 2021, Vol. 42
2021, Vol. 42



2. 中国科学院南京地理与湖泊研究所湖泊与环境国家重点实验室, 南京 210008
2. State Key Laboratory of Lake Science and Environment, Nanjing Institute of Geography and Limnology, Chinese Academy of Sciences, Nanjing 210008, China
浮游植物是指在水体中浮游生活的光合自养生物, 是水体初级生产者[1], 其粒径结构影响着湖泊初级生产光合作用效率和食物网功能, 进而影响到整个湖泊生态系统的物质循环和能量传输, 是定量评价湖泊生态系统状况的重要生态指标[2~4].浮游植物吸收是水体总吸收的重要组成部分, 对于浮游植物光合作用和固碳、水质参数高精度遥感反演、水体初级生产力的估算以及全球碳循环等研究均具有重要的意义[5~15].浮游植物吸收与比吸收系数随着湖泊营养状况、季节更替和浮游植物群落结构而变化, 因此不同湖泊浮游植物吸收特性及其控制因素存在较大差异. Riddick等[16]在对巴拉顿湖的研究中发现, 浮游植物吸收系数与生物量存在明显的幂函数正相关, 蓝藻是影响浮游植物吸收特性的优势物种. Zhang等[17]通过对斜生栅藻(绿藻门)和铜绿微囊藻(蓝藻门)的实验室培养以及对太湖春夏浮游植物吸收特性的研究, 发现春季至夏季太湖浮游植物群落以大细胞绿藻为主演替为以小细胞蓝藻为主, 浮游植物比吸收系数显著降低.但是目前国内外此类研究主要集中在大洋水体[18~21], 针对中富营养水平湖泊的研究较少, 而湖泊水体浮游植物群落结构和粒径结构与大洋水体之间存在巨大差异, 大洋水体的浮游植物吸收特性不能简单的用于湖泊.此外, 多数相关研究主要以粒级指数(SI)来定量描述浮游植物粒径的变化[22~23], 或者进行藻类室内培养实验来独立研究细胞大小与吸收特性的关系[20, 24], 而直接基于湖泊浮游植物群落结构演替的研究较少.因此, 开展浮游植物群落结构及粒径对吸收和比吸收系数的影响研究, 对于深入理解内陆湖泊浮游植物吸收特性, 优化湖泊生物光学模型具有重要的科学意义.
本研究通过对天目湖2013年逐月(不包括2月和12月)浮游植物及其他相关因子的监测, 分析了浮游植物群落结构和吸收特性的季节变化特征, 阐明了浮游植物丰度、生物量以及粒径与吸收系数、比吸收系数之间的关系, 一方面可以为水色遥感数据反演浮游植物群落及粒径结构提供理论支持, 拓展水色遥感反演参数, 同时还可为天目湖生态系统长期监测及水质管理提供科学依据.
1 材料与方法 1.1 研究区与采样点分布天目湖位于江苏省溧阳市, 地处东经119.40°~119.45°, 北纬31.23°~31.32°, 水域面积11.6 km2, 集水面积152 km2, 平均水深7 m[25], 呈东北-西南走向, 南端有河流汇入, 北部修筑大坝, 为水库型深水湖泊, 气候属于亚热带季风气候, 雨热同季, 四季分明.自1962年建成以来, 流域内渔业、农业和旅游业先后兴起发展, 为地方带来经济效益的同时, 湖泊面源污染日益严重, 水质下降和藻类异常繁殖等富营养化问题严重[26, 27].近年来, 天目湖作为饮用水水源保护地, 流域的环境保护和治理工作加强, 地方采取封湖禁渔、生态补偿、区域限批、生态清淤和封山育林等措施, 湖泊水质得到改善, 现常年保持国家地表水Ⅱ类标准, 水体中浮游植物群落结构发生改变.
本研究在天目湖自北向南共布设7个采样点(图 1), 于2013年1月和3~11月每月下旬使用有机玻璃采水器采集表层水样, 于当日分别使用直径47 mm和25 mm GF/F膜对水样进行过滤, 具体过滤体积视水样颗粒物浓度而定.滤后留膜低温保存, 用于实验室后续测定叶绿素a浓度和浮游植物吸收系数.同时, 每个点保留1 L水样, 现场加入10 mL的鲁格试剂固定, 用于实验室浮游植物群落结构的鉴定.

|
图 1 天目湖采样点分布示意 Fig. 1 Distribution of sampling stations in Lake Tianmu |
水样静置沉淀48 h后, 使用虹吸管吸除表层清液, 然后定容到30 mL.浮游植物鉴定计数时, 应先将样品瓶摇匀, 迅速吸取0.1 mL水样于20 mm×20 mm计数框中, 利用40倍显微镜鉴定、计数, 所有样品重复观测计数两次, 取其均值为该样点的浮游植物细胞密度[28, 29].由于浮游植物的细胞密度接近于水的密度, 1 mm3相当于1 mg生物量, 故生物量的测定采用体积转化法[30, 31].
浮游植物的优势属依据优势度(Yi)来确定[32]:
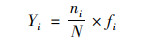
|
(1) |
式中, ni为第i个浮游植物属种的生物量, N为总生物量, fi为第i个属种的出现频次.当优势度(Yi)大于0.02时, 确定为优势属.
