 2024, Vol. 45
2024, Vol. 45

 , LIU Hui
, DONG Fang-yong
, LIU Hui
, DONG Fang-yong
 , ZOU Xi
, LI Jian-yong
, ZHU Bin
, CHEN Feng
, MA Pei-ming
, ZHAO Xian-fu
, ZHENG Zhi-wei
, TANG Hai-bin
, ZOU Xi
, LI Jian-yong
, ZHU Bin
, CHEN Feng
, MA Pei-ming
, ZHAO Xian-fu
, ZHENG Zhi-wei
, TANG Hai-bin
三江源区面积广大, 是长江、澜沧江及黄河的发源地, 被誉为“中华水塔”, 生物多样性丰富, 具有重要的生态及气候调节功能, 是我国重要的生态安全屏障区域, 也是全球生态恢复研究的热点区域. 三江源区具有典型的高原气候特征, 气温低, 风速大, 紫外线辐射强, 氧气稀薄, 昼夜温差大. 为了适应极端的恶劣环境, 三江源植物经过长期的演化, 形成了独特的生态适应机制[1, 2], 可能表现为单一适应机制, 也可能表现为综合适应策略. 即使在世界范围内来看, 三江源植物类群也是独树一帜, 异彩纷呈, 堪称高寒地区的植物博物馆. 三江源土层浅薄, 保水性差, 土壤肥力呈现“少磷、缺氮和富钾等特征[3]. 近些年来, 三江源区生态环境保护受到越来越多科学家的关注. 但大多数研究较为宏观, 多为通过遥感解译的大尺度研究, 侧重于三江源植物区系[4]、高寒草地退化机制[5]、生态服务价值评价[6]和气候变化对三江源生态系统的影响等方面的研究, 这种大尺度的研究无法分辨河岸带的优势植物类群分布和土壤生境特征, 土壤养分尤其是土壤磷形态及其含量是否显著影响着河岸带的优势植物, 还需深入研究.
河岸带是陆地生态系统和水生态系统的交互界面, 河岸带为高低水位之间的河床及高水位之上, 直至河水影响完全消失为止的地带[7]. 由于河岸带对水陆生态系统间的物质流、能量流和信息流起到廊道、过滤器和屏障的作用, 其生态系统结构和服务功能具有特异性, 成为国内外生态学和环境科学研究的热点之一[8]. 尽管很多学者对河岸带植物群落结构和退化河岸带的生态恢复及管理等方面进行了大量研究[9, 10], 但研究对象侧重于人为干扰频繁, 严重退化的河岸带, 三江源河岸带的研究还较为欠缺, 尤其是河岸带植物群落特征及其成因机制不够明晰. 主要是因为三江源地广人稀, 交通不便, 到达河边较为困难. 虽然人类活动对三江源河岸带的干扰少, 除城镇外, 大部分河段呈自然状态, 但河岸带是三江源河流生态系统的重要组成部分, 具有稳定河岸、减少土壤侵蚀和拦截面源污染进入水体等多种生态功能. 深入理解河岸带植被特征及土壤特征及其对水文变化的响应有利于三江源生物多样性保护. 因此, 本文通过实地样带调查, 揭示三江源河岸带植物群落及土壤养分的空间分布特征, 深入探讨优势植物与土壤养分、气候因子和河流地形地貌的关系, 以期为科学认知三江源河岸带植物群落特征的演变规律提供数据支撑.
1 材料与方法 1.1 植物群落调查及多样性分析根据三江源植物群落特征、土壤生境的典型性和到达河岸带调查样地的难易程度等, 于2022年8月设置14个调查样地, 其中长江源10个, 澜沧江源2个, 黄河源2个(图 1和表 1). 长江源调查样地位于楚玛尔河治多县段(楚玛尔河与京拉线交汇处, S1)、沱沱河格尔木市段(唐古拉山镇处, S2)、通天河干流治多县段(囊极巴陇处, S3)、通天河支流莫曲治多县索加乡段(S4)、支流当曲治多县段(S5)、查旦湿地治多县段(S6)、通天河支流口前曲治多县段(S7)、通天河支流聂恰曲治多县城段(S8)、通天河干流曲麻莱县段(S9)和通天河干流称多县段(直门达水文站处, S10). 澜沧江源调查样地位于扎曲杂多县段(S11)和吉曲(又称解曲)囊谦县段(S12). 黄河源调查样地位于黄河源干流玛多县段(黄河大桥处, S13)和黄河源支流格曲玛沁县段(雪山乡恰布龙特大桥处, S14).

|
图 1 河岸带植物及土壤生境调查样地位置示意 Fig. 1 Location of survey sample plots of plants and soil habitat in the riparian zone |
|
|
表 1 三江源调查样地基本信息 Table 1 Basic information of survey sample plots in the Three-River Headwaters region |
为揭示河流水文情势对河岸带植物群落的影响, 每个调查样地设置2个样带, 即从水面开始, 垂直河水流向向岸上延伸5~10 m区域设置样带A, 延伸20~25 m设置样带B, 每个样带4个植物群落调查样方(1 m×1 m), 合计调查植物群落样方112个. 现场测量每个样方内每种植物的高度和盖度. 为获得更多的物种分布信息, 同步开展植物种类踏查工作, 范围为:沿河岸带长150~200 m(与水流方向平行), 宽50~100 m(垂直河流方向), 记录植物种类. 依据《中国植物志》确定所有植物的名称、科、属及其生活型.
