 2017, Vol. 38
2017, Vol. 38



消落带是陆生环境和水生环境之间的过渡带,具有过滤和缓冲的作用[1, 2],在维护水生和陆生环境的生态健康与生态安全方面具有极其特殊的地位[3].三峡大坝修建后,形成面积为350 km2的消落带.三峡消落带主要由城镇消落带、农村消落带、库中岛屿消落带组成[4].其中城市消落带的面积约52 km2,约占整个库区消落带的15%[5].三峡消落带是一个高度脆弱的生态系统[6].为改善三峡消落带的生态环境,生态修复措施在消落带得以推广和实施[7~9].大量研究表明,三峡消落带植被恢复重建后土壤生态系统结构和养分条件得到改善,生态系统的生物多样性和稳定性增加[10],尤其是土壤微生物群落的明显变化.
土壤微生物是土壤生态系统中的分解者,主要由细菌、真菌和放线菌组成,对土壤中物质循环、能量流通都有着极其重要的作用[11~13].放线菌主要是革兰氏阳性细菌,有高GC含量,其在土壤生态系统中广泛存在[14].放线菌已知有亚纲:Rubrobacteridae、Coriobacteridae、Acidimicrobidae、Actinobacteridae和Sphaerobacteridae[15, 16].放线菌作为土壤中的分解者,可分解一些难分解化合物,例如纤维素[17]、半纤维素、几丁质[18]等;同时放线菌可产生大量种类繁多的胞外酶和抗生素[19, 20],可抑制土壤中致病菌的传播[21, 22],从而保持土壤的生态健康.有研究表明土壤放线菌群落受到多种因素影响,例如土壤pH[23]、海拔[24]、土壤养分[25]等因素,更有发现土壤污染程度和放线菌数量呈正相关[26].土壤放线菌作为保持土壤健康的重要因子,在三峡城乡消落带的植被修复后土壤中还未得到研究.因此,本文以三峡库区典型城乡消落带 (重庆江北嘴大剧院消落带和忠县汝溪河流域) 为研究区域,选取人工生态修复后的典型的植被类型——林地和草地,通过分析土壤理化性质和放线菌多样性以及其之间的关系,以期从放线菌群落角度为三峡库区消落带植被修复后的生态保护提供科学理论依据.
1 材料与方法 1.1 研究区概况本研究的城市消落带位于重庆江北区嘉陵江与长江干流交汇处 (106°34′47″~106°34′48″E,29°34′22″~29°34′25″ N).消落带土壤基质以沉积泥沙为主,有少量建筑废弃物和砾石,多为淤积泥土或沙土生境[27].为进行消落带生态恢复,于2009年在江北嘴大剧院消落带区域构建示范基地,种植扁穗牛鞭草 (Hemarthria compressa (L. f.) R. Br.)、狗牙根 (Cynodon dactylon (L.) Pers)、池杉 (Taxodium ascendens) 和枫杨 (Pterocarya stenoptera C. DC) 等植被,同时人为活动十分频繁.
本研究的农村消落带位于三峡库区重庆忠县石宝镇的汝溪河流域消落带.忠县石宝镇位于重庆市中部 (108°08′03″~108°08′21″E,30°24′16″~30°24′56″N),距离忠县县城32 km,位于忠县、石柱、万州三县 (区) 交界处.其消落带原生植被破坏殆尽,现有植被以草本为主;土壤主要为中性紫色土,水土流失、崩塌现象较严重[28].为进行三峡库区消落带的生态植被恢复与重建,本课题组于2012年3月在该地区消落带165~175 m海拔区间按1 m×1 m的株行距栽种柳树 (Salix matsudana)、落羽杉 (Taxodium distichum) 和池杉护岸林;其中按0.3 m×0.5 m栽种扁穗牛鞭草和狗牙根.苗木栽种的岸坡样地土壤均为紫色土,平均坡度为26°,岸坡上接受光照辐射强度大致相同,土壤预热条件基本一致.
1.2 样地设置和样品采集在坡度相似、坡向一致、接受光照辐射强度大致相同、土壤预热条件基本一致的消落带区域内,选定具有典型代表性的样地开展实验研究. 2015年6月,通过实地踏查,选定城乡消落带区域165~175 m海拔区间的林地和草地.根据试验地实际情况、植被类型和受干扰情况,在城市和乡村消落带分别设置林地和草地重复样地各3个,共12个样地,所有样地尽量保持在同一海拔高度.其中城市消落带林地以落羽杉、枫杨为优势种,总盖度约为90%;乡村消落带林地以落羽杉、柳树和池杉为优势种,总盖度约为93%;城乡消落带草地的优势物种均为狗牙根、扁穗牛鞭草.
在每个样地内随机设3条样带,样带中按S形5点取样.采样点从总体上控制整个样地,避免在堆肥料地和田埂、沟边等特殊地形部位采样.在样点清理地被物后,采取0~20 cm、20~40 cm和40~60 cm的土壤,5点同层土壤等质量混合,利用四分法保留土壤样品.其中1 kg土壤室温风干,用于土壤理化性质分析. 20 g混合土样装入无菌的自封袋,低温运输,-80℃保存,用于土壤放线菌多样性分析.
