 2024, Vol. 45
2024, Vol. 45


2. 宝鸡文理学院地理与环境学院,宝鸡 721013;
3. 陕西省灾害监测与机理模拟重点实验室,宝鸡 721013
2. Department of Geography and Environment, Baoji University of Arts and Sciences, Baoji 721013, China;
3. Key Laboratory of Disaster Monitoring and Mechanism Simulation of Shaanxi Province, Baoji 721013, China
近年来土壤重金属污染是世界各国普遍面临的环境问题. 在我国许多地区的土壤和地下水重金属积累问题持续加重[1], 重金属污染土壤已达333万hm2, 且这个数字不断在增长, 重金属污染会对生态安全、食品安全和人类健康造成严重的潜在威胁[2, 3]. 重金属污染具有累积性、隐蔽性、危害滞后性、持久性、难降解性、不可逆性、难治理与难修复等特点[4]. 因此, 控制环境中重金属的污染, 对保护人类健康、减少其潜在危害具有重要意义. 目前诸多的重金属污染土壤修复技术中, 最受青睐的是植物修复技术, 植物修复是一种绿色的原位修复技术, 不会对环境造成二次污染[5~7]. 植物修复相当于一个自然过程, 不破环土壤原有的生态环境, 能使土壤基本保持好的结构和肥力[8, 9]. 植物修复的核心是超富集植物种类的选择, 迄今为止国际上已发现的重金属超富集植物种质资源有限, 而且已发现的重金属超富集植物绝大多数都存在生物量小、富集能量有限、生长缓慢、适应性弱、仅对单一的重金属污染物表现出超富集性能等特点[10, 11], 因此国内外学者对重金属超富集植物的研究主要集中在单一重金属超富集植物上, 对Pb、Cd和Zn复合金属超富集植物的研究鲜有报道, 而自然界中的重金属往往是以复合形式存在的.
宝鸡市是我国四大铅锌产地之一, 在长期的开采、浮选、精炼及加工的过程中, 产生的废气、废水和固废等使得周边土壤Pb、Zn和Cd复合重金属污染问题较为凸出[12~14]. 通常, 在矿区生长的植物长期受高含量重金属污染环境的驯化和自然筛选, 会存在一些优势植物对重金属具有较高的耐受性和潜在的超富集能力[15, 16]. 因此, 本研究对宝鸡市凤县至太白区域铅锌采矿区及尾矿库、凤县铅锌冶炼厂和凤翔铅锌冶炼厂周围生长的植物进行野外调查, 采集优势植物, 分析其对重金属的富集能力, 拟筛选出对复合重金属具有潜在的超富集能力的植物, 并通过室内土培盆栽试验, 研究其对Pb、Zn和Cd复合重金属的胁迫响应特征, 揭示其耐性机制;再经过野外大田试验进行验证, 最终筛选出Pb、Zn和Cd复合重金属污染超富集植物, 通过丰富超富集植物的种质资源, 以期为陕西省矿区土壤重金属污染的植物修复技术提供技术支持, 也为全国同类的重金属土壤污染修复提供借鉴.
1 材料与方法 1.1 研究区域概况宝鸡市凤县是陕西省典型的金属矿产资源分布区, Pb和Zn矿贮量达450万t, 其中已探明350万t, 占陕西省的80%, Pb和Zn资源/储量分别占全省Pb和Zn总资源/储量的26.31%和51.58%, 是中国四大铅锌矿产资源基地之一[17~19]. 近年来随着秦岭保护政策的出台与实施, 秦岭采矿和浮选企业逐渐退出和关闭, 大量的尾矿、矿渣被遗留在矿区和尾矿库区域, 被遗留的重金属污染问题非常严重.
本研究采样区域选取宝鸡市凤县至太白区域铅锌采矿区与尾矿库;以及凤县铅锌冶炼厂区域(106°24'54″~107°07'30″E, 33°34′50″~34°18′13″N), 该冶炼厂于2000年开始建设, 2001年运行, 年精炼Pb和Zn约6万t和生产硫酸12万t[18]. 另外, 选择凤翔铅锌冶炼厂(107°13′41″~107°14′22″E, 34°27′35″~34°28′43″N), 该厂于2006年建成, 年精炼30万t铅锭和锌锭、70万t焦化产品[20]. 采样区域土壤重金属含量情况如表 1所示. 研究区域内各层土壤中重金属含量几乎均已超过陕西省土壤背景值, 各层土壤中Zn和Cd含量已大大超过国家农用地土壤二级污染风险筛选值.
|
|
表 1 土壤中重金属含量统计分析1)/mg·kg-1 Table 1 Descriptive statistical summary of heavy metal content in soils/mg·kg-1 |
1.2 样品采集与处理 1.2.1 样品采集
植物样品选取宝鸡市太白-凤县沿线地带的铅锌矿与尾矿库、凤县铅锌冶炼厂和凤翔铅锌冶炼厂周边区域出现频率高、数量相对多、生长旺盛且生物量大的草本植物, 分别在不同位置随机采集3~16株植物, 包括地上和地下部分及根部根际0~20 cm的土壤, 共采集了40种植物, 320个植物样品和320个土壤样品. 将采集好的植物和土壤样品装袋, 并进行编号, 带回实验室.
野外大田试验, 3月下旬种植, 10月上旬采集, 每个区块随机采集10株完整植株带回实验室.
1.2.2 样品处理将采集的野外优势植物和大田试验植株清除杂质, 根茎分开, 冲洗附带的泥土等杂质, 并用水浸泡10 min后再冲洗2~3次, 然后将其置于2%的NaCl水溶液中浸泡约30 min, 用去离子水将植株反复清洗干净后挂起风干水分, 置于烘箱中于105℃下杀青约30 min, 杀青后在70~75℃下烘干至恒重, 将地上和地下部分分别用植物粉碎机粉碎后装袋备用.
由于植物酶的胁迫响应研究须使用植物鲜样, 从植物活体上采集后需要迅速处理, 以免酶失活, 条件要求相对较为苛刻, 故土培盆栽植物均在活体上随机采集部分叶片称重后, 在冰盒内迅速剪碎至冷冻研钵中, 立即用液氮冷冻后加入提取液快速研磨, 将研磨后的匀浆转移至容量瓶中, 按标准方法进行测定.
1.2.3 样品消解称取粉碎的植物样品约0.500 0 g, 并做好记录, 置于聚四氟乙烯消解管中, 用洗瓶加入少量超纯水待植物样品润湿后加入10.00 mL浓HNO3, 置于消解仪上在180℃下消煮30 min后取下, 待温度降低并远离消解炉加入8.00 mL HClO4后置于消解炉上, 在180℃下继续消煮至近干, 取下后加入5.00 mL硝酸消煮约10 min, 再加入20.0 mL超纯水煮15 min后取下, 待降至室温后将其转移到50.00 mL容量瓶中, 用少量超纯水将消解管冲洗2~3次一并转入, 并定容至刻度线, 摇匀后用注射器+0.45 μm水系过滤膜过滤后, 取10 mL装入离心管贴好标签储存于冰箱冷藏待测.
