期, 本课题组对比分析了我国三大粮食作物秸秆:麦秸、稻草和玉米秆对猪场废水主要污染物去除效率.结果表明, 在6个月试验周期中, 麦秸对COD、TN和TP的去除效果最好, 去除率分别为32.1%、40.9%和33.3%, 稻草对NH4+-N的去除效果最好达43.4%, 且通过干物质(DM)及纤维成分(CF)的降解过程分析, 建议在湖南地区基质材料更换周期为5~6个月[9].本文延续之前的工作, 通过为期1 a的野外试验, 研究秸秆材料处理养殖废水中氮的周年去除效果, 并补充分析其相关微生物功能基因丰度动态变化, 这对于推动秸秆材料生态治理养殖废水技术具有重要实际指导意义.
1 材料与方法 1.1 研究区概况试验区位于湖南省长沙县金井镇湘丰村的中国科学院长沙农业环境观测研究站, 区内为典型亚热带湿润季风气候, 多年平均气温为17.5℃, 最低气温为-5.2℃, 最高气温为40.1℃.
1.2 试验设置试验小区由3个同等大小水泥池串联(长×宽×深:100 cm×50 cm×50 cm)组成, 设置稻草、麦秸、玉米秆和空白对照共4个处理, 每级池内填充12.5 kg大小均匀风干处理的作物秸秆, 并设置3次重复.进水由蠕动泵24 h连续泵入基质池内, 水力停留时间为7 d.
试验时间为2017年9月~2018年9月.进水方式、生物基质材料填装和后端人工湿地如图 1所示.

|
图 1 野外试验现场示意 Fig. 1 Schematic diagram of the field experiments |
本试验进水为白沙乡大花养猪场经过厌氧处理的沼液废水, 水质具有一定的波动性, 各主要污染物成分(TN、NH4+-N和NO3--N)全年浓度变化范围分别为285.5~400.7、249.8~329.0和0.48~0.82 mg ·L-1.
稻草、玉米秆取自中国科学院长沙农业环境站附近牛场, 麦秸购自河南省开封市, 3种秸秆常规营养成分含量见表 1.
|
|
表 1 秸秆主要组成成分含量/g·kg-1 Table 1 Components of different straw materials/g·kg-1 |
1.4 样品采集与分析 1.4.1 水样的采集与测定
每周采集一次水样, 每次水样采集储水池及三级基质池出水口水样, 共42个样品.每次使用200 mL采样瓶采集水面下5 cm处水样, 立即带回实验室分析或放于-20℃冰箱保存.分析测定指标包括:pH、温度(T)、氧化还原电位(Eh)、溶解氧(DO)、NH4+-N、NO3--N和TN.其中, 指标(pH、T、Eh和DO)用便携式DO测定仪对每次进水和收集后的瞬时出水进行测量, 水样中NH4+-N与NO3--N的浓度直接取50 mL左右的水样离心后取上清液进行稀释, 用流动注射仪(AA3, 德国SEAL公司)测定, 剩余的水样(未离心的水样)用于TN的测定.TN采用碱性过硫酸钾消解-紫外分光光度法测定.严格按照文献[10]方法测定.
1.4.2 基质材料样品的采集与测定控制试验于2017年9~12月每月采集两次, 2018年1~9月每月采集1次.取预先填埋于基质池中的白色尼龙网袋, 清洗后放于信封袋内, 于105℃烘箱中杀青30 min, 然后于80℃下烘至恒重, 称干重.分别称取并记录约0.2 g样品于100 mL锥形瓶内, 用于测定秸秆材料中TN含量.并用弯颈小漏斗倒扣在瓶口防止液体飞溅.测定方法为浓硫酸双氧水(H2SO4-H2 O2)法消解, TN取上清液使用AA3流动分析仪采用凯氏定氮法测定[11].
1.4.3 微生物样品的采集与测定每3个月采集一次, 取储水池及各级基质池内混匀水样100 mL, 立即进行抽滤(水样浑浊时先低速离心5 min后对上清液进行抽滤), 滤膜使用美国Millipore混合纤维素酯滤膜(AAWP04700, 孔径0.45 μm).将滤膜和离心后的废渣一同转移到试剂盒(PowerWater DNA Isolation kit)内进行DNA提取操作.并利用1%琼脂糖凝胶电泳检测抽提的基因组DNA, 核对基因组DNA的完整性与浓度, 并用NanoDrop核酸蛋白测定仪(ND-1000)测定DNA的浓度和质量.采用表 2所示的反应体系(Taq PCR MasterMix天根, 中国)、上下游引物和反应条件PCR扩增目的基因[12, 13], DNA模板约200 ng.
|
|
表 2 质粒构建1) Table 2 Plasmid construction |
构建基因的标准曲线, 首先构建克隆文库, 然后选择阳性克隆子, 从中吸取10 μL菌液于1 mL含氨苄青霉素LB培养基中, 37℃摇床培养10 h, 提取重组质粒并测定浓度, 进一步换算成单位体积的拷贝数. 10倍梯度稀释作为标准曲线的模板, 标准曲线的范围在102~108 copies ·μL-1.
