 2024, Vol. 45
2024, Vol. 45


2. 湖南省第二测绘院,长沙 410009
2. Second Surveying and Mapping Institute of Hunan Province, Changsha 410009, China
植被净初级生产力(net primary productivity, NPP)是指绿色植物通过光合作用在单位时间、单位面积上产生的有机物质总量扣除自养呼吸消耗后所剩余的有机物部分.作为生态系统碳循环的重要组成部分, 植被NPP除了直接表征地表植被的自然生产能力和气候变化、人类活动对陆地生态系统的影响外, 还是判定植被碳源/碳汇和生态系统固碳能力的重要指标[1 ~ 4].由于植被NPP的时空变化和空间格局受区域气候差异、地形因素和人类活动等的影响, 因此在“双碳”目标背景下, 查清区域陆地植被NPP的时空变化及其驱动因素对正确评估植被碳汇功能、科学调控区域碳平衡以及应对全球气候变化具有重要参考价值.
目前, 国内外学者从不同视角对植被NPP的时空变化及其驱动因素开展了大量的研究, 归纳起来主要集中在两个方面.一是基于野外监测和多源遥感数据对全球和不同区域尺度植被NPP的时空格局特征展开研究. 结果发现不同时空尺度上陆地植被NPP的时空变化存在明显差异[5 ~ 12].如在全球尺度上, 植被NPP呈缓慢上升趋势, 而在中国区域植被NPP则以3.13 g∙(m2∙a)-1的速率呈快速上升[13], 长江中下游地区增速则更为明显[14].二是聚焦陆地植被NPP变化的驱动因素及其对植被NPP变化的相对贡献研究. 多数结果认为, 不同区域的植被NPP因受自身生理特性差异的影响, 对气候变化、地形因子和人类活动(如土地利用变化)等的响应关系与敏感程度存在明显的空间异质性[9 ~ 18].例如Ge等[13]的研究发现, 气候变化与人类活动主导中国植被NPP变化的面积分别为60.06%和39.94%, 并且气候因子对植被NPP的影响主要起正向作用, 而人类活动则起负向影响;王原等[19]的研究发现, 人类活动对上海农田生态系统NPP的影响大于气候变化;但在广东人类活动对植被NPP的影响远小于气候变化和其它非气候因素[20].上述研究表明驱动植被NPP变化的因素多而复杂, 以致使研究结果存在分歧.
尽管国内外学者对植被NPP的研究取得了丰硕成果, 但仍存在进一步完善的空间.具体表现在:一是在时空尺度上, 现有的研究主要集中在国家层面[21]、区域尺度(如东北、西北、西南)[22]和大流域(长江流域、黄河流域和黄土高原等)[23 ~ 28]以及省域尺度[29, 30], 而对作为改善生态环境、缓解碳减排压力的城市群绿心区植被NPP展开研究的很少;再者, 已有的研究时间尺度多为年际变化, 而从月尺度探讨植被NPP变化的相对较少.二是从驱动因素看, 现有的研究主要集中在植被NPP变化与气候因子和人类活动等的相互关系上, 认为气候变化和人类活动是影响植被NPP的主要因素[31, 32], 而对地形因子对植被NPP变化的影响关注较少[33].尽管也有学者[34]探讨过大渡河流域植被NPP与地形因子之间的关系, 认为地形会通过影响气温和降水的再分配(如同一区域内海拔较高的地点的气温和降水与海拔较低的地点的气温和降水会出现截然不同的模式)间接作用于植物的光合作用和呼吸作用, 进而影响植被的固碳能力和NPP, 但他们在分析地形因子对植被NPP变化的影响时, 多数没有对海拔、坡度和坡向对植被NPP变化的影响进行定量分离.此外, 现有研究在探究气候因子对陆地植被NPP变化的影响时仅从气候因子中选取气温和降水两个指标, 而对影响光照强度和热量分布的太阳辐射考虑甚少, 这无疑会增加研究结果的不确定性.
长株潭城市群生态绿心区(长株潭绿心区)是长株潭“两型”社会综合配套改革的城市绿地保护区, 其植被NPP的变化对区域生态系统稳定和城市群经济发展具有重要影响.目前关于长株潭绿心区的研究主要集中在绿心区规划治理以及生态服务价值等方面, 而针对绿心区植被NPP时空变化及其与气候、地形因子关系的研究较少.本文基于土地利用和气象等遥感数据, 运用改进的CASA(carnegie ames stanford approach)模型估算长株潭绿心区2011~2020年逐月的植被NPP, 借助ArcGIS10.2软件和统计分析探讨植被月NPP的时空变化及其驱动因素, 以期为长株潭绿心区制定生态管理和植被增汇措施提供数据支撑.
1 研究区概况长株潭绿心区位于湖南省中部偏东北部, 地处长沙、湘潭和株洲三市的交汇核心地带(图 1), 区域范围介于112°53'~113°18'E和27°43'~28°08'N之间, 总面积约528.32 km2(长沙占58.46%, 株洲占15.76%, 湘潭占25.78%), 是目前中国最大的城市群绿心区, 也是长株潭城市群重要的生态屏障[35].绿心区属中亚热带大陆季风性湿润气候, 光热资源丰富, 年平均气温为16.9℃, 1月和7月平均气温分别为5.26℃和29.01℃, 年日照时数为1 796 h;降水充沛, 全年降雨量1 348.9 mm, 4~8月降水量约占全年降水量的60%以上.绿心区地貌类型以低山丘陵为主, 大部分地区海拔高度在4~292 m之间, 地势由中部向东西两侧逐渐降低, 坡度变化在0.27°~57.94°之间.土壤类型以红壤为主, 土层厚度约35~100 cm.绿心区生境较为复杂, 生态类型多样, 河溪、塘库、洲滩、农田、灌草和森林并存, 森林植被以常绿阔叶林、落叶阔叶林、针阔混交林和针叶林为主, 樟树、枫香、马尾松和杉木等树种所占比例较高, 其复杂的自然生境和多样的植被类型形成了中亚热带独具一格的植被景观格局.

