 2016, Vol. 37
2016, Vol. 37


2. 河海大学水文水资源学院, 南京 210098;
3. 新疆大学资源与环境科学学院, 乌鲁木齐 830000
2. College of Hydrology and Water Resources, Hohai University, Nanjing 210098, China;
3. College of Resource and Environmental Sciences, Xinjiang University, Urumqi 830000, China
应用农田排水沟渠拦截、 处理农业非点源污染的研究是人工湿地水处理技术的推广和延伸. 1993年,Meuleman 等[1]研究指出,天然沟渠能够吸收水体中氮污染物,可以利用天然沟渠改善受污染的水质; Kröger等[2, 3]研究发现,一条长约400 m,宽约7 m农田排水沟对可溶性无机氮的截留率可以达到57%; Strock等[4]认为农田排水沟渠在生态学和物理学上的功能与线性湿地的功能相似; Needelman等[5]将沟渠系统概括为具有河流和湿地特征的独特工程化生态系统. 1999年,晏维金等[6]通过两条排水沟对比实验,结果表明沟渠湿地系统具有双重性质:一方面,沟渠湿地系统能有效地截留氮磷污染物; 另一方面,沟渠湿地系统本身又是一个潜在的污染源; 姜翠玲等[7]在南京地区开展的沟渠湿地对农业非点源污染物净化能力研究,也得出相似结果; 2005年,杨林章在研究太湖流域农田非点源污染问题时,结合当地实际情况提出了“生态拦截型沟渠系统”概念[8],以此为基础近年又提出了农业非点源污染“4R”控制体系[9]; 李强坤等[10]以青铜峡灌区为例,应用水动力学基本方程和污染物迁移转化方程对非点源溶质在排水沟的运移进行了模拟,并进行了典型排水沟水污特征解析[11],近年,张树楠等[12]研究了沟渠底泥属性对非点源溶质的吸附特性,李如忠等[13]研究进一步细化,分析了排水沟渠溶质传输的暂态存储影响及参数灵敏性,此外,也有学者将沟渠湿地对农业非点源污染物降解的研究进一步扩展到沟塘湿地进行实验研究,彭世彰等[14]以现场监测实验为基础,开展了灌区沟塘湿地对稻田排水中氮磷的原位削减效果及机制研究[15, 16]; 更有学者将沟渠化及景观型灌排系统布设与农业非点源污染防治结合在一起进行探讨[17, 18]等.
综合上述国内外关于农田排水沟渠系统净化农业非点源污染物的相关研究,可得出以下结论:①农田排水沟渠水-底泥-植物组成的复合生态结构对非点源溶质具有明显的净化效果,但也潜伏着诱发内源污染甚至进一步扩散的威胁; ②应用农田排水沟渠“拦截”农业非点源污染尚未形成完善的理论体系,当前研究主要集中于局部或单条、 多条沟渠的对比实验、 水生植物备选及生态结构形式探讨等层面,关于农田排水沟渠水-底泥-植物系统内各介质间非点源溶质的迁移转化机制尚不清楚,农田排水沟渠非点源污染溶质净化与内源污染形成机制也有待进一步研究[19, 20],这也是阻碍沟渠湿地进一步调控运用的焦点和技术难点. 基于当前研究现状,本文以模拟排水沟渠静态实验为基础,以典型非点源溶质氮素为例,分析了农田排水沟渠水-底泥-植物不同介质内氮的迁移动态过程,以期为进一步揭示农田沟渠水-底泥-植物各介质间非点源溶质迁移转化机制奠定基础.
1 材料与方法 1.1 实验布置模拟沟渠实验布置在黄河水利科学研究院节水与农业生态实验基地(河南省新乡市). 制作4个相同规格的PVC实验模拟箱(长×宽×高:2 m×0.65 m×0.65 m,见图 1),其中1号模拟箱作为空白进行对比,仅加入水样; 2号模拟箱在箱底均匀铺设20 cm底泥,实验底泥取自外业实验地——河南省人民胜利渠灌区典型农田排水沟渠,并经自然阴干、 粉碎、 去除杂质; 3号模拟箱在铺设20 cm底泥的基础上,移栽已经培育好的芦苇,初始种植密度为10株 ·m-2(根据成活情况,后期依然按此密度移栽),4号模拟箱为培育多年的水体-底泥-芦苇实验箱.