全部样本按照正式试验条件进行, 每个样本3个重复, 将同一样本的PCR产物混合后用2%琼脂糖凝胶电泳检测, 使用AxyPrepDNA凝胶回收试剂盒(AXYGEN公司)切胶回收PCR产物, Tris-HCl洗脱; 2%琼脂糖电泳检测.参照电泳初步定量结果, 将PCR产物用QuantiFluorTM-ST蓝色荧光定量系统进行检测定量, 之后按照每个样品的测序量要求, 进行相应比例的混合, -80℃保存待后续测量.
1.5 数据处理与统计 1.5.1 基质处理系统污染物拦截计算各污染指标去除率(r)、进水负荷(m0)、出水负荷(m1)和去除负荷(p)分别采用如下公式计算:
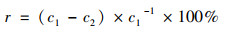
|
(1) |
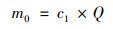
|
(2) |
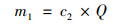
|
(3) |
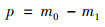
|
(4) |
式中, c1和c2为基质池进、出水浓度(mg ·L-1); Q为基质池进水流量(L ·d-1).
1.5.2 秸秆TN含量的计算秸秆中全氮含量(cN)的计算公式为:
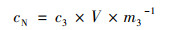
|
(5) |
式中, m3为样品质量(g); c3为反应体系TN浓度(mg ·L-1); V为反应体系体积(L), 一般是0.1 L; cN表示每kg秸秆中TN含量(g ·kg-1).
1.5.3 基因丰度的计算质粒拷贝数通过下列公式计算:
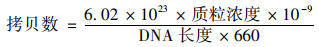
|
(6) |
式中, 拷贝数用(copies ·μL-1)表示; 质粒浓度用(ng ·μL-1)表示; DNA长度为质粒碱基对数(bp)[17].由于上机数据得出的为CT值, 因此将质粒模板实际CT值与式中计算求得的拷贝数的对数值整理并计算二者的标准曲线, 建立CT值与拷贝数对数值的回归方程.
计算每L废水中的拷贝数(copies ·L-1):
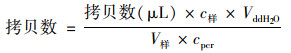
|
(7) |
式中, c样为样品DNA浓度(ng ·μL-1); VddH2O为稀释质粒所加水的体积(μL); V样为进行抽滤用于提取DNA所用样品体积, 一般为0.05 L; cpcr为上机稀释后DNA浓度(一般是将样品DNA稀释100倍)[18, 19].这样将样品CT值代入标准曲线后可得到实际拷贝数的对数值, 再次转化后求得样品基因的拷贝数.
DNA纯度D260/D280在1.6~2.0之间(等于1.8较好), 若<1.6时表明有蛋白质或酚污染; 若>2.0时表明可能有异硫氰酸残存[20].
1.5.4 统计分析采用Microsoft Excel 2013进行数据分析, Origin作图, 并用SPSS 19进行one-way ANOVA单因素方差分析, 在显著性水平P<0.05下表示差异显著, P<0.01表示差异极显著.数据均在重复试验中取平均值与标准差.
2 结论与分析 2.1 生物基质系统周年脱氮效果对猪场废水不同形态N含量测定分析表明, 进水TN、NH4+-N和NO3--N污染负荷分别为24.83~26.78、29.73~2.66和0.05~0.06 g ·d-1时, 3种秸秆材料均能对不同形态氮进行去除(表 3). 3种作物秸秆对养殖废水TN和NH4+-N等主要污染物的去除效果均以前6个月为最佳.添加前6个月麦秸对TN的去除效果最好, 最高达40.9%, 稻草对NH4+-N的去除效果最佳, 最高达43.4%.
|
|
表 3 基质系统周年进出水浓度及去除效率 Table 3 Annual influent/effluent concentrations and removal efficiency in straw material system |
2.1.1 总氮去除效果动态分析
从基质池进出水TN浓度的周年动态变化情况分析可见图 2.试验开展后, 前36 d不同基质材料对TN的去除效果呈波动上升趋势, 随后麦秸与稻草材料的去除作用略有降低, 但玉米秆继续上升, 在试验进行77~99 d左右, 秸秆材料对污染物去除率出现下降变化, 其中稻草材料下降趋势较为平缓.前5个月, 稻草材料对TN的去除作用较玉米秆和麦秸更加稳定, 去除率能够稳定在40%左右, 最高可去除达45%以上.但后期下降较为明显, 最低降至14%以下.相比之下, 玉米秆材料对TN去除效果较差, 麦秸材料对TN具有较强去除能力的周期较短, 去除率高于40%的情况可持续2个月左右, 进入冬季后去除效果能保持在30%, 经过6个月后随气温升高略有提高.分析周年结果得出, 经基质系统处理后ρ(TN)由285.5~400.7 mg ·L-1下降到187.5~294.8 mg ·L-1, 显著低于对照处理(226.1~301.9 mg ·L-1).