1.3 土壤理化性质测定土壤含水量 (water content, WC) 采用烘干法测定,土壤密度 (soil density, SD) 采用比重瓶法测定,土壤总孔隙度 (soil total porosity, SP) 采用计算法测定.土壤pH值测定采用IQ150土壤原位pH计 (IQ Scientific Instruments, Inc., San Diego, CA, USA) 原位测定;土壤有机质 (organic matter, OM) 采用重铬酸钾外加热法测定;土壤碱解氮 (available nitrogen,AN) 含量采用碱解-扩散法测定;速效钾 (available potassium, AK) 含量采用电感耦合等离子体发射光谱法 (ICP-OES, Thermo Fisher iCAP 6300, UK) 测定;速效磷 (available phosphorus, AP) 含量采用钼锑抗比色法 (UV-2550, Shimadzu., CHN) 测定,以上测试方法均参照文献[29].
1.4 土壤放线菌多样性测定本实验土壤放线菌多样性测定方法为限制性末端长度多态性 (T-RFLP)[30].具体测定步骤如下.
1.4.1 DNA提取、扩增和酶切用Ultra CleanTM Soil Isolation DNA Kit (MOBIO,USA) 提取土壤总DNA,用1%的琼脂糖凝胶电泳检测所提DNA片段,并用UV spectrophotometer (SMA1000, Meriton) 测定DNA浓度.放线菌的扩增引物为243F (5′-GGATGAGCCCGCGGCCTA-3′)/1378R (5′-CGGTGTGTACAAGGCCCGGGAACG-3′)[31, 32],且正向引物5′端用6-FAM进行荧光标记.本实验的所用引物由深圳华大基因公司合成并标记.放线菌50 μL的PCR反应体系组成如下:25 μL 2×Taq Master Mix,2 μL DNA模板,243F/1378R (10 μmol·L-1) 各0.5 μL,ddH2O补水至50 μL. PCR反应条件如下:95℃,5 min. 30个循环为: 94℃,30 s;55℃,30 s;72℃,1 min;最后72℃保温10 min.每个样品重复2管.扩增产物经1%琼脂糖凝胶电泳检测,按照PCR产物纯化试剂盒 (TIANGEN,China) 说明书进行PCR产物纯化,-20℃保存备用.
纯化产物用限制性内切酶HhaⅠ消化,反应体系: 10×Buffer 2 μL,HhaⅠ (10 U·μL-1) 1 μL,DNA 200 ng,ddH2O补足至30 μL.将酶切样品置于37℃水浴中温育3 h,酶切完毕后65℃水浴10 min终止反应.每个样品按上述同样条件各重复3次.酶切产物送至上海Invitrogen生命技术公司 (Life Technologies) 进行基因扫描,得到T-RFLP图谱.
1.4.2 T-RFLP分析在限制性片段 (terminal restriction fragment, T-RF) 的选择中,删除引物峰小于50 bp和荧光值小于50 RFU的T-RFs,并去除OTU (operational taxonomic unit) 丰度 < 1%的T-RFs,T-RFs片段大小±1 bp被认为是同一个OTU[33, 34].
1.5 数据处理以T-RFLP图谱中T-RF为一个OTU,以该T-RF的相对峰面积作为对应的OTU的丰度并计算以下多样性指数.
物种丰度 (S)=图谱中T-RF的总数,即OTU的总数.
Shannon-Wiener指数 (H):
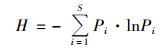
|
Simpson指数 (D):
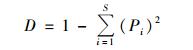
|
Pielou指数 (E):
E=H/lnS
其中,Pi为某个峰的峰面积占总峰面积的比例.
采用双因素方差分析 (Two-way ANOVA) 分析区域因子和土层以及其交互作用对放线菌多样性的影响,并用Duncan检验法检验每个指标在城乡消落带林地和草地之间的差异显著性 (α=0.05).利用冗余分析 (redundancy analysis,RDA) 和蒙特卡罗置换检验 (Monte-Carlo permutation test) 分析环境变量对土壤放线菌群落变异解释度的显著性 (α=0.05).用Origin 8.5和Cano Draw for Windows 4.5软件制图.
2 结果与分析 2.1 城乡消落带林地和草地土壤放线菌多样性差异采用双因素方差分析分析区域因子、土层及其交互作用对城区消落带土壤中的放线菌丰度、香农指数、辛普森指数和均匀度指数的影响. 表 1显示,土壤放线菌多样性在不同土层之间无显著差异,城乡区域和土层的交互作用对放线菌多样性对香农指数产生显著性影响 (P < 0.05),但对物种丰度、辛普森指数和均匀度指数未产生显著性影响.但城乡消落带土壤放线菌的物种丰度、香农指数和均匀度指数之间有显著差异 (P < 0.05).
|
|
表 1 土壤放线菌多样性的双因素方差分析1) Table 1 Two-way analysis of variance (ANOVA) for soil actinobacterial diversity |
图 1显示,城市消落带土壤放线菌物种丰度、香浓指数和辛普森指数均呈现出高于乡村消落带的总体趋势;城乡消落带中,相同土层的林地放线菌的丰度和多样性均高于或等同于相应草地.在消落带0~20 cm土壤中,城市草地土壤放线菌物种丰度、香农指数和辛普森指数显著高于乡村草地;同时城市林地土壤放线菌的香农指数显著高于乡村林地 (P < 0.05).在消落带20~40 cm土壤中,城市林地和草地土壤放线菌物种丰度、香农指数和辛普森指数与农村林地和草地之间分别无显著差异;但均匀度指数显著低于乡村草地 (P < 0.05).在40~60 cm土壤中,城市林地和草地与农村林地和草地的土壤放线菌物种丰度、香农指数和辛普森指数分别均无显著差异;但城市林地土壤放线菌均匀度指数显著低于乡村林地 (P < 0.05).