|
图 1 长株潭绿心区位置及样点分布示意 Fig. 1 Location and distribution of sampling sites in the Chang-Zhu-Tan Green Heart area |
估算植被月NPP及其影响因素分析所需的数据包括2011~2020年的土地利用数据、气象数据、NDVI数据和用于提取海拔、坡度以及坡向的DEM数据.其中, 土地利用数据来自中国科学院资源环境科学与数据中心(https://www.resdc.cn/), 空间分辨率为30 m, 参考中国土地利用遥感监测数据集(CNLUCC)的分类方法, 将土地利用数据重分类得到耕地、林地、草地、水域、建设用地和未利用地这6种土地利用类型;气象数据来自国家气象科学数据网(http://data.cma.cn), 主要包括逐月气温、降水和太阳辐射等.由于长株潭绿心区气象站点较少, 本文基于克里金插值法对气象站点数据进行空间插值, 得到空间分辨率为30 m的栅格数据;NDVI数据和DEM数据均来自地理空间数据云平台(https://www.gscloud.cn/), 空间分辨率为30 m, NDVI数据通过Landsat8 OLI_TIRS遥感影像获取.此外, 用于验证CASA模型估算精度的NPP数据来自美国国家航空航天局(NASA)公布的MYD17A3数据集(https://search.earthdata.nasa.gov/), 空间分辨率为500 m, 利用ArcGIS10.2软件对原始数据进行镶嵌、投影变换和掩膜处理后, 重采样得到空间分辨率为30 m的长株潭绿心区植被NPP数据, 同时将坐标系统一为WGS84.
2.2 研究方法 2.2.1 植被NPP估算在区域尺度上对NPP实施全覆盖监测显然不现实, 本文选取广泛用于区域尺度的改进的CASA模型[21], 借助ENVI5.6软件通过代入月均气温、月降水量、月总太阳辐射和不同植被的NDVI数据估算绿心区2011~2020年各月的NPP, 具体计算公式如式(1).

|
(1) |
式中, ε(x, t)表示像元x在t月植被的实际光能利用率(以C计, g∙MJ-1);APAR(x, t)表示像元x在t月植被吸收的光合有效辐射[以C计, g∙(m2∙month)-1], 由像元x在t月内接收的太阳总辐射SOL[MJ∙(m2∙month)-1]和植被对太阳入射光合有效辐射吸收的比例(FPAR)以及光合有效辐射与太阳总辐射之比决定, 通常用式(2)表示.
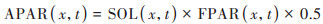
|
(2) |
由于FPAR(x, t)与植被归一化指数NDVI存在线性关系, 于是FPAR(x, t)可用式(3)进行计算.
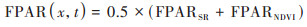
|
(3) |
式中, FPARSR和FPARNDVI分别用式(4)和式(5)进行计算.
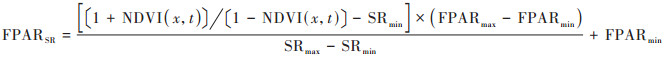
|
(4) |
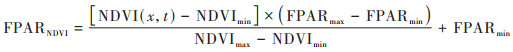
|
(5) |
式中, NDVI(x, t)表示像元x在t月的归一化植被指数;NDVImax和NDVImin表示第i种植被类型NDVI的最大值和最小值, 分别取概率分布为95%和5%所对应的值;FPARmax和FPARmin表示与植被类型无关的最大和最小光合有效辐射比例, 分别取0.95和0.001;SRmax和SRmin表示第i种植被类型NDVI简单比率的最大值和最小值[36], 同样取概率分布为95%和5%时所对应的值.
实际光能利用率ε(x, t)主要受气温、降水和植被类型的影响, 本文采用植被最大光能利用率、水分胁迫因子和气温胁迫因子的乘积利用式(6)对其进行计算.
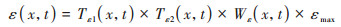
|
(6) |
式中, Tε1(x,t)和Tε2(x,t)分别表示低温和高温对植被光能利用率的胁迫影响;Wε(x,t)表示水分条件对植被光能利用率的胁迫作用, 通常通过降水和蒸散影响植被的光能利用率, 取值范围为0.5(极端干旱)~1(极端湿润)之间;εmax表示最大光能利用率, 其取值与植被类型有关, 参考Zhu等[36]的研究成果, 得到长株潭绿心区不同植被类型的最大光能利用率(表 1);Tε1(x, t)、Tε2(x, t)和Wε(x, t)采用式(7)~(9)进行计算.
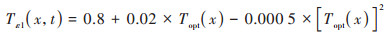
|
(7) |
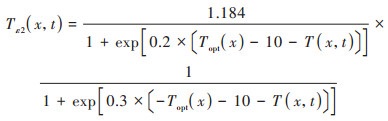
|
(8) |
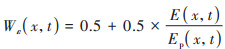
|
(9) |
|
|
表 1 长株潭绿心区不同植被类型的最大光能利用率 Table 1 Maximum light utilization efficiency of different vegetation types in the Chang-Zhu-Tan Green Heart area |
式中, Topt(x)表示植被生长的最适气温, 即一年内某一区域NDVI达到最大时的月平均气温;T(x, t)表示月平均气温, 当T(x, t)≤ -10℃时, 植被光能利用率为0, 即Tε1(x,t) = 0;当T(x, t)比Topt(x)高10℃或低13℃时, Tε2(x,t)等于Topt(x)时Tε2(x,t)值的一半;E(x,t)和Ep(x,t)分别表示研究区的实际蒸散量(mm)和潜在蒸散量(mm), 由周广胜等[37]的区域蒸散模型求得.