1.3 试验方法 1.3.1 试验设计(1)室内盆栽试验设计 由于抗氧化酶和其他生理特征测定需要在活体采集鲜叶, 即刻测定, 为了获得有一定梯度的胁迫浓度, 本研究采用室内人工模拟重金属污染土壤进行盆栽试验, 将供试土壤剔除石块和植物根系等杂质, 捣碎大颗粒并过筛, 设置Pb、Zn和Cd 6个梯度复合重金属污染处理(见表 2)和空白处理CK, 共7组处理, 每组处理设置3个重复, 将Pb、Zn和Cd复合重金属水溶液与土壤搅拌均匀, 装入塑料桶陈化4个月, 期间每月倒出搅拌一次, 陈化结束后测定各梯度的土壤含水率, 并折算成干重, 分别称取陈化好的各梯度模拟土壤各12 kg分别装入对应的花盆(上缘外口径40 cm, 底部外直径20.3 cm, 高37 cm, 每个花盆带托盘底座)中. 每个梯度平行种植3盆. 聚合草为切根繁殖植物, 将直径均匀且不小于0.5 cm粗细的聚合草肉质根系切成约3 cm长的根段, 每个梯度每盆中埋入3段;同时选择有经济价值、生物量大的蓖麻作为共同研究对象, 选择籽粒饱满的8粒蓖麻种子种植在每组盆中, 待出苗后留4株旺盛苗. 一次浇透, 采取少量多次浇水, 确保有60%~70%的土壤持水量, 防止浇水过量导致重金属随水流失, 待盆栽植物发芽露出土面生长4个月后开始测定其抗氧化酶和其他生理指标, 6个月后全部收获.
|
|
表 2 盆栽试验土壤梯度设置/mg·kg-1 Table 2 Setting gradient of soil for pot cultivation/mg·kg-1 |
(2)野外大田验证试验设计 在室内模拟土培盆栽胁迫响应特征研究的基础上, 选取宝鸡某冶炼厂厂界外围3个不同区域, 每个区域设置约5 m2作为一个试验区, 并翻挖平整, 清理杂草及植物根系、石块等, 进行野外大田验证观测试验, 研究目标植物对Pb、Zn和Cd复合重金属污染的富集效果. 将切好段的聚合草根段和蓖麻种子按5行×5列种植, 行列间距约为25~30 cm. 观测试验完全依靠野外自然条件和环境生长, 不做任何人为干扰. 试验区自然土壤重金属污染状况见表 3.
|
|
表 3 大田试验区域土壤重金属含量1)/mg·kg-1 Table 3 Heavy metal content in soil in field experiment area/mg·kg-1 |
1.3.2 测定项目及方法
植物叶绿素含量的测定:95%的乙醇提取法, 用分光光度计分别在波长665、649和470 nm下测定吸光度[21, 22], 具体操作过程见文献[22], 叶绿素含量计算方法如下:
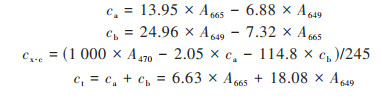
|
按照上式分别计算得到叶绿素a、叶绿素b、类胡萝卜素和叶绿素总浓度ca、cb、cx·c和ct后按照下式分别计算各色素含量(用mg·g-1鲜重表示):
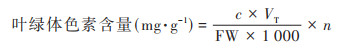
|
式中, c为叶绿体色素的浓度(mg·L-1);FW为鲜重(g);VT为提取液总体积(mL);n为稀释倍数.
酶的测定:植物的超氧化物歧化酶(SOD)、过氧化氢酶(CAT)、过氧化物酶(POD)、丙二醛(MDA)以及脯氨酸等5种酶的测定按照文献[22, 23]的试验方法进行测定.
本研究中植物叶绿素含量和5种酶的测定均通过随机采集土培盆栽试验各梯度中植物活体上新鲜植物叶片进行测定, 叶片采摘时避开植株尖端嫩叶和底部老叶区.
植物样品中重金属总量:利用Nexion 350电感耦合等离子体质谱仪(ICP-MS, 珀金埃尔默Perkin Elmer, 美国), 土壤样品中重金属总量的测定利用岛津AA-6800型原子吸收分光光度计进行测定.
植物重金属转移系数(translocation factor, TF)和富集系数(bioconcentration factors, BCF)的计算方法为[24, 25]:
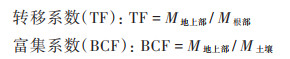
|
式中, M为重金属含量, mg·kg-1.
1.4 数据处理测定结果用Excel 2003、Origin 7.5和SPSS10.0对其结果进行统计分析.
2 结果与分析 2.1 潜在土著超富集植物筛选 2.1.1 研究区优势植物种类通过野外采集调查发现, 在重金属含量比较高的矿区及其尾矿库上生长旺盛、分布频率较高的植物以菊科植物最多, 其次是禾本科植物, 主要有菊科植物13种, 禾木科植物8种, 蓼科植物3种, 藜科、茄科和荨麻科植物各2种, 紫草科、豆科、苋科和木贼科等其它10科植物各1种, 总计40种, 见表 4.
|
|
表 4 本研究采集的植物种类 Table 4 Plant species collected in this study |
2.1.2 优势植物对复合重金属的富集特征
国际上重金属超富集植物的筛选主要通过富集量、富集系数(BCF)和转移系数(TF)这3个指标来判断, 且3个指标缺一不可.
本研究调查阶段从研究区域共采集了40种植物, 320个植物样品, 根据采集植物的地上部分重金属含量的测定结果, 剔除部分重金属含量较低的植物, 选择对Pb、Zn和Cd复合重金属有较高耐性的前10种植物(见表 5). 结合重金属超富集植物的限值要求, 单从植物地上部分对重金属富集量而言, 聚合草地上部分对Pb、Zn和Cd的富集量(mg·kg-1)分别可达到(1 385.82 ± 8.65)(> 1 000)、(13 764.81 ± 14.53)(> 10 000)和(338.05 ± 7.21)(> 100), 即聚合草同时能满足Pb、Zn和Cd各自超富集植物富集量(mg·kg-1)指标的限制(Pb富集量 > 1 000、Zn富集量 > 10 000、Cd富集量 > 100)要求. 水蒿、雪里蕻、芭茅、刺儿菜、土大黄、反枝苋、栝楼和商陆这8种植物地上部分对Cd的富集量也达到Cd超富集量限值要求. 另外, 雪里蕻虽然对锌的富集量不能满足超富集植物对锌富集量指标的限值要求, 但其地上部分对锌的富集量也高达(5 437.17 ± 3.51)mg·kg-1.
