 2025, Vol. 46
2025, Vol. 46

2. 中国科学院生态环境研究中心环境水质学国家重点实验室, 北京 100085
 , LIANG Yuan-fu1,2
, LIANG Yuan-fu1,2 , SUN Zhe2 , LI Meng-kai2 , WANG Jia-le2 , LI Wen-tao2 , BEN Wei-wei2 , ZHANG Guo-zhen1 , QIANG Zhi-min2
, SUN Zhe2 , LI Meng-kai2 , WANG Jia-le2 , LI Wen-tao2 , BEN Wei-wei2 , ZHANG Guo-zhen1 , QIANG Zhi-min2
2. State Key Laboratory of Environmental Aquatic Chemistry, Research Center for Eco-Environmental Sciences, Chinese Academy of Sciences, Beijing 100085, China
饮用水生物安全与人类的健康生活息息相关[1 ~ 3], 据世界卫生组织(WHO)的数据显示, 每年约数百万人死于介水疾病[4, 5]. 因此, 高效、可靠的消毒技术是人民健康的重要保障. 紫外线(UV)消毒是一种新型绿色消毒技术, 具有广谱高效、运维简单和不产生消毒副产物等优势[6], 可以有效的灭活水中绝大部分病原微生物, 目前已经广泛应用于饮用水与污水等消毒领域[7 ~ 11].
基于波长可将UV分为3类[12, 13]:UV-A(320~390 nm)、UV-B(280~320 nm)和UV-C(200~280 nm). 水消毒采用具有高灭活效果的UV-C, 低压汞灯是应用最为广泛的光源, 可输出254 nm UV(UV254). UV254属于常规UV-C波段(250~280 nm), 通过损伤病原微生物内部核酸, 使其无法正常完成复制, 进而失去活性[14 ~ 17]. 此外, 新型光源KrCl准分子灯可输出222 nm UV(UV222), 属于远UV-C波段(200~230 nm)[13, 18]. UV222可通过损伤病原微生物蛋白质(如细菌中的酶和病毒的蛋白衣壳)破坏其生理活动, 实现病原微生物消毒[16, 23]. 结合远UV-C和常规UV-C, 同时破坏病原微生物的蛋白质和核酸[24], 具有协同强化灭活的潜力, 但目前相关研究还十分有限[15, 23, 25].
此外, 病原微生物经常规UV-C灭活后, UV-C辐照受损的DNA会经光、暗复活[26 ~ 30]两种机制进行修复导致复活, 造成较高的饮用水消毒安全风险[31 ~ 36]. DNA修复酶是病原微生物复活不可或缺的部分[33], 其主要成分是蛋白质. 鉴于远UV-C可以对蛋白质进行破坏[29], 双波长UV-C存在抑制病原微生物复活的潜力, 但目前尚没有相关研究.
因此, 本研究搭建了一套配装了KrCl准分子灯和低压汞灯的细管流光反应系统, 可单独/同时输出UV222和UV254, 并进行准确的UV剂量测定;之后以大肠杆菌(Escherichia coli, E. coli)作为模式微生物探究了双波长UV-C对微生物的协同灭活作用, 以及对后续光、暗复活的影响, 并分析了相关机制, 以期为高效和安全水消毒技术的研发提供理论依据.
1 材料与方法 1.1 实验装置由于传统的准平行光束仪难以安装多个UV光源, 本研究搭建了一套配装了KrCl准分子灯(输出UV222)和低压汞灯(输出UV254)的细管流光反应系统, 如图 1所示:KrCl准分子灯以及低压汞灯平行于石英管放置, 提供UV-C辐照, 并通过遮挡管调节实际辐照长度;荧光微探头[37](MFSD)实时监测各UV光源输出的稳定性;风扇控制装置内温度. 在实际实验过程中微生物悬浊液被蠕动泵输送至石英管中接受UV-C辐照, 通过调节泵流量和辐照长度控制辐照时间.

|
图 1 配装KrCl准分子灯和低压汞灯的细管流光反应系统 Fig. 1 Mini-fluidic photoreactor system equipped with KrCl excimer and low-pressure mercury lamps |
装置搭建好后使用辐照计(ILT 960)测定UV222和UV254的输出光谱, 并用KI/KIO3化学感光剂测定相应波长下的UV剂量, 其中1 L的KI/KIO3感光剂的组成为99.6 g KI, 21.4 g KIO3和3.81 g Na2B4O7·10H2O, 测定KI/KIO3感光剂经过细管流光反应系统前后UV352的变化, 利用公式(1)计算得到细管反应器中UV222和UV254的剂量率[38].