2.2.2 植被NPP估算精度验证验证CASA模型估算植被NPP的精度一直是遥感反演的难题.本文利用2020年的实测数据和MYD17A3NPP数据检验CASA模型估算绿心区植被NPP的可靠性.首先利用2020年7~8月在研究区设置的大小为30 m×30 m的30个样方(图 1), 采用对角线法得到地上和地下生物量(地下生物量采用根冠比进行计算), 再根据生物量与NPP的关系式NPP = B/(cT+dB)(式中, B表示单位面积上的生物量;T表示植被年龄;c和d表示对应的植被类型常数)得到实测NPP[38];然后根据GPS实测样方的地理位置提取相应CASA模型估算的NPP值, 进而进行精度验证.图 2(a)显示, 基于改进的CASA模型估算的NPP与实测NPP呈极显著正相关, 平均绝对误差为19.68 g∙(m2∙a)-1, 平均相对误差为8.72%, 表明利用改进的CASA模型估算长株潭绿心区植被NPP具有可靠性.图 2(b)显示, 基于CASA模型估算的NPP与MYD17A3产品的NPP也较为吻合, 平均相对误差为14.20%, R 2为0.779 4, 再次表明使用CASA模型估算植被NPP是可行的.

|
图 2 CASA模型估算的NPP值与实测值和MODIS NPP产品的比较 Fig. 2 Comparison of NPP values estimated by CASA model with measured values and MODIS NPP product |
基于估算的逐月NPP, 采用线性回归模型式(10)分析植被月NPP的年内变化及其与相应气候因子(月均气温、月降水量和月太阳辐射)的关系;同时随机选取研究区30个像元2011~2020年的月平均NPP, 探讨相应地形因子(海拔、坡度和坡向)对植被月NPP的影响.
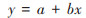
|
(10) |
式中, y表示植被月NPP或月平均NPP, x表示气候和地形因子或月份, a和b表示拟合参数.
另外, 基于偏相关分析以2011~2020年研究区栅格像元各月的平均NPP作为响应变量, 以同时期相应栅格各月的平均气候因子值和地形因子值作为解释变量, 在R-Studio软件中分别计算植被月NPP与气候因子和地形因子的相关系数, 然后利用terra模块和ppcor模块求取植被月NPP与气候因子和地形因子的偏相关系数.相关系数和一阶偏相关系数的计算公式分别见式(11)和式(12).
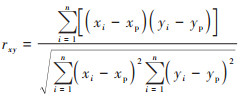
|
(11) |
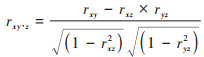
|
(12) |
式中, rxy、rxz和ryz分别表示变量x与y、x与z以及变量y与z之间的相关系数(当探讨植被月NPP与气候因子的一阶偏相关时, y和z表示气温、降水和太阳辐射中的任意两个变量;当探讨NPP与地形因子的一阶偏相关时, y和z则表示海拔、坡度和坡向中的任意两个变量);rxy∙z表示固定气候因子或地形因子中的变量z后变量x与变量y之间的一阶偏相关系数;xi表示像元i各月的平均NPP, xp表示所有像元在相应月份NPP的平均值;yi表示像元i相应月份的平均气候因子或地形因子值, yp表示所有像元在相应月份变量y的平均值;n为像元数.植被月NPP与气候因子或地形因子的二阶偏相关系数可通过一阶偏相关系数利用式(13)进行计算.
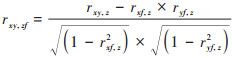
|
(13) |
式中,rxy,zf表示固定气候因子或地形因子中的任意两个变量后植被月NPP与气候或地形因子中剩余变量y之间的二阶偏相关系数;rxf,z、ryf,z与rxy∙z的定义相似.
采用t检验对偏相关分析结果在α = 0.05水平下进行显著性检验. 根据偏相关系数和显著性水平将植被NPP与气候因子和地形因子的相关关系划分为高度正(负)相关(|r| ≥ 0.8,P<0.01)、显著正(负)相关(0.5 ≤ |r|<0.8,0.01 ≤ P<0.05)和不显著正(负)相关(|r|<0.5,P ≥ 0.05).
3 结果与讨论 3.1 长株潭绿心区植被月NPP的时空变化 3.1.1 时间变化特征对长株潭绿心区2011~2020年的月平均NPP进行时间序列分析, 发现10 a间植被的月平均NPP总体呈先增加后减少的单峰型变化规律[图 3(a)].其中, 4~9月植被的NPP整体处于高位, 月值均高于全年平均值(以C计, 下同)28.93 g∙(m2∙month)-1, 这6个月的NPP约占全年总量的74.77%. 分时段看, 1~7月NPP表现出极显著的上升趋势, 在7月达到最大值[(50.40±9.81)g∙m-²], 1月出现最小值[(7.47±3.29)g∙m-²], 此时段NPP的月均上升速率为5.75g∙(m²∙month)-1;而在随后的8~12月, 植被月NPP呈极显著的下降趋势, 月均下降速率为2.64 g∙(m²∙month)-1.绿心区植被月NPP的这种变化特征与该区气候因子的年内变化[图 3(b)]以及植物生长规律相吻合.据报道[39], 亚热带山地植物生长季节通常呈抛物线变化, 即从4月开始进入生长季, 其地上生物量和净初级生产力逐渐增加, 至夏季末达到最大值, 此后下降.由于长株潭绿心区属亚热带季风气候, 雨热同期, 本研究期间1~7月太阳辐射、气温和降水均呈上升趋势(降水在1~6月呈上升趋势), 春夏季充足的水热条件通过调节植物新陈代谢有效促进了植物生长, 从而导致植被NPP在此阶段不断增加;而自7月后, 绿心区月均气温、太阳辐射和月均降水均呈减少趋势, 较差的水热条件使植被生长受到限制, 进而导致绿心区植被NPP出现下降.