分析植物群落特征及物种的重要程度常采样如下公式[11, 12].

|
(1) |

|
(2) |

|
(3) |
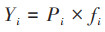
|
(4) |
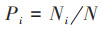
|
(5) |

|
(6) |
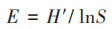
|
(7) |
式中, Yi为物种i的优势度, Ni为物种i的重要值, N为样方内所有物种重要值之和, fi为物种i在各个采样点出现的频率, H'为Shannon-Wiener多样性指数, E为Pielou均匀度指数, S为样方内的物种数.
1.2 土壤理化性质测定调查植物后, 每个样方内用手动取土钻钻取3个表层土壤样品(0~20 cm), 混合为一个样品后放入封口袋, 每个样带1个土壤样品, 合计28个. 带回实验室用玛瑙研钵研磨土壤样品, 测定土壤pH、有机质(SOM)和全氮(TN)[13].
采用改进的SMT(standard measurements testing program of european union)进行土壤各形态磷的提取, OP含量根据差减法得到[14~16]. 通过使用不同性质的提取剂HCl、NaOH提取, 将沉积物磷分为无机磷(IP)、有机磷(OP)、NaOH提取态磷(NaOH-P, 主要为铁铝结合态磷)、HCl提取态磷(HCl-P, 主要为钙镁结合态磷)及3.5 mol∙L-1 HCl提取总磷(HCl-TP). 每个提取步骤后, 离心机离心15 min, 分离固液相, 采用钼锑抗分光光度法测定提取液磷浓度.
1.3 地形地貌及气候因子数据来源样地海拔采用GPS(集思宝MG838)测定, 坡度采用测距仪(图帕斯Trupulse360)测定, 河流的河床比降、径流深、年均气温、年均降雨量和年均蒸发量等数据来源于《玉树藏族自治州水利志》[17]、《中国河湖大典(黄河卷)》[18]和国家青藏高原科学数据中心的三江源及邻近区域国家标准气象站逐月气象数据(1957~2015年)[19].
1.4 数据统计分析利用SPSS 18.0计算植物群落特征的平均值和标准差. 采用单因素方差分析Tukey法两两比较, 分析样地之间和河流之间的植物盖度、高度、多样性和土壤养分是否存在显著差异. 采用独立T检验分析样带A和样带B之间是否存在显著差异. 采用Canoco5.0分析地形坡度、土壤养分和气候等环境因子对植物群落组成的影响. 先将物种数据和环境数据导入软件, 根据软件提示, 对物种数据进行lg(X+1). 因为三江源大部分地区年均气温为负值, 为确保能够统一方式转换, 将环境因子进行lg(X/10+1)转换. 分析环境因子对物种重要值的影响时, DCA分析结果显示最大轴特征值为4.37, 因此选择DCCA分析. 多重检验时, 采用FDR(false discovery rate)校正P值[20].
采用Canoco 5.0中的Var-part-3groups-Simple-effects-tested功能模块将显著的环境因子(P < 0.05)进行方差分解分析(variance partitioning analysis), 确定指定的环境因子对群落结构变化的解释比例.
2 结果与分析 2.1 三江源河岸带植物群落特征 2.1.1 河岸带植物群落组成三江源河岸带植被以高寒草原和高寒草甸为主. 共采集到维管束植物54种, 隶属于46属、24科;其中唇形科植物种类最多, 为6种, 其次为菊科5种, 禾本科5种, 毛茛科4.
按照《中国植物志》的描述进行生活型分类, 以多年生草本植物种类最多(39种, 占72.2%), 其次为一年生草本植物(11种, 占20.4%), 灌木有4种, 呈零星分布. 其中一年生草本植物主要有:香青兰(Dracocephalum moldavica)、黏毛鼠尾草(Salvia roborowskii)、臭蒿(Artemisia hedinii)、假水生龙胆(Gentiana pseudoaquatica)、水蓼(Persicaria hydropiper)和水苋菜(Ammannia baccifera)等. 多年生草本植物主要有:白苞筋骨草(Ajuga lupulina)、白花枝子花(Dracocephalum heterophyllum)、紫花针茅(Stipa purpurea)、冰草(Agropyron cristatum)、三裂碱毛茛(Halerpestes tricuspis)、麻花艽(Gentiana straminea)和高山嵩草(Carex parvula)等. 灌木主要有:沙棘(Hippophae rhamnoides)、甘青铁线莲(Clematis tangutica)、鬼箭锦鸡儿(Caragana jubata)和金露梅(Dasiphora fruticosa)等.
2.1.2 河岸带植物群落优势种分布不同样带区域的植物群落优势种存在着一定的差异(表 2). 14个调查样地的样带A区域, 紫花针茅优势最为明显, 在10个样带中为优势种, 其次为高山嵩草、冰草和麻花艽.样带B中, 紫花针茅优势也很明显, 在10个样带中为优势种, 其次为急弯棘豆、蕨麻和冰草等.
|
|
表 2 不同样带优势种1) Table 2 Dominant species (dominance) of different sample belts |
2.1.3 河岸带植物群落盖度和高度特征
三江源河岸带植物群落盖度和高度均值分别为(64.4 ± 23.6)%和(15.8 ± 10.7)cm, 变幅分别为21.0%~134.3%和2.0~40.0 cm[图 2(a)和图 2(b)].