|
|
表 5 植物样品体内地上部分和地下部分重金属富集量1) Table 5 Enrichment content of heavy metals in aboveground and underground parts of plant samples |
重金属超富集植物另外两个不可少的判断指标是BCF和TF. BCF即富集系数, 是反映植物对沉积物中重金属的富集能力的指标, 而TF为转移系数, 反映的是植物体内重金属的吸收、转运和分布能力, 由植物体内不同的细胞、组织及器官对重金属物质吸收、运输和分布能力所决定. BCF值越大其吸收富集重金属的能力就越强;TF值越大, 吸收、转运重金属的能力就越强. 本研究前10种植物对Pb、Zn和Cd的BCF和TF计算结果见表 6, 所采集的植物中, 仅聚合草对Pb、Zn和Cd的BCF值和TF值均大于1;另外, 雪里蕻和水蒿对Zn和Cd的BCF和TF值均大于1;刺儿菜对Zn的BCF和TF值均大于1;龙葵对Pb的BCF和TF值均大于1. 根据超富集植物指标限值要求, 结合对重金属富集量的情况, 筛选出比较有应用价值的Pb、Zn和Cd复合超富集植物为聚合草. 聚合草为丛生型多年生草本, 高30~90 cm, 叶大而肥厚, 富含蛋白质和纤维素等, 耐寒、抗旱且抗病虫害, 适应性广, 生物量大且产量高, 是一种优质高产畜禽饲料作物, 若能作为重金属超富集植物, 可克服现有超富集植物的许多不足之处. 同时, 选择本地生长繁殖旺盛、生物量大且具有经济价值的土著植物蓖麻作为供试植物, 进行土培盆栽试验, 对其富集机制进一步探索.
|
|
表 6 植物样品对Pb、Zn和Cd富集与转移系数1) Table 6 Heavy metal enrichment and transfer coefficient of plant samples for Pb, Zn, and Cd |
2.2 Pb、Zn和Cd复合重金属胁迫下目标植物相应生理特征 2.2.1 Pb、Zn和Cd复合胁迫对聚合草光合色素的影响
不同梯度Pb、Zn和Cd复合重金属对聚合草叶中叶绿素a、叶绿素b、类胡萝卜素和总叶绿素含量的影响如图 1. 随着Pb、Zn和Cd复合重金属胁迫梯度的提升, 聚合草叶绿素a和b以及总叶绿素含量均表现出先降低, 当胁迫梯度大于等于Ⅳ时, 则呈现升⁃降⁃升的波动状态, 说明逆境胁迫超过了耐受阈值使其叶绿素系统紊乱;而类胡萝卜素均低于CK, 且随重金属复合胁迫梯度的增加较为稳定, 无明显变化特征;此外, 叶绿素a、叶绿素b和总叶绿素含量在不同Pb、Zn和Cd复合重金属胁迫梯度处理下显著性差异不明显, 部分梯度之间无显著性差异, 类胡萝卜素含量在各处理梯度之间无显著性差异. 另外, 试验结果显示, 在不同Pb、Zn和Cd复合重金属胁迫下, 聚合草均呈现出:叶绿素a含量 > 叶绿素b含量 > 类胡萝卜素含量;且随Pb、Zn和Cd复合重金属胁迫梯度的变化, 叶绿素a的含量最为敏感, 变化较大, 其次是叶绿素b的含量, 而类胡萝卜素含量受Pb、Zn和Cd复合重金属胁迫的变化影响较小.

|
不同处理之间不同小写字母表示各处理间差异显著(P < 0.05), 相同字母表示不显著, 下同 图 1 Pb、Zn和Cd复合重金属对聚合草光合色素的影响 Fig. 1 Effect of Pb, Zn, and Cd on photosynthesis pigments in the leaves of Symphytum officinale L. |
由蓖麻叶片光合色素响应特征可知(图 2), 在一定的Pb、Zn和Cd复合重金属胁迫梯度范围内, 随着胁迫梯度的增加, 蓖麻叶片叶绿素a、叶绿素b、类胡萝卜素和总叶绿素含量总体呈现逐渐升高的趋势, 但是当胁迫大于等于Ⅴ梯度时, 开始下降, 随后又升高, 说明蓖麻叶绿素呈现“低促高拟”特征, 当植物体内富集重金属的量超过植物耐受阈值时, 会激发植物体内其他酶保护系统, 降低了重金属对植物的毒害;而类胡萝卜素含量随重金属复合梯度的增加其值较为稳定, 无明显变化;另外, 各处理梯度之间叶绿素a、叶绿素b、类胡萝卜素和总叶绿素含量均存在显著性差异(P < 0.05);此外, 在不同梯度Pb、Zn和Cd复合重金属胁迫下, 均表现出:叶绿素a含量 > 叶绿素b含量 > 类胡萝卜素含量.

|
图 2 Pb、Zn和Cd复合重金属对蓖麻光合色素的影响 Fig. 2 Effect of Pb, Zn, and Cd on photosynthesis pigments in the leaves of Ricinus communis L. |
Pb、Zn和Cd复合胁迫下聚合草和蓖麻SOD酶响应特征显示(表 7), 与空白对照样相比, 随着Pb、Zn和Cd复合重金属污染胁迫增加, 聚合草和蓖麻SOD酶活性[U‧(g·h)-1]变化范围分别为(11 420.27 ± 1.45)~(38 221.66 ± 2.62)和(11 947.88 ± 77.81)~(21 665.00 ± 126.00);两者叶片中SOD酶活性均呈现先增加后降低再增加的变化趋势. 此外, 不同梯度Pb、Zn和Cd复合重金属污染胁迫下聚合草和蓖麻的SOD酶活性均存在显著性差异(P < 0.05).
|
|
表 7 不同梯度Pb、Zn和Cd复合重金属污染对聚合草和蓖麻SOD酶活性的影响1)/U·(g·h)-1 Table 7 Effects of Pb, Zn, and Cd composite heavy metal pollution at different gradients on SOD activity of Symphytum officinale L. and Ricinus communis L./U·(g·h)-1 |
2.2.4 Pb、Zn和Cd复合胁迫对目标植物CAT酶活性的影响
Pb、Zn和Cd复合胁迫下聚合草和蓖麻叶片中CAT酶响应特征显示(表 8), 相同逆境下聚合草和蓖麻体内CAT酶活性不同, 而且同一植物在不同梯度复合重金属胁迫环境中生长, 其体内CAT酶活性也有不同程度的变化. 在不同梯度Pb、Zn和Cd复合重金属胁迫下, 聚合草和蓖麻叶片中的CAT酶活性[U·(g·min)-1]变化范围分别为(43.663 ± 0.83)~(105.543 ± 1.68)和(2.284 ± 0.12)~(11.321 ± 0.56). 聚合草体内CAT酶活性表现出先升后降的趋势, 而蓖麻体内CAT酶活性呈现逐渐降低的趋势. 另外, 不同梯度Pb、Zn和Cd复合重金属污染胁迫下聚合草和蓖麻的CAT酶活性均存在显著性差异(P < 0.05).