|
单位: cm 图 1 实验箱体及取样点布设 Fig. 1 Layout of experiment boxes and sampling point |
实验中采用水体中添加尿素[CO(NH2)2]和磷酸氢胺[(NH4)2HPO4]混合溶液调控水体中氮、 磷浓度. 实验开始时,4月1日4个模拟箱统一加水0.39m3,并在1、 2、 3号模拟箱中添加CO(NH2)2和(NH4)2HPO4混合液,调控水体中总氮浓度为15 mg ·L-1,4号模拟箱没有加入任何药剂,按加水后现有氮素浓度进入实验.
之后随着实验进行,模拟箱中水位和水体中氮素浓度降低,分别在6月13日、 7月29日和10月9日向1、 2、 3号模拟箱中分别加入CO(NH2)2和(NH4)2HPO4混合液,并依据3、 4号模拟箱中水量情况控制,向4个模拟箱加入同样水量,为保证实验一致性同时调控1、 2、 3号模拟箱中总氮浓度为8mg ·L-1,4号模拟箱依然不加入任何药剂.
为使模拟实验尽可能接近自然状态,实验过程中4个模拟箱统一放置在露天环境,气温、 蒸发及雨雪等各实验条件完全一致. 考虑到NH4+离子易转化为气态的NH3,挥发进入大气,因此氮的挥发在实验中统一不计入分析.
1.3 样本提取和分析水样每10 d取样一次,分别在每个模拟箱的前、 中、 后这3个断面分别设置取样点[详见图 1(a)],采集水体深度1/2处水样,每次取样500 mL,分析指标包括总氮、 氨氮、 硝氮等,文中取3个水样的平均值进行分析.
底泥样品每月采集一次,采用PSC-600A型活塞式柱状沉积物采样器,在每个模拟箱的前、 中、 后这3个断面分别设置取样点[详见图 1(b)],分别提取底泥表层、 中层、 底层(即0-5、 5-10、 10-15 cm深)的底泥样品,样品取出后放入风干袋自然风干,然后磨碎、 过100目筛进行分析,分析指标包括总氮、 氨氮、 硝氮等,同样文中取前、 中、 后这3个断面样品的平均值进行分析.
植物样品在芦苇长到一定株高后,6月开始取样,每月取样一次,分别提取同株芦苇的根、 茎、 叶,并按根、 茎、 叶分类放入风干袋自然风干、 粉碎后进行分析,分析指标为总氮.
各项样品提取按照文献[21]中相关规定进行必要的前处理后进行实验室化验分析,主要分析仪器为Smartchem 140全自动化学分析仪.
2 结果与讨论 2.1 水体中氮素变化特征水体中氮素浓度除降雨、 温度等外界自然条件影响外,还受底泥吸附及植物吸收和根系活动层分解影响. 选取2014年4月3日至2014年12月25日时段实验资料(下同),点绘4个模拟箱中水体氨氮、 硝酸盐氮浓度分别见图 2、 图 3.