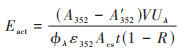
|
(1) |
式中, Eact为KI/KIO3感光剂测得的UV剂量率, mW·cm-2;A'352和A352分别为感光剂溶液在UV⁃C辐照t时间前后352 nm处的吸光度, cm-1;V为溶液体积, mL;Uλ为相应波长下的UV光子的摩尔能量, U222和U254分别为539 213 J·Einstein-1和471 527 J·Einstein-1;ϕλ为KI/KIO3光化学反应的量子产率, ϕ222和ϕ254分别为0.94 mol·Einstein-1和0.72 mol·Einstein-1;ε352为产物I3-的摩尔吸光系数, ε352= 27 636 L·(mol·cm)-1;Acs为辐照截面面积, cm2;t为辐照时间, s;R为溶液界面对UV的反射率, (1–R)为溶液吸收的UV的比例.
1.2 微生物的培养和定量本实验所用的菌种为E. coli(ATCC 11303), 购买自北京科展生物科技有限公司. 选取在含有营养琼脂平板上的E. coli, 将菌落接种至高压灭菌(121 ℃ 15 min)后的胰蛋白胨酵母膏葡萄糖肉汤(TYGB)中, 在37 ℃的恒温摇床中培养12 h后得到高浓度的E. coli悬浊液[37], 丰度约为107 CFU·mL-1.
将培养好的E. coli悬浊液离心(10 min, 3000 g)后弃去上清液, 用灭菌磷酸盐缓冲溶液(PBS, 0.01 mol·L-1, pH 7.2~7.4)清洗3次后重新悬浮于PBS中(丰度约为107 CFU·mL-1), 用于后续实验.
E. coli的定量采用平板计数法[39]:采用无菌PBS对UV-C辐照后的水样进行梯度稀释, 将100 μL稀释后的悬浊液接种在营养琼脂平板上, 平板凝固后倒置放入37℃恒温培养箱中培养12 h, 记录形成的菌落数. 营养琼脂平板成分:胰蛋白胨10 g·L-1, 氯化钠8 g·L-1, 氯化钙0. 294 g·L-1, 维生素B1 10 mg·L-1, 酵母抽提物1 g·L-1, 葡萄糖1 g·L-1, 琼脂粉15 g·L-1, 实验均使用超纯水.
1.3 协同机制的研究为验证双波长辐照协同灭活机制, 使用2, 7-Dichlorodihydrofluorescein diacetate(DCFH-DA)作为探针, 检测细胞内活性氧(ROS)[23, 40, 41]. 取10 mL的E. coli悬浮液于离心管中, 加入20 μL的DCFH-DA, 37℃恒温培育10 min, 于5000 g离心5 min后弃上清液, 用PBS清洗2次去除多余的染料, 将E. coli重新悬浮于PBS中, 使用荧光分光光度计测定488 nm激发波长、525 nm发射波长下的荧光强度, 即可得到细胞内ROS的水平.
1.4 实验过程 1.4.1 实验步骤在UV灭活实验中, 首先预热UV光源(低压汞灯20 min, 准分子灯5 min)至MFSD信号稳定, 将微生物悬浊液泵入细管反应器, 并调整遮光管和泵流量以达到目标辐照时间. 在出水中微生物浓度达到稳定, 即出水体积达到3倍系统管路内液体体积(75 mL)后进行采样, 测定E. coli丰度. 灭活实验中采用的UV波长分别为UV222+254、UV254和UV222, 辐照时间分别为5、10、15、20和25 s, 实验重复3次.
对于复活实验, 分别采集20 mL UV-C辐照后的样品于培养皿和50 mL离心管中, 其中培养皿置于两组荧光灯(15 W, Phillips)下20 cm, 于4.2×103 lx光强下进行光复活, 离心管置于黑箱中进行暗复活, 经1、2、4、6和8 h复活后的样品通过平板计数法测定E. coli丰度. 复活实验中采用的UV波长分别为UV222+254、UV254和UV222, 辐照时间为15 s, 实验重复3次.
1.4.2 灭活速率常数的计算使用Chick-watson模型拟合计算反应系统中单波长与多波长UV-C消毒后的灭活速率常数[26, 42], 对数lg(灭活率)采用公式(2)计算.