|
数据为2011~2020年相应月份的平均值±标准偏差 图 3 长株潭绿心区植被月NPP与气候因子的月变化 Fig. 3 Monthly changes in vegetation NPP and climate factors in the Chang-Zhu-Tan Green Heart area |
长株潭绿心区2011~2020年各月平均植被NPP的空间分布和各月平均NPP的最大值、最小值以及空间变异系数(c.v.)如图 4所示.从中可知, 绿心区月平均NPP的变化范围为0.04~56.48 g∙(m²∙month)-1, 各月NPP的分布整体上表现出较大的空间差异性, 变异系数的变化范围为34.52%(11月)~48.76%(5月), 即在1~5月变异系数呈波动上升, 而在6~12月则呈波动下降.其中, 变异系数小于40.0%的月份有2、3、10、11和12月, 说明在以上月份NPP的空间分布差异较小, 即NPP的变化相对稳定.变异系数大于40.0%的月份有1月和4~9月, 表明在这7个月份植被NPP的空间分布差异较大, NPP的变化易受自然或人为因素的干扰, 植被NPP的稳定性较低.从空间区域看, 各月的平均植被NPP的高值区主要集中在绿心区的中部和东部等地, 低值区则主要分布在绿心区西部的湘江沿岸以及南部的狭长地带, 总体上绿心区各月的平均NPP呈现出“中东部高、西南部低”的空间分布格局.究其原因, 可能与长株潭生态绿心区覆盖度呈中东部高、西南部低的空间格局有关[40].这是因为长株潭绿心区的中部和东部属于绿心区的生态核心地带, 拥有昭山、东风水库郊野公园、石燕湖森林公园和嵩山寺植物园以及九郎山森林公园等大型生态斑块, 植被类型以常绿落叶阔叶混交林为主, 因而植被覆盖度较高, 加之在亚热带植物生长季(4~8月)水热资源丰富且雨热同期, 同时绿心区中东部因海拔相对较高, 人类活动对植被的扰动较少, 故而植被生长旺盛, 固碳能力较强, 从而NPP积累较多, 进而导致各月的平均NPP在此区域出现高值.如绿心区2011~2020年的8月平均NPP的最高值(56.48 g∙m-²)就出现在中部的东风水库郊野公园和石燕湖森林公园, 次高值则出现在九郎山森林公园和五云峰森林公园.而在绿心区西部的湘江沿岸和南部的狭长地带, 这里地势低平, 人类活动范围和干扰强度较大[41], 特别是城镇建设、产业分布等大量侵占林草和耕地, 加上京广铁路和城际铁路等交通干道从该区经过, 从而使湘江沿岸和南部狭长地带林草覆盖度低, 进而使各月的NPP在此区域出现低值, 如8月平均NPP的最低值(7.20 g∙m-²)就出现在绿心区西部的湘江沿线附近.

|
max、min和mean分别表示最大值、最小值和平均值, 单位均为g∙m-2 图 4 2011~2020年长株潭绿心区各月平均NPP的空间分布 Fig. 4 Spatial distribution of monthly average NPP in the Chang-Zhu-Tan Green Heart area from 2011 to 2020 |
图 5显示, 绿心区2011~2020年月NPP与月太阳总辐射量、月均气温和月降水量呈极显著或显著的线性关系, 相关系数分别为0.879、0.872和0.257, 各气候因子对月NPP的解释力从大到小依次为月太阳总辐射、月均气温和月降水量, 说明绿心区植被NPP在月尺度变化上, 月太阳总辐射和月均气温是主要控制因子.这与潘竟虎等[10]对疏勒河流域植被NPP与气候因子关系研究的结果类似, 即在疏勒河流域植被月NPP受当月气温的影响要大于受当月降水的影响, 不同的是他们没有探讨太阳辐射对植被月NPP的作用.

|
数据为2011~2020年相应月份的NPP值 图 5 长株潭绿心区植被月NPP与气候因子的回归关系 Fig. 5 Regression relationship between monthly vegetation NPP and climate factors in the Chang-Zhu-Tan Green Heart area |
为了区分气温、太阳辐射和降水对植被月NPP变化的影响, 利用偏相关分析和显著性检验得到绿心区栅格像元植被月NPP与气候因子的偏相关显著性空间分布(图 6).总体上, 绿心区植被月NPP与气候因子的偏相关显著程度具有明显的空间分异性, 正的偏相关性均表现为“中部高、四周低”由中心向四周减弱的态势, 其中植被月NPP与月太阳总辐射的偏相关性最强, 气温次之, 而与月降水量呈正的偏相关性较弱.具体而言, 绿心区植被月NPP与月总太阳辐射的偏相关系数变化在-0.996 ~ 0.975之间, 平均偏相关系数为0.447.月NPP与月总太阳辐射呈正相关的面积占绿心区总面积的90.21%, 其中, 显著正相关(P < 0.05)和高度正相关(P < 0.01)的地区高达203.71 km2, 且以绿心区中部为中心呈片状广泛分布, 而显著负相关(P < 0.05)和高度负相关(P < 0.01)的地区不足1.0 km2, 主要呈点状分布在同升片区和跳马片区的交界处[图 6(a)].这说明绿心区植被月NPP对太阳辐射表现出高度的正敏感性, 月太阳总辐射是控制植被生长的关键气候因子, 这与前述的植被月NPP与月太阳辐射呈极显著相关性的结果一致.图 6(b)显示, 绿心区植被月NPP与月均气温呈正相关的面积明显要小于呈负相关的面积, 消除降水和太阳辐射影响后的偏相关系数变化在-0.878 ~ 0.874之间, 平均偏相关系数为0.349.其中呈显著正相关(P < 0.05)和高度正相关(P < 0.01)的区域占研究区总面积的25.85%, 主要分布在绿心区的昭山片区、暮云片区、东部的云龙片区和荷塘片区以及柏加-镇头片区的交界部位, 说明以上区域植被NPP受气温的影响较大, 气温是其主要影响因素;显著负相关(P < 0.05)和高度负相关(P < 0.01)的区域面积占14.43%, 主要分布在跳马片区以及其它片区的边缘, 说明随着月均气温的增加, 以上区域的植被月NPP呈下降趋势.原因是在各片区的边缘植被分布零散且较少, 随着气温的增加植物呼吸作用增强, 导致植被固碳能力下降, 从而使NPP表现为降低.相较于月均气温, 植被月NPP与月降水量之间的偏相关系数变化在-0.807 ~ 0.607之间, 平均偏相关系数为0.042, 植被月均NPP与降水量整体呈正相关趋势, 且呈正相关的面积稍大于呈负相关的面积[图 6(c)].其中, 显著正相关(P < 0.05)和高度正相关(P < 0.01)的区域占2.45%, 主要分布在绿心区的昭山片区和跳马片区的交界附近以及云龙片区与柏加-镇头片区的交界部位.此外, 有5.46%的区域呈显著负相关(P < 0.05)和高度负相关(P < 0.01), 主要集中在绿心区湘江沿线附近、同升片区以及南部的天元片区和易俗河片区, 说明以上区域月均NPP对降水量的变化具有一定的负敏感性, 其原因可能是上述区域地势较低, 丰沛的降水在一定程度上对植被的生长产生了一定的胁迫作用[12, 42].