1.3 等效粒径计算由于浮游植物群落组成、生物量和细胞粒径差别非常大, 要非常准确地测定和计算其粒径大小非常困难, 为反映每一次采样中浮游植物细胞的平均粒径, 笔者创新性地提出等效粒径(equivalent sphere diameter, ESD)概念, 其计算公式如下:
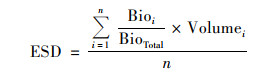
|
(2) |
式中, n是每个样本中浮游植物的种类数, Bioi是第i个物种的生物量, BioTotal是样本总生物量, Volumei是第i物种显微镜下观测到的粒径大小.
1.4 浮游植物吸收系数测定与比吸收系数计算悬浮颗粒物的吸收采用Yentsch[33]提出的定量滤膜技术(quantitative filter technique, QFT)测定, 使用同样浸泡过的空白滤膜作参比, 在岛津UV-2500紫外分光光度计下采用光透射-光反射(T-R)方法测定颗粒物在350~800 nm间每1 nm的吸光度.采用Tassan等[34]提出的公式进行放大因子校正:
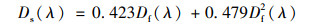
|
(3) |
式中, Ds(λ)为校正后的滤膜上悬浮颗粒物吸光度; Df(λ)为直接在仪器上测定的滤膜上悬浮颗粒物吸光度.
滤膜上悬浮颗粒物的光谱吸收系数ap(λ)按Cleveland提出的公式计算得出[35]:
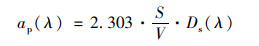
|
(4) |
式中, S为沉积在滤膜上的颗粒物有效面积, V为过滤水样的体积.
用0.1%的次氯酸钠溶液萃取掉滤膜上的色素, 使用同样的方法测定得非藻类颗粒物的吸收系数.由于悬浮颗粒物的吸收是非藻类颗粒物和浮游植物的简单线性相加, 因而浮游植物的光谱吸收系数aph(λ)就等于总悬浮颗粒物吸收系数ap(λ)减去非藻类颗粒物的吸收系数ad(λ), 即:
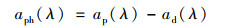
|
(5) |
浮游藻类的比吸收系数a*ph(λ)由以下公式计算得:
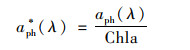
|
(6) |
式中, Chla为叶绿素a浓度, 采用分光光度法测定[36].使用90%的热酒精萃取, 过滤得到的清液在岛津UV-2500紫外分光光度计上测定665 nm和750 nm处吸光度, 并滴入1滴稀盐酸酸化, 计算得叶绿素a浓度.
1.5 统计分析方法使用Excel 2010和IBM SPSS 23进行统计分析, 包括平均值、标准差计算和线性拟合.使用回归分析法研究浮游植物丰度、生物量、等效粒径与吸收系数和比吸收系数之间的关系, 经F检验, 当P≤0.05时为显著.
2 结果与讨论 2.1 浮游植物群落结构季节变化特征采样期间共鉴定出浮游植物7门11纲15目29科42属, 其中蓝藻门3纲5目6科10属, 隐藻门1纲1目1科2属, 金藻门1纲1目1科1属, 甲藻门1纲1目2科2属, 裸藻门1纲1目1科2属, 硅藻门2纲2目7科8属, 绿藻门2纲4目11科17属.天目湖浮游植物丰度以蓝藻最多, 月平均丰度为922.93×104个·L-1, 占浮游植物月平均总数的73.29%;其次为硅藻、绿藻和隐藻, 金藻、裸藻和甲藻的月平均丰度较低, 均小于10×104个·L-1.浮游植物月平均生物量最大的是硅藻, 为1.04 mg·L-1, 占浮游植物月平均总数的35.87%, 其次为隐藻, 生物量为0.88 mg·L-1, 占浮游植物月平均总数的30.45%, 金藻的生物量最低, 为0.04 mg·L-1, 占浮游植物月平均总数的1.49%(表 1).
|
|
表 1 浮游植物丰度、生物量及其所占百分比 Table 1 Percentages of abundance and biomass of phytoplankton |
表 2显示了每月的主要优势属, 包括隐藻门中蓝隐藻属和隐藻属、金藻门中钟罩藻属、裸藻门中裸藻属、硅藻门中小环藻属、针杆藻属和曲壳藻属以及绿藻门中卵胞属、衣藻属和丝藻属. 3~6月以小环藻最具优势, 其优势度范围是0.365~0.528; 1月和7~10月隐藻最具优势, 其优势度范围是0.290~0.486.
|
|
表 2 天目湖浮游植物优势属及其优势度1) Table 2 Dominant genera and dominance index of phytoplankton in Lake Tianmu |
由图 2可知, 浮游植物生物量和丰度存在明显的季节变化.全年中浮游植物生物量最大值为4.49 mg·L-1, 出现在10月, 最小值仅为1.08 mg·L-1, 出现在1月.温度是影响天目湖浮游植物群落结构变化的主要原因, 当气温达到16℃时, 硅藻开始大量繁殖, 气温达26℃时, 硅藻的生物量达到最大, 而超过该温度时, 硅藻生物量开始下降[37].所以春季天目湖水温上升, 硅藻生物量随之增加, 夏季气温过高抑制硅藻生长, 其生物量急剧下降, 秋季气温降低, 硅藻生物量缓慢增加.但是小环藻的最佳生长温度更高, 其生物量总体上随温度的增加而增加.另外, 隐藻与水温呈显著正相关且生态位较宽, 对水体的适应性较强, 因此夏秋两季隐藻生物量较大, 冬春季节较小.

