 2024, Vol. 45
2024, Vol. 45



2. 同济大学环境科学与工程学院, 污染控制与资源化研究国家重点实验室, 上海 200092
2. State Key Laboratory of Pollution Control and Resources Reuse, School of Environmental Science and Engineering, Tongji University, Shanghai 200092, China
微塑料(MPs)通常是指尺寸小于5 mm的塑料颗粒碎片[1], 目前, 微塑料被定义为国际上受广泛关注的四大新污染物之一[2], 由于微塑料的广泛使用和其难以降解的特性, 微塑料污染问题已然成为全球环境保护的重要问题[3]. 微塑料不仅广泛存在于空气、水体、沉积物、冰和雪中, 在人体中也发现了微塑料的存在. Leslie等[4]首次在人体血液中检测到了微塑料的污染, 在近80%的实验受试者样本中均发现了微塑料的存在. Aves等[5]首次在南极洲的新降雪中发现了微塑料. 截至目前, 陆地及水生生态系统中存在的MPs已经对其生态环境的健康构成威胁, 环境中遍布的MPs已经成为全球性的环境保护问题[6].
微塑料可分为初级微塑料和次生微塑料, 环境中初级微塑料的主要来源为家庭和工业生产制造的原材料, 常见的初级微塑料有牙膏和化妆品等含有塑料滚珠的个人护理产品, 温室材料以及塑料覆盖物, 例如农业生产中经常运用到的地膜等塑料制品的不规范处理, 经过生物或者非生物过程变成尺寸更小的微塑料, 这是环境中次生微塑料的主要来源[7]. 环境中常见的微塑料种类主要有聚乙烯(PE)、聚丙烯(PP)、聚氯乙烯(PVC)和聚苯乙烯(PS)等[8], 微塑料通过本身有毒物质的释放以及吸附环境中化合物的能力对生态环境的健康造成威胁.
由于微塑料难降解的物理特性, 其在土壤沉积物中的累计含量会逐渐增加, 土壤沉积物系统及其微生物环境会受到直接或间接的危害[9]. 微塑料由于其较小的分子特性, 很容易在淡水生态系统中富集, 并随着食物链在各个营养级之间进行传递. 微塑料通过降水沉降, 废水灌溉等方式流入土壤以及沉积物之中, 微塑料对土壤的理化性质, 例如透气性、肥力、微生物活性和pH等产生影响[10]. 微塑料在自然降解过程中, 会不断地释放双酚a、多溴二苯醚和邻苯二甲酸盐等有毒污染物, 直接影响土壤沉积物以及植物根际微生物群落, 改变其群落结构和微生物代谢通路等功能[11]. 沉积物在水体中充当储存或者释放各种化合物的媒介, 沉积物与水体营养交换发生在沉积物-水界面, 因此, 微塑料对沉积物的作用也间接影响水体生理状况[12]. 小粒径的微塑料还会堵塞植物根孔以及与外界交换的通道, 影响其生长状况. Lian等[13]研究表明, 在微塑料的胁迫下, 植物的生理性状、光合作用机制和抗氧化代谢都受到影响. 肖甫[14]的研究表明, 在高浓度(2, 5 μg·mL-1)的纳米氨基-聚苯乙烯颗粒(PS-NH2)与磺酸基修饰的聚苯乙烯(PS-SO3H)胁迫作用下, 对浮萍的生物量增长均有抑制作用, 且引起了显著的氧化应激反应. 可以看出, 微塑料对淡水生态系统中的植物产生一定的负面影响. 然而, 淡水生态系统除了水生植物还包含水体以及沉积物等两大基本要素, 而微生物群落又是沉积物中的重要组成部分, 目前的研究多集中于微塑料对于水生植物的影响, 对于淡水生态系统中的水质以及微生物群落结构等进行全面分析的报道较少.