|
图 2 基质池全年TN进出水浓度及去除率动态分析 Fig. 2 Dynamic analysis of TN annual influent/effluent concentrations and removal efficiency in straw material system |
秸秆材料前期对NH4+-N的去除规律与TN相似, 分析全年数据发现3种秸秆材料对NH4+-N的去除效果为:稻草>麦秸>玉米秆.对NH4+-N去除效果动态分析见图 3, 可以看出前3周先迅速上升而后下降, 秸秆材料经3~4周的稳定期后去除效果较为明显, 4~8周后脱氮作用逐渐增强, 最高可去除47%以上的NH4+-N.经过6个月试验周期, 秸秆材料作为基质填料, 对高浓度养殖废水处理效果明显, 麦秸、稻草和玉米秆对NH4+-N去除率平均值分别为42.2%、43.4%和41.7%, 而对照组对NH4+-N去除率平均值仅为11.96%.进入春季温度上升后, 玉米秆对NH4+-N的去除效果有缓慢回升, 相比稻草与麦秸材料去除效果呈逐渐下降的趋势.进入试验后期, 基质池进出水浓度已无明显改善, 去除效果直线下降.试验末期时, 玉米秆对NH4+-N拦截效率低至7.4%, 稻草及麦秸有效去除10%左右的NH4+-N.因此, 定期向基质池内补充秸秆材料可以增强对污染物的去除效果.

|
图 3 基质池全年NH4+-N进出水浓度及去除率动态分析 Fig. 3 Dynamic analysis of NH4+-N annual influent/effluent concentrations and removal efficiency in straw material system |
基质池进出水NO3--N浓度较低如图 4, 进水ρ(NO3--N)为0.08~1.63 mg ·L-1, 基质材料对其去除效果的波动变化较为剧烈.基质池内DO较低, 硝化过程中氧气不充足产生NO2--N.对数据分析后发现, 麦秸对NO3--N去除效果最高达53.3%, 最低为11.3%, 稻草对NO3--N去除效率范围在12.3% ~64%之间, 高于玉米杆的去除范围(4.5% ~12.0%).前6个月麦秸、稻草和玉米秆对NO3--N去除率平均值分别为38%、40.7%和31.56%, 处理间差异显著, 年去除率平均值依次为:玉米秆[(33.78±15.37)%]>麦秸[(32.12±14.88)%]>稻草[(31.95±13.6)%], 显著高于空白对照组去除率20%以上.

|
图 4 基质池全年NO3--N进出水浓度及去除率动态分析 Fig. 4 Dynamic analysis of NO3--N annual influent/effluent concentrations and removal efficiency in straw material system |
基质池系统中T、pH、Eh和DO的变化特征见图 5.本试验期间, 年平均气温为18.5℃, 最高气温为40.4℃, 少数低气温天气达-3.2℃, 无霜期约占全年的75%.基质池内废水最高温度为26.8℃, 最低温度为5.7℃.前6个月中养殖废水流经基质池过程中Eh和DO的变化范围为-37.4~-29.3 mV和0.04~0.17 mg ·L-1, 两者均表现出随梯级的增加而不断上升的变化趋势.废水流经基质池前DO<0.2 mg ·L-1, 长期处于厌氧条件, 经基质池处理后, 废水中溶解氧逐渐增加.系统中pH值的变化范围为7.05~7.74, 呈弱碱性条件, 随着基质池梯级的增加缓慢降低.各级基质池Eh、pH及DO差异显著(P<0.05).后6个月随温度上升, 基质池内DO含量下降, 添加秸秆材料的各处理间差异逐渐减小; pH值受温度及时间影响波动不大, 后6个月pH值在逐级基质池间差异缩小, 改变量低于前期; Eh受环境改变产生变化, 如秸秆分解的影响及其他环境因子的影响, 试验后期Eh升高, 还原性减弱.

|
图 5 主要水质理化指标分析 Fig. 5 Physicochemical properties in straw material system |
利用实时定量PCR(Q-PCR)测定了三级基质系统废水中氮循环微生物功能基因丰度, 包括hzsB、nirK、nirS、AOA、AOB和16S rRNA, 见图 6.