|
|
表 8 不同梯度Pb、Zn和Cd复合重金属污染对聚合草和蓖麻CAT酶活性的影响/U·(g·min)-1 Table 8 Effects of Pb, Zn and Cd composite heavy metal pollution at different gradients on CAT activity of Symphytum officinale L. and Ricinus communis L./U·(g·min)-1 |
2.2.5 Pb、Zn和Cd复合胁迫对目标植物POD酶活性的影响
在不同梯度Pb、Zn和Cd复合重金属污染物胁迫下(表 9), 聚合草和蓖麻植物叶片中POD酶活性[U·(g·min)-1]变化范围分别为(0.247 ± 0.01)~(0.761 ± 0.03)和(4.041 ± 0.06)~(10.088 ± 0.14). 聚合草体内POD酶活性先降后升, 而蓖麻体内POD酶活性呈现先升后降再升的特征, 且不同梯度Pb、Zn和Cd复合重金属污染胁迫下聚合草和蓖麻体内POD酶活性均存在一定的显著性差异(P < 0.05).
|
|
表 9 不同梯度Pb、Zn和Cd复合重金属污染对聚合草和蓖麻POD酶活性的影响/U·(g·min)-1 Table 9 Effects of Pb, Zn, and Cd composite heavy metal pollution at different gradients on POD activity of Symphytum officinale L. and Ricinus communis L./U·(g·min)-1 |
2.2.6 Pb、Zn和Cd复合胁迫对目标植物MDA含量的影响
随着不同梯度Pb、Zn和Cd复合重金属污染胁迫, 植物叶片中MDA含量发生了相应的变化(表 10). 聚合草和蓖麻叶片中MDA含量(mmol·g-1)变化范围分别为(0.680 ± 0.04)~(1.820 ± 0.09)和(1.950 ± 0.05)~(3.060 ± 0.07). 聚合草叶片中MDA含量呈现先降低后略升的特征, 但总体呈现下降趋势;而蓖麻叶片中MDA含量呈先增加后降低再升高特征, 总体呈上升特征, 且不同梯度Pb、Zn和Cd复合重金属污染胁迫下聚合草和蓖麻的MDA含量均存在显著性差异(P < 0.05).
|
|
表 10 不同梯度Pb、Zn和Cd复合重金属污染对聚合草和蓖麻MDA含量的影响/mmol·g-1 Table 10 Effects of Pb, Zn, and Cd composite heavy metal pollution at different gradients on MDA content of Symphytum officinale L. and Ricinus communis L./mmol·g-1 |
2.2.7 Pb、Zn和Cd复合胁迫对目标植物脯氨酸含量的影响
随着Pb、Zn和Cd复合重金属胁迫增加, 聚合草和蓖麻叶片中脯氨酸含量(µg·g-1)变化(表 11)范围分别为:(0.097 ± 0.01)~(0.126 ± 0.02)和(0.200 ± 0.01)~(0.401 ± 0.01). 聚合草叶片中脯氨酸含量呈现逐渐降低, 而蓖麻叶片中脯氨酸含量呈现先升高后降低的趋势;另外, 不同Pb、Zn和Cd复合重金属胁迫梯度之间聚合草和蓖麻的脯氨酸含量均存在显著性差异(P < 0.05).
|
|
表 11 不同梯度Pb、Zn和Cd复合重金属污染对聚合草、蓖麻脯氨酸含量的影响/µg·g-1 Table 11 Effects of Pb, Zn and Cd composite heavy metal pollution at different gradients on praline content of Symphytum officinale L. and Ricinus communis L./µg·g-1 |
2.3 聚合草和蓖麻对Pb、Zn和Cd复合重金属的富集特征
野外大田试验进一步验证了聚合草和蓖麻对Pb、Zn和Cd复合重金属的富集能力, 试验统计结果显示(表 12和表 13), 不同污染程度的3个试验区中聚合草和蓖麻对重金属Pb、Zn和Cd的富集量存在差异, 重金属高污染区富集量较高, 而低污染区的富集量较低, 高污染区聚合草地上部分对Pb、Zn和Cd的富集量同时能达到超富集植物富集量指标限值, 而低污染一区和二区仅对Cd的富集量能达到超富集植物限值要求;聚合草对复合重金属Pb、Zn和Cd的富集能力优于蓖麻, 且富集量大小均为:Zn > Pb > Cd;另外, 不同试验区之间同一重金属富集量之间存在显著性差异(P < 0.05), 而同一试验区不同重金属之间差异性不显著(P ≥ 0.05).
|
|
表 12 聚合草和蓖麻对重金属Pb、Zn和Cd富集量统计/mg·kg-1 Table 12 Enrichment statistics of Symphytum officinale L. and Ricinus communis L. for Pb, Zn, and Cd/mg·kg-1 |
|
|
表 13 聚合草和蓖麻对重金属Pb、Zn和Cd的TF和BCF Table 13 TF and BCF of Symphytum officinale L. and Ricinus communis L. for Pb, Zn, and Cd |
从TF及BCF来看, 3个试验区中聚合草对Pb、Cd和Zn的TF及BCF同时能满足超富集植物限值> 1的要求, 而蓖麻仅部分区域的指标能满足. 同样, 聚合草对复合重金属Pb、Zn和Cd的TF及BCF显示, 不同试验区之间同一重金属TF及BCF之间存在显著性差异(P < 0.05), 而同一试验区不同重金属的TF及BCF之间差异性不显著(P < 0.05).
综合分析, 大田试验显示, 聚合草对Pb、Zn和Cd复合重金属的富集能力可以达到国际上超富集植物指标限值要求, 可被认为是一种Pb、Zn和Cd复合重金属的潜在超富集植物资源;蓖麻对Pb、Zn和Cd复合重金属富集能力虽然不能达到超富集植物指标限值要求, 但从富集量来看, 可以作为Pb、Zn和Cd复合重金属污染土壤修复的备选高富集植物资源.
3 讨论 3.1 Pb、Zn和Cd复合胁迫下植物光合色素的响应叶绿素是光合作用的主要色素, 主要由叶绿素a和叶绿素b等组成, 其含量的高低在一定程度上反映了植物光合作用的强弱, 在植物光合作用中起吸收和转化光能的作用, 通常情况重金属胁迫对植物的光合作用有抑制作用, 且其降低效应与胁迫程度成正相关[26~28]. 本研究中, 在Pb、Zn和Cd复合重金属低污染梯度胁迫下, 随着污染程度的增加, 聚合草叶中叶绿素a、叶绿素b和总叶绿素含量逐渐降低, 说明了Pb、Zn和Cd复合重金属胁迫抑制了光合作用, 当污染梯度大于等于Ⅳ时, 复合重金属的胁迫毒害造成了聚合草光合系统的紊乱, 该研究结果与吴月燕等[27]和石慧芳等[29]的研究结果一致. 而部分植物在重金属胁迫下其叶绿素呈现“低促高抑”现象, 即蓖麻在低梯度复合重金属胁迫下对其光合作用有一定的促进作用, 但是随着胁迫梯度的提高达到其耐受阈值后蓖麻的叶绿素a、叶绿素b和总叶绿素出现了拟制作用, 过高则激发了自身其他酶保护系统使得毒害减轻, 这与权佳惠[30]的研究结果一致.