Scanes等[15]的研究表明, 微塑料可通过迁移转化等途径进入海洋软体动物体内;微塑料因其较大的比表面积, 可作为各种污染物的载体, Rout等[16]的研究表明, 微塑料的自然老化开裂和生物膜的形成可以增强放射性碘的吸附;影响海底生态系统功能[17]. 然而目前对于微塑料的热点研究多集中在对海洋生态系统的影响, 对于淡水生态系统的研究较少. 因此研究淡水生态系统中的微塑料对其生态风险评价极其有意义. PE-MPs对水-苦草-沉积物系统中水质理化指标、沉水植物形态特征、生理性状、抗氧化系统和沉积物中微生物群落结构的研究结果可为水生态系统中微塑料的生态风险评价提供基础数据和科学依据.
1 材料与方法 1.1 样品收集与制备本实验土壤沉积物以及水体采集自上海海洋大学校湖(东经121°54'5.451″, 北纬30°53'23.373″), 海拔5.15 m. 土壤采集后, 暂存在聚乙烯收纳箱内, 并立即运回实验室. 采集到的土壤经过60目的不锈钢筛子筛除底栖生物、植物细根和残体等物质. 校湖采集的水样使用孔径0.45 μm的滤膜进行抽滤. 苦草幼苗购自上海某园艺公司, 用0.5%次氯酸钠溶液表面消毒10 min, 用超纯水冲洗5次, 在实验装置中驯养3 d用以适应实验室条件. PE-MPs(80目粒径)购自河南某商贸有限公司, 样品用紫外线进行灭菌处理.
1.2 实验设计为了避免塑料容器对实验的干扰, 选用玻璃容器(长30 cm, 宽18 cm, 高20 cm)作为实验装置, 容器在使用前经纯水仔细冲洗. 每个玻璃容器内随机选择并放置相等间距的8株苦草(无分株, 高度为10 cm), 初始生物量平均SD为0.165 g. 沉积物厚度为4 cm, 上覆水深度为15 cm.
PE-MPs共分为0%、1%、2%和5%(沉积物湿重质量分数)4个质量分数梯度, 每组设置3个平行组, 添加PE-MPs后与底泥进行均匀搅拌. 本实验在室温下进行(23℃ ± 2℃), 光照强度为8 500 lux, 光照周期为亮光12 h/暗光12 h.
1.3 样品分析 1.3.1 水体理化指标分析在第3、6、9、12、15、18和21 d使用便携式多参数水质分析仪(HQ40 d, HACH)对上覆水进行理化性质分析(盐度、pH、溶解氧和电导率), 以及测定了水体的总氮(TN)、总磷(TP)、氨氮(NH4+-N)和高锰酸盐指数.
1.3.2 苦草的形态和生理性状分析在实验开始与结束时测量植物的株高根长和鲜重, 用以量化其实验周期的生长. 使用宝如亿(北京)生物技术有限公司叶绿素(Chlorophyll)检测试剂盒(比色法-RL7812)测定叶绿素a和b的含量, 称取0.1 g叶片研磨匀浆. 叶绿素用试剂盒中的提取粉剂和Chlorophyll Assay Buffer进行提取, 避光放置直至组织残渣接近白色, 即为提取完全. 用分光光度计在665 nm和649 nm处测量吸光度. 用如下公式计算叶绿素含量.
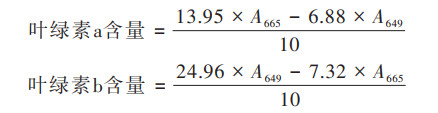
|
植物在遭受环境胁迫时会在体内产生活性氧自由基(ROS), 对植物体内的蛋白质产生破坏作用, 损伤细胞膜, 进而影响植物的生长发育. 抗氧化酶系统是植物体内在受胁迫条件下激发的保护酶系统, 能够消除植物体内多余的活性氧自由基, 从而达到保护的作用[18]. 丙二醛(MDA)含量反映出植物质膜损伤的程度, 植物体内的抗氧化酶主要有过氧化氢酶(CAT)、超氧化物歧化酶(SOD)和过氧化物酶(POD). 采用市售试剂盒(MDA-R21870、CAT-A007-1-1、SOD-A001-4-1和POD-RL7652)测定了植物中MDA、CAT、SOD和POD的含量.