|
图 2 浮游植物生物量和丰度的逐月变化 Fig. 2 Monthly variations in phytoplankton biomass and abundance |
浮游植物丰度11月最大, 为3 422.86×104个·L-1, 1月浮游植物丰度最小, 为402.86×104个·L-1.秋季(9~11月)浮游植物丰度较多, 主要是因为蓝藻门中包括席藻和束丝藻的大量繁殖.浮游植物的生物量和丰度变化趋势并不完全相同, 这是由于生物量的大小除了与浮游植物数量相关外, 还与其个体细胞大小密切相关.硅藻的细胞体积较蓝藻大很多, 5月和6月硅藻丰度较多, 因而浮游植物生物量在春末夏初形成一个次高峰.
2.2 浮游植物等效粒径变化特征鉴定结果表明, 天目湖浮游植物等效粒径的物种组成多样, 分布跨度较大.按照分类标准, 微微型浮游植物包含20个属, 其中蓝藻门10属, 绿藻门9属, 硅藻门1属; 微型浮游植物共13个属, 其中隐藻门1属, 硅藻门6属, 绿藻门6属; 网采浮游植物共9属, 其中隐藻门1属, 金藻门1属, 甲藻门2属, 裸藻门2属, 硅藻门2属, 绿藻门1属.总的来说, 微微型浮游植物的比例占47.6%, 微型浮游植物的比例为31.0%, 是天目湖大部分浮游植物等效粒径的主要分类.
隐藻的等效粒径变化最大, 变化范围是28.43~1 024.78 μm, 平均等效粒径为531.73 μm; 其余按粒径大小排序依次是裸藻、甲藻、硅藻、金藻、绿藻和蓝藻, 其中硅藻、裸藻和甲藻的变化较小, 绿藻和金藻的季节差异不明显; 蓝藻的等效粒径变化幅度最小, 变化范围是0.05~0.62 μm, 平均等效粒径为0.28 μm[图 3(a)].图 3(b)展示了天目湖7个采样点浮游植物等效粒径及其平均值的季节变化, 结果表明所有采样点呈现出较统一的变化趋势, 3~8月, 天目湖浮游植物等效粒径波动上升, 8~11月开始波动下降; 其中夏季(7月)平均等效粒径最大, 为64.83 μm, 秋季(10月)平均等效粒径较小, 为50.92 μm, 其次为冬季(1月), 为46.09 μm, 春季(4月)平均等效粒径最小, 为29.54 μm, 存在明显的季节变化.

|
图 3 主要浮游植物粒径组成及等效粒径逐月变化 Fig. 3 Monthly variations in the size composition of the main phytoplankton and equivalent sphere diameter |
图 4为天目湖7个采样点浮游植物吸收系数曲线, 不同采样时间的浮游植物光谱吸收系数存在较大差异, 具有明显的季节变化.整体而言, 秋季(10月)浮游植物吸收系数最大, aph(440)和aph(675)的变化范围分别是0.49~1.03 m-1和0.22~0.53 m-1, 均值分别为(0.66±0.18)m-1和(0.33±0.10)m-1.其次为冬季(1月), aph(440)和aph(675)的变化范围分别是0.29~0.37 m-1和0.14~0.19 m-1, 均值分别为(0.33±0.03)m-1和(0.16±0.02)m-1; 春季(4月)和夏季(7月)浮游植物吸收系数偏小, 春季aph(440)和aph(675)的变化范围分别是0.11~0.52 m-1和0.05~0.27 m-1, 均值分别为(0.21±0.13)m-1和(0.11±0.07)m-1; 夏季aph(440)和aph(675)的变化范围分别是0.14~0.21 m-1和0.06~0.10 m-1, 均值分别为(0.17±0.02)m-1和(0.08±0.01)m-1.不同季节的浮游植物光谱吸收曲线具有相似特征, 所有采样点在440 nm和675 nm附近存在两个吸收峰, 440 nm的吸收峰是叶绿素a和其他辅助色素共同作用的结果, 675 nm的吸收峰是叶绿素a作用的结果[38].

|
图中曲线表示7个采样点的吸收系数 图 4 不同季节天目湖浮游植物吸收光谱 Fig. 4 Absorption spectra of phytoplankton during different seasons in Lake Tianmu |
天目湖浮游植物比吸收系数季节变化较小(图 5), 冬春季节浮游植物比吸收系数最大, 春季aph*(440)和aph*(675)的变化范围分别是0.04~0.10 m2·mg-1和0.02~0.05 m2·mg-1, 均值分别为(0.07±0.02)m2·mg-1和(0.04±0.01)m2·mg-1, 其对应的叶绿素a浓度范围是1.65~5.20 μg·L-1; 冬季aph*(440)和aph*(675)的变化范围分别是0.05~0.12 m2·mg-1和0.02~0.04 m2·mg-1, 均值分别为(0.07±0.02)m2·mg-1和(0.03±0.01)m2·mg-1, 其对应的叶绿素a浓度范围是4.14~7.63 μg·L-1.其次为秋季, aph*(440)和aph*(675)的变化范围分别是0.04~0.05 m2·mg-1和0.02~0.03 m2·mg-1, 均值分别为(0.05±0.004)m2·mg-1和(0.02±0.001)m2·mg-1, 其对应的叶绿素a浓度范围是9.00~20.07 μg·L-1.夏季浮游植物比吸收系数最小, aph*(440)和aph*(675)的变化范围分别是0.03~0.04 m2·mg-1和0.01~0.02 m2·mg-1, 均值分别为(0.03±0.004)m2·mg-1和(0.01±0.002)m2·mg-1, 其对应的叶绿素a浓度范围是3.93~6.37 μg·L-1.

