 2024, Vol. 45
2024, Vol. 45

2. 河南农业大学国家小麦工程研究中心, 郑州 450046;
3. 河南农业大学林学院, 郑州 450046
 , JIAO Qiu-juan1 , XU Zheng-yang1 , FAN Li-na1 , JIANG Ying1 , SONG Jia1 , HUA Dang-ling1 , LI Ge-zi2 , LIN Di3 , LIU Hai-tao1
, JIAO Qiu-juan1 , XU Zheng-yang1 , FAN Li-na1 , JIANG Ying1 , SONG Jia1 , HUA Dang-ling1 , LI Ge-zi2 , LIN Di3 , LIU Hai-tao1
2. National Engineering Research Center for Wheat, Henan Agricultural University, Zhengzhou 450046, China;
3. College of Forestry, Henan Agricultural University, Zhengzhou 450046, China
随着冶矿产业的发展、人们长期不合理地使用含镉(Cd)农用化学品以及污水灌溉等, 导致我国土壤Cd污染形势严峻[1]. Cd是植物非必需的元素, 有剧毒, 且土壤中的Cd易被作物吸收并通过食物链进入人体, 进而威胁人类健康[2, 3]. 已有的研究表明, 高含量的Cd会导致植株发生一系列生理毒害反应, 抑制光合作用和叶绿素合成[4, 5]、干扰能量供应和代谢过程[6]、破坏细胞内氧化还原稳态、引起氧化损伤、扰乱正常的细胞结构和功能[7, 8], 最终使得植株生长发育受阻、生物量减少、甚至萎黄死亡[9, 10].
Qin等[8]研究发现, 高Cd胁迫下小麦幼苗的生长受到严重抑制, 植株的光合参数、类胡萝卜素、根鲜重及可溶性蛋白含量显著降低, 而丙二醛(MDA)及可溶性糖含量随着Cd浓度的增加而降低. Liu等[7]研究发现, Cd胁迫可显著降低小麦幼苗的株高、地上部生物量、叶绿素a、叶绿素b和总叶绿素含量, 类胡萝卜素含量反而升高. 小麦(Triticum aestivum L.)是世界上广泛种植的口粮作物, 也是我国第二大谷类作物, 是人类蛋白质、能量摄入的主要来源之一[7, 11]. 然而, 相对于玉米和水稻而言, 小麦更易富集累积Cd在其籽粒中, 这主要是由于其将Cd从根系转运到茎叶的能力更强[7, 12]. 因此, 缓解Cd胁迫对小麦生长的影响、提高小麦抗Cd胁迫的能力已成为保障粮食安全的重要课题. 目前, 提高小麦抗Cd胁迫的方法主要有:农艺措施如水分管理或施用调理剂等[1]、遗传育种的手段如筛选耐Cd或Cd低积累小麦品种[12]、外源物质提高幼苗抗Cd能力[7]等, 其中, 外源施用化学调控物质因具有效率高且成本低等特点而被广泛应用.
壳聚糖(chitosan, CTS)是自然界中唯一的天然碱性多糖, 可从甲壳类(虾、蟹)动物中提取获得, 因具有环境友好、价廉和无毒可降解等特点被广泛用于农业生产中[13~15]. Ahmad等[16]研究表明, 壳聚糖可显著提高柠檬草的株高、根长、干重等, 促进其生长. Rahman等[17]研究表明, 壳聚糖可提高草莓叶片中类胡萝卜素含量, 增强其光合作用. 王聪等[18]研究表明, 外源壳聚糖可显著降低NaCl胁迫下菜用大豆叶绿体H2O2的含量, 提高其抗氧化酶活性. 曲丹阳等[13]研究证实, 外源壳聚糖可提高玉米幼苗的抗氧化能力, 缓解Cd胁迫对玉米幼苗的生长抑制. 然而, 目前将壳聚糖作为外源物质应用到小麦抗Cd胁迫领域的研究尚不多见. 因此, 本试验通过水培法研究Cd胁迫下壳聚糖对小麦幼苗生长特征、光合特性、抗氧化酶活性和Cd积累特征的影响, 并结合偏最小二乘回归模型确定影响幼苗Cd积累的关键因子指标, 加深对壳聚糖缓解小麦幼苗Cd毒性机制的认识, 以期为壳聚糖应用于降低植物Cd毒害提供理论基础和技术参考.