1.3.4 沉积物及苦草根际附着微生物群落多样性分析采用2组重复的沉积物以及植物根际样品进行混合处理. 在实验结束时, 运用五点取样法采集沉积物样品, 采集到的样品在无菌密封袋中-80℃保存. 将植物带根拔起, 抖落根周围松散的泥土, 剪取约15 g的带土根系置于灭菌的含有0.86% NaCl溶液的25 mL离心管中, 将离心管放在冰中每隔5 min取出摇匀1次, 总计30 min. 去除离心管中植株根系, 转速4 000 r·min-1, 4℃低温离心30 min, 然后去除上清液, 将沉淀物保存在无菌EP管中, -80℃保存. DNA提取和高通量测序由上海派森诺生物科技有限公司(中国上海)进行. 采用Illumina平台对群落DNA片段进行双端(Paired-end)测序. 引物对338F(ACTCCTACGGGA GGCAGCA)和806R(GGACTACHVGGGTWTCTAAT)扩增V3-V4, 运用Illumina MiSeq平台进行微生物群落结构的分析, 并分析沉积物与根系微生物群落组成.
1.4 数据统计与分析对每组变量进行shaprio-wilk test正态性检验, 运用单因素方差分析(one-way ANOVA)和非参数检验(nonparametric or mixed)进行统计处理, 其中P < 0.05时, 组间存在显著差异. 使用Excel 2016作为数据记录软件, GraphPad Prism9作为作图以及统计分析软件.
2 结果与分析 2.1 水体理化指标对PE-MPs的响应苦草的存在会影响水体理化指标, 微塑料的存在会改变苦草的生理性状, 从而间接影响到水质的变化. 在不同微塑料浓度处理条件下水体盐度, 电导率的变化如图 1所示. 在21 d的实验期间, 盐度与电导率呈现出的大致趋势为:2%PE-MPs处理组为最高, 1%PE-MPs处理组比0% PE-MPs对照组要高, 5% PE-MPs处理组的盐度与电导率始终低于对照组的状况. 且2% PE-MPs处理组与5% PE-MPs处理组差异极显著, 盐度在第18 d最大相差30%, 电导率在第18 d与第21 d最大相差27.8%. 微塑料处理组对溶解氧与pH有一定的影响, 如图 1(e)~1(h)所示, 数值均比空白对照组高, 但无显著性差异.

|
**表示P < 0.01 图 1 水体理化指标变化情况 Fig. 1 Changes in physical and chemical indexes of water body |
本实验开始前后的TN数据如图 2(a)所示, 实验结束后各组对TN均有50%以上的去除效果, 1% PE-MPs处理组去除效率最高达62.4%. NH4+-N的去除情况与TN类似, 各实验组对NH4+-N的去除率均超过70%, 表明水-苦草-沉积物系统对NH4+-N的去除效果较理想, 0% PE-MPs对照组对NH4+-N的去除效果最佳, 为81.7%的去除率, 其余受PE-MPs胁迫的各组去除率受到影响均低于对照组, 5% PE-MPs处理组的去除率最低. 相比之下, TP与高锰酸盐指数的变化不明显, 2% PE-MPs处理组对TP的去除率达48.0%, 其余组均没有很好的去除率, 0% PE-MPs对照组实验结束后的TP浓度不降反升. 高锰酸盐指数在实验开始与结束时的变化不明显, 整个水-苦草-沉积物系统对于高锰酸盐指数没有显著的影响作用.

|
图 2 实验开始与结束时水质变化情况 Fig. 2 Changes in water quality at the beginning and end of the experiment |
PE-MPs对苦草的株高和根长的影响如图 3(a)和3(b)所示, 各PE-MPs处理组的株高均受到显著影响, 与对照组差异均极显著. 1%、2%和5% PE-MPs处理组的株高增长仅为对照组的47.44%、33.33%和59.62%. 其中, 2% PE-MPs处理组株高的增长受到最严重的抑制, 其次为1% PE-MPs处理组. 5% PE-MPs处理组的根长增长为对照组的162.50%, 高浓度的PE-MPs有利于根系的生长;而低浓度的PE-MPs则对根系的生长起到抑制作用, 1%和2% PE-MPs处理组中植物根长的增长仅为对照组的68.75%和81.25%.