|
图 2 模拟实验箱水体中氨氮浓度变化 Fig. 2 Variation of nitrogen concentration in water in simulated experiment boxes |

|
图 3 模拟实验箱水体中硝酸盐氮浓度变化 Fig. 3 Variation of nitrate concentration in water in simulated experiment boxes |
图 2、 图 3中可以看出,水体中氮素浓度变化具有以下几个特征: ①在4月1日向1、 2、 3号箱中注入等量CO(NH2)2和(NH4)2HPO4混合溶液,4月3日实验开始取样后,1、 2、 3号模拟箱中硝酸盐氮浓度已经升高,分别达到12.97、 11.09 和11.39 mg ·L-1,而水体中氨氮浓度提升则相对滞后,在第二个取样日即4月14日才达到第一个峰值,浓度也远低于硝酸盐氮浓度,这同文献[22, 23]在大田实验中所监测的施肥后农田排水中硝酸盐氮和氨氮浓度变化具有相似特征; ②1、 2、 3号模拟箱水体中氨氮和硝酸盐氮浓度变化具有几乎相同的趋势,4月1日向1、 2、 3号箱中注入等量CO(NH2)2
和(NH4)2HPO4混合液后,浓度升高,之后开始下降,6月13日、 7月29日及10月9日再次注入CO(NH2)2和(NH4)2HPO4混合液后,也表现出相似的升高-下降过程; ③9月之前,无论是水体中的氨氮浓度还是硝酸盐氮浓度,1号箱高于2号箱,2号箱又高于3号箱; 9月中旬之后,2、 3号模拟箱水体中的氨氮浓度和硝酸盐氮浓度开始逐渐高于1号模拟箱,进一步比较3号箱又高于2号箱; ④ 4号模拟箱水体中没有加入任何药剂,氨氮和硝酸盐氮浓度表现出的是自然状态下水体-底泥-植物间的氮素运移特征,图 2、 图 3中可以看出,实验期内水体中氨氮和硝酸盐氮浓度有所波动,但变化范围不大. 在10月9日最后一次加水之后,水体中氨氮浓度在11月13日之后、 硝酸盐氮浓度在12月8日之后依然呈现出上升趋势.
1号模拟箱和2号模拟箱的差别在于有无底泥,在同等实验条件下水体中氮素浓度的差异可看做是因底泥吸附作用引起; 2号模拟箱和3号模拟箱区别在于箱体内移栽的芦苇,箱内水体中氮素浓度差异可认为是芦苇作用造成. 因此,引入净化率W做进一步分析.
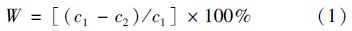
分别计算实验期内底泥净化率(2号模拟箱相对于1号模拟箱)、 植物净化率(3号模拟箱相对于2号模拟箱),并点绘于图 4.

|
图 4 模拟箱水体中氮素净化率变化过程 Fig. 4 Changing process of nitrogen purification rate in simulated experiment boxes |
图 4中可以看出,无论是底泥对水体中氮素的吸附净化,还是植物的吸收净化,基本上可以10月为界划分为两个过程,10月之前各净化率基本均大于零,表现出对水体中氮素的吸收和净化作用,10月之后基本接近于零或小于零,其中在9月28日底泥对氨氮的净化率、 植物对硝氮的净化率已小于零,11月之后除植物对氨氮净化效率接近于零外其他3种净化率均小于零,表现出对箱内水体的“负净化”作用,对水体实质上是一个再污染过程,形成“内源污染”或“二次污染”. 文献[24]也对此有过研究,结果基本相似. 具体原因除文献已经提出的底泥解吸、 芦苇生长能力减弱甚至部分根茎叶枯败作用外,还应该包括温度降低、 微生物作用减弱等因素.
2.2 底泥中氮素含量变化特征点绘2号、 3号、 4号模拟箱底泥中氨氮、 硝酸盐氮含量变化过程见图 5.

|
图 5 模拟实验箱底泥中硝酸盐氮和氨氮浓度变化 Fig. 5 Variation of nitrate and nitrogen concentration in sediment in simulated experiment boxes |
可以看出,2号、 3号模拟箱底泥中氨氮、 硝酸盐氮含量变化过程基本一致,其中氨氮含量在6月之前有一个小的升降波动,6月之后直到11月之前是一个明显的上升过程,之后开始下降,在整个升降过程中,3号模拟箱底泥中氨氮含量几乎全低于2号箱底泥中氨氮含量; 底泥中硝酸盐氮的含量,2号、 3号模拟箱不仅变化过程一致,而且整体上都呈现出下降的趋势,其差异体现在不同的时间段下降的幅度不同. 4号模拟箱底泥中氨氮、 硝酸盐氮含量变化趋势与2号、 3号模拟箱具有一定差异,大致呈“V”字形变化,7月中旬以前是一个整体下降过程,之后则是一个快速上升又缓慢下降的过程.