|
图中曲线表示7个采样点的吸收系数 图 5 不同季节天目湖浮游植物比吸收光谱 Fig. 5 Specific absorption spectra of phytoplankton during different seasons in Lake Tianmu |
不同的研究学者发现, 无论是远海一类水体还是湖泊、河流等二类水体[39, 40], aph(440)和aph(675)与叶绿素a浓度存在很好地线性或非线性关系, 且675 nm处的相关性要好于440 nm.图 6为天目湖10个月70个采样点在440和675 nm处浮游植物吸收系数和比吸收系数与叶绿素a浓度的关系, aph(440)和aph(675)与叶绿素a浓度存在显著幂函数正相关关系, aph*(440)和aph*(675)与叶绿素a浓度相关性较弱, 但存在负相关关系.

|
图 6 浮游植物吸收系数、比吸收系数和叶绿素a浓度的关系 Fig. 6 Relationships between phytoplankton absorption, specific absorption coefficients, and chlorophyll a concentrations |
如上所述, 天目湖浮游植物群落结构存在明显的季节变化特征.图 7展示了浮游植物在440 nm和675 nm处的吸收系数随生物量和丰度的变化, 二者存在显著线性正相关.米氏理论可解释这两组参量间的关系, 多分散性的颗粒群吸收光谱可表示为[41]:
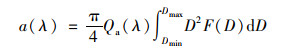
|
(7) |

|
图 7 浮游植物的生物量和丰度与吸收系数的关系 Fig. 7 Relationships between phytoplankton biomass, abundance, and absorption coefficients |
式中, Qa(λ)为平均吸收效率因子, D为颗粒直径, F(D)dD表示D到D+dD粒径范围内的粒子数.又因细胞密度即丰度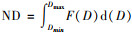
天目湖浮游植物吸收系数春、秋高而冬、夏低, 这与浮游植物生物量与丰度的季节变化特征一致.从冬末到春季随着天目湖湖水的升温, 硅藻开始大量繁殖, 丰度占比由6.7%升至38.2%, 总丰度比冬季增加了105.3%, 因此浮游植物吸收系数在春季形成一个次高峰.夏季的高温抑制了硅藻的生长, 其丰度占比降至5.0%, 而高温适宜蓝藻的生长, 其丰度由48.6%升至56.9%, 但总丰度减少了4.8%, 因而夏季与冬季的浮游植物吸收系数相对较低.秋季蓝藻迅速繁殖, 丰度占比升至80.6%, 总丰度增加了237.0%, 所以秋季的浮游植物吸收系数远大于其他季节.气温不同而引起的硅藻、蓝藻丰度的变化是造成天目湖浮游植物吸收系数季节变化的主要原因.
图 8表明440 nm和675 nm处浮游植物比吸收系数与等效粒径之间呈线性负相关, 随着等效粒径的增加, 浮游植物比吸收系数减小.研究表明, 大型浮游植物叶绿体体积较大, 自蔽效应(self-shading)较强, 对光能的利用率较低, 浮游植物包裹效应是造成比吸收系数空间和时间差异的主要原因, 以包裹因子来表示[43, 44], 公式为:
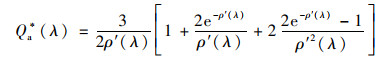
|
(8) |

|
图 8 浮游植物比吸收系数和等效粒径的关系 Fig. 8 Relationships between specific absorption coefficients and equivalent sphere diameter |
式中, ρ′(λ)=asol*cselld, asol*为悬浮状态下的比吸收系数; ccell为细胞内叶绿素a的浓度; d为细胞的直径.由以上公式可见, 细胞内叶绿素a浓度和浮游植物粒径是影响比吸收系数的共同因素, 这也解释了440 nm处由于辅助色素的影响, 包裹效应的体现不如675 nm.