|
*表示P < 0.05, **表示P < 0.01, ***表示P < 0.001, ****表示P < 0.0001 图 3 PE-MPs对植物生理性状以及叶绿素的影响 Fig. 3 Effects of PE-MPs on plant physiological characters and chlorophyll |
微塑料分别对苦草的总生物量和相对生长率的影响结果如图 3(c)和3(d)所示, 1%和2% PE-MPs处理组总生物量的变化与对照组有明显差异, 经过实验周期总生物量的增长仅为对照组的38.39%和38.76%, 与株高以及相对生长率的受抑制情况一致, 低浓度的PE-MPs更能抑制总生物量的增长.
图 3(e)和3(f)的结果表明, 不同浓度的PE-MPs对植物叶片中的叶绿素含量也会有影响. 实验周期结束后, 测定植物叶片中叶绿素a和叶绿素b的含量. 结果表明, 1% PE-MPs处理组中叶绿素a的合成受到明显的抑制, 叶绿素a含量为对照组的81.04%, 其余处理组叶绿素a含量均高于对照组, 叶绿素b也呈现出相同的规律.
2.3 苦草氧化应激指标和抗氧化酶系统对PE-MPs的响应植物受胁迫时产生的过氧化氢(H2O2)可以被CAT分解为水和氧气, 保护细胞质膜不受伤害, 减轻植物体内自由基对大分子物质如DNA、蛋白质的伤害[19]. 如图 4(b)结果显示, 1% PE-MPs处理组CAT活性明显升高, 较对照组升高了233.70%. 2%和5% PE-MPs处理组较对照组分别升高了95.73%和60.92%. 在为期21 d的实验周期内, 从图 4(a)中可看出, 苦草抗氧化酶系统中的CAT活性呈现出先升高再降低而后升高的趋势, 整体酶活性表现为有波动地上升.

|
*表示P < 0.05, **表示P < 0.01, ***表示P < 0.001 图 4 PE-MPs对苦草抗氧化酶系统的影响 Fig. 4 Effect of PE-MPs on antioxidant enzyme system of Vallisneria natans |
MDA是细胞膜脂质过氧化的产物, 通过测定MDA含量可以判断出逆境条件下膜脂过氧化反应的程度, MDA含量越高, 植物体内积累的H2O2越多, 过量的H2O2会对细胞质膜产生严重的损伤. 图 4(d)结果表明, 1%和2% PE-MPs处理组较对照组皆具有明显的差异, MDA含量分别较对照组升高了117.82%和114.98%, 5% PE-MPs处理组与对照组无明显统计学差异. 结果表明, 在低浓度PE-MPs的刺激下MDA含量的升高更为敏感. 如图 4(c)所示, 整体MDA含量跟随实验周期均呈现出整体向上的趋势, 1%和2% PE-MPs处理组的升高尤为明显.
POD的作用与CAT类似, POD通过将H2O2作为电子受体来进一步催化底物的氧化, 增强将H2O2转化为水的过程, 降低过量H2O2对植物脂膜过氧化的伤害.
如图 4(f)所示, 1%和2% PE-MPs处理组与对照组差异极显著, 其POD活性分别上升了61.62%和42.22%, 结果与MDA类似, 均为低浓度处理组的反应更敏感. 5% PE-MPs处理组比之对照组上升了19.30%, 无明显统计学差异. 实验周期内的趋势变化如图 4(e)所示, 总体趋势为先急速上升, 回落后稳步上升.
SOD是植物抗氧化酶系统的第一道防线, 能够催化超氧自由基发生歧化反应产生O2和H2O2, 从而去除植物体内的活性氧(ROS), 从而保护植物细胞膜的正常功能与机制[19]. 如图 4(h)所示, 2% PE-MPs处理组与对照组有显著差异, SOD活性上升了65.35%, 1%和5% PE-MPs处理组与对照组相比分别提升了53.75%和36.48%. 实验周期内的趋势变化如图 4(g)所示, 总体呈现出先升高再缓慢稳定降低的趋势.