底泥中氮素含量变化以及不同形式氮素之间的转化相对较为复杂[25]. 首先是底泥对水体中氮素的吸附,尤以对还原态氨氮的吸附能力较强,当水体中氨氮浓度较高时,底泥可以通过阳离子交换吸附去除水溶液中的铵离子,以塑造自身新的平衡. 实验初期、 6月13日、 7月29日及10月9日再次向模拟箱注入CO(NH2)2和(NH4)2HPO4混合液,图 5可以看出2、 3号模拟箱底泥中氨氮含量的增加; 其次是植物对底泥中氨氮和硝酸盐氮含量的影响,由于生长需求芦苇根系吸收并向茎叶传递以同化为自身物质,3号模拟箱底泥中氨氮和硝酸盐氮含量在整个实验过程中一直低于2号模拟箱; 另外,由于底泥基质中发育着大量的好氧、 厌氧及兼性的微生物,硝化作用与反硝化作用交替进行,从图 5底泥内氨氮和硝酸盐氮含量变化趋势初步分析,2、 3号模拟箱底泥中应以反硝化作用为主.
鉴于底泥中氮素含量变化及不同形式氮素之间转化的复杂性,点绘2、 3号模拟箱底泥中总氮含量及芦苇对底泥中总氮的吸收效率[计算同式(1)]见图 6. 从中可以看出,2、 3号箱底泥中总氮含量在7月29日注入CO(NH2)2和(NH4)2HPO4混合液后进入平台期,10月之后持续呈下降趋势; 芦苇对底泥中的总氮的影响效率也比较明显,在根系生长期的4-7月,芦苇净化影响效率较大,8-11月大致是一个平衡期,11月之后对水体影响也是“负净化”.

|
图 6 底泥中总氮含量及芦苇对底泥中总氮的净化效率 Fig. 6 Total nitrogen content in sediment and the purification efficiency of reeds |
作为水-底泥-植物复合生态体的重要组成,随着植物体的生长-衰败循环,植物体内的氮素含量也在不停地变化,点绘3号、 4号模拟箱内芦苇体不同时期根茎叶总氮含量变化过程见图 7.

|
图 7 植物体内总氮含量变化 Fig. 7 Variation of total nitrogen content in plants |
由于3号模拟箱是新移栽芦苇,4月植入后直到6月才开始取样. 图 7可以看出,6-11月,3号箱叶系总氮含量最高,9月最高含量达到38 g ·kg-1,之后开始下降直到11月芦苇叶脱落; 4号箱叶系总氮含量从6月开始上升,7月达到17 g ·kg-1,8-9月一直稳定在这个水平,10月开始下降直至芦苇叶体脱落; 芦苇根茎总氮含量远低于叶系总氮含量,其中在6月芦苇开始生长期,根系总氮含量略高于茎系含量,7-9月芦苇生长旺盛期,茎系总氮含量高于根系含量,并且在此期间,3号模拟箱新栽芦苇根茎系总氮含量在10月之前一直高于4号箱多年生芦苇根茎系含量.
文献[24, 26]等提出,应该定期收割芦苇,以带走植物体本身从底泥及水体中吸收的氮素,消减氮污染源. 从3、 4号模拟箱对比可以看出,定期收割芦苇不仅可带走氮营养源,同时考虑到第二年新生芦苇的“生长拉力”作用,还可以刺激新生芦苇对底泥及水体中氮素的吸收和同化能力,提高芦苇对水体的净化水平.
2.4 水体-底泥-芦苇氮素含量响应分析农田排水沟渠中水体-底泥-水生植物构成一个复合生态体,单一介质内部及不同介质间氮素的迁移和转化都不是孤立的. 不考虑外界环境因素影响的前提下,系统内水体中氮素浓度的升高或降低,必然受底泥或水生植物介质内或介质间氮素迁移转化影响. 以3号模拟箱和4号模拟箱为例,分别点绘水体-底泥-芦苇不同时期介质内总氮含量见图 8、 图 9. 其中芦苇体总氮含量取根、 茎、 叶总氮含量的平均值.

