 2025, Vol. 46
2025, Vol. 46


2. 河海大学环境学院, 南京 210024
2. College of Environment, Hohai University, Nanjing 210024, China
塑料产品因其质轻、耐用及价廉等优点在日常生活中大量应用. 据统计, 目前全球塑料产量达到了每年4.6亿t, 并预计到2060年增加至12.3亿t[1]. 但由于极低的回收利用率, 大部分塑料产品未经处理即直接排入水环境, 并在物理、化学和生物等自然力作用下裂解为粒径小于5 mm的微塑料(microplastics, MPs)[2]. 由于粒径小、重量轻和难降解等特点, MPs极易随水和大气循环等过程在海洋、河流和湖泊中长距离迁移和持久赋存[3~6]. 有研究表明, MPs因其尺寸和形态等因素, 极易被水生生物作为食物所误食, 进而累积在水生生物体内并产生毒性效应, 包括诱导氧化应激、促进细胞死亡和引起肠道损伤等反应, 乃至造成个体死亡和种群演替[7~10]. 因此, 水环境中MPs的潜在生态风险引起了广泛关注, 解析MPs的生物累积过程及生物效应成为评估其生态风险的重要前提.
浮游动物是水生食物网的基础, 其一方面以浮游植物为食, 一方面又是许多更高营养级捕食者的重要食物来源. 水环境中的MPs经摄食和黏附等途径进入浮游动物体后, 极易通过食物链传递途径影响更高营养级的水生生物[11, 12], 甚至进入人体组织影响生命健康[13~15]. 因此, MPs在浮游动物体内的累积和生物效应在评估MPs对水生态系统健康的影响方面起着重要作用. 在已有研究中, 聚苯乙烯(polystyrene, PS)MPs可以在浮游动物大型溞(Daphnia magna)体内产生显著累积, 进而损伤其食物过滤器, 导致消化道堵塞和滤水率、摄食率降低等[10]. 刘加强等[16]也证实可降解聚乳酸MPs明显抑制了大型溞的游泳活力和摄食行为, 改变了其生活史和繁殖能力. 尽管PS-MPs暴露未对丰年虾(Artemia saline)产生明显的急性和慢性毒性作用, 但其长时间暴露仍导致了肠道细胞出现异常, 影响了丰年虾的营养吸收和能量代谢过程[17, 18]. 以上研究表明, MPs对浮游动物的毒性效应可能与其自身特性等多种因素有关. 然而, MPs自身特性变化对其在浮游动物体内的生物累积动力学影响如何仍少有报道, 其生物效应研究仍主要集中在氧化应激响应等方面, 微生物组成和代谢功能等直接反映浮游动物吸收和代谢能力的生物效应研究尚需进一步拓展.
丰年虾是一种典型的滤食性浮游动物, 也是生态毒理学领域使用的模式物种之一[19]. 丰年虾主要生活在高盐度水域中, 具有极强的环境适应能力和繁殖能力, 其休眠卵抗逆性强, 并可在短时间内孵化成无节幼虫. 因具有高蛋白质和丰富的营养物质, 丰年虾被广泛作为水产动物苗种的开口饵料, 在水产养殖及其传染病治疗中广泛应用. 因此, 本研究以丰年虾作为典型浮游动物, 通过分析不同MPs特性(浓度、粒径)及营养条件下典型PS-MPs在丰年虾无节幼虫体内的摄入与清除情况, 解析其生物浓缩系数(bioconcentration factor, BCF)和半衰期(half-life period, T1/2)变化, 评估不同条件下PS-MPs在丰年虾体内的生物累积能力. 同时, 结合微生物组学及代谢组学分析, 解析PS-MPs暴露对丰年虾体内菌群构成和代谢功能的影响, 以期为水环境中MPs的风险评估和管理提供科学支撑.
1 材料与方法 1.1 试验材料本文选取PS-MPs为研究对象, 其作为日常生活中常见的塑料之一, 在水环境中频繁检出[20]. 同时, PS-MPs也是目前研究MPs生物胁迫效应常用的塑料种类之一[20]. 为便于检测, 绿色荧光标记的PS-MPs用于累积清除试验, 其最大激发波长和最大发射波长分别为488 nm和518 nm, 荧光染料采用包被法埋藏在MPs内部, 且在暴露和后续处理过程中没有发生泄漏. 非荧光标记的PS-MPs用于后续生物毒性效应暴露试验. 所有PS-MPs均购自赛尔群科技有限公司(天津, 中国). 所有PS-MPs在使用前均用纯水超声清洗20 min, 以去除其表面的化学试剂. 为保证MPs的分散性, 所有暴露溶液均现配现用.
1.2 受试生物试验用丰年虾卵购自潍坊禾羽田公司, 其在孵化前存储于-20℃冰箱中. 为保持溶液的新鲜并排除其他污染物干扰, 采用过滤的人工海水孵化丰年虾卵, 盐度保持为30 g·L-1. 孵化溶液pH为8.1±0.1, 溶解氧浓度≥8 mg·L-1, 孵化时间为24 h[21]. 孵化后, 立即将丰年虾无节幼体(instar I)转移至(25±1)℃的光照培养箱中培养24 h, 设置光照度为3 000 lx, 光/暗周期为比为12∶12. 由于孵化时间较短, 无节幼体在培养期间不喂食. 随后, 选择健康、有活力的丰年虾幼虫(instar II)进行后续的累积和毒性暴露试验.