2.4 沉积物及苦草根际微生物对PE-MPs添加的响应如图 5所示, 不同浓度PE-MPs的添加对底泥微生物的影响较小, 根系微生物群落结构和底泥微生物群落结构差异性不显著. 变形菌门(Proteobacteria)占总体细菌的50%以上, 为优势主导菌. 在植物根际微生物群落中, 放线菌门(Actinobacteria)随着微塑料浓度的增加呈现出减少的趋势, 零浓度对照组最高, 5%PE-MPs处理组最低. 在底泥微生物群落中, 放线菌门则呈现出完全相反的规律, 底泥中PE-MPs浓度越高, 放线菌占比越高.

|
A0、A1、A2和A5为0%、1%、2%和5% PE-MPs处理组中底泥微生物结构群落;B0、B1、B2和B5为0%、1%、2%和5% PE-MPs处理组中植物根际微生物结构群落, 下同 图 5 PE-MPs处理组层次聚类分析 Fig. 5 Hierarchical cluster analysis of PE-MPs treatments |
图 6为物种组成热图, 可以看出, 底泥与植物根际微生物群落优势种占比呈现完全不同的状况, 绿弯菌门、放线菌门、厚壁菌门、黏胶球形菌门、螺旋菌门和酸杆菌门在底泥微生物群落结构中占主导地位, 疣微菌门、变形菌门、拟杆菌门和硝化螺旋菌门在植物根际微生物群落结构中占主导地位, 表明底泥中的微生物群落与植物根际的微生物群落结构大不相同.

|
图 6 PE-MPs处理组物种组成热图 Fig. 6 Thermal map of species composition of PE-MPs treatment |
不同浓度的PE-MPs对植物的生理性状产生影响, 微塑料的存在严重影响植株的生长, 如图 3(a)所示, 微塑料对沉水植物株高的影响有极其显著的差异. 其原因可以归结为以下3方面:①微塑料可以通过静电吸附力的作用附着在沉水植物的根茎叶表面, 影响植物与外界物质交换和代谢过程, 粒径较小的微塑料颗粒能直接堵塞植物体表的气孔, 影响光合作用速率, 抑制其生长[20]. ②微塑料颗粒由于拥有较大比表面积的特性, 很容易作为水中细菌病原体以及抗生素等有毒污染物的载体, 通过吸附作用累积毒素直接影响植物的生长发育[21]. ③淡水沉积物由于污水排放等原因积聚微塑料从而容易形成污染热点区域, 微塑料通过影响底泥微生物的氮循环而间接对植物根系产生影响[12]. 在本研究中, PE-MPs处理组显著降低了植物的株高, 对根长、总生物量和叶绿素a的生成起到抑制作用. 且1% PE-MPs处理组苦草的受抑制程度均比其他对照组高, 从另一方面表明, 在低浓度PE-MPs的刺激下, 苦草对于微塑料胁迫的响应更为敏感[1].
3.2 PE-MPs对苦草抗氧化系统的影响本实验所测3种酶活和1种氧化应激指标属于植物抗氧化系统的一部分, 实验所得的数据结论可以大致理解为对植物抗氧化系统的影响. 植物所处环境无污染, 能够正常生长发育的情况下, 植物的抗氧化系统和体内的活性氧(ROS)达成动态平衡以此维持正常发育. 当所处环境对植物造成胁迫时, 植物体内的抗氧化系统会消除体内增多的ROS以免对植物本身造成一定的毒害[22]. 从图 4可看出, 低浓度处理组如1%和2% PE-MPs处理组对3种酶和MDA的响应更为迅速和敏感, 低浓度处理组能够更好地激发植物抗氧化系统以应对自身所受到的胁迫效应. 当PE含量增加到5%泥沙湿重时, 植物所产生的抗氧化酶并不能完全消除体内产生的大量ROS[23], 植物生长受到抑制反过来影响抗氧化系统的运作, 抗氧化酶活性和氧化应激指标显著低于低浓度处理组, 说明5% PE-MPs处理组的植物中的MPs超过了胁迫阈值, 导致其应对外界变化的能力减弱, 植物即将发生凋零[24]. 在为期21 d的实验周期内, 除了SOD以外, POD和CAT的活性以及MDA含量均呈现逐步上升的趋势, 表明在微塑料胁迫下能够产生较好的应对策略, 只有SOD在实验的中段左右开始出现了下降的趋势, 由于SOD是抗氧化系统的第一道防线[25], 推测可能是因为1% PE-MPs已经远远大于环境中的微塑料浓度, 植物受到较为强烈的刺激, 在一开始催化超氧自由基发生歧化反应时并不能完全消除植物体内多余的ROS, 植物在处于长时间和高强度的胁迫下不能对SOD的缺失做到及时补充, 另外因为SOD合成结构遭到破坏, 导致SOD含量逐渐减少[26], 陈晶莹等[26]对铜绿微囊藻做的研究结果也是SOD活性均先升高后降低, 与本文的研究结果一致.