|
图 8 3号模拟箱水体-底泥-芦苇总氮含量变化 Fig. 8 Variation of total nitrogen in water-sediment-reeds in box 3 |

|
图 9 4号模拟箱水体-底泥-芦苇总氮含量变化 Fig. 9 Variation of total nitrogen in water-sediment-reeds in box 4 |
图 8中可以看出,整个实验期内3号模拟箱内水体中总氮浓度的变化呈现出高-低-高的“凹”形变化趋势,底泥内总氮含量则呈现出低-高-低的微“凸”形变化特征,而芦苇因受生长期影响由高到低. 10月之前,即使人为调控注入CO(NH2)2和(NH4)2HPO4混合液,但由于底泥的吸附和芦苇的生长吸收,水体中总氮浓度总体上依然呈下降趋势; 而10月之后,水体中总氮浓度再次升高直至实验期结束,同时底泥中总氮含量下降,一定程度上反映出水体-底泥-芦苇介质间氮素的迁移转化动态及其对水体中氮素浓度的影响机制.
图 9为4号模拟箱内水体-底泥-芦苇不同介质内总氮含量变化过程. 整个实验期内,水体中总氮浓度和底泥中总氮含量的变化趋势和3号模拟箱相类似,但变动的幅度要小一些. 可以看出,水体中总氮浓度的变化同时伴随着底泥中总氮含量的波动,10月之前,水体内总氮浓度和底泥中总氮含量循环升降,但其过程并不同步,水体中总氮浓度升降在前,底泥中总氮含量波动在后,一定程度上反映出底泥对水体中氮素的吸附,中间同时具有多年生芦苇对底泥及水体中氮素的吸收与同化作用; 10月之后,水体中总氮浓度持续上升,而底泥中的总氮含量则持续缓慢下降,芦苇体同时进入衰败期体内总氮下降. 也反映了水体-底泥-芦苇介质间氮素的迁移转化对水体中氮素浓度的影响机制.
3 结论(1)水体中氮素浓度受底泥吸附及植物吸收和根系活动层分解影响,10月之前这种影响表现为促使水体中氮素浓度降低,是水体的氮净化过程; 11月之后则表现为促使水体中氮素浓度升高,易形成二次氮污染.
(2)受人工调控、 微生物及芦苇根系活动影响,底泥中的总氮实验期整体上可划分为升-平衡-下降这3个阶段; 芦苇对底泥中总氮影响比较明显,根系生长期4-7月,芦苇净化影响效率较大,8-11月大致是一个平衡期,11月之后可能引起底泥总氮含量上升.
(3)芦苇体中叶系氮素含量最高,定期收割芦苇不仅可带走氮营养源,同时还可以刺激第二年新生芦苇对底泥及水体中氮素的吸收和同化能力,提高芦苇对水体的净化水平.
(4)模拟沟渠水-底泥-芦苇生态体内氮迁移联系紧密,任一介质内或介质间氮素迁移转化都会引起水体中氮素浓度相应调整.
| [1] | Meuleman A F M, Beltman B. The use of vegetated ditches for water quality improvement[J]. Hydrobiologia, 1993, 253 (1-3): 375. |
| [2] | Kröger R, Holland M M, Moore M T, et al. Hydrological variability and agricultural drainage ditch inorganic nitrogen reduction capacity[J]. Journal of Environmental Quality, 2007, 36 (6): 1646-1652. |
| [3] | Kröger R, Holland M M, Moore M T, et al. Agricultural drainage ditches mitigate phosphorus loads as a function of hydrological variability[J]. Journal of Environmental Quality, 2008, 37 (1): 107-113. |
| [4] | Strock J S, Dell C J, Schmidt J P. Managing natural processes in drainage ditches for nonpoint source nitrogen control[J]. Journal of Soil and Water Conservation, 2007, 62 (4): 188-196. |