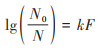
|
(2) |
式中, N0为UV消毒前的E. coli丰度, CFU·mL-1;N为UV消毒后的E. coli丰度, CFU·mL-1;k为E. coli的灭活速率常数, cm2·mJ-1, F为UV剂量, mJ·cm-2.
1.4.3 协同系数的计算使用公式(3)计算不同UV-C辐照时间下灭活的协同系数[15].
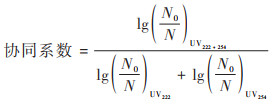
|
(3) |
为评估光、暗复活的效果[27, 43], 采用公式(4)计算lg(修复率), 用于评估已失活(lgN0-lgN)细菌中的复活(lgNt-lgN)程度.
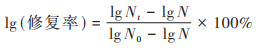
|
(4) |
式中, Nt为光复活后的E. coli丰度, CFU·mL-1.
1.4.5 统计学分析通过t检验比较E. coli在不同光、暗处理下是否存在复活现象[23, 27], 通过协方差分析(ANCOVA)[44]比较不同波长UV-C辐照后E. coli的lg(修复率)是否存在显著差异, P < 0.05为显著不同.
2 结果与讨论 2.1 细管流光反应系统表征细管流光反应系统中配装的KrCl准分子灯和低压汞灯的输出光谱如图 2所示, 输出波长的峰值分别在222 nm和254 nm处, 表明该系统中可以同时输出远UV-C和常规UV-C.

|
图 2 KrCl准分子灯和低压汞灯的输出光谱 Fig. 2 Emission spectra for KrCl excimer lamp and low-pressure mercury lamp |
为了表征细管流光反应系统不同UV光源输出的稳定性, 测定了各光源单独/同时开启时的相对辐照强度和系统温度变化(图 3). KrCl准分子灯和低压汞灯分别在开启5 min和20 min后输出达到稳定. 当KrCl准分子灯和低压汞灯分别单独开启时, 体系温度最终稳定在20.6 ℃±0.3 ℃和20.3 ℃±0.5 ℃, 而当双UV光源同时开启时, 由于产生的热量较高, 体系内温度略微升高, 最终稳定在21.4 ℃±0.4 ℃. 结果表明搭建的细管流光反应系统可以提供稳定的KrCl准分子灯和低压汞灯的单独/同时辐照.

|
(a)UV222辐照强度和系统温度变化, (b)UV254辐照强度和系统温度变化, (c)UV222+254辐照强度和系统温度变化 图 3 KrCl准分子灯和低压汞灯单独/同时开启时的相对输出和系统温度变化 Fig. 3 Variations in relative output and system temperature under individual/simultaneous UV exposure by KrCl excimer lamp and low-pressure mercury lamp |
通过KI/KIO3感光剂分别测定了UV222和UV254在辐照时间为5、10、15、20和25 s时的剂量, 并绘制成图 4. 根据公式(1)计算得到UV222和UV254的Eact分别为0.731 mW·cm-2和0.671 mW·cm-2, R2分别为0.999和0.995. 这表明, 该系统可以获得准确的Eact测定结果, 以及基本一致的UV222和UV254辐照剂量. 此外, UV222+254的Eact值等于UV222+UV254[45], 因此UV222+254的UV辐照强度为1.402 mW·cm-2.

|
图 4 UV222、UV254和UV222+254的剂量 Fig. 4 Fluence rates of UV222, UV254, and UV222+254 |
单波长与双波长UV-C对E. coli的灭活情况如图 5所示. E. coli在单波长UV-C下的灭活剂量响应如图 5(a)所示, 图中横坐标为不同的UV剂量, 纵坐标为E. coli的lg(灭活率), UV222的灭活效果略低于UV254, 灭活速率常数k分别为0.067 3 cm2·mJ-1和0.098 1 cm2·mJ-1. 这表明较UV222而言E. coli更容易被UV254灭活. 此外, 由于初始悬浊液E. coli丰度约为107 CFU·mL-1, 因此E. coli的lg(灭活率)检测限约为7.