3.3 PE-MPs对沉积物及苦草根际微生物群落结构的影响PE-MPs能改变沉积物中微生物群落结构, 与对照组相比, PE-MPs处理组的放线菌门和绿弯菌门丰度明显增加, 而占优势地位的变形菌门以及酸杆菌门则呈现丰度减少的状况. Sun等[27]的研究表明, 经过PBS和PLA改良的土壤微生物组表现出更高的吸收外源性碳水化合物和氨基酸的潜力, 但是会降低相关代谢功能的能力. 研究指出常规微塑料处理组与无微塑料对照组在代谢组学上无显著差异, 可能的原因是可生物降解颗粒对促进土壤微生物的多功能性起到积极的作用, 在有水分的条件下, PBS聚合物主链中的酯键通过水解降解[28], 同样PLA聚合物中的酯键也能通过水解并形成水溶性低分子量低聚物, 这可作为微生物同化的额外碳源而影响其群落结构的改变[29, 30], 而PE和PS微塑料对土壤微生物的功能多样性没有影响, 与本实验的结果一致. 微塑料颗粒还参与生物地球化学循环, Su等[30]评估了微塑料颗粒在河口生物与非生物反硝化和硝化作用中的潜力, 微塑料颗粒表面生物膜的形成提供了有利于反硝化菌形成的缺氧条件, 使得微塑料颗粒具有更高的反硝化活性. Seeley等[31]的研究结果表明, 微塑料可作为微生物群落的有机碳基质, 不同的微塑料可显著影响沉积物中氮循环的过程, PUF和PLA改性的沉积物相较于无微塑料对照组在硝化反硝化过程中起促进作用, 而PVC改性的沉积物则对硝化反硝化过程起到抑制作用. 微塑料其较大比表面积的特性决定了其能够为硅藻、异养细菌和真菌等提供生态位, 在水环境中驱动生物地球化学循环[32].
4 结论(1) 不同PE-MPs处理组对系统中的盐度与电导率存在不同程度的影响, 且处理组之间的盐度和电导率存在显著差异. PE-MPs处理组的溶解氧与pH均比对照组高, 但处理组之间无显著性差异, 受PE-MPs胁迫的各组对NH4+-N的去除率均低于对照组.
(2) PE-MPs对苦草的株高、根长、总生物量和相对生长率存在不同程度的抑制或促进作用. PE-MPs影响叶片叶绿素的含量, 1% PE-MPs处理组中叶绿素含量低于对照组, 其余处理组叶绿素含量均高于对照组. PE-MPs显著影响苦草抗氧化酶系统, 在实验周期内, 处理组中的MDA含量、CAT、POD和SOD活性较对照组均有不同程度的上升.
(3) PE-MPs的添加对底泥微生物的影响较小, 根系微生物和底泥微生物群落结构中的放线菌门对PE-MPs的响应呈现相反的规律. 底泥与植物根际微生物群落优势种占比存在显著差异.