|
图 6 2011~2020年长株潭绿心区月平均NPP与同期气候因子的偏相关空间分布 Fig. 6 Spatial distribution of the partial correlations between monthly average NPP and climate factors in the Chang-Zhu-Tan Green Heart area from 2011 to 2020 |
地形在一定程度上影响物质循环和能量分配, 进而影响植被的生长和分布, 最终影响植被的NPP.图 7是基于栅格像元利用偏相关分析和显著性检验得到的植被月NPP与地形因子的偏相关空间分布结果. 总体而言, 绿心区植被月NPP与地形因子的偏相关分布也存在明显的空间异质性, 显著或高度正相关的区域与显著或高度负相关的区域大多呈零散分布, 表明地形因子对植被NPP的影响较为复杂.其中, 植被NPP与海拔和坡度呈显著或高度正相关的区域要明显小于植被NPP与坡向呈显著或高度正相关的区域, 说明在长株潭绿心区地形因子中的坡向对植被月NPP空间分布的正向影响要大于海拔和坡度, 意味着坡向对植被NPP的解释能力要大于坡度和海拔.具体而言, 植被月NPP与海拔的偏相关系数变化在-0.996 ~ 0.997之间, 平均值为-0.119.其中, 显著正相关(P < 0.05)和高度正相关(P < 0.01)的区域只占绿心区总面积的1.64%, 且呈点状零星分布;另有12.63%的区域呈显著负相关(P < 0.05)和高度负相关(P < 0.01), 主要呈带状、块状分布在绿心区中部、北部和东部各片区的交界边缘[图 7(a)].植被月NPP与坡度的偏相关系数区间为-0.986 ~ 0.992, 平均值为-0.146.其中, 植被月NPP与坡度呈显著正相关和高度正相关的区域占全区总面积的2.73%, 亦呈零散分布;呈显著负相关和高度负相关的区域约占16.37%, 分布范围与植被月NPP和海拔呈显著或高度负相关的区域大体相似, 以条带状和块状分布为主[图 7(b)].绿心区植被月NPP与坡向的偏相关系数变化在-0.864 ~ 0.996之间, 平均偏相关系数为0.148, 显著正相关(P < 0.05)和高度正相关(P < 0.01)的面积约占研究区总面积的6.42%, 并且呈点、块状分散分布;呈显著负相关(P < 0.05)和高度负相关(P < 0.01)的区域主要分布在绿心区跳马片区的西北部和东北部以及柏加-镇头片区的北部, 但面积不到研究区总面积的0.42%, 说明在月尺度上绿心区植被NPP的空间分布受坡向的负向影响较小[图 7(c)].

|
图 7 2011~2020年长株潭绿心区月平均NPP与地形因子的偏相关空间分布 Fig. 7 Spatial distribution of the partial correlations between monthly average NPP and terrain factors in the Chang-Zhu-Tan Green Heart area from 2011 to 2020 |
尽管研究区30个像元的月均NPP与地形因子均存在显著回归关系(图 8), 但对比植被月NPP与气候因子和地形因子的回归关系与偏相关分布, 发现气候因子对绿心区植被月NPP的影响程度明显要大于地形因子.这是因为气温、太阳辐射和降水决定了水热和能量的总体输入, 是控制植被生长的主导自然因素, 而地形因子在一定程度上仅对水热和能量产生再分配作用[43, 44].其次, 长株潭绿心区最高海拔只有292 m, 地表高程起伏不大, 地势相对较平坦, 因而较低的海拔和狭窄的坡度范围不足以通过影响气温和降水进而对植被NPP产生显著作用.另外, 本文得到的地形因子对植被月NPP的影响与前人得到的结果不完全吻合.如黄兰鹰等[34]对大渡河流域植被NPP与地形因子的关系研究发现, 随海拔和坡度增加植被NPP分别表现为先上升后降低和显著增加的趋势, 但对坡向变化不敏感;而本文的结果显示, 坡向对绿心区植被月NPP的影响大于海拔和坡度.造成上述差异的原因可能与二者研究区的地形特征不同有关.前者的研究区域属于川西高山峡谷地带, 海拔高度超过5 000 m, 地表高低起伏较大, 加之大渡河流域属于干旱半干旱河谷分布区, 自北向南由高原气候逐渐过渡到亚热带湿润气候;而本研究区的最大海拔仅有292 m, 且以亚热带低山丘陵为主.前者较大的海拔高差和地形起伏有可能掩盖或削弱坡向对植被NPP的影响, 加之特殊气候因子的叠加作用最终有可能使坡向对植被NPP的影响不显著[33].

|
图 8 2011~2020年长株潭绿心区栅格像元月平均NPP与地形因子的回归关系 Fig. 8 Regression relationship between monthly average NPP and terrain factors of grid pixels in the Chang-Zhu-Tan Green Heart area from 2011 to 2020 |
值得指出的是, 在不考虑人类活动影响的情况下, 植被NPP受气候和地形等自然因素的综合影响.而本文在估算植被NPP时仅考虑了植被指数和气候因素, 在使用气象数据进行插值时并未考虑地形因素对气温、降水和太阳辐射的再分配作用, 因而利用改进的CASA模型得到的结果必然与实际情况存在偏差.因此, 在后续研究中如何将地形因子、气候因子以及人类活动纳入同一框架体系探讨它们对植被NPP的耦合影响将是下一步努力的方向.