天目湖浮游植物比吸收系数具有冬春较大, 夏秋偏小的特点, 与浮游植物等效粒径冬春较小, 夏秋较大的变化趋势正好相反.隐藻、小环藻和针杆藻等浮游植物全年占优势且等效粒径变化不大, 因而对比吸收系数的季节变化影响较小.春季硅藻大量繁殖, 曲壳藻成为优势属之一, 其等效粒径为46.32 μm, 此外蓝隐藻也是优势属之一, 其等效粒径仅为5.68 μm, 因而春季的浮游植物比吸收系数偏大.夏季, 卵胞藻和多甲藻成为优势属, 其等效粒径分别为49.50 μm和365.89 μm, 导致与春季相比, 夏季浮游植物比吸收系数较小.秋季, 天目湖优势属为丝藻和裸藻, 其等效粒径分别为16.08 μm和373.35 μm, 所以秋季浮游植物比吸收系数较夏季有所上升但仍旧偏小.冬季以适宜低温生长的蓝隐藻和钟罩藻为优势属, 其等效粒径分别为10.95 μm和74.47 μm, 粒径较小, 所以浮游植物比吸收系数受其影响继续增加直至与春季相当.浮游植物群落的季节演替, 尤其是优势属的组成变化, 其粒径大小是引起包裹效应的主要因素, 进而影响天目湖浮游植物比吸收系数的变化.
上述分析表明, 浮游植物群落组成的不同以及细胞大小的分布对浮游植物的吸收特性有着显著的影响.表 3整理了国内外典型湖泊的浮游植物吸收系数与比吸收系数, 明显地, 天目湖浮游植物吸收系数要远低于表中所列的6个湖泊, 而浮游植物比吸收系数较高; 与以往的研究一致, 浮游植物吸收系数与叶绿素浓度存在较好的相关性, 但是天目湖浮游植物比吸收系数与叶绿素a浓度相关性较弱, 这与Riddick等[16]在巴拉顿湖和Yoshimura等[47]在卡西高罗湖的研究结果相似.已有研究表明[48~50], 湖泊营养水平影响叶绿素a浓度和浮游植物比吸收系数进而改变二者之间的相关性, 但巴拉顿湖、卡西高罗湖、太湖和滇池同为重富营养湖泊, 而太湖、滇池其浮游植物比吸收系数与叶绿素a浓度却具有较好的相关性. Tomioka等[51]的研究发现光利用率是决定辅助色素和包裹效应的重要因素之一, Fujiki等[52]通过研究6种浮游植物在不同辐照度下比吸收和细胞大小的关系, 发现弱光条件下, 细胞大小对浮游植物比吸收的影响更稳健.由于光照条件的影响因素复杂多样, 如地理位置、天气状况、采样时间等, 所以浮游植物比吸收系数与叶绿素a浓度之间的关系在不同湖泊中表现出巨大的差异性.另外, Yentsch等[39]在查斯科姆湖的研究中发现, 浮游植物群落与生物量没有明显的季节变化, 与浮游植物吸收光谱的变化保持一致. Riddick等[16]的研究发现, 浮游植物吸收系数aph(675)与总生物量之间相关性较好(R2=0.882), 并进一步地发现了蓝藻和隐藻的生物量之和与浮游吸收系数同样呈指数正相关.与天目湖相比, 巴拉顿湖浮游植物生物量高且东西部湖区差距明显, 差值大于4.5×102 mg·L-1, 浮游植物吸收系数随生物量的增加急剧增大, 表明富营养湖泊浮游植物吸收系数受生物量影响更加明显. Zhang等[17]的研究结果表明, 浮游植物吸收系数[aph(675)]、比吸收系数a*ph(675)与细胞大小和叶绿素a浓度具有显著的相关性(R2=0.99, R2=0.99).浮游植物群落中个体藻类对吸收信号的贡献各不相同, 因而不同浮游植物其吸收特性与细胞大小、叶绿素a浓度之间的拟合效果不同, 这也是本研究中相关系数较低的原因.
|
|
表 3 6个湖泊的生物光学参数1) Table 3 Bio-optical parameters of six different lakes |
3 结论
本研究基于浮游植物群落结构和吸收系数的鉴定和测定, 探讨了群落结构演替对浮游植物比吸收系数的影响机制, 创新性地提出了表征全部浮游植物的平均粒径大小的等效粒径概念.本研究发现天目湖浮游植物生物量、丰度及优势属存在明显季节变化, 春季优势属以小环藻、隐藻、针杆藻和曲壳藻为主, 夏季优势属以隐藻、卵胞藻、多甲藻和衣藻为主, 秋季优势属以隐藻、针杆藻、丝藻和小环藻为主, 冬季优势属以隐藻、蓝隐藻、针杆藻和小环藻为主; 随着群落结构发生变化, 浮游植物等效粒径季节变化明显, 呈现夏季>秋季>冬季>春季; 吸收系数与叶绿素a浓度呈幂函数正相关关系, 但比吸收系数与叶绿素a浓度具有较弱的显著负相关, 表明随着藻类生物量增加, 色素包裹效应增加, 比吸收系数下降; 浮游植物比吸收系数与等效粒径存在显著线性负相关, 随等效粒径的增大而减小.隐藻、小环藻和针杆藻等全年优势属对比吸收系数的影响较小, 曲壳藻、卵胞藻、多甲藻、丝藻、裸藻和钟罩藻等优势属的季节更替引起的等效粒径的变化, 是影响浮游植物比吸收系数变化的重要因素.