|
同层土壤中,不同字母表示在P < 0.05水平上差异显著,下同 图 1 城乡消落带土壤放线菌多样性的差异 Fig. 1 Difference in soil actinobacterial biodiversity of urban and rural hydro-fluctuation zone |
在T-RFLP图谱中,每一个T-RF至少代表一种放线菌,因此T-RF的多样性代表放线菌物种的多样性.如图 2所示,柱状堆叠图分别表示不同样点土壤中放线菌优势物种的相对丰度.从T-RFLP图谱可知,经过HhaI酶切后共得到34个优势末端限制性片段 (图 2). 图 2中110、150和225 bp在土壤中普遍存在,但其丰度在不同用地方式土壤中表现不同.在不同土地利用方式下,优势片段的种类与丰度均不同,且随土壤深度呈现不同的变化趋势.在乡村消落带中,145 bp大量存在乡村消落带林地的各层土壤中,但仅少量存在于乡村消落带的草地中.在城市消落带林地和草地存在的优势片段593 bp在乡村消落带中并未发现.

|
RGH代表乡村草地0~20 cm土壤;RGM代表乡村草地20~40 cm土壤;RGD代表乡村草地40~60 cm土壤;RWH代表乡村林地0~20 cm土壤;RWM代表乡村林地20~40 cm土壤;RWD代表乡村林地40~60 cm土壤;UGH代表城市草地0~20 cm土壤;UGM代表城市草地20~40 cm土壤;UGD代表城市草地40~60 cm土壤;UWH代表城市林地0~20 cm土壤;UEM代表城市林地20~40 cm土壤;UWD代表城市林地40~60 cm土壤 图 2 土壤放线菌群落优势T-RF的相对丰度 Fig. 2 Relative abundance of predominant T-RFs of soil actinobacterial communities |
表 2显示,城乡消落带土壤的各理化指标有显著差异 (P < 0.05).其中乡村消落带中,林地含水量均高于其草地;但在城市消落带中,在0~20 cm土壤中林地的含水量高于草地,其他土层则相反,为草地的含水量高于其林地.在城乡消落带0~20 cm土壤中草地土壤密度均高于林地,而其它层的土壤密度则相互之间无显著差异;与此同时,林地的孔隙度在各土层均未出现显著低于草地的现象.
|
|
表 2 城乡消落带林地和草地土壤理化性质 (平均值±标准误) Table 2 Soil physical-chemical properties in the urban and rural hydro-fluctuation zone (Means±SE) |
在0~20 cm土壤中,乡村消落带林地AK和AN含量显著高于草地,但pH则相反,而OM和AP的含量无显著差异;城市消落带林地土壤OM含量显著高于草地,但pH、AK、AN和AP无显著差异.在20~40 cm土壤中,乡村消落带林地AK和AN含量显著高于草地,但OM则相反,pH和AP的含量没有显著性差异;在城市消落带草地土壤OM和AN的含量显著高于林地,pH、AK和AP无显著差异.在40~60 cm土壤中,乡村消落带草OM和AN的含量显著高于林地,pH、AK和AP无显著差异;城市消落带林地pH和OM的含量显著高于草地,AN、AP和AK无显著差异.在不同土层中,乡村消落带林地和草地AP的含量显著高于相应城市消落带林地和草地.
2.2.3 土壤理化性质与放线菌多样性之间的关系图 3反映城乡消落带土壤放线菌多样性与环境因子之间的关系.从中可知,在城乡消落带中,林地和草地之间的放线菌群落有明显差异.对乡村消落带分析显示[图 3(a)],第1序轴解释所有信息的54.70%,第2序轴解释所有信息的25.90%,前两轴可解释80.60%.蒙特卡罗检验显示WC (F=11.50,P=0.002)、AP (F=9.69,P=0.004)、AN (F=5.02,P=0.038) 和OM (F=4.96,P=0.024) 对乡村消落带放线菌群落多样性的影响达到显著水平 (P < 0.05).对城市消落带RDA分析显示[图 3(b)],第1序轴解释所有信息的67.70%,第2序轴解释所有信息的4.20%,前两轴可解释71.90%.蒙特卡罗检验显示WC (F=10.05,P=0.008)、AN (F=8.39,P=0.008) 对城市消落带放线菌群落多样性的影响达到显著水平 (P < 0.05).