|
图 2 三江源河岸带植物群落特征 Fig. 2 Plant community characteristics in riparian zone of the Three-River Headwaters region |
不同样地之间比较, 样地S5植物盖度均值最大[(115.7 ± 26.4)%], 其次为S8[(81.7 ± 16.0)%]和S13[(74.5 ± 19.6)%], S14最小[(36.3 ± 4.2)%], 但差异不显著. S11处植物高度最大, 为(35.7 ± 6.0)cm, 显著大于S5和S6(F = 4.763, P = 0.003).
不同样带之间比较, 样带A的盖度均值为(57.3 ± 27.3)%, 低于样带B[(71.5 ± 17.5)%].样带A的高度均值为(13.8 ± 6.0)cm, 低于样带B[(15.3 ± 7.4)cm], 但差异不显著.
不同河流之间比较, 长江源植物盖度均值最大[(67.3 ± 25.5)%], 黄河源次之[(62.25 ± 21.11)%], 澜沧江源最小[(48.50 ± 24.60)%], 但差异不显著. 长江源植物群落高度最大, 澜沧江源次之, 黄河源最小, 但差异不显著.
2.1.4 河岸带植物群落多样性特征三江源河岸带消落区植物群落Shannon-Wiener多样性指数和Pielou均匀度指数均值分别为1.31 ± 0.42和0.84 ± 0.08, 变幅分别为0.51~2.05和0.72~0.99[图 2(c)和图 2(d)].
不同样地之间, 不同河流之间植物群落多样性指数差异均不显著. 其中澜沧江源植物群落Shannon-Wiener指数均值最大, 长江源次之, 黄河源最小. 澜沧江源和黄河源植物群落Pielou均匀度指数相近, 略大于长江源.
2.2 三江源河岸带土壤生境特征 2.2.1 河岸带土壤pH、有机质、全氮分布特征三江源河岸带土壤pH、ω(有机质)和ω(全氮)均值分别为8.43 ± 0.40、(21.28 ± 26.87)g·kg-1和(1.35 ± 1.55)g·kg-1, 变化范围分别为7.65~9.08、1.13~108.15 g·kg-1和0.08~5.36 g·kg-1, 变异系数分别为4.70%、126.29%和114.83%. 有机质和全氮的平均值大于中位数, 而pH的均值小于中位数[图 3(a)~3(c)].

|
图 3 河岸带土壤pH、有机质、全氮、全磷、钙镁结合态磷和铁铝结合态磷分布 Fig. 3 Distribution of soil pH, organic matter, total nitrogen, total phosphorus, calcium-magnesium bound phosphorus, and iron-aluminum bound phosphorus in riparian zone |
不同样地之间和不同样带之间比较, 土壤pH、有机质和全氮均值差异均不显著. 不同河流之间比较, 土壤pH显著大于澜沧江源(F = 5.192, P = 0.013), 但有机质(F = 5.443, P = 0.011)和全氮均值(F = 4.489, P = 0.022)显著低于澜沧江源.
2.2.2 河岸带土壤各形态磷分布特征三江源河岸带土壤ω(全磷)、ω(铁铝结合态磷)、ω(钙镁结合态磷)和ω(有机磷)均值分别为(693.48 ± 393.39)、(50.30 ± 52.06)、(437.12 ± 306.07)和(243.47 ± 166.44)mg·kg-1, 变化范围分别为311.66~1735.16、11.87~272.88、118.00~1350.35和333.5~574.93 mg·kg-1, 变异系数分别为56.73%、103.50%、70.02%和68.36%. 全磷、铁铝结合态磷、钙镁结合态磷和有机磷的均值大于中位数[图 3(d)~3(f)和图 4]. 河岸带土壤磷的主要形态为钙镁结合态磷, 占全磷的质量分数为63.89%.

|
图 4 河岸带土壤有机磷分布 Fig. 4 Distribution of soil organic phosphorus in riparian zone |
不同样地之间、不同样带之间比较, 土壤全磷、铁铝结合态磷、钙镁结合态磷和有机磷含量差异均不显著. 长江源河岸带土壤铁铝结合态磷含量均值显著低于澜沧江源(F = 3.717, P = 0.039).
2.2.3 影响河岸带植物群落特征的关键环境因子DCCA结果显示, 14个环境因子共可解释植物群落结构变异的63.09%, 其中第一和第二轴的解释率分别为10.98%和7.62%. 气温对物种重要值单独效应最大(8.1%), 其次为降雨量(7.8%)、海拔(7.2%)、有机磷(6.5%)、径流深(6.2%)、河床比降(6.2%)、蒸发量(6.1%)和pH(6.0%)(Padj < 0.05). 但通过Monte Carlo检验环境因子(Padj < 0.05)只有气温(F = 2.4, Padj = 0.014)、土壤有机磷(F = 1.9, Padj = 0.036)和土壤pH值(F = 2.0, Padj = 0.037), 表明影响三江源河岸带植物群落结构的环境因子主要是气温、土壤有机磷和pH值(21.49%), 其中第一和第二轴的解释率分别为9.8%和6.28%(图 5).