|
不同大写字母表示各级基质池间具有显著差异(P<0.05), 不同小写字母表示不同处理间差异 图 6 不同处理hzsb, nirK, nirS, AOA, AOB和16S rRNA基因丰度分析 Fig. 6 Abundance of hzsb, nirK, nirS, AOA, AOB, and 16S rRNA genes under different treatments |
生物基质系统处理养殖废水过程中, 各级基质池内的微生物功能基因丰度存在显著的统计差异(P<0.05), 每L废水中hzsB、nirK、nirS、AOA、AOB和16S rRNA基因丰度均表现为:一级>二级>三级.
对同级间不同处理组的分析得出:添加秸秆90 d后, 1 L废水中nirK基因的拷贝数由1.94×106 copies升至1.80×109 copies(麦秸>稻草>玉米秆), 进行3~6个月时, 添加秸秆材料后nirK基因丰度增至2.06×109 copies ·L-1, 进行9~12个月时从6.45×109 copies ·L-1降至4.45×109 copies ·L-1, 且随试验进行, 对全年结果分析得添加秸秆的3个处理组间差异极显著(P<0.01); nirS基因由1.54×106 copies ·L-1升至1.58×109 copies ·L-1(麦秸>玉米秆>稻草), 且三者在前3个月产生显著差异(P<0.05), 后期差异逐渐消失, 其丰度由6.18×109 copies ·L-1降至4.18×109 copies ·L-1; AOA和AOB基因丰度在3个月时均表现为(麦秸>玉米秆>稻草), 其中AOA由1.49×106copies ·L-1升至6.85×108 copies ·L-1; AOB由1.62×106 copies ·L-1升至1.31×109 copies ·L-1.周年分析得出3种秸秆在试验前9个月时, 各基质池废水中AOA(P<0.01)和AOB(P<0.05)差异显著, 试验后期差异性降低, AOA和AOB平均基因丰度由6.12×1010 copies ·L-1和4.93×109 copies ·L-1, 降至4.12×1010 copies ·L-1和3.93×109 copies ·L-1; hzsB由1.20×106 copies ·L-1升至2.31×109 copies ·L-1(稻草>玉米>麦秸).此外, 3种秸秆中hzsB功能基因在基质池内无明显差异, 试验后期平均丰度在1.71×108~3.55×108 copies ·L-1; 16S rRNA基因丰度由1.94×107 copies ·L-1升至8.90×1010 copies ·L-1(麦秸>稻草>玉米秆), 试验后期处理组间产生显著差异(P<0.01), 并且各基因丰度在逐级基质池内递减.添加麦秸的处理中含AOA、AOB、nirK和16S rRNA基因的微生物数量最高, 含hzsB基因和nirS基因的微生物分别在稻草和麦秸中最多.其中CK组各基因丰度均与其他处理组产生明显差异, 后期添加3种秸秆的处理组间差异性逐渐降低.结果表明, 与对照组相比, 添加秸秆材料的处理组基因的丰度均显著增加.
3.2 氮循环微生物功能基因丰度与环境因子相关性分析将全年水质环境因子(pH、Eh、DO)和废水中部分氮循环微生物功能基因丰度进行Pearson分析(见表 4), 结果表明二者显著相关且系统内微生物受DO及温度影响较大, AOA和AOB功能基因丰度与T呈显著正相关(P<0.05), 与DO呈显著正相关(P<0.05), 且与Eh、pH正相关, hzsB功能基因丰度与DO呈极显著负相关(P<0.01), 且与T、Eh和pH正相关.
|
|
表 4 养殖废水微生物功能基因丰度与环境因子间相关性分析1) Table 4 Correlation analysis between abundance of microbial functional genes and environmental factors |
同时, 相关分析表明功能基因丰度与废水中NH4+-N、TN和NO3--N浓度相关.AOA、AOB、nirK和hzsB基因均与NH4+-N和TN浓度呈正相关关系(P<0.05), 与NO3--N呈负相关关系, nirS基因与废水中NH4+-N和TN浓度呈显著负相关关系(P<0.05), 与NO3--N呈显著正相关关系.
4 讨论本研究选择我国3种主要农作物秸秆作为生物基质填料, 是由于农村地区秸秆作为农业副产品年产量较大, 如何通过廉价的加工程序就能使其再次发挥作用, 是秸秆资源化利用的有效方式.而将秸秆材料作为养殖废水的治污材料既符合农户的需求又符合秸秆资源化利用理念并解决其二次污染对环境造成的危害.有研究表明, 传统无机基质材料需向基质池投加不同类型的液体有机碳源, 而与液态碳源相比, 固体有机物质可以避免反硝化过程碳源过量或不足量的问题, 易于控制剂量且价格低廉[21].而针对农村地区高负荷畜禽养殖废水的C/N比较低的情况, 来源广且成本低的农作物秸秆可作为基质材料的首选.玉米秆、稻草和麦秸等木质纤维类物质, 纤维素、木质素和半纤维素, 纤维素所占比例最大, 未加工的玉米秆、稻草和麦秸等生物基质材料也可在微生物的作用下转化, 释放出单糖为反硝化提供碳源, 且渗透系数高[22, 23].同时, 作物秸秆与水生植物茎叶类似, 其表面的附着层为反硝化作用提供了微环境和物质基础[24, 25].因此, 它作为一种天然可再生的高分子材料, 具有多重优势应得到充分的利用.