|
图 3 环境因素对土壤放线菌群落结构特征影响的冗余分析 Fig. 3 Redundancy analysis of soil actinobacterial biodiversity structure and environmental factors |
三峡消落带的生态健康对整个三峡流域至关重要,其中生态系统组分的完整性是其保证生态健康的基础.消落带植被恢复后,林地、草地和混合种植区域内土壤养分得到改善[10, 35],物种多样性以及土壤酶活性增加[36].放线菌作为生态系统中的一部分,可分解纤维素等难分解的化合物,溶解致病菌细胞壁中几丁质或抑制其产生,因而具有生物防治病菌土传的能力[37, 38],所以放线菌多样性是保持土壤生态系统的健康和质量的重要因素[39].
3.1 城乡消落带林地和草地土壤放线菌多样性差异本研究发现城乡消落带放线菌多样性有差异,城市消落带土壤放线菌物种丰度、香浓指数和辛普森指数均呈现出高于乡村消落带的总体趋势;且城乡消落带中,相同土层的林地放线菌的丰度和多样性均高于或等同于相应草地,与Shirokikh等[40]研究结果相似;但是土层对放线菌多样性的影响没有达到显著性影响,这与前人研究中发现的土壤微生物群落多样性是随着土壤深度增加而减少[41]不同,这主要是由不同微生物类群对外界环境变化的敏感度不同所致.有研究表明放线菌对于环境变化的敏感程度低于细菌和真菌[26],在本研究的土壤深度所导致的微环境变化不足以引起放线菌多样性的显著变化,使得土层对土壤放线菌群落的影响未达到显著.
本研究中城市消落带土壤放线菌多样性呈现高于乡村消落带的趋势,这有可能人为干扰会造成土壤微生物多样性的波动[42].有研究发现城市土壤微生物数量要高于农村土壤[43],城市化会显著影响土壤放线菌群落及多样性[44].在城乡消落带中,区域因子对表层0~20 cm土壤放线菌多样性的影响比20~40 cm和40~60 cm更为显著.这可能是由于在城市消落带中有着频繁的人为活动和外来污染物的输入,而这种人为干扰和污染会直接作用于土壤表层,使得表层土壤放线菌群落有较为明显的波动.
3.2 土壤放线菌优势菌群的丰度土壤微生物的种群的数量、种类、分布等特征均会受到外界环境因子 (如土壤水热状况、肥力条件以及植被类型) 的影响,因此导致所得到的T-RFLP图谱有所差别[11, 45, 46].本文中样品来自典型的城市消落带和乡村消落带,相对丰度较高的优势种110、150和225 bp在所有样品中都大量存在,其优势种组成的差异主要来自其他相对丰度较小的优势种,如450 bp、593 bp等;这种差异可能是地理隔离或者放线菌所生存的微环境差异所造成的[47].放线菌作为微生物中的一部分,也受大气氮沉降、土壤pH、土壤类型等的影响[48, 49].植被对于土壤微生物的影响也是不可忽略的,根系特定的分泌物会限制微生物的生长,从而导致微生物菌群的特异化[50].消落带林地和草地的地表植被相差甚远,有可能根系分泌物的差异导致土壤放线菌群落之间的差异.同时土壤放线菌主要以植被凋落物为食,分解木质素、纤维素和半纤维素等,地表凋落物也会对放线菌群落有选择作用,从而导致优势菌群之间的差异.
3.3 消落带土壤放线菌和环境因子之间的关系在城乡消落带土壤理化性质 (表 2) 中,城乡消落带土壤的各理化指标有显著差异.在表层土壤中,城乡消落带林地土壤有机质、速效钾、速效磷的含量均高于草地.这可能是由于是林地冠层对于雨水的拦截使得地表径流变小,土壤表层有机质以及矿质营养得以保留,流失较少[51].同时,与草地相比,林地中产生的较多的林木根系和枯落物,林地有机物的输入量高于草地,也会有助于土壤营养元素含量的提高.
本研究发现城乡消落带放线菌多样性的影响因子有差异.乡村消落带中含水量、速效磷、速效氮、有机质的含量显著影响放线菌多样性,而城市消落带中含水量、速效氮的含量显著影响放线菌多样性,该结果和Singh等[52]的研究结果相似.在陆地生态系统中,土壤含水量显著的影响放线菌多样性.本研究中,消落带土壤面临着干旱-水淹-干旱交替的特殊水文变化影响,土壤放线菌群落受到双重水分胁迫.土壤含水量可通过影响水压和土壤养分含量影响放线菌群落[53].有研究表明,土壤含水量可能成为放线菌物种的选择因子.在土壤含水量相对较低的旱季,耐旱的物种的孢子可以萌发,完成生活史从而被保留[54].同时研究表明,放线菌中的Streptomyces、Micromonospora、Actinomadura、Saccharopolyspora和Microbispora 会在不同的水分条件下发生不同程度的隔离[55];其中Streptomyces 更倾向于在高水分土壤中生长,在低水分的土壤中其生长会受到限制[56].同时土壤养分是土壤微生物生长活动的能源,调节土壤微生物群落结构和活动的重要因子[57~59].当土壤中养分达到均衡时,放线菌达到最适生长[60].土壤中有机质和速效氮主要来源于地上植被的凋落物和植被根系分泌物[61],其含量直接影响土壤放线菌群落.放线菌作为异养微生物,主要以有机碳作为其能源.城市消落带由于水流相对较缓,每年泥沙沉积时,植被残体被滞留在土壤中[62],特别是草本植株根系的附着从而被保留于泥沙中.但乡村消落带水流冲刷会剥离表层土壤和残体,使得土壤表层的有机质流失,土壤的碳氮储备量下降,土壤中微生物的生长和次生代谢都较低[63, 64],在多重影响下最终表现为城市消落带土壤放线菌多样性高于乡村消落带,且影响因素不同.