4 结论(1)时间上, 长株潭绿心区2011~2020年平均植被月NPP总体呈先增加后下降的趋势, 最高、最低值分别出现在7月和1月.其中1~7月植被NPP月均增速为5.75 g∙(m²∙month)-1, 8~12月植被NPP月均降速为2.64 g∙(m²∙month)-1;植被月NPP的变化与气候因子的年内变化和植被生长规律一致.
(2)空间上, 长株潭绿心区各月的NPP具有明显的空间差异性, 变异系数在1~5月呈上升趋势, 6~12月呈下降趋势.各月NPP的高值出现在绿心区中部和东部的生态核心带, 低值集中分布在绿心区西部的湘江沿岸以及南部的狭长地带, 植被NPP“中东部高、西南部低”的分布特征与植被盖度分布格局和人为活动有关.
(3)气候与地形因子对绿心区植被月NPP的影响存在正向促进与负向抑制作用, 但总体上均呈正相关关系且空间差异明显.绿心区植被月NPP与月太阳总辐射的偏相关性最强, 月均气温次之, 月降水量的偏相关性最小;正的偏相关性由中心向四周减弱, 太阳辐射是植被NPP变化的主控因子.地形因子对绿心区植被NPP的影响相对较弱, 显著或高度正相关的区域与负相关的区域大多呈零散分布, 坡向对植被NPP的正向影响大于海拔和坡度.
| [1] | Huang M, Wang Z S, Wang S Q, et al. Global vegetation productivity responses to the West Pacific Warm Pool[J]. Science of the Total Environment, 2019, 655: 641-651. DOI:10.1016/j.scitotenv.2018.11.170 |
| [2] | Michaletz S T, Cheng D L, Kerkhoff A J, et al. Convergence of terrestrial plant production across global climate gradients[J]. Nature, 2014, 512(7512): 39-43. DOI:10.1038/nature13470 |
| [3] |
刘军会, 高吉喜, 王文杰, 等. 我国典型陆地生态系统固碳重要区范围界定[J]. 环境科学研究, 2016, 29(12): 1782-1789. Liu J H, Gao J X, Wang W J, et al. Delimiting important carbon sequestration areas for typical terrestrial ecosystems in China[J]. Research of Environmental Sciences, 2016, 29(12): 1782-1789. |
| [4] | Wei X D, Yang J, Luo P P, et al. Assessment of the variation and influencing factors of vegetation NPP and carbon sink capacity under different natural conditions[J]. Ecological Indicators, 2022, 138. DOI:10.1016/j.ecolind.2022.108834 |
| [5] |
苏胜涛, 曾源, 赵旦, 等. 中国陆地植被净初级生产力估算模型优化与分析——基于中国生态系统研究网络数据[J]. 生态学报, 2022, 42(4): 1276-1289. Su S T, Zeng Y, Zhao D, et al. Optimization of net primary productivity estimation model for terrestrial vegetation in China based on CERN data[J]. Acta Ecologica Sinica, 2022, 42(4): 1276-1289. |
| [6] | Schuur E A G, Bockheim J, Canadell J G, et al. Vulnerability of permafrost carbon to climate change: implications for the global carbon cycle[J]. Bioscience, 2008, 58(8): 701-714. DOI:10.1641/B580807 |
| [7] |
刘凤, 曾永年. 2000—2015年青海高原植被碳源/汇时空格局及变化[J]. 生态学报, 2021, 41(14): 5792-5803. Liu F, Zeng Y N. Analysis of the spatio-temporal variation of vegetation carbon source/sink in Qinghai Plateau from 2000—2015[J]. Acta Ecologica Sinica, 2021, 41(14): 5792-5803. |
| [8] |
朴世龙, 何悦, 王旭辉, 等. 中国陆地生态系统碳汇估算: 方法、进展、展望[J]. 中国科学: 地球科学, 2022, 65(6): 1010-1020. Piao S L, He Y, Wang X H, et al. Estimation of China's terrestrial ecosystem carbon sink: methods, progress and prospects[J]. Science China Earth Sciences, 2022, 65(4): 641-651. |
| [9] |
徐勇, 黄海艳, 戴强玉, 等. 西南地区陆地植被生态系统NPP时空演变及驱动力分析[J]. 环境科学, 2023, 44(5): 2704-2714. Xu Y, Huang H Y, Dai Q Y, et al. Spatial-temporal variation in net primary productivity in terrestrial vegetation ecosystems and its driving forces in Southwest China[J]. Environmental Science, 2023, 44(5): 2704-2714. |
| [10] |
潘竟虎, 黄克军, 李真. 2001—2010年疏勒河流域植被净初级生产力时空变化及其与气候因子的关系[J]. 生态学报, 2017, 37(6): 1888-1899. Pan J H, Huang K J, Li Z. Spatio-temporal variation in vegetation net primary productivity and its relationship with climatic factors in the Shule River basin from 2001 to 2010[J]. Acta Ecologica Sinica, 2017, 37(6): 1888-1899. |
| [11] |
徐勇, 黄雯婷, 郑志威, 等. 基于空间尺度效应的西南地区植被NPP影响因子探测[J]. 环境科学, 2023, 44(2): 900-911. Xu Y, Huang W T, Zheng Z W, et al. Detecting influencing factor of vegetation NPP in Southwest China based on spatial scale effect[J]. Environmental Science, 2023, 44(2): 900-911. |
| [12] |
闫妍, 覃金华, 房磊, 等. 湖南省植被净初级生产力时空动态及其与气候因素的关系[J]. 生态学杂志, 2022, 41(8): 1535-1544. Yan Y, Qin J H, Fang L, et al. Spatiotemporal dynamics of vegetation net primary productivity and its relationships with climatic factors in Hunan Province[J]. Chinese Journal of Ecology, 2022, 41(8): 1535-1544. |
| [13] | Ge W Y, Deng L Q, Wang F, et al. Quantifying the contributions of human activities and climate change to vegetation net primary productivity dynamics in China from 2001 to 2016[J]. Science of the Total Environment, 2021, 773. DOI:10.1016/j.scitotenv.2021.145648 |
| [14] | Yang H F, Zhong X N, Deng S Q, et al. Assessment of the impact of LUCC on NPP and its influencing factors in the Yangtze River basin, China[J]. Catena, 2021, 206. DOI:10.1016/j.catena.2021.105542 |
| [15] |