对废水中污染物去除效果分析发现, 试验开始时, 秸秆材料对TN去除率呈缓慢上升后波动下降的变化趋势, 有可能试验前期以秸秆的吸附作用为主, 后期以微生物作用为主.在秸秆利用的有效期限内, 秸秆材料对NH4+-N去除效果高于TN与秸秆材料向水体释放有机物及无机氮的协同作用有关[26, 27].随试验进行, 3种秸秆材料稻草、麦秸与玉米秆中干物质由初始的86.33%、83.48%和81.05%降至20.62% ~28.79%、30.75% ~37.55%和38.78% ~47.07%, 有效成分已在前6个月降解[7], 导致秸秆材料在后6个月对污染物去除效果下降, 污染物周年去除率均在25.8% ~35.3%.进入试验末期, 3种基质材料对TN的去除效果降至10%左右, 显著低于有效期限内的净化效果, 微生物作用缓慢是受温度影响.并且结合秸秆分解过程中参与硝化、反硝化及厌氧氨氧化过程中所可利用的能源物质降低, 微生物脱氮作用减弱, 污染物去除效率迅速下降.其中空白对照组对NH4+-N的去除率在10%左右, 且受温度影响较大, 赵聪芳等[29]发现未添加秸秆材料的对照组N含量发生降低是由于底泥微生物参与硝化反硝化作用且存在氨挥发的发生.此外, 3种秸秆中TN含量表现出“快速-平缓”下降的变化趋势, 9月背景值含量最高(稻草>麦秸>玉米秆), 12月下降最快最高, 试验进行6个月后TN含量基本持平.麦秸中TN含量由初始时8.60 g ·kg-1降至2.00 g ·kg-1, 下降76.74%; 稻草中TN初始含量由最高12.39 g ·kg-1降至3.19 g ·kg-1, 下降74.25%; 玉米秆中TN含量由初始时7.98 g ·kg-1降至1.63 g ·kg-1下降最高达79.57%.
同时, 生物基质材料在处理养殖废水过程中的脱氮机制是复杂的, 本研究试图通过微生物硝化、反硝化和厌氧氨氧化过程功能基因丰度的变化初步解释其脱氮机制.试验初期正值长沙地区较热时期, 在微生物挂膜期间长沙步入冬季, 因此挂膜时期受温度影响变长, 进入冬季后虽然温度降低, 但也在微生物适应范围内, 且营养物质充足微生物缓慢生长.次年春天, 温度上升后微生物活性逐渐增加, 但试验后期由于营养成分的限制, 微生物的生长繁殖同时受到影响. T和DO是影响微生物活性的重要因素, 在最适温度范围内, T升高可显著提升微生物活性, 与国内外的研究结果一致[29~31].试验过程中, 基质池内废水中的DO不高于0.3 mg ·L-1适宜厌氧氨氧化细菌的繁殖, 随着废水在逐级基质池中处理净化, DO升高, hzsB基因丰度降低, 但各级基质池间pH变化范围不大, 均适宜微生物繁殖, 因此各功能基因受pH变化影响不显著.有研究表明[32, 33], 当人工湿地基质内DO<2 mg ·L-1时, 有利于污水中硝态氮的去除.Liu等[34]的研究也发现高DO能提高湿地系统中NH4+-N的去除率.由于当湿地系统中DO增加时, 好氧硝化细菌的硝化速率增强, 进而有更多的NH4+-N被氧化, 湿地系统中TN去除率随之增高.而基质池内为厌氧环境, 添加秸秆材料在湿地前端不仅为参与硝化/反硝化微生物提供附着位点及能量使其数量显著增加, 还证实基质池内发生厌氧氨氧化作用.因而, 尽管基质池内DO低至0.05~0.21 mg ·L-1基质池中仍可进行脱氮作用, 并表明厌氧氨氧化过程是基质消纳系统高浓度养殖废水氮去除途径之一.此外, 由于进水中NO3--N浓度较低且随着处理系统内含量仍保持稳定, 因此猜测系统内存在好氧反硝化菌和异养硝化菌[35]为同步硝化反硝化提供了有利条件.