4 结论城乡消落带土壤放线菌多样性和优势物种有显著性差异,呈现城市消落带土壤放线菌多样性高于乡村消落带,且林地土壤放线菌多样性高于草地的趋势;放线菌多样性在土层之间无显著差异.经冗余分析和蒙特卡罗置换检验表明,土壤含水量、速效氮含量显著影响城市消落带放线菌多样性,土壤含水量、速效氮、速效钾和有机质的含量显著影响乡村消落带土壤放线菌多样性.由此说明城市化会影响土壤放线菌群落,增加消落带土壤放线菌多样性,这可能主要是由于城市消落带受到更多的人为干扰和土壤养分含量增加所致.建议减少城市消落带的人为干扰,以保护消落带生态环境.
| [1] | Kenwick R A, Shammin M R, Sullivan W C. Preferences for riparian buffers[J]. Landscape and Urban Planning, 2009, 91(2): 88–96. DOI: 10.1016/j.landurbplan.2008.12.005 |
| [2] | 程瑞梅, 王晓荣, 肖文发, 等. 消落带研究进展[J]. 林业科学, 2010, 46(4): 111–119. Cheng R M, Wang X R, Xiao W F, et al. Advances in studies on water-level-fluctuation zone[J]. Scientia Silvae Sinicae, 2010, 46(4): 111–119. DOI: 10.11707/j.1001-7488.20100417 |
| [3] | 张建春, 彭补拙. 河岸带研究及其退化生态系统的恢复与重建[J]. 生态学报, 2003, 23(1): 56–63. Zhang C J, Peng B Z. Study on riparian zone and the restoration and rebuilding of its degraded ecosystem[J]. Acta Ecologica Sinica, 2003, 23(1): 56–63. |
| [4] | 范小华, 谢德体, 魏朝富. 三峡水库消落区生态环境保护与利用对策研究[J]. 水土保持学报, 2006, 20(2): 165–169. Fan X H, Xie D T, Wei C F. Study on countermeasures for ecological environmental protection and utilization of riparian zone of Three Gorges Reservoir[J]. Journal of Soil and Water Conservation, 2006, 20(2): 165–169. |
| [5] | 杜立刚, 方芳, 郭劲松, 等. 三峡库区城市消落带生态规划与保护探讨[J]. 长江流域资源与环境, 2012, 21(6): 726–731. Du L G, Fang F, Guo J S, et al. Ecological planning and protection of urban water-level-fluctuation zone in the Three Gorges Reservoir[J]. Resources and Environment in the Yangtze Basin, 2012, 21(6): 726–731. |
| [6] | 马骏, 李昌晓, 魏虹, 等. 三峡库区生态脆弱性评价[J]. 生态学报, 2015, 35(21): 7117–7129. Ma J, Li C X, Wei H, et al. Dynamic evaluation of ecological vulnerability in the Three Gorges Reservoir Region in Chongqing Municipality, China[J]. Acta Ecologica Sinica, 2015, 35(21): 7117–7129. |
| [7] | Webb A A, Erskine W D. A practical scientific approach to riparian vegetation rehabilitation in Australia[J]. Journal of Environmental Management, 2003, 68(4): 329–341. DOI: 10.1016/S0301-4797(03)00071-9 |
| [8] | 王勇, 刘义飞, 刘松柏, 等. 三峡库区消涨带植被重建[J]. 植物学通报, 2005, 22(5): 513–522. Wang Y, Liu Y F, Liu S B, et al. Vegetation reconstruction in the water-level-fluctuation zone of the Three Gorges Reservoir[J]. Chinese Bulletin of Botany, 2005, 22(5): 513–522. |
| [9] | Ye C, Cheng X L, Zhang Q F. Recovery approach affects soil quality in the water level fluctuation zone of the Three Gorges Reservoir, China:implications for revegetation[J]. Environmental Science and Pollution Research, 2014, 21(3): 2018–2031. DOI: 10.1007/s11356-013-2128-5 |
| [10] | 杨予静, 李昌晓, 马朋. 三峡水库城区消落带人工草本植被土壤养分含量研究[J]. 草业学报, 2015, 24(4): 1–11. Yang Y J, Li C X, Ma P, et al. Nutrient content of soils under artificial grass vegetation in the urban hydro-fluctuation belt of the Three Gorges Reservoir region[J]. Acta Prataculturae Sinica, 2015, 24(4): 1–11. DOI: 10.11686/cyxb20150401 |
| [11] | Unger I M, Goyne K W, Kremer R J, et al. Microbial community diversity in agroforestry and grass vegetative filter strips[J]. Agroforestry Systems, 2013, 87(2): 395–402. DOI: 10.1007/s10457-012-9559-8 |
| [12] | Lienhard P, Terrat S, Mathieu O, et al. Soil microbial diversity and C turnover modified by tillage and cropping in Laos tropical grassland[J]. Environmental Chemistry Letters, 2013, 11(4): 391–398. DOI: 10.1007/s10311-013-0420-8 |
| [13] | Bardgett R D, Freeman C, Ostle N J. Microbial contributions to climate change through carbon cycle feedbacks[J]. The ISME Journal, 2008, 2(8): 805–814. DOI: 10.1038/ismej.2008.58 |
| [14] | Lechevalier H. Actinomycetes and their products:a look at the future[J]. World Journal of Microbiology and Biotechnology, 1992, 8(1): 72–73. |
| [15] | Stackebrandt E, Rainey F A, Ward-Rainey N L. Proposal for a new hierarchic classification system, Actinobacteria classis nov.[J]. Internatoinal Journal of Systematic Bacteriology, 1997, 47(2): 479–491. DOI: 10.1099/00207713-47-2-479 |