|
(a)单波长UV-C对E. coli的灭活动力学, (b)双波长UV-C对E. coli的协同灭活效果 图 5 单/双波长UV-C对E. coli的灭活效果 Fig. 5 Inactivation of E. coli by single- and dual-wavelength UV-C |
双波长UV-C对E. coli的协同灭活如图 5(b)所示, 其中横坐标为不同的辐照时间, 左侧纵坐标为lg(灭活率), 不同辐照时间下左侧的数据柱是UV222、UV254的lg(灭活率)叠加, 右侧的数据柱为UV222+254的lg(灭活率);右侧纵坐标为不同辐照时间下双波长UV-C灭活E. coli的协同系数, 虚线为实验中细管流光反应系统的检测限. 从图 5(b)中可以看出, 当辐照时间超过10 s后UV222+254的消毒效果优于UV222和UV254灭活率的相加, lg(灭活率)提高了约1倍, 最大协同系数为2.2, 表明双波长UV-C对E. coli具有明显的协同灭活作用, 这与Kang[15]和Ramsay等[46]观测到的现象一致.
2.3 单/双波长UV-C对光复活的影响选取单/双波长UV-C辐照15 s后的E. coli样品进行不同时间的光复活实验, 结果如图 6所示, 图 6(a)中横坐标为UV-C辐照后E. coli光复活的时间, 左侧纵坐标为E. coli经不同复活时间后的lg灭活率, 不同光复活时间下左侧的数据柱是UV222、UV254的lg灭活率叠加, 右侧的数据柱为UV222+254的lg(灭活率);右侧纵坐标为经不同复活时间后的协同系数. 从图 6(a)中可以看出:经过UV222辐照后的E. coli并未出现明显的光复活现象, 对数灭活率在0.33~0.43之间波动;经过UV254辐照后的E. coli均表现出了明显的光复活作用, 在复活8 h后共修复了0.39;经UV222+254辐照后的E. coli光复活更为明显, 在8 h后共修复0.9. 此外, 双波长UV-C的协同灭活系数在1.71~2.23之间, 经过光复活后并未出现明显下降, 表明光复活并未对双波长UV-C的协同灭活效果产生明显影响.

|
(a)对数灭活率和协同系数, (b)对数修复率 图 6 单/双波长UV-C对E. coli光复活的影响 Fig. 6 Effects of single- and dual-wavelength UV-C on the subsequent photoreactivation of E. coli |
为更好对比单/双波长UV-C对光复活的影响, 根据公式(4)计算了E. coli在不同复活时间后的lg(修复率), 如图 6(b)所示:图中横坐标为E. coli光复活时间, 纵坐标为lg(修复率). 为明确E. coli是否存在光复活, 通过t检验对E. coli在不同光复活时间下的修复率和初始修复率进行了对比分析, 发现UV222辐照后的E. coli未出现光复活(P > 0.05);UV254和UV222+254辐照后E. coli出现明显的光复活(P < 0.05), 在1、2、4、6和8 h时UV222+254的lg(修复率)均小于UV254, lg(修复率)随复活时间的延长而上升, 最终在8 h后lg(修复率)达到50.8%±11.0%;UV222+254辐照后的E. coli也展现出了明显的光复活现象, 且随复活时间的延长而增长, 但增长速率低于UV254, 在8 h后lg(修复率)达到36.1%±3.8%. 其次, 为明确E. coli在UV254以及UV222+254辐照后的光复活是否存在差异, 采用一阶线性关系拟合了lg(修复率)随时间的变化, 修复率分别为5.98 h-1(UV254)和4.75 h-1(UV222+254), ANCOVA的分析结果表明二者之间存在显著性差异(P < 0.01), UV254的光复活现象相较于UV222+254更为明显. 因此, 当UV222和UV254剂量大致相同时, 双波长UV-C会对E. coli的光复活过程产生抑制作用, 削弱光复活的效果.
2.4 单/双波长UV-C对暗复活的影响选取单/双波长UV-C辐照15 s后的E. coli样品进行不同时间的暗复活实验, 结果如图 7(a)所示, 图中横坐标为UV-C辐照后E. coli暗复活的时间, 左侧纵坐标为E. coli经不同复活时间后的lg(灭活率), 不同光复活时间下左侧的数据柱是UV222、UV254的lg(灭活率)叠加, 右侧的数据柱为UV222+254的lg(灭活率);右侧纵坐标为经不同复活时间后的协同系数. 结果表明单/双波长UV-C辐照后的E. coli均未出现明显的暗复活, 其中UV222+254的对数灭活率进一步从2.48升高至2.80, 呈现暗凋零现象. 此外, 双波长UV-C的协同系数经过暗复活后从2.09上升至了2.47, 表明协同效应在暗复活过程中进一步加强, 这可能是由于暗凋零现象导致的, 这与Quek等[43]观测到的现象一致.