| [1] | Zhang J W, Huang D Y, Deng H, et al. Responses of submerged plant Vallisneria natans growth and leaf biofilms to water contaminated with microplastics[J]. Science of the Total Environment, 2022, 818. DOI:10.1016/j.scitotenv.2021.151750 |
| [2] | Tang Y Q, Liu Y G, Chen Y, et al. A review: Research progress on microplastic pollutants in aquatic environments[J]. Science of the Total Environment, 2021, 766. DOI:10.1016/j.scitotenv.2020.142572 |
| [3] | Zhou J, Wen Y, Marshall M R, et al. Microplastics as an emerging threat to plant and soil health in agroecosystems[J]. Science of the Total Environment, 2021, 787. DOI:10.1016/j.scitotenv.2021.147444 |
| [4] | Leslie H A, Van Velzen M J M, Brandsma S H, et al. Discovery and quantification of plastic particle pollution in human blood[J]. Environment International, 2022, 163. DOI:10.1016/j.envint.2022.107199 |
| [5] | Aves A R, Revell L E, Gaw S, et al. First evidence of microplastics in Antarctic snow[J]. The Cryosphere, 2022, 16(6): 2127-2145. DOI:10.5194/tc-16-2127-2022 |
| [6] | Corinaldesi C, Canensi S, Dell'Anno A, et al. Multiple impacts of microplastics can threaten marine habitat-forming species[J]. Communications Biology, 2021, 4(1). DOI:10.1038/s42003-021-01961-1 |
| [7] |
施庆还, 林子增, 季钰浩. 环境中微塑料的来源、转移机制及降解方法研究[J]. 应用化工, 2022, 51(11): 3327-3332. Shi Q H, Lin Z Z, Ji Y H. Study on the source, transfer mechanism and degradation methods of microplastics in the environment[J]. Applied Chemical Industry, 2022, 51(11): 3327-3332. DOI:10.3969/j.issn.1671-3206.2022.11.043 |
| [8] |
宋亚丽, 李帅斌, 李紫燕, 等. 水体中微塑料种类、来源、影响与处理方法[J]. 塑料, 2023, 52(2): 72-76, 133. Song Y L, Li S B, Li Z Y, et al. Types, sources, effect and removal of microplastic in aqueous environment[J]. Plastics, 2023, 52(2): 72-76, 133. |
| [9] | Ya H, Jiang B, Xing Y, et al. Recent advances on ecological effects of microplastics on soil environment[J]. Science of the Total Environment, 2021, 798. DOI:10.1016/j.scitotenv.2021.149338 |
| [10] | Zhao T T, Lozano Y M, Rillig M C. Microplastics increase soil pH and decrease microbial activities as a function of microplastic shape, polymer type, and exposure time[J]. Frontiers in Environmental Science, 2021, 9. DOI:10.3389/fenvs.2021.675803 |
| [11] | Meng J, Xu B L, Liu F, et al. Effects of chemical and natural ageing on the release of potentially toxic metal additives in commercial PVC microplastics[J]. Chemosphere, 2021, 283. DOI:10.1016/j.chemosphere.2021.131274 |
| [12] | Yu H W, Liu M M, Gang D, et al. Polyethylene microplastics interfere with the nutrient cycle in water-plant-sediment systems[J]. Water Research, 2022, 214. DOI:10.1016/j.watres.2022.118191 |
| [13] | Lian J P, Wu J N, Xiong H X, et al. Impact of polystyrene nanoplastics (PSNPs) on seed germination and seedling growth of wheat (Triticum aestivum L.)[J]. Journal of Hazardous Materials, 2020, 385. DOI:10.1016/j.jhazmat.2019.121620 |
| [14] |
肖甫. 浮萍对纳米聚苯乙烯颗粒的吸收与积累机制[D]. 济南: 山东大学, 2020. Xiao P. Absorption and accumulation mechanism of duckweed on nano-polystyrene particles[D]. Jinan: Shandong University, 2020. |
| [15] | Scanes E, Wood H, Ross P. Microplastics detected in haemolymph of the Sydney rock oyster Saccostrea glomerata [J]. Marine Pollution Bulletin, 2019, 149. DOI:10.1016/j.marpolbul.2019.110537 |