3.2 Pb、Zn和Cd复合胁迫下植物抗氧化酶的响应SOD、POD和CAT是植物为了应对重金属胁迫造成毒害作用而产生的抗氧化剂, 是植物体应对外界胁迫的重要保护酶系统, 组成了植物体内有效的活性氧清除系统, SOD可以及时有效地清除自由基, POD可催化活性氧等进行氧化分解, CAT能清除植物细胞中的H2O2, 它们均保护了细胞免受氧化胁迫的伤害, 是反映重金属污染胁迫的灵敏指标[31]. 其中SOD是活性氧清除反应过程中第1个发挥作用的酶, 能歧化O2生成H2O2和过氧化物, 以此减少具有毒性的高活性氧化剂羟自由基(·OH)的形成, 而CAT和POD共同作用能把H2O2和过氧化物转化为H2O与分子氧, 3种酶协同作用, 使植物免受自由基损害[27, 32, 33]. 本研究中, 随着Pb、Zn和Cd复合重金属胁迫的增加, 聚合草和蓖麻体内的SOD活性均呈现先增加后降低再增加的趋势. 这主要是由于植物受到复合重金属逆境胁迫后, 活性氧代谢失去平衡造成体内活性氧增加, 从而诱导SOD活性增加, 但随着逆境胁迫的继续增强, SOD活性下降, 当逆境胁迫超过植物承受阈值后SOD系统紊乱, 这与顾艾博[34]的研究结果基本一致. CAT是一种以铁卟啉为辅基的结合酶, 含四聚血红素, 主要位于过氧化物酶体中, 可将2H2O2转化为O2+2H2O, 使细胞内H2O2维持在一个正常水平, 保护细胞膜[35]. 本研究中, 随着Pb、Zn和Cd复合重金属胁迫的增加, 聚合草体内CAT活性表现先升后降的趋势, 呈现低促进效应与高抑制效应, 这与袁红艳[36]的研究结果一致;而蓖麻体内CAT活性呈现持续降低的趋势, 说明蓖麻逆境中耐胁迫能力较弱, 在复合重金属胁迫下, 产生的H2O2消耗了CAT, 同时拟制了其产生. POD是植物抗氧化酶系统的重要组成部分, 它通过过氧化氢促进酚类物质的降解, 可以有效清除植物在氧化应激下产生的ROS和酚类物质[37, 38]. 本研究中, 随着Pb、Zn和Cd复合重金属胁迫的增加, 聚合草体内POD活性呈现略降低后逐渐升高, 这主要是由于, 在较低逆境胁迫下, POD及时启动, 清除植物在氧化应激下产生的ROS和酚类物质而消耗一部分POD, 但随着复合重金属胁迫梯度增加, 植物体内活性氧代谢失去平衡造成体内活性氧增加, 从而诱导POD活性增加, 总体呈现增加趋势, 使得在逆境胁迫下聚合草抵抗代谢过程产生的有害物质对细胞的伤害表现出一定的抗逆性;而蓖麻体内POD则呈现出先升高后降低再升高的特征. 另外, 本研究结果显示在不同含量Pb、Zn和Cd复合重金属污染胁迫下, 聚合草和蓖麻的几种抗氧化酶的酶活性均存在显著性差异(P < 0.05). 总之, 复合重金属逆境胁迫与其他形式的氧化逆境胁迫相似, 会抑制植物体内一些保护酶的活性, 从而导致对植物体内部细胞及其相应功能的损伤, 植物体内的SOD、CAT及POD这三者组成了一个有效的活性氧清除系统, 其值的高低在一定程度上反映了植物耐逆境胁迫能力的高低. 一般情况下, 低浓度的重金属胁迫诱导下保护酶活性会上升, 但当重金属胁迫超过植物承受阈值后, 其体内的活性氧含量超过了正常的歧化能力, 会对植物组织和细胞的多种功能膜及酶系统造成破坏, 反过来抑制了植物体内SOD、POD和CAT这3种酶的抗氧化能力, 使植物细胞受到损害[39].
3.3 Pb、Zn和Cd复合胁迫下植物其他生理指标的响应MDA是植物细胞膜脂过氧化的最终分解产物, 常作为植物细胞膜脂过氧化指标, 反映细胞膜脂过氧化程度和植物对逆境条件反应的强弱[40, 41]. 植物在重金属胁迫下, 会产生大量活性氧化物(ROS), 与细胞膜中的脂肪酸发生过氧化反应, 产生MDA, 对植物体产生破坏性作用[28]. 本研究中, 随着Pb、Zn和Cd复合重金属胁迫量的增加, 聚合草叶片中MDA含量总体呈现下降趋势, 而蓖麻叶片中MDA含量总体呈上升特征;聚合草叶片中MDA含量降低可能是由于植物体内自身防御调节机制, 削弱了膜脂过氧化所带来的损伤, 缓解了植物体在重金属胁迫下受到的膜脂过氧化作用[28, 42]. 而蓖麻叶片中MDA含量上升, 主要是由于随着Pb、Zn和Cd复合重金属胁迫的增加, 复合胁迫所诱导的氧自由基造成了脂质过氧化物水平的相应升高, 形成了更多的膜脂过氧化物, 表现为MDA含量升高, 这与张义贤等[43]的研究结果是一致的, 这也说明了聚合草的抗胁迫能力较强.
脯氨酸是植物体内细胞质中一种少有的游离氨基酸, 也是植物细胞内部主要的细胞渗透调节物质, 其主要成分是蛋白质. 在重金属胁迫中, 脯氨酸的作用是多方面的, 但主要是抗氧化胁迫方面的作用, 作为一种抗氧化剂保护细胞避免因受重金属胁迫而产生自由基的毒害[44]. 本研究中, 随着Pb、Zn和Cd复合重金属胁迫的增加, 蓖麻叶片中脯氨酸含量先升高后降低, 这表明在一定范围内, 蓖麻能通过增加游离脯氨酸来抵抗复合重金属的胁迫, 但胁迫过高可能会进一步破坏蓖麻的细胞膜系统, 导致其应激能力降低, 因此游离脯氨酸含量有降低的现象, 该研究结果与刘朝荣等[45]对重金属胁迫下的珙桐幼苗研究结果一致;有研究表明植物体内脯氨酸积累是由于植物对胁迫的反应[46], 当胁迫解除后, 脯氨酸含量立即下降, 聚合草叶片中脯氨酸含量随着复合重金属胁迫的增加逐渐降低, 表明聚合草中脯氨酸在复合重金属胁迫条件下拟制了胁迫产生的毒害作用, 有效地调节渗透平衡, 维持蛋白质、膜和亚细胞结构的稳定, 清除活性氧等对植物细胞的伤害, 说明聚合草具有一定的抗逆性.