周妍妍, 朱敏翔, 郭晓娟, 等. 疏勒河流域气候变化和人类活动对植被NPP的相对影响评价[J]. 生态学报, 2019, 39(14): 5127-5137. Zhou Y Y, Zhu M X, Guo X J, et al. Relative effects of climate change and human activities on net primary productivity in Shule River Basin[J]. Acta Ecologica Sinica, 2019, 39(14): 5127-5137. |
| [16] | Dai E F, Huang Y, Wu Z, et al. Analysis of spatio-temporal features of a carbon source/sink and its relationship to climatic factors in the Inner Mongolia grassland ecosystem[J]. Journal of Geographical Sciences, 2016, 26(3): 297-312. |
| [17] | Arain M A, Xu B, Brodeur J J, et al. Heat and drought impact on carbon exchange in an age-sequence of temperate pine forests[J]. Ecological Processes, 2022, 11(1). DOI:10.1186/s13717-021-00349-7 |
| [18] | Wang M M, Zhao J, Wang S Q, et al. Detection and attribution of positive net ecosystem productivity extremes in China's terrestrial ecosystems during 2000-2016[J]. Ecological Indicators, 2021, 132. DOI:10.1016/j.ecolind.2021.108323 |
| [19] |
王原, 黄玫, 王祥荣. 气候和土地利用变化对上海市农田生态系统净初级生产力的影响[J]. 环境科学学报, 2010, 30(3): 641-648. Wang Y, Huang M, Wang X R. Impacts of land use and climate change on agricultural productivity in Shanghai[J]. Acta Scientiae Circumstantiae, 2010, 30(3): 641-648. |
| [20] |
姜春, 吴志峰, 程炯, 等. 气候波动和土地覆盖变化对广东省植被净初级生产力的相对影响[J]. 热带亚热带植物学报, 2016, 24(4): 397-405. Jiang C, Wu Z F, Cheng J, et al. Relative effects of climatic fluctuation and land cover changes on net primary productivity of vegetation in Guangdong Province[J]. Journal of Tropical and Subtropical Botany, 2016, 24(4): 397-405. |
| [21] |
朱文泉, 潘耀忠, 张锦水. 中国陆地植被净初级生产力遥感估算[J]. 植物生态学报, 2007, 31(3): 413-424. Zhu W Q, Pan Y Z, Zhang J S. Estimation of net primary productivity of Chinese terrestrial vegetation based on remote sensing[J]. Journal of Plant Ecology, 2007, 31(3): 413-424. |
| [22] |
张筠, 张春华, 张安定, 等. 水热波动和土地覆盖变化对东北地区植被NPP的相对影响[J]. 生态学报, 2020, 40(21): 7733-7744. Zhang J, Zhang C H, Zhang A D, et al. Relative effects of hydrothermal fluctuation and land cover changes on vegetation net primary productivity in Northeast China[J]. Acta Ecologica Sinica, 2020, 40(21): 7733-7744. |
| [23] |
徐勇, 郑志威, 郭振东, 等. 2000~2020年长江流域植被NDVI动态变化及影响因素探测[J]. 环境科学, 2022, 43(7): 3730-3740. Xu Y, Zheng Z W, Guo Z D, et al. Dynamic variation in vegetation cover and its influencing factor detection in the Yangtze River Basin from 2000 to 2020[J]. Environmental Science, 2022, 43(7): 3730-3740. |
| [24] |
田智慧, 张丹丹, 赫晓慧, 等. 2000—2015年黄河流域植被净初级生产力时空变化特征及其驱动因子[J]. 水土保持研究, 2019, 26(2): 255-262. Tian Z H, Zhang D D, He X H, et al. Spatiotemporal variations in vegetation net primary productivity and their driving factors in Yellow River Basin from 2000 to 2015[J]. Research of Soil and Water Conservation, 2019, 26(2): 255-262. |
| [25] |
陈强, 陈云浩, 王萌杰, 等. 2001—2010年黄河流域生态系统植被净第一性生产力变化及气候因素驱动分析[J]. 应用生态学报, 2014, 25(10): 2811-2818. Chen Q, Chen Y H, Wang M J, et al. Change of vegetation net primary productivity in Yellow River watersheds from 2001 to 2010 and its climatic driving factors analysis[J]. Chinese Journal of Applied Ecology, 2014, 25(10): 2811-2818. |
| [26] |
刘铮, 杨金贵, 马理辉, 等. 黄土高原草地净初级生产力时空趋势及其驱动因素[J]. 应用生态学报, 2021, 32(1): 113-122. Liu Z, Yang J G, Ma L H, et al. Spatial-temporal trend of grassland net primary production and their driving factors in the Loess Plateau, China[J]. Chinese Journal of Applied Ecology, 2021, 32(1): 113-122. |
| [27] |
戴尔阜, 李双元, 吴卓, 等. 中国南方红壤丘陵区植被净初级生产力空间分布及其与气候因子的关系——以江西省泰和县为例[J]. 地理研究, 2015, 34(7): 1222-1234. Dai E F, Li S Y, Wu Z, et al. Spatial pattern of net primary productivity and its relationship with climatic factors in Hilly Red Soil Region of southern China: a case study in Taihe county, Jiangxi province[J]. Geographical Research, 2015, 34(7): 1222-1234. |
| [28] |
徐勇, 卢云贵, 戴强玉, 等. 气候变化和土地利用变化对长江中下游地区植被NPP变化相对贡献分析[J]. 中国环境科学, 2023, 43(9): 4988-5000. Xu Y, Lu Y G, Dai Q Y, et al. Assessment of the relative contribution of climate change and land use change on net primary productivity variation in the middle and lower reaches of the Yangtze River Basin[J]. China Environmental Science, 2023, 43(9): 4988-5000. |
| [29] |
张峰, 周广胜, 王玉辉. 基于CASA模型的内蒙古典型草原植被净初级生产力动态模拟[J]. 植物生态学报, 2008, 32(4): 786-797. Zhang F, Zhou G S, Wang Y H. Dynamics simulation of net primary productivity by a satellite data-driven casa model in Inner Mongolian typical steppe, China[J]. Journal of Plant Ecology, 2008, 32(4): 786-797. |