|
显示权重前20的物种, 其中Aj lu为白苞筋骨草, De ta为康定翠雀花, Dr he为白花枝子花, Cl ta为甘青铁线莲, St ch为狼毒, Ge st为麻花艽, Dy sc为菊叶香藜, Pe ve为轮叶马先蒿, Ar an为蕨麻, Ar he为臭蒿, El da为披碱草, St by为绵毛水苏, Ca pa为高山嵩草, Aj pr为细裂亚菊, Ha tr为三裂碱毛茛, Pe hy:水蓼, Pe fl为白草, Ag cr为冰草, An ta为垫状点地梅, St pu为紫花针茅 图 5 河岸带植物重要值与环境变量的关系 Fig. 5 Relationship among the important value of plant species and environmental variables in riparian zone |
解释方差调整后, 气温、土壤有机磷和pH共同解释了河岸带植物群落结构变化的11.68%(pseudo-F = 2.2, P = 0.001), 气温、土壤有机磷和pH分别单独揭示了河岸带植物群落结构变化的5.8%、3.7%和3.2%(图 6).

|
图 6 河岸带植物重要值与环境变量的方差分解 Fig. 6 Variation partitioning results showing the explanatory power of environmental variables for the important value of plant species |
对植物群落多样性特征的DCA分析最大轴特征值为0.02, 因此选择RDA分析, 结果显示, 14个环境因子共可解释植物群落多样性变异的88.61%, 其中第一和第二轴的解释率分别为78.55%和8.73%. 土壤钙镁结合态磷对多样性指数的单独效应最大(47.7%), 其次为pH(39.2%)、土壤全磷(37.7%)、铁铝结合态磷(32.1%)、有机质(23.6%)和海拔(23.0%)(Padj < 0.05). 但通过Monte Carlo检验环境因子(Padj < 0.05)只有土壤钙镁结合态磷(F = 23.7, Padj = 0.016), 表明影响三江源河岸带植物群落多样性的关键环境因子为土壤钙镁结合态磷(图 7).

|
图 7 河岸带植物群落多样性与环境变量的关系 Fig. 7 Relationship among the plant community diversity and environmental variables in riparian zone |
气候因子(年均气温和年均降水量等)能够很好地解释物种分布、多样性及生产力的大尺度格局[21, 22]. Michaletz等[23]认为气候是通过影响植物个体大小、群落年龄结构、群落生物量和生长季长度等间接影响生产力, 而不是温度和降水的代谢动力学效应直接决定群落生产力. 本研究也表明了年均气温和降雨量对三江源河岸带植物重要值的单独效应显著(Padj < 0.05), 排在第1位和第2位(图 5).
三江源区具有典型的高原气候特征, 气温低、降雨量小、风速大、紫外线辐射强和昼夜温差大, 极大限制了植物的生存、生长和空间分布, 显著影响着植物的外在形态和适应策略[24]. 垫状植被是三江源植被覆盖的一个鲜明特色, 也是高山植物种间互助的重要证据和典型代表[25], 主要有垫状点地梅、垫状棱子芹、独一味、蕨麻、雪灵芝和钻叶风毛菊等(表 2), 均贴着地面生长, 小枝多呈辐射性生长, 形成紧密簇生球形或半球形结构, 具有一定的保温作用, 能够改善枝叶覆盖下的土壤容重和含水量等指标, 为其自身及其他物种创造并维持了一个条件更加优越的微生境[26]. 年降雨量 < 100 mm时会显著威胁垫状植物自身的生存[27]. 垫状植被对气候变化非常敏感, 全球气候变暖导致低海拔物种向高海拔迁移, 势必挤压垫状植被的生态空间. 降雨量的年内及年际变化显著影响着河流的水文条件(丰枯变化、流量大小、水位高低、水淹时间等)的变化, 而水文变化对河岸带植物形态、分布、繁殖和生存策略的影响是最显著的[28], 还能间接影响河岸带土壤氮磷迁移转化规律和养分循环[29].
植株密披绒毛或具多刺是三江源植被的另一重要形态适应机制. 三江源河岸带很多植物的茎干、叶片和花序的表面上生有厚厚的绒毛, 具有保温和抗紫外线作用, 以适应冬季的极端低温环境和强紫外线辐射环境, 主要典型植物有火绒草、鬼箭锦鸡儿和绵毛水苏等(表 2). 还有些植物枝具多刺, 叶小型化, 以适应三江源降雨量小, 夏季高温干旱, 土壤贫瘠等环境, 主要典型植物有沙棘和节毛飞廉等.
花色艳丽, 图案多样是三江源常见植物的繁殖适应策略之一. 三江源植物花瓣中产生大量类胡萝卜素和花青素可以有效反射阳光中的紫外线, 以减轻紫外辐射的伤害, 这两种物质使花冠呈现蓝色、紫色、黄色和红色, 从而成为高山植物的常见花色, 主要典型植物有微孔草、天蓝韭、假水生龙胆、草甸雪兔子、紫花针茅、缘毛紫菀、康定翠雀花和独一味等(表 2). 这些花色也是“迎合”传粉者偏好的特有适应性状, 其中熊蜂本身偏爱拜访紫蓝色花[30]. 白苞筋骨草和塔黄均具有高度特化的黄色半透明苞片, 相互交错, 层层叠叠, 越往上越小, 整体呈塔状, 具有增温、防风和防雨等作用, 还为蕈蚊提供交配场所, 为成虫产卵和幼虫发育提供适宜的微环境[31].