由上述分析结果可知, 3种秸秆添加对基质池N转化微生物的影响略有差异, 且不同的外部环境因子对于生物基质消纳系统中微生物功能基因丰度均有着影响, 与王欢等[36]和宋壮壮等[37]研究结果较为一致.因此, 对于猪场沼液的处理, 创造合理的环境条件, 促进微生物特别是功能菌群的增加, 可以最大程度发挥生物基质材料及水体中微生物脱氮的功能, 是提高生物基质系统处理效果的有效途径之一.
筛选比较3种作物秸秆作为生物基质材料不仅在一定程度上能够降低废污染物浓度, 有利于末端生态湿地植物的生长, 而且达到养殖废水营养物质植物转化与生态治理的目的, 是基质系统中参与脱氮微生物挂膜的良好载体, 为人工湿地前端处理提供新的基质材料选择.
5 结论(1) 添加前6个月, 3种秸秆材料对NH4+-N和TN有较好的去除效果, 后6个月去除效果呈下降趋势.稻草与麦秸对TN及NH4+-N的去除效果最好, 年均去除率均高于32%.生物基质材料可显著提高养殖废水脱氮效果.
(2) 生物基质材料有利于系统内微生物增加, 添加麦秸材料的处理中AOA、AOB、nirK及16S rRNA基因丰度最高, hzsB基因与nirS基因丰度分别在稻草和麦秸中最多, 证实了基质池内有厌氧氨氧化过程.
(3) N循环相关功能基因丰度与水质环境因子之间存在显著相关, T及DO是影响微生物活性的重要因素.同时也受废水中NH4+-N、TN及NO3--N浓度影响.
致谢: 课题组与实验室的老师、同学和试验站的人员在采样和试验方面提供帮助, 在此一并致谢!
| [1] | 第二次全国污染源普查公报[J]. 环境保护, 2020, 48(18): 8-10. |
| [2] | Zhou S L, Zhang Y R, Huang T L, et al. Microbial aerobic denitrification dominates nitrogen losses from reservoir ecosystem in the spring of Zhoucun reservoir[J]. Science of the Total Environment, 2019, 651(Pt 1): 998-1010. |
| [3] | Hu B, Quan J N, Huang K, et al. Effects of C/N ratio and dissolved oxygen on aerobic denitrification process: a mathematical modeling study[J]. Chemosphere, 2021, 272. DOI:10.1016/j.chemosphere.2020.129521 |
| [4] |
张燕, 阎百兴, 刘秀奇, 等. 农田排水沟渠系统对磷面源污染的控制[J]. 土壤通报, 2012, 43(3): 745-750. Zhang Y, Yan B X, Liu X Q, et al. Measures for controlling phosphorus from agricultural non-point source pollutions in drainage ditch systems[J]. Chinese Journal of Soil Science, 2012, 43(3): 745-750. |
| [5] | Li X, Li Y Y, Lv D Q, et al. Nitrogen and phosphorus removal performance and bacterial communities in a multi-stage surface flow constructed wetland treating rural domestic sewage[J]. Science of the Total Environment, 2020, 709. DOI:10.1016/j.scitotenv.2019.136235 |
| [6] |
赵远哲, 董伟羊, 王海燕, 等. 气水比对A/O-BF处理低碳氮比农村生活污水脱氮的影响[J]. 环境科学学报, 2021, 41(2): 451-459. Zhao Y Z, Dong W Y, Wang H Y, et al. Effects of air-water ratio on the nitrogen removal of A/O-BF for the treatment of rural domestic sewage with low C/N ratio[J]. Acta Scientiae Circumstantiae, 2021, 41(2): 451-459. |
| [7] |
陈亮, 刘锋, 肖润林, 等. 人工湿地氮去除关键功能微生物生态学研究进展[J]. 生态学报, 2017, 37(18): 6265-6274. Chen L, Liu F, Xiao R L, et al. Research advances in microbial ecology for N-removal in constructed wetlands[J]. Acta Ecologica Sinica, 2017, 37(18): 6265-6274. |
| [8] |
王晓玲, 宋铁红, 殷宝勇, 等. 利用主要缺氧段ORP作为连续流单污泥污水脱氮除磷系统调控参数[J]. 环境科学, 2015, 36(7): 2617-2625. Wang X L, Song T H, Yin B Y, et al. ORP in the main anoxic stage as the control parameter for nitrogen and phosphorus removal in the single sludge system with a continuous flow[J]. Environmental Science, 2015, 36(7): 2617-2625. |
| [9] |
刘铭羽, 夏梦华, 李远航, 等. 3种基质材料对高浓度养殖废水处理效果及降解过程[J]. 环境科学, 2019, 40(8): 3650-3659. Liu M Y, Xia M H, Li Y H, et al. Treatment of highly concentrated swine wastewater and its degradation processes using three matrix Materials[J]. Environmental Science, 2019, 40(8): 3650-3659. |
| [10] |
王丽莎, 李希, 甘蕾, 等. 亚热带丘陵区湿地水生植物组合模式拦截氮磷的研究[J]. 生态环境学报, 2017, 26(9): 1577-1583. Wang L S, Li X, Gan L, et al. Study on the aquatic plant combination patterns for intercepting nitrogen and phosphorus in wetland of subtropical hilly region[J]. Ecology and Environmental Sciences, 2017, 26(9): 1577-1583. |