| [30] |
邵嘉豪, 李晶, 闫星光, 等. 基于地理探测器的山西省2000~2020年NPP时空变化特征及驱动力分析[J]. 环境科学, 2023, 44(1): 312-322. Shao J H, Li J, Yan X G, et al. Analysis of spatiotemporal variation characteristics and driving forces of NPP in Shanxi province from 2000 to 2020 based on geodetector[J]. Environmental Science, 2023, 44(1): 312-322. |
| [31] |
单振东, 刘顿, 骆汉, 等. 气候变化背景下人类活动对承德接坝区植被净初级生产力的影响[J]. 环境科学, 2023, 44(11): 6215-6225. Shan Z D, Liu D, Luo H, et al. Impacts of human activities on the net primary productivity of vegetation in Chengde's transitional region from plateau to plain in the context of climate change[J]. Environmental Science, 2023, 44(11): 6215-6225. |
| [32] |
肖晶, 饶良懿. 2001~2020年乌梁素海流域植被NPP时空变化及驱动因素分析[J]. 环境科学, 2024, 45(8): 4744-4755. Xiao J, Rao L Y. Spatiotemporal variation characteristics and driving factors of vegetation NPP in the Ulansuhai Nur Basin from 2001 to 2020[J]. Environmental Science, 2024, 45(8): 4744-4755. |
| [33] | Sundqvist M K, Sanders N J, Wardle D A. Community and ecosystem responses to elevational gradients: processes, mechanisms, and insights for global change[J]. Annual Review of Ecology, Evolution and Systematics, 2013, 44: 261-280. |
| [34] |
黄兰鹰, 杨育林, 彭涛, 等. 大渡河流域植被净初级生产力的时空变化及其地形分异特征[J]. 东北林业大学学报, 2023, 51(11): 63-67, 73. Huang L Y, Yang Y L, Peng T, et al. Spatial and temporal variation of vegetation NPP and its topographic differentiation characteristics in the Dadu River Basin[J]. Journal of Northeast Forestry University, 2023, 51(11): 63-67, 73. |
| [35] |
王瑞禛, 王光军. 长株潭城市绿心区公益林生物量遥感反演[J]. 湖南林业科技, 2020, 47(4): 68-78. Wang R Z, Wang G J. Modelling of public welfare forest biomass in city green heart area of Changsha-Zhuzhou-Xiangtan in Hunan province based on remote sensing data[J]. Hunan Forestry Science & Technology, 2020, 47(4): 68-78. |
| [36] | Zhu W Q, Pan Y Z, He H, et al. Simulation of maximum light use efficiency for some typical vegetation types in China[J]. Chinese Science Bulletin, 2006, 51(4): 457-463. |
| [37] |
周广胜, 张新时. 中国气候-植被关系初探[J]. 植物生态学报, 1996, 20(2): 113-119. Zhou G S, Zhang X S. Study on Chinese climate-vegetation relationship[J]. Acta Phytoecologica Sinica, 1996, 20(2): 113-119. |
| [38] |
王斌, 刘某承, 张彪. 基于森林资源清查资料的森林植被净生产量及其动态变化研究[J]. 林业资源管理, 2009(1): 35-43. Wang B, Liu M C, Zhang B. Dynamics of net production of Chinese forest vegetation based on forest inventory data[J]. Forest Resources Management, 2009(1): 35-43. |
| [39] |
周寿荣, 干友民, 蒲朝龙, 等. 四川盆地西部亚热带山地草地植物生物量季节动态的研究[J]. 生态学杂志, 1989, 8(4): 1-4, 12. Zhou S R, Gan Y M, Pu C L, et al. Seasonal dynamics of phyto-biomass in subtropical mountainous grassland in western Sichuan Basin[J]. Journal of Ecology, 1989, 8(4): 1-4, 12. |
| [40] |
邓梦达, 游健荣, 李家湘, 等. 长株潭城市群生态绿心地区主要植被类型的群落特征[J]. 植物生态学报, 2020, 44(12): 1296-1304. Deng M D, You J R, Li J X, et al. Community characteristics of main vegetation types in the ecological "green-core" area of Changzhutan urban cluster[J]. Chinese Journal of Plant Ecology, 2020, 44(12): 1296-1304. |
| [41] |
王本礼, 王也, 曾超群. 长株潭绿心地区碳汇用地时空演变研究[J]. 经济地理, 2023, 43(6): 176-182, 210. Wang B L, Wang Y, Zeng C Q. Spatiotemporal evolution of carbon sink land in Changsha-Zhuzhou-Xiangtan greenheart area[J]. Economic Geography, 2023, 43(6): 176-182, 210. |
| [42] |
徐勇, 郑志威, 戴强玉, 等. 顾及时滞效应的西南地区植被NPP变化归因分析[J]. 农业工程学报, 2022, 38(9): 297-305. Xu Y, Zheng Z W, Dai Q Y, et al. Attribution analysis of vegetation NPP variation in Southwest China considering time-lag effects[J]. Transactions of the Chinese Society of Agricultural Engineering, 2022, 38(9): 297-305. |
| [43] | Zhang M, Yuan N Q, Lin H, et al. Quantitative estimation of the factors impacting spatiotemporal variation in NPP in the Dongting Lake wetlands using Landsat time series data for the last two decades[J]. Ecological Indicators, 2022, 135. DOI:10.1016/j.ecolind.2022.108544 |
| [44] |
崔嵩, 贾朝阳, 郭亮, 等. 不同海拔梯度下极端气候事件对松花江流域植被NPP的影响[J]. 环境科学, 2024, 45(1): 275-286. Cui S, Jia Z Y, Guo L, et al. Impacts of extreme climate events at different altitudinal gradients on vegetation NPP in Songhua River Basin[J]. Environmental Science, 2024, 45(1): 275-286. |