1.3 暴露试验根据现有研究中MPs在不同水环境中的最高检出质量浓度(7 mg·L-1)和水生毒理试验常用暴露浓度[22, 23], 选择相同数量级的10 mg·L-1为暴露浓度, 开展不同PS-MPs暴露粒径试验(0.1、1、5和10 μm), 探究不同暴露粒径PS-MPs在丰年虾中的累积动力学. 根据粒径暴露试验结果, 选择累积明显且属于微塑料尺寸范围的1 μm粒径PS-MPs为暴露对象, 设置浓度梯度暴露试验(0.1、1、10、20和50 mg·L-1), 探究不同暴露浓度PS-MPs在丰年虾中的累积动力学. 基于PS-MPs特性试验, 选择1 μm PS-MPs(10 mg·L-1)为暴露对象, 开展不同营养条件下的暴露试验(无喂食、暴露期给食和清除期给食物), 食物为0.02 g干酵母粉, 探究不同营养条件下PS-MPs在丰年虾中的累积动力学. 累积暴露周期设置为24 h, 之后用超纯水反复清洗丰年虾3次, 以去除附着在丰年虾表面的PS-MPs, 并将其置换到人工模拟海水中进行清除试验24 h. 本试验期间, 所有处理组均在光照培养箱中进行暴露, 光/暗周期、温度、pH和溶解氧等培养参数均与孵化阶段一致. 除营养试验外, 其他处理组均不给予食物. 每个暴露试验均包括一个空白对照组, 每个处理组均设置3个平行.
结合累积试验, 利用非荧光标记的1 μm PS-MPs开展毒性暴露试验. 暴露组别仍设置为空白对照组和PS-MPs处理组, 暴露浓度为10 mg·L-1. 其余暴露条件设置与累积试验相同, 暴露时间为24 h, 随后采集丰年虾用于随后的微生物多样性和代谢组学分析.
1.4 MPs累积清除分析在累积期间(2、4、8、12、24 h)和清除期间(25、26、28、32、48 h), 分别从各处理组中随机取出20只丰年虾, 用超纯水反复清洗3次, 以去除其表面粘附的MPs. 随后, 将丰年虾移至1.5 mL冷冻管中称重. 基于文献[24], 采用KOH法对丰年虾进行消解, 在60℃下振荡消解至溶液澄清. 随后, 采用荧光分光光度计(Hitachi F7000, 日本)测定累积MPs的荧光强度, 激发和发射波长设置为488 nm和518 nm, 采用外标法定量MPs的暴露浓度和累积含量.
1.5 累积动力学模拟采用“双箱模型”对丰年虾中PS-MPs的累积与清除过程进行一级动力学模拟, 该模型可用于计算生物体内污染物浓度未达到稳态时的动力学参数, 计算过程如下.

|
(1) |
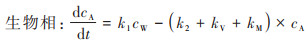
|
(2) |
式中, k1为吸收速率常数, k2为清除速率常数, kM为代谢速率常数, kV为挥发速率常数, cW为水中MPs浓度, cA为生物体内MPs浓度.
由于暴露周期短且不发生生物代谢, 生物体的PS-MPs自然挥发和生物代谢可以忽略, 即kM=0, kV=0, 由公式(1)和(2)可得:
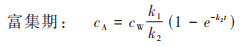
|
(3) |
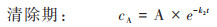
|
(4) |

|
(5) |
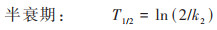
|
(6) |
毒性暴露24 h后, 每个处理组随机挑选200只丰年虾, 共3个平行用于微生物组成和多样性分析. 丰年虾中的总DNA通过HiPure Soil DNA Kits(基迪奥生物, 广州)进行提取, 并采用Nanodrop 2000微量分光光度计(Thermo Fisher, 美国)进行浓度及纯度检验. 随后, 用带有barcode的特异引物扩增16S rRNA的V3和V4区. 引物序列为341F:CCTACGGGNGGCWG CAG;806R:GGACTACHVGGGTATCTAAT. 扩增系统为ABI StepOnePlus实时荧光PCR系统(Life Technologies, 美国). 扩增过程为:95 ℃下预变性3 min后, 在95 ℃变性30 s, 随后50 ℃下退火30 s, 并于72 ℃下延伸45 s, 共计27个循环, 最后在72 ℃下维持10 min, 并在4 ℃终止. 而后对同一样本的PCR扩增产物进行切胶回收并定量检测. 测序文库构建后利用HiSeq2500 PE250平台(Illumina, 美国)的进行测序.
1.7 代谢组学分析毒性暴露试验结束后, 每个处理组随机选取200只丰年虾, 共5个平行, 用于代谢组学分析. 经清水冲洗3次后, 选取的丰年虾样品置于冻存管中冷冻处理, 随后加入提取液[甲醇∶乙腈=1∶1(体积比)]进行代谢物提取. 样品提取液在混匀后置于冰水浴中超声10 min, 并在冷冻条件下静置1 h, 随后在4 ℃下离心15 min(12 000 r·min-1), 取上清液于2 mL进样瓶中并利用超分辨率液质联用仪(Q Exactive HFX, Thermo Fisher, 美国)进行上机检测. 其色谱条件为柱温保持在25℃, 流速为0.5 mL·min-1, 进样量为2 μL, 流动相为A(水+25 mmol·L-1乙酸铵+25 mmol·L-1氨水)和B(乙腈)组成. 结合使用负离子模式和正离子模式进行全扫描, 以确保代谢物处于高覆盖程度. 其质谱条件为:离子源温度600℃, 产物离子扫描范围:25~1 000, 产物离子的单个光谱扫描累积时间为0.05 s. 基于采集的一、二级质谱数据进行非靶向代谢组学分析.
1.8 数据分析PS-MPs在丰年虾体内的累积与清除数据处理分析采用SPSS 26软件, 数据的正态性和同质性采用Shapiro-Wilk和Levene检验进行分析, 利用单因素方差分析(ANOVA)中Tukey事后检验方法确定数据之间的显著差异性, 所有显著性水平为P<0.05. 数据的可视化由OriginPro 2022软件完成. 微生物组学分析在Omicsmart实时交互平台(http://www.omicsmart.com)分析. 对微生物菌群多样性及丰富度信息, 利用Uparse软件对所有样品的全部Effective Tags序列聚类, 以97%的一致性将序列聚类成为OTU(operational taxonomic unit). 随后进行微生物alpha多样性分析和物种结构分析. 非靶向代谢组学信息同样在Omicsmart实时交互平台(http://www. omicsmart.com)进行分析, 使用基于XCMS的R程序包对高分辨质谱数据进行提取和分析, 并基于质谱数据库进行数据注释, 得到组间差异代谢物, 并通过应用超几何检验, 找出差异代谢物表达显著性富集的路径, 注释到KEGG数据库种进行差异代谢通路富集分析.