致谢: 周万平帮助鉴定浮游植物群落, 邓建明帮助计算等效粒径, 在此一并表示感谢.
| [1] |
胡韧, 蓝于倩, 肖利娟, 等. 淡水浮游植物功能群的概念、划分方法和应用[J]. 湖泊科学, 2015, 27(1): 11-23. Hu R, Lan Y Q, Xiao L J, et al. The concepts, classification and application of freshwater phytoplankton functional groups[J]. Journal of Lake Sciences, 2015, 27(1): 11-23. |
| [2] | Guidi L, Stemmann L, Jackson G A, et al. Effects of phytoplankton community on production, size, and export of large aggregates: a world-ocean analysis[J]. Limnology and Oceanography, 2009, 54(6): 1951-1963. DOI:10.4319/lo.2009.54.6.1951 |
| [3] | Marañón E, Holligan P M, Barciela P, et al. Patterns of phytoplankton size structure and productivity in contrasting open-ocean environments[J]. Marine Ecology Progress Series, 2001, 216: 43-56. DOI:10.3354/meps216043 |
| [4] | Mouw C B, Yoder J A, Doney S C. Impact of phytoplankton community size on a linked global ocean optical and ecosystem model[J]. Journal of Marine Systems, 2012, 89(1): 61-75. DOI:10.1016/j.jmarsys.2011.08.002 |
| [5] | Bissett W P, Schofield O, Glenn S, et al. Resolving the impacts and feedback of ocean optics on upper ocean ecology[J]. Oceanography, 2001, 14(3): 30-53. DOI:10.5670/oceanog.2001.22 |
| [6] | Morel A, Antoine D. Heating rate within the upper ocean in relation to its bio-optical state[J]. Journal of Physical Oceanography, 1994, 24(7): 1652-1665. DOI:10.1175/1520-0485(1994)024<1652:HRWTUO>2.0.CO;2 |
| [7] | Nakamoto S, Kumar S P, Oberhuber J M, et al. Chlorophyll modulation of sea surface temperature in the Arabian Sea in a mixed-layer isopycnal general circulation model[J]. Geophysical Research Letters, 2000, 27(6): 747-750. DOI:10.1029/1999GL002371 |
| [8] | Bidigare R R, Prézelin B B, Smith R C. Bio-optical models and the problems of scaling[A]. In: Falkowski P G, Woodhead A D, Vivirito K (Eds.). Primary Productivity and Biogeochemical Cycles in the Sea[M]. Boston: Springer, 1992. 175-212. |
| [9] | Morel A, Antoine D, Babin M, et al. Measured and modeled primary production in the northeast Atlantic (EUMELI JGOFS program): The impact of natural variations in photosynthetic parameters on model predictive skill[J]. Deep Sea Research Part Ⅰ: Oceanographic Research Papers, 1996, 43(8): 1273-1304. DOI:10.1016/0967-0637(96)00059-3 |
| [10] | O'Reilly J E, Maritorena S, Mitchell B G, et al. Ocean color chlorophyll algorithms for SeaWiFS[J]. Journal of Geophysical Research, 1998, 103(C11): 24937-24953. DOI:10.1029/98JC02160 |
| [11] | Hirata T, Aiken J, Hardman-Mountford N, et al. An absorption model to determine phytoplankton size classes from satellite ocean colour[J]. Remote Sensing of Environment, 2008, 112(6): 3153-3159. DOI:10.1016/j.rse.2008.03.011 |
| [12] | Wu G F, Cui L J, He J J, et al. Comparison of MODIS-based models for retrieving suspended particulate matter concentrations in Poyang Lake, China[J]. International Journal of Applied Earth Observation and Geoinformation, 2013, 24: 63-67. DOI:10.1016/j.jag.2013.03.001 |
| [13] |
王桂芬, 曹文熙, 周雯, 等. 基于南海北部海区浮游植物吸收光谱斜率变化的粒级结构反演[J]. 热带海洋学报, 2010, 29(2): 25-32. Wang G F, Cao W X, Zhou W, et al. Retrieval of phytoplankton size structure based on the spectral slope of phytoplankton absorption in the northern South China Sea[J]. Journal of Tropical Oceanography, 2010, 29(2): 25-32. DOI:10.3969/j.issn.1009-5470.2010.02.005 |
| [14] | Li L H, Li L, Song K S, et al. An inversion model for deriving inherent optical properties of inland waters: establishment, validation and application[J]. Remote Sensing of Environment, 2013, 135: 150-166. DOI:10.1016/j.rse.2013.03.031 |
| [15] | Naik P, D'Sa E J, Gomes H D R, et al. Light absorption properties of southeastern Bering Sea waters: analysis, parameterization and implications for remote sensing[J]. Remote Sensing of Environment, 2013, 134: 120-134. DOI:10.1016/j.rse.2013.03.004 |
| [16] | Riddick C A L, Hunter P D, Tyler A N, et al. Spatial variability of absorption coefficients over a biogeochemical gradient in a large and optically complex shallow lake[J]. Journal of Geophysical Research: Oceans, 2015, 120(10): 7040-7066. DOI:10.1002/2015JC011202 |