|
(a)对数灭活率和协同系数, (b)对数修复率 图 7 单/双波长UV-C对E. coli暗复活的影响 Fig. 7 Effects of single- and dual-wavelength UV-C on the subsequent dark repair of E. coli |
E. coli在不同暗复活时间后的lg(修复率)如图 7(b)所示. 由于单波长UV-C对E. coli的灭活率较低, 导致lg(修复率)波动范围较大, 为明确E. coli是否存在暗复活, 通过t检验对E. coli在不同暗复活时间下的修复率和初始修复率进行了对比分析, 发现单波长UV-C辐照后的E. coli未出现暗复活(P > 0.05). 而UV222+254辐照后E. coli的lg(修复率)随复活时间呈负增长趋势, 在4 h后下降至约-10%, 经t检验分析后发现暗处理 > 1 h的E. coli均存在暗凋零现象(P < 0.05).
2.5 双波长UV-C协同灭活与抑制复活的机制通过DCFH-DA染色剂测定了细胞胞内ROS, 经UV222、UV254和UV222+254辐照和未经UV-C辐照的E. coli内部荧光强度(a.u.)分别为38.07±0.29、3.18±0.23、46.85±0.6和2.75±0.24. 可以看出, 双波长UV-C辐照后的E. coli内部荧光强度大于单波长UV-C辐照后的荧光强度之和, 表明双波长UV-C辐照可导致胞内ROS水平上升, 机制为UV222抑制了抗ROS氧化酶的活性[15], 导致UV254光子额外产生的ROS无法被抗ROS氧化酶消耗, 进而对微生物组织(如细胞膜和酶)造成损伤, 从而实现协同灭活. 而这些额外产生的ROS可以对E. coli内部的DNA光修复酶造成损伤, 减少光复活过程中对DNA损伤的修复[47, 48], 进而实现对光复活的抑制. 此外, 这些额外产生的ROS还可以对E. coli内部的细胞组织(如细胞器和酶)等进行破坏, 可能会产生延迟性突变效应[43], 或者在缺少营养来源(PBS无法为E. coli提供营养物质)或光线的情况下逐渐失去活性[48], 最终呈现出暗凋零现象.
2.6 环境应用随着新型UV光源(如准分子灯和UV发光二极管等)的不断发展和应用, 传统准平行光束仪难以安装多个UV光源, 无法满足多光源协同消毒的研究需求. 配装多个UV光源细管流光反应系统可单独/同时输出不同波长的UV-C辐照, 输出稳定且可以获得准确的剂量测试结果, 可以用于多UV光源协同消毒研究. 此外, 双波长UV-C可同时对病原微生物的核酸和蛋白质造成损伤, 实现对病原微生物的协同强化灭活, 但远UV-C易被水中的杂质吸收, 因此双波长UV-C更适合应用于较为洁净的水体(如饮用水)消毒, 以及空气消毒. 最后, 双波长UV-C可以削弱微生物在可见光下进行光复活的能力, 而在黑暗条件下则会呈现暗凋零现象. 因此, 饮用水由于消毒后的输送均在黑暗条件下进行, 经双波长UV-C消毒后可以获得更好的消毒效果.
3 结论(1)配装了KrCl准分子灯和低压汞灯的细管流光反应系统可以单独/同时输出222 nm和254 nm的UV-C辐照, 且保持输出的UV剂量率和系统内温度稳定. 通过化学感光剂标定的UV222和UV254的Eact分别为0.731 mW·cm-2和0.671 mW·cm-2.
(2)双波长UV-C灭活存在明显的协同效应, 协同系数最高可达2.2. 协同机制为:UV222+254中的UV222可以有效抑制E. coli中的抗ROS氧化酶, 导致UV254光子产生的ROS无法被消耗, 进而损伤E. coli细胞组织致使其失活.