3.4 聚合草和蓖麻对Pb、Zn和Cd复合重金属的富集和转移能力重金属超富集植物不仅要求富集量满足限值要求, 同时TF和BCF均要大于1, 二者的值越大, 说明植物地上部分越容易转移和富集累积重金属. 本研究中聚合草源于野外重金属污染区域, 由于其具有超富集潜力而作为本研究的目标植物. 经土培盆栽试验对复合重金属胁迫和其他生理指标响应研究显示, 其对Pb、Zn和Cd复合重金属胁迫具有一定的抗逆性和耐受性;野外大田试验结果显示, 虽然采集的聚合草样品对Pb、Zn和Cd的富集量未能全部满足超富集植物限值的要求, 但是相对中高污染区的70%的样品富集量达到了超富集植物富集量的限制要求, 且TF和BCF均大于1, 所以聚合草是潜在的Pb、Zn和Cd复合重金属的超富集植物, 可以作为Pb、Zn和Cd重金属污染土壤修复的优选植物.
4 结论(1)通过对宝鸡市典型铅锌矿区、浮选厂和冶炼厂区域调查采集的40种植物, 320个植物样品和320个土壤样品进行室内检测分析, 筛选出聚合草对Pb、Zn和Cd的富集量、TF和BCF均满足国际上对超富集植物限值的要求, 是一种潜在的Pb、Zn和Cd复合重金属的超富集植物, 雪里蕻地上对Zn的富集量虽未达到超富集限值, 但也高达(5 437.17 ± 3.51)mg·kg-1, 可认为是一种潜在的Zn高富集植物.
(2)盆栽试验结果可知, 随着Pb、Zn和Cd复合重金属胁迫梯度的增加:聚合草光合色素受到抑制使得叶片中叶绿素a、叶绿素b、总叶绿素含量逐渐降低, 当胁迫梯度大于等于Ⅳ时, 胁迫毒害导致聚合草光合系统紊乱;蓖麻叶片中的叶绿素呈现“低促高抑”现象, 但过高则激发了自身其他酶保护系统使得毒害减轻. 此外, 各胁迫梯度之间聚合草叶片中类胡萝卜素含量无显著性差异(P ≥ 0.5), 而其它光合色素指标含量有一定的显著性差异(P < 0.5);各胁迫梯度之间蓖麻叶片中光合色素各指标含量存在显著性差异(P < 0.5).
(3)植物抗氧化酶研究结果显示, 随着Pb、Zn和Cd复合重金属胁迫梯度的增加, 聚合草和蓖麻体内的SOD酶活性均呈现先增加后降低再增加的趋势;聚合草体内CAT酶活性表现先升后降的趋势, 呈现“低促进和高抑制”效应, 而蓖麻体内CAT酶活性呈现持续降低的趋势;聚合草体内POD酶活性先降低后逐渐升高, 总体呈现增加趋势;蓖麻则体内POD酶活性呈现先升后降低再升高的特征. 另外, 各Pb、Zn和Cd复合重金属胁迫梯度之间, 聚合草和蓖麻的SOD、CAT和POD这3种酶活性均存在显著性差异(P < 0.05).
(4)其他生理指标研究显示, 随着Pb、Zn和Cd复合重金属胁迫梯度的增加:聚合草叶片中MDA含量总体呈现下降趋势, 而蓖麻叶片中MDA含量总体呈上升特征;聚合草叶片中脯氨酸含量呈现逐渐降低, 而蓖麻叶片中脯氨酸含量呈现先升高后降的趋势, 且各Pb、Zn和Cd复合重金属胁迫梯度之间, 聚合草和蓖麻叶片中的MDA含量和脯氨酸含量均存在显著性差异(P < 0.05).
(5)大田试验结果显示, 聚合草对Pb、Zn和Cd的富集能力可以达到超富集植物各项指标限值要求, 是一种潜在的Pb、Zn和Cd复合重金属的超富集植物种质资源, 可以作为Pb、Zn和Cd重金属污染土壤修复的优选植物.
| [1] |
康乐, 彭鑫波, 马延龙, 等. 兰州市耕地表层土壤重金属的积累特征及其影响因素分析[J]. 环境科学, 2023, 44(3): 1620-1635. Kang L, Peng X B, Ma Y L, et al. Accumulation characteristics and influencing factors of heavy metals in cultivated land surface soil in Lanzhou[J]. Environmental Science, 2023, 44(3): 1620-1635. |
| [2] |
胡杰, 赵心语, 王婷婷, 等. 太原市汾河河岸带土壤重金属分布特征、评价与来源解析[J]. 环境科学, 2022, 43(5): 2500-2509. Hu J, Zhao X Y, Wang T T, et al. Distribution characteristics, evaluation, and source analysis of heavy metals in soils of Fenhe riparian zone in Taiyuan city[J]. Environmental Science, 2022, 43(5): 2500-2509. |
| [3] |
李军, 李旭, 李开明, 等. 黄河流域农田土壤重金属污染特征及其优先控制源分析[J]. 环境科学, 2024, 45(3): 1724-1738. Li J, Li X, Li K M, et al. Characteristics and identification priority source of heavy metals pollution in farmland soils in the Yellow River Basin[J]. Environmental Science, 2024, 45(3): 1724-1738. |
| [4] |
周艳, 万金忠, 李群, 等. 铅锌矿区玉米中重金属污染特征及健康风险评价[J]. 环境科学, 2020, 41(10): 4733-4739. Zhou Y, Wan J Z, Li Q, et al. Heavy metal contamination and health risk assessment of corn grains from a Pb-Zn mining area[J]. Environmental Science, 2020, 41(10): 4733-4739. |
| [5] |
杨滨娟, 黄国勤. 植物种植修复土壤重金属污染的模式、技术与效果综述[J]. 生态科学, 2022, 41(4): 251-256. Yang B J, Huang G Q. Review on model, technology and effect of phytoremediation technology on remediation of heavy metal pollution[J]. Ecological Science, 2022, 41(4): 251-256. |
| [6] |
赵晓峰, 雷梅, 陈同斌. 中国镉超富集植物的物种、生境特征和筛选建议[J]. 环境科学, 2023, 44(5): 2786-2798. Zhao X F, Lei M, Chen T B. Species, habitat characteristics, and screening suggestions of cadmium hyperaccumulators in China[J]. Environmental Science, 2023, 44(5): 2786-2798. |
| [7] |
李红霞, 林文波, 李玉庆, 等. 植物修复重金属污染沉积物的模型仿真及验证[J]. 环境科学, 2011, 32(7): 2119-2124. Li H X, Lin W B, Li Y Q, et al. Simulation and verification for model of phytoremediation on heavy metal contaminated sediment[J]. Environmental Science, 2011, 32(7): 2119-2124. |
| [8] |
仪小梅. 国土空间生态修复背景下的重金属污染土壤植物修复及联合技术研究进展[J]. 现代化工, 2023, 43(9): 76-79, 84. Yi X M. Research progress on phytoremediation combination technology for heavy metals-contaminated soil[J]. Modern Chemical Industry, 2023, 43(9): 76-79, 84. |
| [9] |
逯秋源, 樊丽, 姚芸, 等. 植物联合修复土壤重金属污染的研究进展[J]. 四川环境, 2023, 42(4): 319-327. Lu Q Y, Fan L, Yao Y, et al. Research progress on phytoremediation of heavy metal contaminated soils[J]. Sichuan Environment, 2023, 42(4): 319-327. |
| [10] |
林小兵, 何绍浪, 吴多基, 等. 不同程度镉污染农田中粉葛镉富集转运特征[J]. 环境科学, 2023, 44(10): 5769-5778. Lin X B, He S L, Wu D J, et al. Characteristics of cadmium concentration and transport of Pueraria thornsonii in farmland with different cadmium pollution degrees[J]. Environmental Science, 2023, 44(10): 5769-5778. |
| [11] |
何孟轲, 郭俊娒, 杨俊兴, 等. 典型铅锌企业周边农田重金属富集植物筛选[J]. 中国生态农业学报(中英文), 2023, 31(6): 954-966. He M K, Guo J M, Yang J X, et al. Screening of accumulating plants in farmland surrounding typical lead and zinc smelting enterprises[J]. Chinese Journal of Eco-Agriculture, 2023, 31(6): 954-966. |
| [12] |