| [16] | Gao B L, Gupta R S. Conserved indels in protein sequences that are characteristic of the phylum Actinobacteria[J]. International Journal of Systematic and Evolutionary Microbiology, 2005, 55(6): 2401–2412. DOI: 10.1099/ijs.0.63785-0 |
| [17] | Pankratov T A, Dedysh S N, Zavarzin G A. The leading role of actinobacteria in aerobic cellulose degradation in Sphagnum peat bogs[J]. Doklady Biological Sciences, 2006, 410(1): 428–430. DOI: 10.1134/S0012496606050243 |
| [18] | Krsek M, Wellington E M H. Assessment of chitin decomposer diversity within an upland grassland[J]. Antonie van Leeuwenhoek, 2001, 79(3): 261–267. |
| [19] | Gomes R C, Semêdo L T A S, Soares R M A, et al. Chitinolytic activity of actinomycetes from a cerrado soil and their potential in biocontrol[J]. Letters in Applied Microbiology, 2000, 30(2): 146–150. DOI: 10.1046/j.1472-765x.2000.00687.x |
| [20] | Challis G L, Hopwood D A. Synergy and contingency as driving forces for the evolution of multiple secondary metabolite production by Streptomyces species[J]. Proceedings of the National Academy of Sciences of the United States of America, 2003, 100(S2): 14555–14561. |
| [21] | Hyakumachi M, Takahashi H, Matsubara Y, et al. Recent studies on biological control of plant diseases in Japan[J]. Journal of General Plant Pathology, 2014, 80(4): 287–302. DOI: 10.1007/s10327-014-0524-4 |
| [22] | Degtyareva E A, Vinogradova K A, Aleksandrova A V, et al. Soil actinomycetes as potential biofungicides[J]. Moscow University Soil Science Bulletin, 2009, 64(2): 73–77. DOI: 10.3103/S0147687409020045 |
| [23] | Sanyika T W, Stafford W, Cowan D A. The soil and plant determinants of community structures of the dominant actinobacteria in Marion Island terrestrial habitats, Sub-Antarctica[J]. Polar Biology, 2012, 35(8): 1129–1141. DOI: 10.1007/s00300-012-1160-0 |
| [24] | 曾清苹, 何丙辉, 毛巧芝, 等. 缙云山马尾松林和柑橘林土壤微生物PLFA沿海拔梯度的变化[J]. 环境科学, 2015, 36(12): 4667–4675. Zeng Q P, He B H, Mao Q Z, et al. Effects of different altitudes on soil microbial PLFA and enzyme activity in two kinds of forests[J]. Environmental Science, 2015, 36(12): 4667–4675. |
| [25] | Wan X H, Huang Z Q, He Z M, et al. Soil C:N ratio is the major determinant of soil microbial community structure in subtropical coniferous and broadleaf forest plantations[J]. Plant and Soil, 2015, 387(1-2): 103–116. DOI: 10.1007/s11104-014-2277-4 |
| [26] | Větrovsky T, Baldrian P. An in-depth analysis of actinobacterial communities shows their high diversity in grassland soils along a gradient of mixed heavy metal contamination[J]. Biology and Fertility of Soils, 2015, 51(7): 827–837. DOI: 10.1007/s00374-015-1029-9 |
| [27] | 冯义龙, 先旭东, 黄芳. 重庆江北嘴大剧院段消落带植被生态恢复示范区建设[J]. 南方农业, 2011, 5(10): 1–4. |
| [28] | 杨予静, 李昌晓, 丽娜·热玛赞. 基于PSR框架模型的三峡库区忠县汝溪河流域生态系统健康评价[J]. 长江流域资源与环境, 2013, 22(S1): 66–74. Yang Y J, Li C X, Remazan L N. Assessment of ecosystem health in the Ruxi river basin of Zhongxian county in the Three Gorges Reservoir area based on Pressure-State-Response (PSR model)[J]. Resources and Environment in the Yangtze Basin, 2013, 22(S1): 66–74. |
| [29] | 鲍士旦. 土壤农化分析[M]. (第三版). 北京: 中国农业出版社, 2000: 30-109. |
| [30] | Kitts C L. Terminal restriction fragment patterns:a tool for comparing microbial communities and assessing community dynamics[J]. Current Issues in Intestinal Microbiology, 2001, 2(1): 17–25. |
| [31] | Heuer H, Krsek M, Baker P, et al. Analysis of actinomycete communities by specific amplification of genes encoding 16S rRNA and gel-electrophoretic separation in denaturing gradients[J]. Applied and Environmental Microbiology, 1997, 63(8): 3233–3241. |
| [32] | Monciardini P, Sosio M, Cavaletti L, et al. New PCR primers for the selective amplification of 16S rDNA from different groups of actinomycetes[J]. FEMS Microbiology Ecology, 2002, 42(3): 419–429. |