2 结果与讨论 2.1 不同暴露浓度PS-MPs在丰年虾中的累积与清除不同暴露浓度PS-MPs在丰年虾体内的累积如图 1所示. 暴露期间, PS-MPs在丰年虾体内的累积随暴露时间的增加而增加, 从12 h后开始缓慢增长, 并在24 h后达到最大值. 随着PS-MPs暴露浓度的增加, 其在丰年虾体内的累积含量也随之增大, 呈现出明显的浓度依赖性. 清除期间, PS-MPs从丰年虾体内的排出主要发生在前8 h, 其清除率为83.0%~93.2%;浓度越高, 其生物清除率越高. Wang等[25]也发现粒径小于10 μm的MPs均可被水生生物快速清除.

|
图 1 不同暴露浓度PS-MPs在丰年虾体内的累积和清除动力学 Fig. 1 Accumulation and clearance of PS-MPs in brine shrimp treated with different concentrations |
双箱模型模拟结果显示, 随着PS-MPs暴露浓度的增加, 其在丰年虾中的吸收速率常数k1和清除速率常数k2显著提升(表 1). 相较于低暴露浓度组(0.1 mg·L-1), 高暴露浓度中的k1和k2分别提升了1.86~3.40倍和1.87~4.16倍, 表明PS-MPs在生物体内的吸收和清除速率均与暴露浓度密切相关. 对比不同暴露浓度PS-MPs的生物富集能力后发现, 其BCF为0.308~0.440, 均小于1, 表明PS-MPs短时间暴露在丰年虾体内未呈现出生物富集潜能. 进一步对比不同暴露浓度PS-MPs在丰年虾体内的半衰期后发现, 随着暴露浓度的升高, 其半衰期从0.335 h下降至0.080 h, 表明PS-MPs的生物清除能力也呈现出明显的浓度依赖性. 因此, PS-MPs在丰年虾体内的生物富集动力学过程与其暴露浓度显著正相关, 其在丰年虾体内更高的清除速率导致其生物富集潜能明显下降. 已有研究显示, 水环境中的MPs极易通过滤食或摄食等过程被水生动物所摄取[26], 暴露浓度越高, 其被生物体所摄取的含量也越高, 导致的生物累积也越大. 此外, 水环境中的MPs还可通过表面吸附、糅合等过程黏附于水生生物体表面, 并在长期包裹作用后成为生物体的一部分[27]. 水环境中的MPs暴露浓度越高, 其与和水生生物接触的几率也越大, 生物累积潜能也随之增大. 而MPs的生物清除是其吸收过程的逆向, 累积越多, 释放清除越快. 但MPs一旦进入生物体内组织, 其排泄清除过程将变得十分缓慢, 受到外排转运过程的制约[28]. 因此, MPs的环境暴露浓度成为影响其生物累积能力的关键因素.
|
|
表 1 不同暴露浓度MPs在丰年虾体内的生物累积动力学参数 Table 1 Kinetic parameters of PS-MP bioaccumulation in brine shrimp treated with different concentrations |
2.2 不同暴露粒径PS-MPs在丰年虾中的累积与清除
不同暴露粒径PS-MPs在丰年虾体内的累积含量如图 2所示. 累积暴露期间, 不同暴露粒径的PS-MPs在丰年虾体内的累积含量存在着显著性差异, 随着暴露粒径的减小, PS-MPs的生物累积含量明显升高, 其中小暴露粒径(0.1 μm)PS-MPs的生物累积含量是大暴露粒径(10 μm)的8.73倍, 最高可达366.67 μg·mg-1. 结果表明, 小暴露粒径的MPs更容易在生物体内产生累积, 这可能是与小暴露粒径MPs可以通过更多路径(如黏膜、皮肤等)进入生物体内有关, 且小暴露粒径MPs在生物体内更易迁移, 而大暴露粒径的MPs则主要通过摄食途径进入生物体, 进而造成两者之间存在明显差别[29]. 高嘉蔚等[10]也发现, 0.1 μm和5 μm的MPs在大型溞体内的积累量和停留量均高于50 μm的MPs, 大暴露粒径MPs主要黏附在胸肢处, 而小暴露粒径MPs则能在大型溞肠道中积累. 此外, 不同暴露粒径PS-MPs从丰年虾体内清除的速率也存在明显不同, 小暴露粒径远高于大粒径, 这可能与小暴露粒径PS-MPs更易通过细胞膜进出生物体组织有关[30, 31]. 已有研究证实, MPs的暴露粒径是决定其在生物体内累积和产生毒性效应的决定性因素[29], 不同暴露粒径的MPs可能会在水生生物体内诱导产生迥异的生物效应[32]. 因此, 不同暴露粒径MPs在生物体内的累积和生物效应都不可忽视.

|
图 2 不同暴露粒径PS-MPs在丰年虾体内的累积和清除动力学 Fig. 2 Accumulation and clearance of PS-MPs in brine shrimp treated with different particle sizes |
不同暴露粒径PS-MPs在丰年虾体内的生物累积动力学参数如表 2所示. 随着PS-MPs暴露粒径的增大, 其在丰年虾体内的吸收速率常数k1从5.36下降至0.17, 清除速率常数k2也由8.33下降至0.84, 表明PS-MPs在丰年虾体内的累积和清除均与其粒径显著负相关. 不同暴露粒径PS-MPs在丰年虾体内的BCF均小于1未呈现出生物富集潜能, 但小暴露粒径PS-MPs的BCF相比大粒径显著增长了2倍至0.6, 表明小暴露粒径PS-MPs在丰年虾体内的生物富集潜能不可忽视. 进一步对比不同粒径PS-MPs在丰年虾体内的半衰期和清除率后发现, 随着暴露粒径的增大, 其在丰年虾体内清除的越慢. 清除期之后, 大暴露粒径PS-MPs在丰年虾体内的残留需要着重关注. 因此, MPs的暴露粒径也是影响其生物累积能力的关键因素. Ma等[33]认为暴露粒径较大的MPs可通过肠道吸收或表皮渗入等途径进入水生生物体的内部器官, 而对于暴露粒径很小的纳米尺寸MPs, 其也可能通过内吞、细胞旁渗透以及主动转运等多种复杂作用过程而进入生物体, 进而产生比大暴露粒径MPs更高的生物累积潜能. 但本研究中较短的暴露时间也可能导致小暴露粒径MPs难以深入丰年虾的内部组织, 因而极易清除, 而大暴露粒径的MPs则可能因为物理阻塞等作用而难以从消化道中快速清除, 进而产生较低的清除效率.