(3)UV222+254辐照后的E. coli较UV254辐照后的光复活作用被削弱, 最大对数复活率分别为36.1%±3.8%和50.8%±11.0%, 表明双波长UV-C可以抑制E. coli的光复活作用. 而双波长UV-C辐照后的E. coli在黑暗条件下则会出现暗凋零现象, 进一步强化消毒效果.
| [1] | Boretti A, Rosa L. Reassessing the projections of the world water development report[J]. npj Clean Water, 2019, 2(1). DOI:10.1038/s41545-019-0039-9 |
| [2] | Harris A R, Daly S W, Pickering A J, et al. Safe today, unsafe tomorrow: Tanzanian households experience variability in drinking water quality[J]. Environmental Science & Technology, 2023, 57(45): 17481-17489. |
| [3] | Li B, Li X N, Saingam P, et al. Understanding the microbiological quality of drinking water at the point of consumption with citizen science[J]. ACS ES & T Water, 2023, 3(8): 2691-2699. |
| [4] | Lambertini E, Borchardt M A, Kieke B A Jr, et al. Risk of viral acute gastrointestinal illness from nondisinfected drinking water distribution systems[J]. Environmental Science & Technology, 2012, 46(17): 9299-9307. |
| [5] | Song W, Zheng Z L, Alawadhi A H, et al. MOF water harvester produces water from death valley desert air in ambient sunlight[J]. Nature Water, 2023, 1(7): 626-634. DOI:10.1038/s44221-023-00103-7 |
| [6] | Huang R X, Ma C X, Huangfu X L, et al. Preparing for the next pandemic: predicting UV inactivation of coronaviruses with machine learning[J]. Environmental Science & Technology, 2023, 57(37): 13767-13777. |
| [7] | Song K, Mohseni M, Taghipour F. Application of ultraviolet light-emitting diodes (UV-LEDs) for water disinfection: a review[J]. Water Research, 2016, 94: 341-349. DOI:10.1016/j.watres.2016.03.003 |
| [8] | Sun Z, Blatchley E R. Tetraselmis as a challenge organism for validation of ballast water UV systems[J]. Water Research, 2017, 121: 311-319. DOI:10.1016/j.watres.2017.05.052 |
| [9] | Linden K G, Hull N, Speight V. Thinking outside the treatment plant: UV for water distribution system disinfection[J]. Accounts of Chemical Research, 2019, 52(5): 1226-1233. DOI:10.1021/acs.accounts.9b00060 |
| [10] | Ma B, Linden Y S, Gundy P M, et al. Inactivation of coronaviruses and phage Phi6 from irradiation across UVC wavelengths[J]. Environmental Science & Technology Letters, 2021, 8(5): 425-430. |
| [11] |
徐丽梅, 张崇淼, 王晓昌, 等. 紫外线和次氯酸钠对Escherichia coli和Poliovirus的消毒作用[J]. 环境科学, 2017, 38(5): 1928-1935. Xu L M, Zhang C M, Wang X C, et al. Disinfection action of ultraviolet radiation and chlorination on Escherichia coli and Poliovirus[J]. Environmental Science, 2017, 38(5): 1928-1935. |
| [12] | Moreno-Andrés J, Tierno-Galán M, Romero-Martínez L, et al. Inactivation of the waterborne marine pathogen Vibrio alginolyticus by photo-chemical processes driven by UV-A, UV-B, or UV-C LED combined with H2O2 or HSO5-[J]. Water Research, 2023, 232. DOI:10.1016/j.watres.2023.119686 |
| [13] | Blatchley III E R, Brenner D J, Claus H, et al. Far UV-C radiation: an emerging tool for pandemic control[J]. Critical Reviews in Environmental Science and Technology, 2023, 53(6): 733-753. DOI:10.1080/10643389.2022.2084315 |
| [14] |
李静, 王大宁, 曲红梅, 等. 紫外线对大肠杆菌的损伤机制研究[J]. 军事医学, 2016, 40(9): 725-728. Li J, Wang D N, Qu H M, et al. Mechanism of Escherichia coli injury under UV disinfection[J]. Military Medical Sciences, 2016, 40(9): 725-728. |
| [15] | Kang J W, Kim W J, Kang D H. Synergistic effect of 222-nm krypton-chlorine excilamp and mild heating combined treatment on inactivation of Escherichia coli O157:H7 and Salmonella Typhimurium in apple juice[J]. International Journal of Food Microbiology, 2020, 329. DOI:10.1016/j.ijfoodmicro.2020.108665 |