| [11] | 鲍士旦, 戚道光, 薛爱英, 等. 氮素营养对啤酒大麦产量和品质的影响[J]. 土壤, 1993, 25(1): 21-23. |
| [12] | Rotthauwe J H, Witzel K P, Liesack W. The ammonia monooxygenase structural gene amoA as a functional marker: molecular fine-scale analysis of natural ammonia-oxidizing populations[J]. Applied and Environmental Microbiology, 1997, 63(12): 4704-4712. DOI:10.1128/aem.63.12.4704-4712.1997 |
| [13] | Sahan E, Muyzer G. Diversity and spatio-temporal distribution of ammonia-oxidizing Archaea and Bacteria in sediments of the Westerschelde estuary[J]. FEMS Microbiology Ecology, 2008, 64(2): 175-186. DOI:10.1111/j.1574-6941.2008.00462.x |
| [14] |
黄薇, 刘兰英, 吴妙鸿, 等. 养殖废水处理系统中微生物菌群结构及动态变化[J]. 中国环境科学, 2019, 39(2): 839-848. Huang W, Liu L Y, Wu M H, et al. Microbial community structure and dynamics in swine wastewater treatment system[J]. China Environmental Science, 2019, 39(2): 839-848. DOI:10.3969/j.issn.1000-6923.2019.02.049 |
| [15] | 张树楠. 绿狐尾藻湿地对养殖废水氮去除效应及其机理研究[D]. 北京: 中国科学院大学, 2016. |
| [16] | Throbäck I N, Enwall K, Jarvis Å, et al. Reassessing PCR primers targeting nirS, nirK and nosZ genes for community surveys of denitrifying bacteria with DGGE[J]. FEMS Microbiology Ecology, 2004, 49(3): 401-417. DOI:10.1016/j.femsec.2004.04.011 |
| [17] | Dale O R, Tobias C R, Song B. Biogeographical distribution of diverse anaerobic ammonium oxidizing (anammox) bacteria in Cape Fear River Estuary[J]. Environmental Microbiology, 2009, 11(5): 1194-1207. DOI:10.1111/j.1462-2920.2008.01850.x |
| [18] | 杨杰. 亚热带红壤不同施肥处理对根际土壤微生物硝化及反硝化作用的影响研究[D]. 长沙: 湖南师范大学, 2019. |
| [19] |
闫冰, 夏嵩, 桂双林, 等. 厌氧氨氧化菌富集培养过程微生物群落结构及多样性[J]. 环境科学, 2020, 41(12): 5535-5543. Yan B, Xia S, Gui S L, et al. Microbial community structure and diversity during the enrichment of anaerobic ammonium oxidation bacteria[J]. Environmental Science, 2020, 41(12): 5535-5543. |
| [20] | 萨姆布鲁克J, 拉塞尔D W, 著. 黄培堂, 译. 分子克隆实验指南第三版下[M]. 北京: 科学出版社, 2002. |
| [21] | Wang J L, Chu L B. Biological nitrate removal from water and wastewater by solid-phase denitrification process[J]. Biotechnology Advances, 2016, 34(6): 1103-1112. DOI:10.1016/j.biotechadv.2016.07.001 |
| [22] |
周卿伟, 祝惠, 阎百兴, 等. 添加填料的人工湿地反硝化过程研究[J]. 湿地科学, 2017, 15(4): 588-594. Zhou Q W, Zhu H, Yan B X, et al. Denitrification process of constructed wetlands with adding filler[J]. Wetland Science, 2017, 15(4): 588-594. |
| [23] |
凌宇, 闫国凯, 王海燕, 等. 6种农业废弃物初期碳源及溶解性有机物释放机制[J]. 环境科学, 2021, 42(5): 2422-2431. Ling Y, Yan G K, Wang H Y, et al. Release mechanisms of carbon source and dissolved organic matter of Six agricultural wastes in the initial stage[J]. Environmental Science, 2021, 42(5): 2422-2431. |
| [24] | Li X, Zhang M M, Liu F, et al. The significance of Myriophyllum elatinoides for swine wastewater treatment: abundance and community structure of ammonia-oxidizing microorganisms in sediments[J]. PLoS One, 2015, 10(10). DOI:10.1371/journal.pone.0139778 |
| [25] |
赵林丽, 邵学新, 吴明, 等. 人工湿地不同基质和粒径对污水净化效果的比较[J]. 环境科学, 2018, 39(9): 4236-4241. Zhao L L, Shao X X, Wu M, et al. Effects of Different substrates and particle sizes on wastewater purification[J]. Environmental Science, 2018, 39(9): 4236-4241. |