|
|
表 2 不同暴露粒径PS-MPs在丰年虾体内的生物累积动力学参数 Table 2 Kinetic parameters of PS-MP bioaccumulation in brine shrimp treated with different particle sizes |
2.3 营养条件对丰年虾累积与清除PS-MPs的影响
水生生物不仅可以从环境介质中直接摄取MPs, 也可以通过营养转移间接摄入MPs, 营养条件成为影响MPs迁移累积的重要因素[34]. 在本研究中, 不同营养条件下PS-MPs在丰年虾体内的累积与清除动力学过程如图 3所示. 和对照组相比, 暴露期投喂食物和清除期给予食物均没有显著改变PS-MPs在丰年虾体内的累积含量. 进一步分析不同营养条件下PS-MPs在丰年虾体内的生物累积动力学参数后也发现(表 3), 无论是暴露期喂食还是清除阶段喂食, 丰年虾对PS-MPs的吸收速率常数k1和清除速率常数k2均没有显著改变, 丰年虾对PS-MPs的生物累积能力也没有明显变化, 表明短期内营养条件可能不会影响丰年虾对PS-MPs的吸收及清除. 这一结果与Capó等[35]观察到的现象相类似, 其发现在有无食物的情况下, 金枪鱼(Sparus aurata)90 d内对MPs的累积量相差不大, 清除率则均接近100%, 营养条件不是影响MPs生物累积和清除的关键因素. 然而在地中海贻贝(Mytilus galloprovincialis)体内, 食物存在明显降低了其对MPs的净化, 食物丰度成为影响贻贝清除MPs的重要因素[36]. 尽管也有研究关注食物存在对MPs在浮游动物体内的累积和清除, 但结果并不一致, MPs的形态、喂食时间和物种差异都是影响MPs在浮游动物体内累积的重要条件[37], 短期暴露下营养条件的改变很难扰动水生生物的生理过程, 进而干扰其生物吸收和外排效应. 因此, 本研究中较短的喂食时间(24 h)可能也是产生无差异性结果的重要因素, 后续研究需要关注长时间营养条件作用对MPs生物累积和清除的影响, 短期暴露则更需关注pH、盐度等环境条件的变化, 其可能会更迅速地影响MPs的累积动力学过程.

|
图 3 不同营养条件下PS-MPs在丰年虾体内的累积和清除动力学 Fig. 3 Accumulation and clearance of PS-MPs in brine shrimp under different nutritional conditions |
|
|
表 3 不同营养条件下丰年虾体内PS-MPs的生物累积动力学参数 Table 3 Kinetic parameters of PS-MP bioaccumulation in brine shrimp under different nutritional conditions |
2.4 微生物菌群结构响应
生物体内的微生物菌群在维持宿主内环境稳定和机体稳态中发挥着重要作用, 当微生物菌群受到环境等外在因素影响时, 微生物菌群的结构就会被打破, 导致机体菌群紊乱, 从而影响宿主健康[38]. 因此, 生物体内微生物菌群的稳定性在维持机体物质代谢、营养吸收及生长发育等方面具有重要作用. PS-MPs暴露对丰年虾体内微生物菌群alpha多样性的影响如表 4所示. 其中, 所有处理组中的Goods_coverage指数均达到了99.7%, 说明测序能够全面反映几乎所有菌群物种. 经过PS-MPs处理后, 表示丰年虾体内菌群丰富度的Chao1指数与ACE指数分别上升了74%和66%, 而指征菌群多样性的Shannon指数和Simpson指数也分别上升了18%和8%, 表明PS-MPs暴露会显著提升丰年虾体内微生物菌群的丰富度和多样性, 这与Li等[39]的研究结果相一致, 其也发现聚乙烯等MPs可以显著提升丰年虾肠道中菌群丰富度和多样性. 这可能与MPs可以激活生物体内酶的合成和细胞代谢有关, 其通过为微生物提供生长位点而刺激特异性菌群的生长, 进而提升生物体内微生物菌群的多样性和丰富度[40].
|
|
表 4 不同处理组中丰年虾体内微生物菌群alpha多样性指数 Table 4 Index of alpha diversity of bacteria community in brine shrimp in different treatments |
丰年虾体内菌群在门分类水平下的群落组成如图 4(a)所示. 从中可以看出, 厚壁菌门(Firmicutes)、变形菌门(Proteobacteria)和拟杆菌门(Bacteroidetes)是最主要的3个门类. 厚壁菌门下包括众多有益菌种, 例如乳酸杆菌和瘤胃球菌等[41], PS-MPs暴露后丰年虾体内的厚壁菌门细菌占比由84%下降至41%, 表明PS-MPs会抑制丰年虾体内有益菌的生长, 对丰年虾正常的生命代谢活动造成干扰. Zhang等[42]也报道了PS-MPs暴露会显著降低斑马鱼(Danio rerio)体内厚壁菌门的占比. 变形菌门下包含众多致病菌, 如大肠杆菌、埃希氏菌、沙门氏菌、弧菌等, 极易对虾类等节肢类生物的健康与免疫系统产生负面影响[43, 44]. PS-MPs暴露后, 丰年虾体内变形菌门的占比由对照组的13%上升至49%, 成为丰年虾体内的优势菌门, 表明PS-MPs暴露会增加变形菌门的丰度, 这与先前MPs对肠道菌群的影响相一致[45, 46]. 变形菌门细菌丰度的上升可能会刺激丰年虾体内致病菌的大幅增加, 进而诱发疾病的产生, 对丰年虾的正常生长产生不利影响. 在属水平上[图 4(b)], 硝化螺旋菌属(Nitrospira)、单胞菌属(Brevundimonas)、Allorhizobium⁃Neorhizobium⁃Pararhizobium⁃Rhizobium属、芽孢杆菌属(Bacillus)、endosymbionts8属、海葵海洋杆菌属(Pontibacter)、赖氨酸芽孢杆菌属(Lysinibacillus)、微小杆菌属(Exiguobacterium)、不动杆菌属(Acinetobacter)和乳球菌属(Lactococcus)为丰年虾体内前十的菌属. 但在PS-MPs暴露后, 许多菌属的相对占比出现了明显变化. 其中, 芽孢杆菌属和微小杆菌属的相对丰度分别降低了86%和95%;而乳球菌属和不动杆菌属的相对丰度则显著增加了5.6倍和12.3倍. 芽孢杆菌属具有促进生物生长、提升消化酶活性、改善免疫能力等作用, 广泛分布于动物肠道内[47]. 微小杆菌作为有益菌群, 也具有改善生物体生长和免疫的能力[48, 49]. 丰年虾体内两种菌群相对丰度的下降表明, PS-MPs暴露可能会降低其免疫能力并抑制生长. 不动杆菌属含有多种致病菌, 可以引发炎症和获得性感染[50]. 与对照组相比, PS-MPs暴露明显增加了丰年虾体内不动杆菌的相对丰度, 这可能和MPs可以选择性富集不动杆菌属有关[51]. 门和属水平上的菌群结构变化表明, PS-MPs易诱导丰年虾体内产生致病菌, 并抑制有益菌, 进而导致丰年虾体内菌群紊乱, 干扰其正常的生物代谢活动.