| [16] | Kang J W, Kim S S, Kang D H. Inactivation dynamics of 222 nm krypton-chlorine excilamp irradiation on Gram-positive and Gram-negative foodborne pathogenic bacteria[J]. Food Research International, 2018, 109: 325-333. DOI:10.1016/j.foodres.2018.04.018 |
| [17] | Vazquez-Bravo B, Gonçalves K, Shisler J L, et al. Adenovirus replication cycle disruption from exposure to polychromatic ultraviolet irradiation[J]. Environmental Science & Technology, 2018, 52(6): 3652-3659. |
| [18] | Buonanno M, Welch D, Shuryak I, et al. Far-UVC light (222 nm) efficiently and safely inactivates airborne human coronaviruses[J]. Scientific Reports, 2020, 10(1). DOI:10.1038/s41598-020-67211-2 |
| [19] | Sliney D H, Stuck B E. A need to revise human exposure limits for ultraviolet UV-C radiation[J]. Photochemistry and Photobiology, 2021, 97(3): 485-492. DOI:10.1111/php.13402 |
| [20] | Matafonova G, Batoev V. Dual-wavelength light radiation for synergistic water disinfection[J]. Science of the Total Environment, 2022, 806. DOI:10.1016/j.scitotenv.2021.151233 |
| [21] | Keil J N, Kätker H, Wegh R T, et al. Novel bandpass filter for far UV-C emitting radiation sources[J]. Optical Materials, 2023, 140. DOI:10.1016/j.optmat.2023.113866 |
| [22] | Li T, Zhang Y Z, Gan J M, et al. Superiority of UV222 radiation by in situ aquatic electrode KrCl excimer in disinfecting waterborne pathogens: mechanism and efficacy[J]. Journal of Hazardous Materials, 2023, 452. DOI:10.1016/j.jhazmat.2023.131292 |
| [23] | Kang J W, Kang D H. The synergistic bactericidal mechanism of simultaneous treatment with a 222-nanometer krypton-chlorine excilamp and a 254-nanometer low-pressure mercury lamp[J]. Applied and Environmental Microbiology, 2019, 85(1). DOI:10.1128/aem.01952-18 |
| [24] | Hull N M, Linden K G. Synergy of MS2 disinfection by sequential exposure to tailored UV wavelengths[J]. Water Research, 2018, 143: 292-300. DOI:10.1016/j.watres.2018.06.017 |
| [25] | Qiao Y, Chen D Y, Wen D Y. Use of coupled wavelength ultraviolet light-emitting diodes for inactivation of bacteria in subsea oil-field injection water[J]. Science of the Total Environment, 2018, 640⁃641: 757-763. |
| [26] | Li M K, Qiang Z M, Li T G, et al. In situ measurement of UV fluence rate distribution by use of a micro fluorescent silica detector[J]. Environmental Science & Technology, 2011, 45(7): 3034-3039. |
| [27] | Song K, Mohseni M, Taghipour F. Mechanisms investigation on bacterial inactivation through combinations of UV wavelengths[J]. Water Research, 2019, 163. DOI:10.1016/j.watres.2019.114875 |
| [28] |
陆海, 李雪琪, 冯伟豪. 复活光强对大肠杆菌光复活影响的研究进展[J]. 长春工程学院学报(自然科学版), 2021, 22(3): 99-103. Lu H, Li X Q, Feng W H. The research progress on the influence of light resurrection by photoractivation light Intensity to Escherichia coli[J]. Journal of Changchun Institute of Technology (Natural Sciences Edition), 2021, 22(3): 99-103. |
| [29] | Jing Z B, Wang W L, Nong Y J, et al. Suppression of photoreactivation of E. coli by excimer far-UV light (222 nm) via damage to multiple targets[J]. Water Research, 2024, 255. DOI:10.1016/j.watres.2024.121533 |
| [30] | Cheng L H, Wei X H, Gao A L, et al. Performance and mechanism of sequential UV-NaClO disinfection: inactivation and reactivation of antibiotic-resistant bacteria, disinfection byproduct formation and microbial community variation[J]. Journal of Water Process Engineering, 2024, 58. DOI:10.1016/j.jwpe.2024.104824 |
| [31] | Hijnen W A M, Beerendonk E F, Medema G J. Inactivation credit of UV radiation for viruses, bacteria and protozoan (oo)cysts in water: a review[J]. Water Research, 2006, 40(1): 3-22. |