3.2 土壤养分对河岸带植物群落特征的影响河岸带土壤养分决定着植物群落结构和功能的局部尺度格局, 显著影响着植被覆盖度和生物量的大小, 而土壤的有无, 直接决定着植物是否存在[32]. 本研究结果表明, 显著影响河岸带植物群落结构的土壤因子是土壤有机磷含量和pH值(图 5). 已有研究表明土壤养分含量较高的生态系统比土壤养分贫乏的生态系统具有较低的生产力的年际变异, 能够更好地缓冲气候变化的影响[33], 这是因为肥沃的土壤可为植物生长提供更广的生态位. 全球自然陆地生态系统(农田、城市和冰川除外)有43%的区域受到较强的磷限制, 18%的区域受到较强的氮限制, 而其他39%的区域则受氮、磷共同限制或氮、磷任一元素的微弱限制[34]. 在自然生态系统中, 磷添加使地上生产力提高34.9%, 农田仅提高13.9%, 这可能与农田土壤具有更高的磷有效性以及持续的磷肥施用历史有关[35].
土壤有效养分含量决定着植物的初级生产潜力, 但这类的研究还相对较少. 本研究结果表明土壤有机磷含量显著影响着河岸带的植物群落结构(图 5和图 6), 钙镁结合态磷含量对多样性的影响显著(图 7). 对藏北退化草甸植被生长贡献率最高的土壤环境因子为速效氮、有机质和pH[36]. 土壤的酸碱性影响土壤中元素对植物的有效性, 土壤酸化和碱化都降低了荒漠草原群落的多样性和稳定性[37]. 库岸带植物鲜重与土壤养分含量均呈正相关, 其中土壤有效氮含量与植物鲜重呈显著的正相关[38].
3.3 地形地貌对河岸带植物群落特征的影响高程或海拔也是影响河岸带植物种类分布的关键因子之一[29, 39, 40]. 本研究表明海拔(7.2%)与河床比降(6.2%)对河岸带植被的单独影响效应较大(Padj < 0.05). 在局部尺度上, 河岸带高程的微小差异可形成复杂的微地形特征, 影响植物群落的组成[41]. 季节性洪水之所以影响河岸带植被组成和分布格局[42, 43], 是因为河岸带的低高程区域, 洪水淹没和冲刷频繁, 只有耐水淹和根系发达的植物才能生存, 而相对不耐水淹的植物多分布在较高的高程区域[44, 45]. 为了适应周期性与季节性的洪水冲刷, 低高程区域分布的植物高度普遍较低, 本研究结果也证实了这点, 样带A的盖度和多样性均低于样带B. 水位涨落幅度大的库区, 高程对库岸带物种分布和多样性的影响更大[32, 46].
坡度是影响河岸带植被特征的另一个重要地貌因子. 已有研究表明坡度越大, 土层厚度越小, 植被盖度和生物量越低[38]. 本研究结果显示黄河源支流格曲河玛沁县段裸露的陡坡岩质很常见, 既无土壤, 也无植被覆盖, 在岩石缝隙中零星分布有少量的草本植物. 通天河干流治多县段陡坡土质库岸带植被常呈斑块化分布, 土壤长期受涌浪侵蚀, 土层越来越薄, 植被将失去赖以生存生长的基质. 坡的陡峭程度影响着河岸带土壤侵蚀机制和营养物质组成[47], 侵蚀影响河岸植被的演替格局, 导致河岸带植被呈斑块分布[8].
河岸带植被的分布格局是自然因素和人类活动及其综合作用的结果[48, 49]. 三江源区河流密布, 局部气候及土壤养分等自然地理要素对河岸带植被分布的影响占绝对优势. 与其它区域相比, 三江源人口密度小, 人类的社会活动对河岸带的影响相对较小, 主要集中于河流的城集镇段, 如长江源通天河支流巴塘河玉树市段两岸均为硬质化岸坡, 河岸带植被稀疏, 澜沧江源吉曲囊谦县城段河岸带植被多呈斑块化分布.
4 结论(1)三江源河岸带植被以高寒草原、高寒草甸为主, 唇形科植物种类最多, 其次为菊科和禾本科, 多年生草本植物种类最多(72.2%), 其次为一年生草本植物(20.4%)和灌木(7.4%).
(2)长江源河岸带土壤有机质、全氮、铁铝结合态磷含量均值显著低于澜沧江源, 而长江源河岸带土壤pH显著大于澜沧江源. 河岸带土壤磷的主要形态为钙镁结合态磷, 占全磷的质量分数为63.89%.