|
图 4 不同处理组中丰年虾体内微生物菌群在门水平和属水平上的组成和相对丰度 Fig. 4 Species composition and relative abundance distribution of bacterial community in brine shrimp in different treatments at the phylum and genus levels |
为探究PS-MPs对丰年虾体内代谢活动的影响, 进一步采用代谢组学分析生物体内代谢路径的变化. 与对照组相比, PS-MPs暴露诱导丰年虾体内产生了2 311种差异代谢物, 其中上调了1 465种, 下调了846种(图 5). 为探究以上差异代谢物所对应的代谢功能, 进一步应用超几何检验方法并通过KEGG数据库对各组间差异代谢物进行KEGG pathway显著性富集分析, 将各组间最具差异性的前20个代谢通路显示在KEGG pathway通路图中, 并以circos图的形式展示其参与的KEGG注释通路图, 剖析其最主要参与的生物代谢及信号传递途径, 结果如图 5所示.

|
(a)KEGG注释通路, (b)前20差异代谢通路 图 5 丰年虾体内KEGG注释通路和前20差异代谢通路的表达 Fig. 5 Expression of KEGG annotated pathway and the top 20 differential metabolic pathways in brine shrimp treated with PS-MPs |
与对照组相比, PS-MPs暴露后丰年虾体内受到影响的主要代谢合成功能依次为嘧啶代谢、硫酸皮肤素/硫酸软骨素生物合成和硫酸乙酰肝素生物合成等. 嘧啶代谢的重要途径之一是嘧啶还原, 包括尿嘧啶被催化还原成为二氢尿嘧啶, 并逐步水解为β-脲基丙酸酯, 最终形成CO2及β-丙氨酸[52, 53]. 相较于对照组, PS-MPs显著上调了丰年虾体内嘧啶代谢相关代谢物5-羟甲基脱氧胞苷酸盐的丰度, β-丙氨酸代谢量也增长了172%, 尿嘧啶含量则下降了35%, 这表明PS-MPs暴露显著加强了丰年虾体内嘧啶代谢的进程, 导致了嘧啶代谢紊乱的发生. 硫酸软骨素及硫酸皮肤素均属于糖胺聚糖, 在生物体发挥着改善代谢综合征、抗炎、抗癌、抗病毒和调节神经等众多生理作用[54~58]. 在PS-MPs作用下, 丰年虾体内与硫酸皮肤素/硫酸软骨素生物合成相关的代谢产物前列腺素出现了显著下调, 仅为对照组的35%, 表明PS-MPs会通过遏制硫酸皮肤素/硫酸软骨素生物的合成而干扰丰年虾体内正常的生命活动.
生物体内代谢功能的变化一方面受制于其自身, 另一方面也和其体内的微生物菌群密切相关. 已有研究证实, 生物体内的微生物菌群是宿主的隐藏代谢器官, 广泛参与生物体的代谢过程, 生物体内微生物菌群落的失调也往往意味着其代谢功能的紊乱. 其中, 生物体内变形菌门丰度的增加意味着其体内炎症水平的升高[59]. 在本研究中, PS-MPs暴露导致丰年虾体内的变形菌门微生物相对丰度显著增加, 表明PS-MPs暴露可能会导致丰年虾体内产生炎症反应, 这一结果和丰年虾体内的炎症调节相关功能紊乱相一致. 此外, 本研究中发现, PS-MPs作用导致丰年虾体内的嘧啶代谢产物β-丙氨酸含量增长了1.7倍, 其后续主要通过芽孢杆菌属微生物产生的丙酮酸转氨酶和烟酰胺腺嘌呤二核苷酸依赖性脱氢酶进行代谢[53]. 当PS-MPs暴露后, 丰年虾体内的芽孢杆菌属微生物相对丰度下降了86%, 导致其不能通过合成相关酶而代谢去除嘧啶代谢中产生的β-丙氨酸, 从而引起β-丙氨酸含量在丰年虾体内的快速增长, 其嘧啶代谢过程受到严重干扰. 同时, 生物体内的芽孢杆菌还可以通过其丰富的酶解作用分解糖类等物质, 并通过其抗生作用干扰宿主的免疫反应[60]. 本研究中PS-MPs暴露对丰年虾体内芽孢杆菌的抑制可能和其体内果糖和甘露糖代谢功能的减弱以及炎症相关的调节功能的下降密切相关. 因此, PS-MPs暴露也可能会通过改变丰年虾体内的微生物组成而导致代谢功能的紊乱.
3 结论(1)PS-MPs在丰年虾体内的累积与清除与其暴露浓度密切正相关, 但与暴露粒径负相关, 营养条件变化在短时间内不会显著改变PS-MPs在丰年虾中的累积和清除过程.