| [32] | Sanz E N, Dávila I S, Balao J A A, et al. Modelling of reactivation after UV disinfection: effect of UV-C dose on subsequent photoreactivation and dark repair[J]. Water Research, 2007, 41(14): 3141-3151. |
| [33] | Li G Q, Wang W L, Huo Z Y, et al. Comparison of UV-LED and low pressure UV for water disinfection: photoreactivation and dark repair of Escherichia coli[J]. Water Research, 2017, 126: 134-143. |
| [34] | Wang D, Oppenländer T, El-Din M G, et al. Comparison of the disinfection effects of vacuum-UV (VUV) and UV light on Bacillus subtilis spores in aqueous suspensions at 172, 222 and 254 nm[J]. Photochemistry and Photobiology, 2010, 86(1): 176-181. |
| [35] | Ha J W, Kang D H. Effect of intermittent 222 nm krypton-chlorine excilamp irradiation on microbial inactivation in water[J]. Food Control, 2018, 90: 146-151. |
| [36] |
竹涛, 付顺江, 谢蔚, 等. 短波紫外线的消杀机制与影响因素[J]. 中国激光, 2023, 50(9): 169-180. Zhu T, Fu S J, Xie W, et al. UVC sterilization mechanism and influencing factors[J]. Chinese Journal of Lasers, 2023, 50(9): 169-180. |
| [37] |
王佳乐, 李梦凯, 孙喆, 等. 生物剂量法与模型模拟法测定紫外消毒器剂量的对比[J]. 中国给水排水, 2023, 39(9): 33-38. Wang J L, Li M K, Sun Z, et al. Comparison between biodosimetry and model simulation for determining fluence of UV disinfector[J]. China Water & Wastewater, 2023, 39(9): 33-38. |
| [38] | Bolton J R, Stefan M I, Shaw P S, et al. Determination of the quantum yields of the potassium ferrioxalate and potassium iodide–iodate actinometers and a method for the calibration of radiometer detectors[J]. Journal of Photochemistry and Photobiology A: Chemistry, 2011, 222(1): 166-169. |
| [39] | Wang Y Y, Yin R, Tang Z Y, et al. Reactive nitrogen species mediated inactivation of pathogenic microorganisms during UVA photolysis of nitrite at surface water levels[J]. Environmental Science & Technology, 2022, 56(17): 12542-12552. |
| [40] | Wojtala A, Bonora M, Malinska D, et al. Methods to monitor ROS production by fluorescence microscopy and fluorometry[J]. Methods in Enzymology, 2014, 542: 243-262. |
| [41] | Kalyanaraman B, Darley-Usmar V, Davies K J A, et al. Measuring reactive oxygen and nitrogen species with fluorescent probes: challenges and limitations[J]. Free Radical Biology and Medicine, 2012, 52(1): 1-6. |
| [42] | Wen G, Xu X Q, Zhu H, et al. Inactivation of four genera of dominant fungal spores in groundwater using UV and UV/PMS: efficiency and mechanisms[J]. Chemical Engineering Journal, 2017, 328: 619-628. |
| [43] | Quek P H, Hu J. Influence of photoreactivating light intensity and incubation temperature on photoreactivation of Escherichia coli following LP and MP UV disinfection[J]. Journal of Applied Microbiology, 2008, 105(1): 124-133. |
| [44] | Sun Z, Fu J N, Li X, et al. Using Algal Virus Paramecium bursaria chlorella virus as a human adenovirus surrogate for validation of UV treatment systems[J]. Environmental Science & Technology, 2020, 54(23): 15507-15515. |
| [45] | Li M K, Qiang Z M, Wang C, et al. Experimental assessment of photon fluence rate distributions in a medium-pressure UV photoreactor[J]. Environmental Science & Technology, 2017, 51(6): 3453-3460. |
| [46] | Ramsay I A, Niedziela J C, Ogden I D. The synergistic effect of excimer and low-pressure mercury lamps on the disinfection of flowing water[J]. Journal of Food Protection, 2000, 63(11): 1529-1533. |
| [47] | Sinha R P, Häder D P. UV-induced DNA damage and repair: a review[J]. Photochemical & Photobiological Sciences, 2002, 1(4): 225-236. |
| [48] | Nyangaresi P O, Qin Y, Chen G L, et al. Effects of single and combined UV-LEDs on inactivation and subsequent reactivation of E. coli in water disinfection[J]. Water Research, 2018, 147: 331-341. |