| [17] | Zhang Y L, Yin Y, Wang M Z, et al. Effect of phytoplankton community composition and cell size on absorption properties in eutrophic shallow lakes: field and experimental evidence[J]. Optics Express, 2012, 20(11): 11882-11898. DOI:10.1364/OE.20.011882 |
| [18] | Lohrenz S E, Weidemann A D, Tuel M. Phytoplankton spectral absorption as influenced by community size structure and pigment composition[J]. Journal of Plankton Research, 2003, 25(1): 35-61. DOI:10.1093/plankt/25.1.35 |
| [19] | Ciotti A M, Lewis M R, Cullen J J. Assessment of the relationships between dominant cell size in natural phytoplankton communities and the spectral shape of the absorption coefficient[J]. Limnology and Oceanography, 2002, 47(2): 404-417. DOI:10.4319/lo.2002.47.2.0404 |
| [20] | Stuart V, Sathyendranath S, Platt T, et al. Pigments and species composition of natural phytoplankton populations: effect on the absorption spectra[J]. Journal of Plankton Research, 1998, 20(2): 187-217. DOI:10.1093/plankt/20.2.187 |
| [21] | Bricaud A, Claustre H, Ras J, et al. Natural variability of phytoplanktonic absorption in oceanic waters: influence of the size structure of algal populations[J]. Journal of Geophysical Research, 2004, 109(C11). DOI:10.1029/2004JC002419 |
| [22] |
王桂芬, 曹文熙, 许大志, 等. 南海北部藻类粒级结构及色素成分对浮游植物吸收系数的影响[J]. 海洋学报, 2007, 29(1): 38-48. Wang G F, Cao W X, Xu D Z, et al. Effects of size structure and pigment composition of algal population on phytoplankton absorption coefficients in the South China Sea[J]. Acta Oceanologica Sinica, 2007, 29(1): 38-48. DOI:10.3321/j.issn:0253-4193.2007.01.006 |
| [23] |
刘洋洋, 沈芳, 李秀珍. 长江口邻近海域赤潮水体浮游植物光吸收特性分析[J]. 环境科学, 2015, 36(6): 2019-2027. Liu Y Y, Shen F, Li X Z. Phytoplankton Light absorption properties during the blooms in adjacent waters of the Changjiang estuary[J]. Environmental Science, 2015, 36(6): 2019-2027. |
| [24] | Agustí S. Light environment within dense algal populations: cell size influences on self-shading[J]. Journal of Plankton Research, 1991, 13(4): 863-871. DOI:10.1093/plankt/13.4.863 |
| [25] |
孙祥, 朱广伟, 杨文斌, 等. 天目湖沙河水库浮游植物群落结构的时空异质性[J]. 环境科学, 2017, 38(10): 4160-4168. Sun X, Zhu G W, Yang W B, et al. Spatio-temporal variations in phytoplankton community in Shahe Reservoir, Tianmuhu, China[J]. Environmental Science, 2017, 38(10): 4160-4168. |
| [26] |
伊平, 张磊, 包健. 天目湖流域农业面源污染控制研究[J]. 污染防治技术, 2008, 21(6): 38-40, 43. Yi P, Zhang L, Bao J. Study on controlling of the agricultural non-point source pollution in the Tianmu Lake Valley[J]. Pollution Control Technology, 2008, 21(6): 38-40, 43. |
| [27] |
朱广伟, 陈伟民, 李恒鹏, 等. 天目湖沙河水库水质对流域开发与保护的响应[J]. 湖泊科学, 2013, 25(6): 809-817. Zhu G W, Chen W M, Li H P, et al. Response of water quality to the catchment development and protection in Tianmuhu Reservoir, China[J]. Journal of Lake Sciences, 2013, 25(6): 809-817. |
| [28] |
胡鸿钧, 魏印心. 中国淡水藻类——系统、分类及生态[M]. 北京: 科学出版社, 2006. Hu H J, Wei Y X. The freshwater algae of china-systematics, taxonomy and ecology[M]. Beijing: Science Press, 2006. |
| [29] |
胡鸿钧. 水华蓝藻生物学[M]. 北京: 科学出版社, 2011. Hu H J. The biology of water-blooms blue-green algae[M]. Beijing: Science Press, 2011. |
| [30] | 孙军, 刘东艳, 钱树本. 浮游植物生物量研究Ⅰ.浮游植物生物量细胞体积转化法[J]. 海洋学报, 1999, 21(2): 75-85. DOI:10.3321/j.issn:0253-4193.1999.02.010 |
| [31] | 章宗涉, 黄祥飞. 淡水浮游生物研究方法[M]. 北京: 科学出版社, 1991. |
| [32] | Austin M P. Relationships among functional properties of Californian grassland[J]. Nature, 1968, 217(5134). DOI:10.1038/216168b0 |
| [33] | Yentsch C S. Measurement of visible light absorption by particulate matter in the ocean[J]. Limnology and Oceanography, 1962, 7(2): 207-217. DOI:10.4319/lo.1962.7.2.0207 |
| [34] | Tassan S, Ferrari G M. An alternative approach to absorption measurements of aquatic particles retained on filters[J]. Limnology and Oceanography, 1995, 40(8): 1358-1368. DOI:10.4319/lo.1995.40.8.1358 |
| [35] | Cleveland J S, Weidemann A D. Quantifying absorption by aquatic particles: A multiple scattering correction for glass-fiber filters[J]. Limnology and Oceanography, 1993, 38(6): 1321-1327. DOI:10.4319/lo.1993.38.6.1321 |