(2)PS-MPs暴露提高了丰年虾体内微生物菌群的丰富度和多样性. 在菌群结构上, PS-MPs暴露显著下调了丰年虾体内厚壁菌门和芽孢杆菌属等有益菌群的占比, 提升了变形菌门和不动杆菌等含有众多致病菌的菌群占比, 导致丰年虾体内菌群结构紊乱.
(3)PS-MPs存在导致丰年虾体内产出了众多差异性代谢产物, 显著改变了其嘧啶代谢和糖胺聚糖生物合成功能, 严重干扰了丰年虾体内正常的生物代谢功能.
| [1] | OECD. Global plastics outlook: policy scenarios to 2060[EB/OL]. https://www.oecd.org/publications/global-plastics-outlook-aa1edf33-en.htm, 2023-04-07. |
| [2] | Thompson R C, Olsen Y, Mitchell R P, et al. Lost at sea: where is all the plastic?[J]. Science, 2004, 304(5672): 838-838. DOI:10.1126/science.1094559 |
| [3] |
李思琼, 王华, 储林佑, 等. 长江流域微塑料污染特征及生态风险评价[J]. 环境科学, 2024, 45(3): 1439-1447. LI S Q, Wang H, Chu L Y, et al. Pollution characteristics and ecological risk assessment of microplastics in the Yangtze River Basin[J]. Environmental Science, 2024, 45(3): 1439-1447. |
| [4] | Wang C H, Zhao J, Xing B S. Environmental source, fate, and toxicity of microplastics[J]. Journal of Hazardous Materials, 2021, 407. DOI:10.1016/j.jhazmat.2020.124357 |
| [5] |
江为群, 胡启武, 简敏菲, 等. 鄱阳湖典型区铜锈环棱螺体内微塑料分布特征[J]. 环境科学, 2022, 43(5): 2633-2639. Jiang W Q, Hu Q W, Jian M F, et al. Distribution characteristics of microplastics in Bellamya aeruginosa in typical area of Poyang Lake[J]. Environmental Science, 2022, 43(5): 2633-2639. |
| [6] | Nirmala K, Rangasamy G, Ramya M, et al. A critical review on recent research progress on microplastic pollutants in drinking water[J]. Environmental Research, 2023, 222. DOI:10.1016/j.envres.2023.115312 |
| [7] | Kumar R, Sharma P, Manna C, et al. Abundance, interaction, ingestion, ecological concerns, and mitigation policies of microplastic pollution in riverine ecosystem: a review[J]. Science of the Total Environment, 2021, 782. DOI:10.1016/j.scitotenv.2021.146695 |
| [8] |
胡嘉敏, 左剑恶, 李金波, 等. 微塑料对鲫鱼生长、肝脏损伤和肠道微生物组成的影响[J]. 环境科学, 2022, 43(7): 3664-3671. Hu J M, Zuo J E, Li J B, et al. Effects of microplastic exposure on crucian growth, liver damage, and gut microbiome composition[J]. Environmental Science, 2022, 43(7): 3664-3671. |
| [9] |
包亚博, 王成尘, 彭吾光, 等. 微塑料的人体富集及毒性机制研究进展[J]. 环境科学, 2024, 45(2): 1173-1184. Bao Y B, Wang C C, Peng W G, et al. Human accumulation and toxic effects of microplastics: a critical review[J]. Environmental Science, 2024, 45(2): 1173-1184. |
| [10] |
高嘉蔚, 赵莎莎, 李富云, 等. 微塑料对大型溞摄食和抗氧化防御系统的影响[J]. 环境科学研究, 2021, 34(5): 1205-1212. Gao J W, Zhao S S, Li F Y, et al. Effects of microplastics on feeding behavior and antioxidant system of Daphnia magna [J]. Research of Environmental Sciences, 2021, 34(5): 1205-1212. |
| [11] | Chen G L, Li Y Z, Wang J. Occurrence and ecological impact of microplastics in aquaculture ecosystems[J]. Chemosphere, 2021, 274. DOI:10.1016/j.chemosphere.2021.129989 |
| [12] | Carbery M, O'Connor W, Palanisami T. Trophic transfer of microplastics and mixed contaminants in the marine food web and implications for human health[J]. Environment International, 2018, 115: 400-409. |
| [13] | Huang W, Song B, Liang J, et al. Microplastics and associated contaminants in the aquatic environment: a review on their ecotoxicological effects, trophic transfer, and potential impacts to human health[J]. Journal of Hazardous Materials, 2021, 405. DOI:10.1016/j.jhazmat.2020.124187 |
| [14] | Mamun A A, Prasetya T A E, Dewi I R, et al. Microplastics in human food chains: food becoming a threat to health safety[J]. Science of the Total Environment, 2023, 858. DOI:10.1016/j.scitotenv.2022.159834 |
| [15] | Hasan J, Dristy E Y, Anjumanara, et al. Dried fish more prone to microplastics contamination over fresh fish – higher potential of trophic transfer to human body[J]. Ecotoxicology and Environmental Safety, 2023, 250. DOI:10.1016/j.ecoenv.2023.114510 |
| [16] |
刘加强, 蒋园园, 杨杨阳, 等. 聚乳酸微塑料对大型溞的毒性效应[J]. 中国环境科学, 2023, 43(8): 4343-4352. Liu J Q, Jiang Y Y, Yang Y Y, et al. Toxic effects of polylactic acid microplastics to Daphnia magna [J]. China Environmental Science, 2023, 43(8): 4343-4352. |
| [17] | Wang Y, Zhang D, Zhang M X, et al. Effects of ingested polystyrene microplastics on brine shrimp, Artemia parthenogenetica [J]. Environmental Pollution, 2019, 244: 715-722. |
| [18] | Wang Y, Mao Z, Zhang M X, et al. The uptake and elimination of polystyrene microplastics by the brine shrimp, Artemia parthenogenetica, and its impact on its feeding behavior and intestinal histology[J]. Chemosphere, 2019, 234: 123-131. |
| [19] | Kim L, Kim S A, Kim T H, et al. Synthetic and natural microfibers induce gut damage in the brine shrimp Artemia franciscana [J]. Aquatic Toxicology, 2021, 232. DOI:10.1016/j.aquatox.2021.105748 |
| [20] | Zhang P, Lu G H, Sun Y, et al. Metagenomic analysis explores the interaction of aged microplastics and roxithromycin on gut microbiota and antibiotic resistance genes of Carassius auratus [J]. Journal of Hazardous Materials, 2022, 425. DOI:10.1016/j.jhazmat.2021.127773 |