| [5] | Needelman B A, Kleinman P J A, Strock J S, et al. Improved management of agricultural drainage ditches for water quality protection: An overview[J]. Journal of Soil and Water Conservation, 2007, 62 (4): 171-178. |
| [6] | 晏维金, 尹澄清, 孙濮, 等. 磷氮在水田湿地中的迁移转化及径流流失过程[J]. 应用生态学报, 1999, 10 (3): 312-316. |
| [7] | 姜翠玲, 崔广柏, 范晓秋, 等. 沟渠湿地对农业非点源污染物的净化能力研究[J]. 环境科学, 2004, 25 (2): 125-128. |
| [8] | 杨林章, 周小平, 王建国, 等. 用于农田非点源污染控制的生态拦截型沟渠系统及其效果[J]. 生态学杂志, 2005, 24 (11): 1371-1374. |
| [9] | 杨林章, 施卫明, 薛利红, 等. 农村面源污染治理的"4R"理论与工程实践——总体思路与"4R"治理技术[J]. 农业环境科学学报, 2013, 32 (1): 1-8. |
| [10] | 李强坤, 胡亚伟, 李怀恩. 农业非点源污染物在排水沟渠中的模拟与应用[J]. 环境科学, 2011, 32 (5): 1273-1278. |
| [11] | 李强坤, 胡亚伟, 罗良国. 青铜峡灌区典型排水沟水污特征解析[J]. 环境科学, 2012, 33( 5): 1579-1586. |
| [12] | 张树楠, 贾兆月, 肖润林, 等. 生态沟渠底泥属性与磷吸附特性研究[J]. 环境科学, 2013, 34 (3): 1101-1106. |
| [13] | 李如忠, 钱靖, 董玉红, 等. 城郊排水沟渠溶质传输的暂态存储影响及参数灵敏性[J]. 环境科学, 2015, 36 (2): 481-489. |
| [14] | 彭世彰, 高焕芝, 张正良. 灌区沟塘湿地对稻田排水中氮磷的原位削减效果及机理研究[J]. 水利学报, 2010, 41 (4): 406-411. |
| [15] | 潘乐, 茆智, 董斌, 等. 塘堰湿地减少农田面源污染的试验研究[J]. 农业工程学报, 2012, 28 (4): 130-135. |
| [16] | 何军, 崔远来, 吕露, 等. 沟渠及塘堰湿地系统对稻田氮磷污染的去除试验[J]. 农业环境科学学报, 2011, 30 (9): 1872-1879. |
| [17] | 苏文辉, 于晓菲, 王国平, 等. 沟渠化对三江平原湿地铁元素沉积过程的影响[J]. 环境科学 2015, 36 (4): 1431-1436. |
| [18] | 张雅杰, 邵庆军, 李海彩, 等. 生态景观型灌排系统面源污染防治试验及生态响应[J]. 农业工程学报, 2015, 31 (1): 133-138. |
| [19] | 李强坤, 胡亚伟, 宋常吉. 农田排水沟渠净化非点源污染的研究实践和关键问题[A]. 见:中国环境科学学会学术年会论文集[C]. 成都: 中国环境科学学会, 2014. 580-583. |
| [20] | 陆海明, 孙金华, 邹鹰, 等. 农田排水沟渠的环境效应与生态功能综述[J]. 水科学进展, 2010, 21 (5): 719-725. |
| [21] | 国家环境保护总局. 水和废水监测分析方法[M]. (第四版). 北京: 中国环境科学出版社, 2002. |
| [22] | 李强坤, 李怀恩, 胡亚伟, 等. 青铜峡灌区氮素流失试验研究[J]. 农业环境科学学报, 2008, 27 (2): 683-686. |
| [23] | 李强坤, 陈伟伟, 孙娟, 等. 青铜峡灌区氮磷运移特征试验研究[J]. 环境科学, 2010, 31 (9): 2048-2055. |
| [24] | 姜翠玲, 范晓秋, 章亦兵. 农田沟渠挺水植物对N、 P的吸收及二次污染防治[J]. 中国环境科学, 2004, 24 (6): 702-706. |
| [25] | 李强坤, 胡亚伟, 孙娟. 农业非点源污染物在排水沟渠中的迁移转化研究进展[J]. 中国生态农业学报, 2010, 18 (1): 210-214. |
| [26] | 余红兵, 肖润林, 杨知建, 等. 五种水生植物生物量及其对生态沟渠氮、 磷吸收效果的研究[J]. 核农学报, 2012, 26 (5): 798-802. |