(3)影响三江源河岸带植物群落组成的关键环境因子为气温、土壤有机磷含量和pH值, 影响植物群落多样性的关键环境因子为土壤钙镁结合态磷含量.
| [1] |
张婵, 安宇梦, Jäschke Y, 等. 青藏高原及周边高山地区的植物繁殖生态学研究进展[J]. 植物生态学报, 2020, 44(1): 1-21. Zhang C, An Y M, Jäschke Y, et al. Processes on reproductive ecology of plant species in the Qinghai-Xizang Plateau and adjacent highlands[J]. Chinese Journal of Plant Ecology, 2020, 44(1): 1-21. |
| [2] |
林马震, 黄勇, 李洋, 等. 高寒草地植物生存策略地理分布特征及其影响因素[J]. 植物生态学报, 2023, 47(1): 41-50. Lin M Z, Huang Y, Li Y, et al. Geographical distribution characteristics and influencing factors of plant survival strategies in an alpine grassland[J]. Chinese Journal of Plant Ecology, 2023, 47(1): 41-50. |
| [3] | 赵新全. 三江源国家公园生态系统现状、变化及管理[M]. 北京: 科学出版社, 2021. |
| [4] |
马世鹏, 孙海群. 三江源自然保护区湿地种子植物区系分析[J]. 青海大学学报(自然科学版), 2015, 33(4): 17-24. Ma S P, Sun H Q. Flora of seed plants in wetlands of the Sanjiangyuan Natural Reserve[J]. Journal of Qinghai University (Natural Science Edition), 2015, 33(4): 17-24. |
| [5] |
彭景涛, 李国胜, 傅瓦利, 等. 青海三江源地区退化草地土壤全氮的时空分异特征[J]. 环境科学, 2012, 33(7): 2490-2496. Peng J T, Li G S, Fu W L, et al. Temporal-spatial variations of total nitrogen in the degraded grassland of Three-River Headwaters region in Qinghai Province[J]. Environmental Science, 2012, 33(7): 2490-2496. |
| [6] |
吴丹, 邵全琴, 刘纪远, 等. 三江源地区林草生态系统水源涵养服务评估[J]. 水土保持通报, 2016, 36(3): 206-210. Wu D, Shao Q Q, Liu J Y, et al. Assessment of water regulation service of forest and grassland ecosystems in Three-River Headwaters region[J]. Bulletin of Soil and Water Conservation, 2016, 36(3): 206-210. |
| [7] | Nilsson C, Berggren K. Alterations of riparian ecosystems caused by river regulation: dam operations have caused global-scale ecological changes in riparian ecosystems. How to protect river environments and human needs of rivers remains one of the most important questions of our time[J]. BioScience, 2000, 50(9): 783-792. DOI:10.1641/0006-3568(2000)050[0783:AORECB]2.0.CO;2 |
| [8] |
韩路, 王海珍, 于军. 河岸带生态学研究进展与展望[J]. 生态环境学报, 2013, 22(5): 879-886. Han L, Wang H Z, Yu J. Research progress and prospects on riparian zone ecology[J]. Ecology and Environmental Sciences, 2013, 22(5): 879-886. DOI:10.3969/j.issn.1674-5906.2013.05.026 |
| [9] |
申健勇, 吴金清. 嘉陵江河岸带维管植物区系的初步研究[J]. 华中师范大学学报(自然科学版), 2012, 46(6): 733-739, 788. Shen J Y, Wu J Q. Floristic analysis of vascular plants in riparian zone of Jialing River[J]. Journal of Huazhong Normal University (Natural Sciences), 2012, 46(6): 733-739, 788. DOI:10.3969/j.issn.1000-1190.2012.06.020 |
| [10] |
修晨, 欧阳志云, 郑华. 北京永定河-海河干流河岸带植物的区系分析[J]. 生态学报, 2014, 34(6): 1535-1547. Xiu C, Ouyang Z Y, Zheng H. Flora analysis of riparian vegetation in Yongding-Haihe river system, China[J]. Acta Ecologica Sinica, 2014, 34(6): 1535-1547. |
| [11] | Li J P, Dong S K, Yang Z F, et al. Effects of cascade hydropower dams on the structure and distribution of riparian and upland vegetation along the middle-lower Lancang-Mekong River[J]. Forest Ecology and Management, 2012, 284: 251-259. DOI:10.1016/j.foreco.2012.07.050 |
| [12] | Zhang J T, Xiang C L, Li M. Effects of tourism and topography on vegetation diversity in the subalpine meadows of the Dongling Mountains of Beijing, China[J]. Environmental Management, 2012, 49: 403-411. |
| [13] | 鲍士旦. 土壤农化分析[M]. 北京: 中国农业出版社, 2000, 第三版. |
| [14] | Shan B Q, Jie L, Zhang W Q, et al. Characteristics of phosphorus components in the sediments of main rivers into the Bohai sea[J]. Ecological Engineering, 2016, 97: 426-433. DOI:10.1016/j.ecoleng.2016.10.042 |
| [15] | Zhang Z Y, Hu H Q, Wan C Y, et al. Lateral and longitudinal variation in phosphorus fractions in surface sediment and adjacent riparian soil in the Three Gorges Reservoir, China[J]. Environmental Science and Pollution Research, 2018, 25(31): 31262-31271. DOI:10.1007/s11356-018-3087-7 |
| [16] |