| [26] | Li T, Yang X L, Song H L, et al. Alkali-treated cellulose carrier enhancing denitrification in membrane bioreactor[J]. International Biodeterioration & Biodegradation, 2019, 145. DOI:10.1016/j.ibiod.2019.104813 |
| [27] | Xu Z S, Dai X H, Chai X L. Effect of different carbon sources on denitrification performance, microbial community structure and denitrification genes[J]. Science of the Total Environment, 2018, 634: 195-204. DOI:10.1016/j.scitotenv.2018.03.348 |
| [28] | Wu L N, Yan Z B, Huang S, et al. Rapid start-up and stable maintenance of partial nitrification-anaerobic ammonium oxidation treatment of landfill leachate at low temperatures[J]. Environmental Research, 2020, 191. DOI:10.1016/j.envres.2020.110131 |
| [29] |
赵聪芳, 陈坤, 李希, 等. 利用秸秆材料处理养殖废水过程中氮的转化与氨排放特征[J]. 环境工程学报, 2020, 14(4): 993-1002. Zhao C F, Chen K, Li X, et al. Characteristics of nitrogen transformation and ammonia emission in swine wastewater treatment with straw materials[J]. Chinese Journal of Environmental Engineering, 2020, 14(4): 993-1002. |
| [30] |
黄娟, 王世和, 鄢璐, 等. 潜流型人工湿地硝化和反硝化作用强度研究[J]. 环境科学, 2007, 28(9): 1965-1969. Huang J, Wang S H, Yan L, et al. Intensity of nitrification and denitrification in subsurface-flow constructed wetlands[J]. Environmental Science, 2007, 28(9): 1965-1969. DOI:10.3321/j.issn:0250-3301.2007.09.012 |
| [31] | 刘永杰. 基于自养反硝化的沼气脱硫与废水脱氮同步处理技术研究[D]. 北京: 中国地质大学(北京), 2019. |
| [32] | 徐伟锋, 郑淑平, 孙力平, 等. 生物膜法同步硝化反硝化影响因素的分析[J]. 天津城市建设学院学报, 2003, 9(1): 4-7. DOI:10.3969/j.issn.1006-6853.2003.01.002 |
| [33] |
秦彦荣, 袁忠玲, 张明, 等. 部分亚硝化-厌氧氨氧化协同反硝化处理生活污水脱氮除碳[J]. 环境科学, 2021, 42(10): 4853-4863. Qin Y R, Yuan Z L, Zhang M, et al. Partial nitritation and anaerobic ammonia oxidation synergistic denitrification to remove nitrogen and carbon from domestic Sewage[J]. Environmental Science, 2021, 42(10): 4853-4863. |
| [34] | Liu D Z, Li J W, Li C W, et al. Poly (butylene succinate)/bamboo powder blends as solid-phase carbon source and biofilm carrier for denitrifying biofilters treating wastewater from recirculating aquaculture system[J]. Scientific Reports, 2018, 8(1). DOI:10.1038/s41598-018-21702-5 |
| [35] |
梁伟光, 黄廷林, 张海涵, 等. 李家河水库春季分层期nirS型反硝化菌群特征分析[J]. 环境科学, 2022, 43(1): 306-313. Liang W G, Huang T L, Zhang H H, et al. Characteristic analysis of nirS denitrifying bacterial community in Lijiahe reservoir during stratification[J]. Environmental Science, 2022, 43(1): 306-313. |
| [36] |
王欢, 裴伟征, 李旭东, 等. 低碳氮比猪场废水短程硝化反硝化-厌氧氨氧化脱氮[J]. 环境科学, 2009, 30(3): 815-821. Wang H, Pei W Z, Li X D, et al. Removing nitrogen from low-C/N-piggery-wastewater using shortcut nitrification/denitrification-ANAMMOX[J]. Environmental Science, 2009, 30(3): 815-821. DOI:10.3321/j.issn:0250-3301.2009.03.032 |
| [37] |
宋壮壮, 吕爽, 刘哲, 等. 厌氧氨氧化耦合反硝化工艺的启动及微生物群落变化特征[J]. 环境科学, 2019, 40(11): 5057-5065. Song Z Z, Lü S, Liu Z, et al. Start-up of simultaneous ANAMMOX and denitrification process and changes in microbial community characteristics[J]. Environmental Science, 2019, 40(11): 5057-5065. |