| [21] | Kim L, Cui R X, Il Kwak J, et al. Trophic transfer of nanoplastics through a microalgae‐crustacean‐small yellow croaker food chain: inhibition of digestive enzyme activity in fish[J]. Journal of Hazardous Materials, 2022, 440. DOI:10.1016/j.jhazmat.2022.129715 |
| [22] | Yan Z H, Zhao H Z, Zhu P Y, et al. Polystyrene microplastics alter the trophic transfer and biotoxicity of fluoxetine in an aquatic food chain[J]. Journal of Hazardous Materials, 2024, 470. DOI:10.1016/j.jhazmat.2024.134179 |
| [23] | Han M, Niu X R, Tang M, et al. Distribution of microplastics in surface water of the lower Yellow River near estuary[J]. Science of the Total Environment, 2020, 707. DOI:10.1016/j.scitotenv.2019.135601 |
| [24] | Yan Z H, Chen Y F, Bao X H, et al. Microplastic pollution in an urbanized river affected by water diversion: combining with active biomonitoring[J]. Journal of Hazardous Materials, 2021, 417. DOI:10.1016/j.jhazmat.2021.126058 |
| [25] | Wang S X, Hu M H, Zheng J H, et al. Ingestion of nano/micro plastic particles by the mussel Mytilus coruscus is size dependent[J]. Chemosphere, 2021, 263. DOI:10.1016/j.chemosphere.2020.127957 |
| [26] | Pinheiro L M, Ivar do Sul J A, Costa M F. Uptake and ingestion are the main pathways for microplastics to enter marine benthos: a review[J]. Food Webs, 2020, 24. DOI:10.1016/j.fooweb.2020.e00150 |
| [27] | Li Q P, Sun C J, Wang Y J, et al. Fusion of microplastics into the mussel byssus[J]. Environmental Pollution, 2019, 252: 420-426. |
| [28] | Farrell P, Nelson K. Trophic level transfer of microplastic: Mytilus edulis (L.) to Carcinus maenas (L.)[J]. Environmental Pollution, 2013, 177: 1-3. |
| [29] |
李昕桐, 丁平, 孙雅楠, 等. 微塑料对斑马鱼的毒性效应及机制研究进展[J]. 环境化学, 2024, 43(6): 1792-1804. Li X T, Ding P, Sun Y N, et al. Research status of microplastics for zebrafish on the toxicity and mechanism[J]. Environmental Chemistry, 2024, 43(6): 1792-1804. |
| [30] | Elizalde-velázquez A, Carcano A M, Crago J, et al. Translocation, trophic transfer, accumulation and depuration of polystyrene microplastics in Daphnia magna and Pimephales promelas [J]. Environmental Pollution, 2020, 259. DOI:10.1016/j.envpol.2020.113937 |
| [31] | Schür C, Rist S, Baun A, et al. When fluorescence is not a particle: the tissue translocation of microplastics in Daphnia magna seems an artifact[J]. Environmental Toxicology and Chemistry, 2019, 38(7): 1495-1503. |
| [32] | Lin H, Yuan Y Q, Jiang X M, et al. Bioavailability quantification and uptake mechanisms of pyrene associated with different-sized microplastics to Daphnia magna [J]. Science of the Total Environment, 2021, 797. DOI:10.1016/j.scitotenv.2021.149201 |
| [33] | Ma C Z, Chen Q Q, Li J W, et al. Distribution and translocation of micro- and nanoplastics in fish[J]. Critical Reviews in Toxicology, 2021, 51(9): 740-753. |
| [34] | Hasegawa T, Nakaoka M. Trophic transfer of microplastics from mysids to fish greatly exceeds direct ingestion from the water column[J]. Environmental Pollution, 2021, 273. DOI:10.1016/j.envpol.2021.116468 |
| [35] | Capó X, Company J J, Alomar C, et al. Long-term exposure to virgin and seawater exposed microplastic enriched-diet causes liver oxidative stress and inflammation in gilthead seabream Sparus aurata, Linnaeus 1758[J]. Science of the Total Environment, 2021, 767. DOI:10.1016/j.scitotenv.2021.144976 |
| [36] | Chae Y, An Y J. Effects of food presence on microplastic ingestion and egestion in Mytilus galloprovincialis [J]. Chemosphere, 2020, 240. DOI:10.1016/j.chemosphere.2019.124855 |
| [37] |
闫振华, 朱培元, 陆光华. 典型水生生物中微塑料的累积、清除及食物链传递效应[J]. 环境监测管理与技术, 2023, 35(2): 1-7, 11. Yan Z H, Zhu P Y, Lu G H. Bioaccumulation, removal and trophic transfer effect of microplastics in typical aquatic organisms[J]. The Administration and Technique of Environmental Monitoring, 2023, 35(2): 1-7, 11. |
| [38] | Gacesa R, Kurilshikov A, Vich Vila A, et al. Environmental factors shaping the gut microbiome in a Dutch population[J]. Nature, 2022, 604(7907): 732-739. |
| [39] | Li H Y, Chen H W, Wang J, et al. Influence of microplastics on the growth and the intestinal microbiota composition of brine shrimp[J]. Frontiers in Microbiology, 2021, 12. DOI:10.3389/fmicb.2021.717272 |
| [40] | Holguin G, Vazquez P, Bashan Y. The role of sediment microorganisms in the productivity, conservation, and rehabilitation of mangrove ecosystems: an overview[J]. Biology and Fertility of Soils, 2001, 33(4): 265-278. |
| [41] |
刘苗苗, 毕冉冉, 孙玉敬. 果蔬与肠道菌群互作及其健康功效研究进展[J]. 中国食品学报, 2022, 22(4): 387-407. Liu M M, Bi R R, Sun Y J. Recent advances on the interaction between fruits or vegetables and intestinal flora and its effect on health[J]. Journal of Chinese Institute of Food Science and Technology, 2022, 22(4): 387-407. |