张军, 董洁, 梁青芳, 等. 宝鸡市区土壤重金属污染影响因子探测及其源解析[J]. 环境科学, 2019, 40(8): 3774-3784. Zhang J, Dong J, Liang Q F, et al. Heavy metal pollution characteristics and influencing factors in Baoji Arban soils[J]. Environmental Science, 2019, 40(8): 3774-3784. |
| [13] |
何婷婷. 超高含量重金属复合污染土壤中东南景天修复机制研究[D]. 武汉: 武汉理工大学, 2020. He T T. Study on the remediation mechanism of Sedum alfredii in the soil polluted by ultra-high content heavy metals[D]. Wuhan: Wuhan University of Technology, 2020. |
| [14] |
邓忠, 王健, 赵胜军, 等. 广西梧州大黎镇利达铅锌矿区土壤重金属污染评价[J]. 有色金属(矿山部分), 2023, 75(4): 155-160. Deng Z, Wang J, Zhao S J, et al. Evaluation of soil heavy metal pollution in Lida lead-zinc mining area of Dali town, Wuzhou, Guangxi[J]. Nonferrous Metals (Mine Section), 2023, 75(4): 155-160. |
| [15] |
郭松明, 余海波, 袁龙义. 近20年我国重金属超积累植物种质资源筛选研究进展[J]. 生态毒理学报, 2022, 17(2): 96-108. Guo S M, Yu H B, Yuan L Y. Research progress of screening of germplasm resources of heavy metal hyperaccumulator in recent 20 years in China[J]. Asian Journal of Ecotoxicology, 2022, 17(2): 96-108. |
| [16] |
邓焱, 郝喆, 曹明杰, 等. 尾矿库植物生长与重金属含量的相互关系研究[J]. 能源环境保护, 2020, 34(1): 8-14. Deng Y, Hao Z, Cao M J, et al. Study on the relationship between plant growth andheavy metal content in tailings reservoirs[J]. Energy Environmental Protection, 2020, 34(1): 8-14. |
| [17] |
沈锋. 典型铅锌冶炼区农田土壤重金属污染及植物化学联合修复研究[D]. 杨凌: 西北农林科技大学, 2017. Shen F. Heavy metal pollution of farmland soil in a typical Pb/Zn smelting area and phyto-chemistry combined remediation[D]. Yangling: Northwest A & F University, 2017. |
| [18] |
刘智峰. 陕西凤县铅锌冶炼区土壤重金属污染特征及根际促生菌强化植物修复研究[D]. 杨凌: 西北农林科技大学, 2019. Liu Z F. Research on pollution characteristics and enhanced phytoremediation with PGPR of heavy metal contaminated soil in lead-zinc smelting area in Fengxian County, Shaaxi Province[D]. Yangling: Northwest A & F University, 2019. |
| [19] |
周小康, 李维成, 董王仓, 等. 陕西省铅锌矿资源特征及时空分布规律概论[J]. 西北地质, 2020, 53(3): 127-139. Zhou X K, Li W C, Dong W C, et al. A preliminary study of the characteristics of lead-zinc deposits in Shaanxi Province and their distribution laws[J]. Northwestern Geology, 2020, 53(3): 127-139. |
| [20] |
范拴喜. 宝鸡市长青镇冶炼厂周边居住区土壤重金属污染特征与风险评估[J]. 环境污染与防治, 2015, 37(9): 46-54. Fan S X. Pollution and health risk assessment of heavy metals in soil neighborhoods around a smelter in Changqing town of Baoji city[J]. Environmental Pollution and Control, 2015, 37(9): 46-54. |
| [21] |
磨春丽, 谷战英, 黄鹏, 等. 一品红对重金属锌胁迫的耐受性及其富集能力[J]. 西南农业学报, 2023, 36(2): 234-240. Mo C L, Gu Z Y, Huang P, et al. Tolerance and accumulation ability of poinsettia to zinc stress[J]. Southwest China Journal of Agricultural Sciences, 2023, 36(2): 234-240. |
| [22] |
高俊凤. 植物生理学实验指导[M]. 北京: 高等教育出版社, 2006. Gao J F. Experimental guidance for plant physiology[M]. Beijing: Higher Education Press, 2006. |
| [23] |
王学奎, 黄见良. 植物生理生化实验原理和技术[M]. (第3版). 北京: 高等教育出版社, 2015. Wang X K, Huang J L. Principles and techniques of plant physiological biochemical experiment[M]. 3rd ed. Beijing: Higher Education Press, 2015. |
| [24] |
陶理, 王沛芳, 袁秋生, 等. 4种典型沉水植物对去除镉污染底泥的应用效果[J]. 环境科学, 2021, 42(9): 4311-4318. Tao L, Wang P F, Yuan Q S, et al. Application effect of four typical submerged macrophytes on removing cadmium from polluted sediment[J]. Environmental Science, 2021, 42(9): 4311-4318. |
| [25] | Waris M, Baig J A, Talpur F N, et al. Estimation of phytoextraction potential of selected halophytes for accumulation of heavy metals from wetland saline soil[J]. Rendiconti Lincei. Scienze Fisiche e Naturali, 2023, 34(2): 553-562. |
| [26] |
刘倩钰. 铬镉铅复合胁迫下紫金牛的生理响应研究[D]. 长沙: 中南林业科技大学, 2021. Liu Q Y. The research on physiological response of Ardisia japonica under Cr, Cd and Pb combined stress[D]. Changsha: Central South University of Forestry and Technology, 2021. |
| [27] |
吴月燕, 陈赛, 张燕忠, 等. 重金属胁迫对5个常绿阔叶树种生理生化特性的影响[J]. 核农学报, 2009, 23(5): 843-852. Wu Y Y, Chen S, Zhang Y Z, et al. Effect of heavy metal stress on physiological and biochemical characteristics of five evergreen broad-leaved trees[J]. Journal of Nuclear Agricultural Sciences, 2009, 23(5): 843-852. |
| [28] |
左明博, 高天鹏, 万子栋, 等. 复合重金属污染对矿区植物生理指标的影响[J]. 兰州大学学报(自然科学版), 2022, 58(3): 404-411. Zuo M B, Gao T P, Wan Z D, et al. Effects of compound heavy metal pollution on the physiological indexes of plants in mining areas[J]. Journal of Lanzhou University (Natural Sciences), 2022, 58(3): 404-411. |
| [29] |