| [36] | Parsons T R, Maita Y, Lalli C M. A manual of chemical and biological methods for seawater analysis[M]. Oxford: Pergamon Press, 1984. |
| [37] |
朱广伟, 金颖薇, 任杰, 等. 太湖流域水库型水源地硅藻水华发生特征及对策分析[J]. 湖泊科学, 2016, 28(1): 9-21. Zhu G W, Jin Y W, Ren J, et al. Characteristics of diatom blooms in a reservoir-water supply area and the countermeasures in Taihu Basin, China[J]. Journal of Lake Sciences, 2016, 28(1): 9-21. |
| [38] | 周百成, 郑舜琴, 曾呈奎. 几种绿藻、褐藻和红藻的吸收光谱的比较研究[J]. 植物学报, 1974, 16(2): 146-155. |
| [39] | Yentsch C S, Phinney D A. A bridge between ocean optics and microbial ecology[J]. Limnology and Oceanography, 1989, 34(8): 1694-1705. DOI:10.4319/lo.1989.34.8.1694 |
| [40] |
张运林, 秦伯强, 杨龙元. 太湖梅梁湾水体悬浮颗粒物和CDOM的吸收特性[J]. 生态学报, 2006, 26(12): 3969-3979. Zhang Y L, Qin B Q, Yang L Y. Spectral absorption coefficients of particulate matter and chromophoric dissolved organic matter in Meiliang Bay of Lake Taihu[J]. Acta Ecologica Sinica, 2006, 26(12): 3969-3979. DOI:10.3321/j.issn:1000-0933.2006.12.007 |
| [41] |
周雯, 曹文熙, 李彩, 等. 由吸收系数和粒度分布计算浮游植物的散射光谱特征[J]. 光学学报, 2008, 28(8): 1429-1433. Zhou W, Cao W X, Li C, et al. Spectral scattering property of phytoplankton calculated by absorption coefficient and size distribution[J]. Acta Optica Sinica, 2008, 28(8): 1429-1433. DOI:10.3321/j.issn:0253-2239.2008.08.001 |
| [42] |
周雯, 孙兆华, 曹文熙, 等. 浮游植物的吸收-衰减特性及其与粒径间的关系[J]. 光谱学与光谱分析, 2012, 32(12): 3347-3352. Zhou W, Sun Z H, Cao W X, et al. Variations in the optical absorption and attenuation properties of cultured phytoplankton and their relationships with cell size[J]. Spectroscopy and Spectral Analysis, 2012, 32(12): 3347-3352. DOI:10.3964/j.issn.1000-0593(2012)12-3347-06 |
| [43] | Duyens L N M. The flattering of the absorption spectrum of suspensions, as compared to that of solutions[J]. Biochimica et Biophysica Acta, 1956, 19: 1-12. DOI:10.1016/0006-3002(56)90380-8 |
| [44] | Morel A, Bricaud A. Theoretical results concerning light absorption in a discrete medium, and application to specific absorption of phytoplankton[J]. Deep Sea Research Part A. Oceanographic Research Papers, 1981, 28(11): 1375-1393. DOI:10.1016/0198-0149(81)90039-X |
| [45] | Shi K, Li Y M, Li L, et al. Absorption characteristics of optically complex inland waters: Implications for water optical classification[J]. Journal of Geophysical Research: Biogeosciences, 2013, 118(2): 860-874. DOI:10.1002/jgrg.20071 |
| [46] | Pérez G L, Llames M E, Lagomarsino L, et al. Seasonal variability of optical properties in a highly turbid lake (Laguna Chascomús, Argentina)[J]. Photochemistry and Photobiology, 2011, 87(3): 659-670. DOI:10.1111/j.1751-1097.2011.00907.x |
| [47] | Yoshimura K, Zaitsu N, Sekimura Y, et al. Parameterization of chlorophyll a-specific absorption coefficients and effects of their variations in a highly eutrophic lake: a case study at Lake Kasumigaura, Japan[J]. Hydrobiologia, 2012, 691(1): 157-159. DOI:10.1007/s10750-012-1066-4 |
| [48] | Agawin N S R, Duarte C M, Agusti S. Nutrient and temperature control of the contribution of picoplankton to phytoplankton biomass and production[J]. Limnology and Oceanography, 2000, 45(3): 591-600. DOI:10.4319/lo.2000.45.3.0591 |
| [49] | Raven J A. Physiological consequences of extremely small size for autotrophic organisms in the sea[J]. Canadian Bulletin of Fisheries and Aquatic Sciences, 1986, 214: 1-70. |
| [50] | Cao W X, Yang Y Z, Xu X Q, et al. Regional patterns of particulate spectral absorption in the Pearl River estuary[J]. Science Bulletin, 2003, 48(21): 2344-2351. DOI:10.1360/03wd0151 |
| [51] | Tomioka N, Imai A, Komatsu K. Effect of light availability on Microcystis aeruginosa blooms in shallow hypereutrophic Lake Kasumigaura[J]. Journal of Plankton Research, 2011, 33(8): 1263-1273. DOI:10.1093/plankt/fbr020 |
| [52] | Fujiki T, Taguchi S. Variability in chlorophyll a specific absorption coefficient in marine phytoplankton as a function of cell size and irradiance[J]. Journal of Plankton Research, 2002, 24(9): 859-874. DOI:10.1093/plankt/24.9.859 |