| [42] | Zhang J H, Meng H, Kong X C, et al. Combined effects of polyethylene and organic contaminant on zebrafish (Danio rerio): accumulation of 9-nitroanthracene, biomarkers and intestinal microbiota[J]. Environmental Pollution, 2021, 277. DOI:10.1016/j.envpol.2021.116767 |
| [43] | Liao G W, Wu Q P, Mo B H, et al. Intestinal morphology and microflora to Vibrio alginolyticus in pacific white shrimp (Litopenaeus vannamei)[J]. Fish & Shellfish Immunology, 2022, 121: 437-445. |
| [44] | Yin X L, Zhuang X Q, Liao M Q, et al. Transcriptome analysis of Pacific white shrimp (Litopenaeus vannamei) hepatopancreas challenged by Vibrio alginolyticus reveals lipid metabolic disturbance[J]. Fish & Shellfish Immunology, 2022, 123: 238-247. |
| [45] | Liu J, Lv M, Sun A Q, et al. Exposure to microplastics reduces the bioaccumulation of sulfamethoxazole but enhances its effects on gut microbiota and the antibiotic resistome of mice[J]. Chemosphere, 2022, 294. DOI:10.1016/j.chemosphere.2022.133810 |
| [46] | Zhao Y, Qin Z, Huang Z Z, et al. Effects of polyethylene microplastics on the microbiome and metabolism in larval zebrafish[J]. Environmental Pollution, 2021, 282. DOI:10.1016/j.envpol.2021.117039 |
| [47] | Kewcharoen W, Srisapoome P. Probiotic effects of Bacillus spp. from Pacific white shrimp (Litopenaeus vannamei) on water quality and shrimp growth, immune responses, and resistance to Vibrio parahaemolyticus (AHPND strains)[J]. Fish & Shellfish Immunology, 2019, 94: 175-189. |
| [48] | Kim S, Jeon H, Bai S C, et al. Evaluation of Salipiger thiooxidans and Exiguobacterium aestuarii from the Saemangeum reservoir as potential probiotics for pacific white shrimp (Litopenaeus vannamei)[J]. Microorganisms, 2022, 10(6). DOI:10.3390/microorganisms10061113 |
| [49] | Orozco-Medina C, Maeda-Martı́nez A M, López-Cortés A. Effect of aerobic gram-positive heterotrophic bacteria associated with Artemia franciscana cysts on the survival and development of its larvae[J]. Aquaculture, 2002, 213(1-4): 15-29. |
| [50] |
李招吟倩, 谢静玲, 杨葵, 等. 鲍曼不动杆菌感染及治疗的研究进展[J]. 中国抗生素杂志, 2022, 47(8): 766-771. Li Z Y Q, Xie J L, Yang K, et al. Research progresses of Acinetobacter baumannii infection and treatment[J]. Chinese Journal of Antibiotics, 2022, 47(8): 766-771. |
| [51] | Junaid M, Siddiqui J A, Sadaf M, et al. Enrichment and dissemination of bacterial pathogens by microplastics in the aquatic environment[J]. Science of the Total Environment, 2022, 830. DOI:10.1016/j.scitotenv.2022.154720 |
| [52] |
王飞飞. 还原铬梭菌中的铁氧还蛋白依赖性二氢嘧啶还原酶的生物化学研究[D]. 天津: 天津大学, 2020. Wang F F. A ferredoxin-dependent dihydropyrimidine dehydrogenase in Clostridium chromiireducens[D]. Tianjin: Tianjin University, 2020. |
| [53] |
张海彬. 微生物中嘧啶代谢途径的研究进展[J]. 生物技术, 2021, 31(6): 619-624, 566. Zhang H B. Research progress of pyrimidine metabolism in microbes[J]. Biotechnology, 2021, 31(6): 619-624, 566. |
| [54] |
蓝伟, 陈建平. 硫酸软骨素的生物活性及其构效关系研究进展[J]. 食品安全质量检测学报, 2022, 13(15): 4924-4932. Lan W, Chen J P. Research progress on biological activity and structure-activity relationship of chondroitin sulfate[J]. Journal of Food Safety and Quality, 2022, 13(15): 4924-4932. |
| [55] |
胡世伟. 岩藻糖基化海参硫酸软骨素改善胰岛素抵抗及其作用机制的研究[D]. 青岛: 中国海洋大学, 2014. Hu S W. Study of fucosylated chondroitin sulfate isolated from sea cucumber on improving insulin resistance effects and mechanism[D]. Qingdao: Ocean University of China, 2014. |
| [56] | Shmagel A, Demmer R, Knights D, et al. The effects of glucosamine and chondroitin sulfate on gut microbial composition: a systematic review of evidence from animal and human studies[J]. Nutrients, 2019, 11(2). DOI:10.3390/nu11020294 |
| [57] | Rani A, Baruah R, Goyal A. Prebiotic chondroitin sulfate disaccharide isolated from chicken keel bone exhibiting anticancer potential against human colon cancer cells[J]. Nutrition and Cancer, 2019, 71(5): 825-839. |
| [58] | Chen Y M, Wang Y H, Yang S, et al. Glycosaminoglycan from Apostichopus japonicus improves glucose metabolism in the liver of insulin resistant mice[J]. Marine Drugs, 2019, 18(1). DOI:10.3390/md18010001 |
| [59] |
闫振华, 张燕, 包旭辉, 等. 不同官能团微塑料对斑马鱼胚胎菌群和代谢功能的胁迫效应[J]. 环境科学, 2024, 45(3): 1859-1868. Yan Z H, Zhang Y, Bao X H, et al. Biological effect of microplastics with different functional groups on the bacterial communities and metabolic functions of zebrafish (Danio rerio) embryos[J]. Environmental Science, 2024, 45(3): 1859-1868. |
| [60] | Mongkolthanaruk W. Classification of Bacillus beneficial substances related to plants, humans and animals[J]. Journal of Microbiology and Biotechnology, 2012, 22(12): 1597-1604. |