| [33] | Székely A J, Sipos R, Berta B, et al. DGGE and T-RFLP analysis of bacterial succession during mushroom compost production and sequence-aided T-RFLP profile of mature compost[J]. Microbial Ecology, 2009, 57(3): 522–533. DOI: 10.1007/s00248-008-9424-5 |
| [34] | Schütte U M E, Abdo Z, Bent S J, et al. Advances in the use of terminal restriction fragment length polymorphism (T-RFLP) analysis of 16S rRNA genes to characterize microbial communities[J]. Applied Microbiology and Biotechnology, 2008, 80(3): 365–380. DOI: 10.1007/s00253-008-1565-4 |
| [35] | 马朋, 李昌晓, 任庆水, 等. 三峡水库消落带柳树 (Salix matsudana) 种植初期对土壤碳、氮、磷含量的影响[J]. 西南大学学报 (自然科学版), 2014, 36(8): 36–43. Ma P, Li C X, Ren Q S, et al. Soil C, N and P contents after short-term revegetation by Salix matsudana in the water-level-fluctuation zone of the Three Gorges Reservoir[J]. Journal of Southwest University (Natural Science Edition), 2014, 36(8): 36–43. |
| [36] | 马朋, 李昌晓, 雷明, 等. 三峡库区岸坡消落带草地、弃耕地和耕地土壤微生物及酶活性特征[J]. 生态学报, 2014, 34(4): 1010–1020. Ma P, Li C X, Lei M, et al. Responses of soil microorganisms and soil enzyme activities to different land use patterns in the water-level-fluctuating zone of the Three Gorges Reservoir region[J]. Acta Ecologica Sinica, 2014, 34(4): 1010–1020. |
| [37] | El-Tarabily K A, Sivasithamparam K. Non-streptomycete actinomycetes as biocontrol agents of soil-borne fungal plant pathogens and as plant growth promoters[J]. Soil Biology and Biochemistry, 2006, 38(7): 1505–1520. DOI: 10.1016/j.soilbio.2005.12.017 |
| [38] | 宋影, 辜夕容, 严海元, 等. 中亚热带马尾松林凋落物分解过程中的微生物与酶活性动态[J]. 环境科学, 2015, 35(3): 1151–1158. Song Y, Gu X R, Yan H Y, et al. Dynamics of microbes and enzyme activities during litter decomposition of Pinus massoniana forest in mid-subtropical area[J]. Environmental Science, 2015, 35(3): 1151–1158. |
| [39] | Shen W S, Lin X G, Gao N, et al. Land use intensification affects soil microbial populations, functional diversity and related suppressiveness of cucumber Fusarium wilt in China's Yangtze River Delta[J]. Plant and Soil, 2008, 306(1-2): 117–127. DOI: 10.1007/s11104-007-9472-5 |
| [40] | Shirokikh I G, Solov'eva E S, Ashikhmina T Y. Actinomycetes in garden soils of the city of Kirov[J]. Eurasian Soil Science, 2013, 46(5): 565–571. DOI: 10.1134/S106422931305013X |
| [41] | Barness G, Zaragoza S R, Shmueli I, et al. Vertical distribution of a soil microbial community as affected by plant ecophysiological adaptation in a desert system[J]. Microbial Ecology, 2009, 57(1): 36–49. DOI: 10.1007/s00248-008-9396-5 |
| [42] | Zhang X, Gu Q, Long X E, et al. Anthropogenic activities drive the microbial community and its function in urban river sediment[J]. Journal of Soils and Sediments, 2016, 16(2): 716–725. DOI: 10.1007/s11368-015-1246-8 |
| [43] | 赵哲霞, 郭恢财. 城市化对城乡梯度森林土壤微生物数量变化的影响 (英文)[J]. 农业科学与技术, 2010, 11(3): 118–122. Zhao Z X, Guo H C. Effects of urbanization on the quantity changes of microbes in urban-to-rural gradient forest soil[J]. Agricultural Science and Technology, 2010, 11(3): 118–122. |
| [44] | Hill P, Krištůfek V, Dijkhuizen L, et al. Land use intensity controls actinobacterial community structure[J]. Microbial Ecology, 2011, 61(2): 286–302. DOI: 10.1007/s00248-010-9752-0 |
| [45] | Xu H J, Wang X H, Li H, et al. Biochar impacts soil microbial community composition and nitrogen cycling in an acidic soil planted with rape[J]. Environmental Science & Technology, 2014, 48(16): 9391–9399. |
| [46] | Spurgeon D J, Keith A M, Schmidt O, et al. Land-use and land-management change:relationships with earthworm and fungi communities and soil structural properties[J]. BMC Ecology, 2013, 13: 46. DOI: 10.1186/1472-6785-13-46 |