| [16] | Rout S, Yadav S, Joshi V, et al. Microplastics as vectors of radioiodine in the marine environment: A study on sorption and interaction mechanism[J]. Environmental Pollution, 2022, 307. DOI:10.1016/j.envpol.2022.119432 |
| [17] | Ladewig S M, Coco G, Hope J A, et al. Real-world impacts of microplastic pollution on seafloor ecosystem function[J]. Science of the Total Environment, 2023, 858. DOI:10.1016/j.scitotenv.2022.160114 |
| [18] | Dvořák P, Krasylenko Y, Zeiner A, et al. Signaling toward reactive oxygen species-scavenging enzymes in plants[J]. Frontiers in Plant Science, 2021, 11. DOI:10.3389/fpls.2020.618835 |
| [19] | 王磊. 沉水植物苦草(Vallisneria natans)对水位变化的生理生态适应性及机制研究[D]. 南京: 南京师范大学, 2021. |
| [20] | Abduro Ogo H, Tang N, Li X W, et al. Combined toxicity of microplastic and lead on submerged macrophytes[J]. Chemosphere, 2022, 295. DOI:10.1016/j.chemosphere.2022.133956 |
| [21] | Wang L, Gao Y X, Jiang W, et al. Microplastics with cadmium inhibit the growth of Vallisneria natans (Lour.) Hara rather than reduce cadmium toxicity[J]. Chemosphere, 2021, 266. DOI:10.1016/j.chemosphere.2020.128979 |
| [22] | Liang R H, Xua M L, Cao Y, et al. Effect of water level fluctuations on the antioxidant enzyme system and the degree of cell membrane peroxidation in Vallisneria natans [J]. Applied Ecology and Environmental Research, 2022, 20(6): 5251-5262. DOI:10.15666/aeer/2006_52515262 |
| [23] | Chang W J, Zhu X X, Sun J L, et al. Effects of lead pollution on bacterial communities in biofilm attached to submerged plants[J]. Water Science and Technology, 2022, 86(6): 1358-1372. DOI:10.2166/wst.2022.279 |
| [24] | 毛鸿志. 不同粒径微塑料与四环素及其联合作用对沉水植物及其群落的影响[D]. 武汉: 湖北大学, 2022. |
| [25] | Xia W T, Zhu B, Qu X, et al. Effects of salinity on sprouting and growth of three submerged macrophytes[J]. Ecohydrology, 2020, 13(7). DOI:10.1002/eco.2235 |
| [26] |
陈晶莹, 周炜杰, 林果, 等. 沉水态圆叶节节菜的化感抑藻作用[J]. 浙江农林大学学报, 2023, 40(4): 765-772. Chen J Y, Zhou W J, Lin G, et al. Allelopathic and algal inhibition effects of submerged aquatic plant Rotala rotundifolia [J]. Journal of Zhejiang A & F University, 2023, 40(4): 765-772. |
| [27] | Sun Y Z, Duan C X, Cao N, et al. Biodegradable and conventional microplastics exhibit distinct microbiome, functionality, and metabolome changes in soil[J]. Journal of Hazardous Materials, 2022, 424. DOI:10.1016/j.jhazmat.2021.127282 |
| [28] | Gigli M, Negroni A, Soccio M, et al. Influence of chemical and architectural modifications on the enzymatic hydrolysis of poly(butylene succinate)[J]. Green Chemistry, 2012, 14(10): 2885-2893. DOI:10.1039/c2gc35876j |
| [29] | Elsawy M A, Kim K H, Park J W, et al. Hydrolytic degradation of polylactic acid (PLA) and its composites[J]. Renewable and Sustainable Energy Reviews, 2017, 79: 1346-1352. DOI:10.1016/j.rser.2017.05.143 |
| [30] | Su X X, Yang L Y, Yang K, et al. Estuarine plastisphere as an overlooked source of N2O production[J]. Nature Communications, 2022, 13(1). DOI:10.1038/s41467-022-31584-x |
| [31] | Seeley M E, Song B, Passie R, et al. Microplastics affect sedimentary microbial communities and nitrogen cycling[J]. Nature Communications, 2020, 11(1). DOI:10.1038/s41467-020-16235-3 |
| [32] | Rogers K L, Carreres-Calabuig J A, Gorokhova E, et al. Micro-by-micro interactions: How microorganisms influence the fate of marine microplastics[J]. Limnology and Oceanography Letters, 2020, 5(1): 18-36. DOI:10.1002/lol2.10136 |