张文强, 李洁, 金鑫, 等. 基于31P-NMR与SMT方法的海河流域典型河流沉积物中磷形态特征研究[J]. 环境科学学报, 2016, 36(6): 1891-1900. Zhang W Q, Li J, Jin X, et al. Detection of phosphorus components in the sediments of typical river in the Haihe Basin by SMT and 31P-NMR[J]. Acta Scientiae Circumstantiae, 2016, 36(6): 1891-1900. |
| [17] | 玉树藏族自治州水利志(1951-2017)[M]. 郑州: 黄河水利出版社, 2019. |
| [18] | 《中国河湖大典》编纂委员会. 中国河湖大典(黄河卷)[M]. 北京: 中国水利水电出版社, 2014. |
| [19] | 国家气象信息中心, 数据应用服务室. 三江源及邻近区域国家标准气象站逐月气象数据(1957-2015)[EB/OL]. https://doi.org/10.11888/Meteoro.tpdc.270540, 2022-05-27. |
| [20] | Lepš J, Šmilauer P. Multivariate analysis of ecological data using CANOCO: visualizing multivariate data[M]. New York: Cambridge University Press, 2003. |
| [21] | Ponce-Campos G E, Moran M S, Huete A, et al. Ecosystem resilience despite large-scale altered hydroclimatic conditions[J]. Nature, 2013, 494(7437): 349-352. DOI:10.1038/nature11836 |
| [22] | Yao Z Y, Xin Y, Yang L, et al. Precipitation and temperature regulate species diversity, plant coverage and aboveground biomass through opposing mechanisms in large-scale grasslands[J]. Frontiers in Plant Science, 2022, 13. DOI:10.3389/fpls.2022.999636 |
| [23] | Michaletz S T, Kerkhoff A J, Enquist B J. Drivers of terrestrial plant production across broad geographical gradients[J]. Global Ecology and Biogeography, 2018, 27(2): 166-174. DOI:10.1111/geb.12685 |
| [24] |
赵宁. 青藏高原极端环境植物适应性进化分子机制研究进展[J]. 环境生态学, 2022, 4(1): 65-70. Zhao N. Advances in molecular mechanisms of adaptive evolution of plants in extreme environments on the Qinghai-Tibet Plateau[J]. Environmental Ecology, 2022, 4(1): 65-70. |
| [25] | Kikvidze Z, Brooker R W, Butterfield B J, et al. The effects of foundation species on community assembly: a global study on alpine cushion plant communities[J]. Ecology, 2015, 96(8): 2064-2069. DOI:10.1890/14-2443.1 |
| [26] |
杨扬, 陈建国, 宋波, 等. 青藏高原冰缘植物多样性与适应机制研究进展[J]. 科学通报, 2019, 64(27): 2856-2864. Yang Y, Chen J G, Song B, et al. Advances in the studies of plant diversity and ecological adaptation in the subnival ecosystem of the Qinghai-Tibet Plateau[J]. Chinese Science Bulletin, 2019, 64(27): 2856-2864. |
| [27] | de Bello F, Doležal J, Dvorský M, et al. Cushions of Thylacospermum caespitosum (Caryophyllaceae) do not facilitate other plants under extreme altitude and dry conditions in the north-west Himalayas[J]. Annals of Botany, 2011, 108(3): 567-573. DOI:10.1093/aob/mcr183 |
| [28] |
郭二辉, 樊子豪, 张瑞香, 等. 河岸带生态系统植被与土壤对水文变化的响应研究进展[J]. 生态学报, 2021, 41(23): 9164-9173. Guo E H, Fan Z H, Zhang R X, et al. Research progresses on the response of vegetation and soil to hydrological changes in riparian ecosystems[J]. Acta Ecologica Sinica, 2021, 41(23): 9164-9173. |
| [29] |
凌玲, 闫淑君, 关永鑫, 等. 河岸带植被群落物种组成及多样性分布特征[J]. 世界林业研究, 2023, 36(5): 27-33. Ling L, Yan S J, Guan Y X, et al. Species composition and diversity distribution characteristics of vegetation community in riparian zones[J]. World Forestry Research, 2023, 36(5): 27-33. |
| [30] |
彭德力, 张志强, 牛洋, 等. 高山植物繁殖策略的研究进展[J]. 生物多样性, 2012, 20(3): 286-299. Peng D L, Zhang Z Q, Niu Y, et al. Advances in the studies of reproductive strategies of alpine plants[J]. Biodiversity Science, 2012, 20(3): 286-299. |
| [31] | Song B, Stöcklin J, Peng D L, et al. The bracts of the alpine "glasshouse" plant Rheum alexandrae (Polygonaceae) enhance reproductive fitness of its pollinating seed-consuming mutualist[J]. Botanical Journal of the Linnean Society, 2015, 179(2): 349-359. DOI:10.1111/boj.12312 |
| [32] |
张志永, 向林, 万成炎, 等. 三峡水库消落区植物群落演变趋势及优势植物适应策略[J]. 湖泊科学, 2023, 35(2): 553-563. Zhang Z Y, Xiang L, Wan C Y, et al. Evolutionary trend of plant community and adaptive strategies of dominant plants in the water-level-fluctuation zone of the Three Gorges Reservoir[J]. Journal of Lake Sciences, 2023, 35(2): 553-563. |
| [33] | Yan P, Zhang J H, He N P, et al. Functional diversity and soil nutrients regulate the interannual variability in gross primary productivity[J]. Journal of Ecology, 2023, 111(5): 1094-1106. |