| [47] | Zhang B L, Wu X K, Zhang W, et al. Diversity and succession of actinobacteria in the forelands of the Tianshan Glacier, China[J]. Geomicrobiology Journal, 2016, 33(8): 716–723. DOI: 10.1080/01490451.2015.1085468 |
| [48] | Zenova G M, Manucharova N A, Zvyagintsev D G. Extremophilic and extremotolerant actinomycetes in different soil types[J]. Eurasian Soil Science, 2011, 44(4): 417–436. DOI: 10.1134/S1064229311040132 |
| [49] | Eisenlord S D, Zak D R. Simulated atmospheric nitrogen deposition alters actinobacterial community composition in forest soils[J]. Soil Science Society of America Journal, 2010, 74(4): 1157–1166. DOI: 10.2136/sssaj2009.0240 |
| [50] | Berendsen R L, Pieterse C M J, Bakker P A H M. The rhizosphere microbiome and plant health[J]. Trends in Plant Science, 2012, 17(8): 478–486. DOI: 10.1016/j.tplants.2012.04.001 |
| [51] | 刘斌, 罗全华, 常文哲, 等. 不同林草植被覆盖度的水土保持效益及适宜植被覆盖度[J]. 中国水土保持科学, 2008, 6(6): 68–73. Liu B, Luo Q H, Chang W Z, et al. Relationship between percentage of vegetative cover and soil erosion[J]. Science of Soil and Water Conservation, 2008, 6(6): 68–73. |
| [52] | Singh A K, Singh M, Dubey S K. Changes in Actinomycetes community structure under the influence of Bt transgenic brinjal crop in a tropical agroecosystem[J]. BMC Microbiology, 2013, 13: 122. DOI: 10.1186/1471-2180-13-122 |
| [53] | Zvyagintsev D G, Zenova G M, Sudnitsyn I I, et al. Development of actinomycetes in brown semidesert soil under low water pressure[J]. Eurasian Soil Science, 2012, 45(7): 717–723. DOI: 10.1134/S1064229312030155 |
| [54] | Zenova G M, Gryadunova A A, Doroshenko E A, et al. Influence of moisture on the vital activity of actinomycetes in a cultivated low-moor peat soil[J]. Eurasian Soil Science, 2007, 40(5): 560–564. DOI: 10.1134/S1064229307050110 |
| [55] | Zvyagintsev D G, Zenova G M, Doroshenko E A, et al. Actinomycete growth in conditions of low moisture[J]. Biology Bulletin, 2007, 34(3): 242–247. DOI: 10.1134/S1062359007030053 |
| [56] | Goodfellow M, Williams S T. Ecology of actinomycetes[J]. Annual Review of Microbiology, 1983, 37: 189–216. DOI: 10.1146/annurev.mi.37.100183.001201 |
| [57] | Brant J B, Sulzman E W, Myrold D D. Microbial community utilization of added carbon substrates in response to long-term carbon input manipulation[J]. Soil Biology and Biochemistry, 2006, 38(8): 2219–2232. DOI: 10.1016/j.soilbio.2006.01.022 |
| [58] | Brant J B, Myrold D D, Sulzman E W. Root controls on soil microbial community structure in forest soils[J]. Oecologia, 2006, 148(4): 650–659. DOI: 10.1007/s00442-006-0402-7 |
| [59] | Hgberg M N, Hgberg P, Myrold D D. Is microbial community composition in boreal forest soils determined by pH, C-to-N ratio, the trees, or all three?[J]. Oecologia, 2006, 150(4): 590–601. DOI: 10.1007/s00442-006-0562-5 |
| [60] | Zhong W H, Gu T, Wang W, et al. The effects of mineral fertilizer and organic manure on soil microbial community and diversity[J]. Plant and Soil, 2010, 326(1-2): 511–522. DOI: 10.1007/s11104-009-9988-y |
| [61] | 熊顺贵. 基础土壤学[M]. 北京: 中国农业大学出版社, 2001: 118-230. |
| [62] | 沈冰洁, 祝贞科, 袁红朝, 等. 不同种植方式对亚热带红壤微生物多样性的影响[J]. 环境科学, 2015, 36(10): 3839–3844. Shen B J, Zhu Z K, Yuan H Z, et al. Effects of different plantation type on the abundance and diversity of soil microbes in subtropical red soils[J]. Environmental Science, 2015, 36(10): 3839–3844. |
| [63] | Degens B P, Schipper L A, Sparling G P, et al. Decreases in organic C reserves in soils can reduce the catabolic diversity of soil microbial communities[J]. Soil Biology and Biochemistry, 2000, 32(2): 189–196. DOI: 10.1016/S0038-0717(99)00141-8 |
| [64] | Zhong W H, Cai Z C. Long-term effects of inorganic fertilizers on microbial biomass and community functional diversity in a paddy soil derived from quaternary red clay[J]. Applied Soil Ecology, 2007, 36(2-3): 84–91. DOI: 10.1016/j.apsoil.2006.12.001 |