石慧芳, 席溢, 张茂, 等. 铅、锌、镉单一及复合胁迫对多年生黑麦草生长及生理特性的影响[J]. 南方农业学报, 2021, 52(4): 1030-1039. Shi H F, Xi Y, Zhang M, et al. Effects of single and combined stresses of lead, zinc and cadmium on growth and physiological characteristics of perennial ryegrass[J]. Journal of Southern Agriculture, 2021, 52(4): 1030-1039. |
| [30] |
权佳惠. 贝克喜盐草对Zn、Cd胁迫的生理响应[D]. 南宁: 广西大学, 2022. Quan J H. Physiological response of Halophila beccarii to Zn and Cd stress[D]. Nanning: Guangxi University, 2022. |
| [31] |
彭雪婷. 水淹及重金属Cd胁迫对丰都车前(Plantago fengdouensis)根叶结构和生理生化特性的影响[D]. 重庆: 西南大学, 2021. Peng X T. Effects on root-leaf structure and physiological biochemical properties of Plantago fengdouensis under flooding and heavy metal Cd stress[D]. Chongqing: Southwest University, 2021. |
| [32] |
杨雯一. 不同胁迫对各种植物体内抗氧化酶系统的影响综述[J]. 化工管理, 2021(1): 92-93. Yang W Y. A review of the effects of different stresses on antioxidant enzyme systems in various plants[J]. Chemical Engineering Management, 2021(1): 92-93. |
| [33] |
张秀娟, 范玟, 杨乐, 等. 镉胁迫对3种草坪草生理特性及叶片超微结构的影响[J]. 草地学报, 2023, 31(9): 2663-2670. Zhang X J, Fan W, Yang L, et al. Effects of cadmium stresson physiological activities and leaf ultrastructures of three turfgrass species[J]. Acta Agrestia Sinica, 2023, 31(9): 2663-2670. |
| [34] |
顾艾博. 铬、铜胁迫对2种玉簪生长及生理特性的影响[D]. 长春: 吉林农业大学, 2014. Gu A B. Cr, Cu stress on the influence of two Hosta growth and physiological characteristics[D]. Changchun: Jilin Agricultural University, 2014. |
| [35] |
陈孚尧. 类芦修复矿山土壤重金属污染场地的效果与机理研究[D]. 徐州: 中国矿业大学, 2022. Chen F Y. Study on the mechanism and effects of heavy metal remediation with Neyraudia reynaudiana in contaminated soil in mining site[D]. Xuzhou: China University of Mining and Technology, 2022. |
| [36] |
袁红艳. Cu2+、Pb2+、Zn2+胁迫对费菜耐性及积累特性的研究[D]. 苏州: 苏州大学, 2010. Yuan H Y. Studies on tolerance and accumulation of Sedum aizoon L. to Cu2+, Pb2+, Zn2+ stress[D]. Suzhou: Soochow University, 2010. |
| [37] |
万子栋, 高天鹏, 周玉霞, 等. 重金属复合胁迫下碱蓬萌发生长及富集特征[J]. 生物工程学报, 2020, 36(3): 493-507. Wan Z D, Gao T P, Zhou Y X, et al. Seed germination, bud growth and heavy-metal accumulation of Suaeda salsa [J]. Chinese Journal of Biotechnology, 2020, 36(3): 493-507. |
| [38] | Thounaojam T C, Panda P, Mazumdar P, et al. Excess copper induced oxidative stress and response of antioxidants in rice[J]. Plant Physiology and Biochemistry, 2012, 53: 33-39. |
| [39] |
范拴喜. 几种土著植物对Pb、Cd、Zn复合污染的响应及微观机制研究[D]. 杨凌: 西北农林科技大学, 2023. Fan S X. Responses and icro-mechanism of compound of Pb, Cd, Zn on some native plants[D]. Yangling: Northwest Agriculture and Forestry University, 2023. |
| [40] |
孙天国, 沙伟, 刘岩. 复合重金属胁迫对两种藓类植物生理特性的影响[J]. 生态学报, 2010, 30(9): 2332-2339. Sun T G, Sha W, Liu Y. The effects of compound heavy metal stress on some physiological characteristics of two moss species[J]. Acta Ecologica Sinica, 2010, 30(9): 2332-2339. |
| [41] |
王小平, 华春, 吴向华, 等. Cr3+胁迫对油菜幼苗抗氧化酶系统和MDA的影响[J]. 湖北农业科学, 2011, 50(8): 1537-1539. Wang X P, Hua C, Wu X H, et al. Effect of Cr3+ stress on antioxidant enzyme and MDA of rape seedlings[J]. Hubei Agricultural Sciences, 2011, 50(8): 1537-1539. |
| [42] |
赵晶怡. 9种园林树木抗旱性生理指标的测定及评价[D]. 临汾: 山西师范大学, 2012. Zhao J Y. Studies and on several physiological indexes of the drought-resistance of nine species of gardening trees and comprehensive evaluation[D]. Linfen: Shanxi Normal University, 2012. |
| [43] |
张义贤, 张丽萍. 重金属对大麦幼苗膜脂过氧化及脯氨酸和可溶性糖含量的影响[J]. 农业环境科学学报, 2006, 25(4): 857-860. Zhang Y X, Zhang L P. Effects of heavy metals on membrane lipid peroxidation, proline and soluble sugar in roots of Hordeum vulgare [J]. Journal of Agro-Environment Science, 2006, 25(4): 857-860. |
| [44] |
徐超. 脯氨酸与重金属耐性和富集的研究进展[J]. 中国资源综合利用, 2018, 36(2): 80-83. Xu C. A review of amino acid in heavy metal tolerance and accumulation[J]. China Resources Comprehensive Utilization, 2018, 36(2): 80-83. |
| [45] |
刘朝荣, 张柳青, 杨艳, 等. 珙桐幼苗生理生化指标对重金属铅、镉胁迫的响应[J]. 广西植物, 2021, 41(9): 1401-1410. Liu C R, Zhang L Q, Yang Y, et al. Effects of lead and cadmium on physiology and biochemical indexes of Davidia involucrata seedlings[J]. Guihaia, 2021, 41(9): 1401-1410. |
| [46] |
张浩, 陆宁, 钱晓刚, 等. 不同类型土壤重金属胁迫对烟叶脯氨酸含量的影响[J]. 贵州农业科学, 2014, 42(1): 127-131. Zhang H, Lu N, Qian X G, et al. Effects of different concentrations of heavy metals on tobacco proline content in four soil types[J]. Guizhou Agricultural Sciences, 2014, 42(1): 127-131. |