| [34] | Du E Z, Terrer C, Pellegrini A F A, et al. Global patterns of terrestrial nitrogen and phosphorus limitation[J]. Nature Geoscience, 2020, 13(3): 221-226. |
| [35] | Hou E Q, Luo Y Q, Kuang Y W, et al. Global meta-analysis shows pervasive phosphorus limitation of aboveground plant production in natural terrestrial ecosystems[J]. Nature Communications, 2020, 11(1). DOI:10.1038/s41467-020-14492-w |
| [36] |
彭艳, 孙晶远, 马素洁, 等. 藏北不同退化阶段高寒草甸植物群落特征与土壤养分特性[J]. 草业学报, 2022, 31(8): 49-60. Peng Y, Sun J Y, Ma S J, et al. Plant community composition and soil nutrient status of degraded alpine meadow sites in Northern Tibet[J]. Acta Prataculturae Sinica, 2022, 31(8): 49-60. |
| [37] | Liu K, Liu Z C, Zhou N, et al. Diversity-stability relationships in temperate grasslands as a function of soil pH[J]. Land Degradation & Development, 2022, 33(10): 1704-1717. |
| [38] |
张志永, 程郁春, 程丽, 等. 三峡库区万州段消落带植被及土壤理化特征分析[J]. 水生态学杂志, 2016, 37(2): 24-33. Zhang Z Y, Chen Y C, Cheng L, et al. Characteristics of vegetation and soil in the water level fluctuation zone of the Wanzhou Region of Three Gorges Reservoir[J]. Journal of Hydroecology, 2016, 37(2): 24-33. |
| [39] |
许莎莎, 孙国钧, 刘慧明, 等. 黑河河岸植被与环境因子间的相互作用[J]. 生态学报, 2011, 31(9): 2421-2429. Xu S S, Sun G J, Liu H M, et al. The relationship between riparian vegetation and environmental factors in Heihe River Basin[J]. Acta Ecologica Sinica, 2011, 31(9): 2421-2429. |
| [40] |
桂东伟, 雷加强, 曾凡江, 等. 中昆仑山北坡策勒河流域生态因素对植物群落的影响[J]. 草业学报, 2010, 19(3): 38-46. Gui D W, Lei J Q, Zeng F J, et al. Effect of ecological factors on plant communities of the Cele River Basin on the north slope of the middle Kunlun mountains[J]. Acta Prataculturae Sinica, 2010, 19(3): 38-46. |
| [41] |
江明喜, 邓红兵, 唐涛, 等. 香溪河流域河岸带植物群落物种丰富度格局[J]. 生态学报, 2002, 22(5): 629-635. Jiang M X, Deng H B, Tang T, et al. On Spatial pattern of species richness in plant communities along riparian zone in Xiangxi River watershed[J]. Acta Ecologica Sinica, 2002, 22(5): 629-635. |
| [42] | Vervuren P J A, Blom C W P M, De Kroon H. Extreme flooding events on the Rhine and the survival and distribution of riparian plant species[J]. Journal of Ecology, 2003, 91(1): 135-146. |
| [43] | Goebel P C, Pregitzer K S, Palik B J. Influence of flooding and landform properties on riparian plant communities in an Old-Growth Northern Hardwood Watershed[J]. Wetlands, 2012, 32(4): 679-691. |
| [44] | Lenssen J P M, De Kroon H. Abiotic constraints at the upper boundaries of two Rumex species on a freshwater flooding gradient[J]. Journal of Ecology, 2005, 93(1): 138-147. |
| [45] | van Eck W H J M, Lenssen J P M, van De Steeg H M, et al. Seasonal dependent effects of flooding on plant species survival and zonation: a comparative study of 10 terrestrial grassland species[J]. Hydrobiologia, 2006, 565(1): 59-69. |
| [46] |
江维薇, 李文涛, 肖衡林. 澜沧江流域糯扎渡水库消落带植被的物种组成、空间分布特征及地形解释[J]. 湖泊科学, 2022, 34(6): 2025-2038. Jiang W W, Li W T, Xiao H L. Species composition, spatial distribution characteristics and topographic attribution of vegetation in the water-level-fluctuating zone of the Nuozhadu Reservoir in the Lancang River Basin[J]. Journal of Lake Sciences, 2022, 34(6): 2025-2038. |
| [47] |
张媛媛, 王晓锋, 王轶浩, 等. 三峡消落带不同坡度狗牙根及实生土壤生态化学计量特征[J]. 生态学报, 2023, 43(11): 4798-4811. Zhang Y Y, Wang X F, Wang Y H, et al. Ecological stoichiometry and interrelation of Cynodon dactylon and soil in the three Gorges hydro-fluctuation zone under different slopes[J]. Acta Ecologica Sinica, 2023, 43(11): 4798-4811. |
| [48] | You X G, Liu J L, Zhang L L. Ecological modeling of riparian vegetation under disturbances: a review[J]. Ecological Modelling, 2015, 318: 293-300. |
| [49] | Henriques M, Mcvicar T R, Holland K L, et al. Riparian vegetation and geomorphological interactions in anabranching rivers: a global review[J]. Ecohydrology, 2022, 15(2). DOI:10.1002/eco.2370 |