1 材料与方法 1.1 供试材料与试验设置供试冬小麦品种为百农207, Cd和壳聚糖源分别为氯化镉(CdCl2)和羧化壳聚糖(CAS 9012-76-4, 阿拉丁), 营养液为霍格兰(Hoagland)营养液. 本试验共设置3个Cd水平(0 µmol·L-1、10 µmol·L-1和25 µmol·L-1)和两个CTS水平(0 mg·L-1和100 mg·L-1), 共5个处理:0 µmol·L-1 Cd + 0 mg·L-1 CTS(对照组, 只添加营养液)、10 µmol·L-1 Cd + 0 mg·L-1 CTS、25 µmol·L-1 Cd + 0 mg·L-1 CTS、10 µmol·L-1 Cd + 100 mg·L-1 CTS、25 µmol·L-1 Cd + 100 mg·L-1 CTS, 分别记作CK、Cd10、Cd25、Cd10 + CTS、Cd25 + CTS. 本试验于河南农业大学人工气候室内进行, 每个处理设置3个重复.
1.2 幼苗培养试验选取颗粒饱满、大小均一的小麦种子, 首先用5%H2O2消毒15 min后, 用去离子水冲洗4~5次, 于25℃遮光催芽24 h后放入育苗盘并转入温室培养, 待种子出芽1 cm左右转入植物光照培养箱. 待种子长出一叶一心后, 将长势一致的幼苗移栽到含有2 L营养液的水培盆中. 为使小麦幼苗更好地适应水培环境, 移栽后先用1/8浓度的营养液进行培养, 再依次逐渐增加至1/4、1/2和完全浓度的营养液. 期间, 每3 d更换一次营养液, 待幼苗长至两叶一心时对其进行处理. 保持人工气候室相对湿度60%~65%, 昼夜时间16 h/8 h, 昼夜温度25℃/20℃, 光照强度400 μmol·(m2·s)-1. 在处理后的第14 d采集幼苗样本并分离幼苗植株的地上部和地下部. 采样前, 利用便携式光合仪(LI-6400, 美国)测定植株的光合参数. 将根系于20 mmol·L-1的Na2-EDTA溶液中浸泡15 min, 用去离子水反复冲洗4~5次, 将一部分鲜样装入信封中, 放入烘箱杀青30 min(105℃), 再于低温(70℃)烘干至恒重, 用于测定根系与地上部的生物量, 并取适量样品粉碎备用;另一部分鲜样用液氮进行处理并储存在冰箱(-80℃)中, 用于生理生化指标的测量.
1.3 生理生化指标测定取样前, 利用便携式光合仪(Li-6400, LICOR Inc., 美国)对小麦植株顶部的第一片完全展开叶进行叶片气体参数的测量:每个处理选3株, 光照强度控制在1 000 µmol·(m2·s)-1、温度控制在25℃, 测定指标包括净光合速率(net photosynthetic rate, Pn)、蒸腾速率(transpiration rate, Tr)、胞间CO2浓度(intercellular CO2 concentration, Ci)和气孔导度(stomatal conductance, Gs). 采用根系扫描仪(V700 PHOTO, Epson, 日本)测定小麦幼苗根系的总根长(total root length, RL)、根表面积(root surface area, SA)、根体积(root volume, RV)、根平均直径(root average diameter, RD)和总根尖数(root tips, RT). 采用95%乙醇提取法测定叶片叶绿素含量[19];采用硫代巴比妥酸比色法测定丙二醛(malondialdehyde, MDA)含量[20, 21];采用硫酸钛比色法测定过氧化氢(H2O2)含量[22];采用氮蓝四唑(NBT)还原法测定超氧化物歧化酶(superoxide dismutase, SOD)活性[21];参照愈创木酚法测定过氧化物酶(peroxidase, POD)活性[23].
1.4 Cd含量的测定烘干粉碎后的植株样用硝酸∶高氯酸= 3∶1(体积比)进行消解, 用原子吸收光谱仪(PinAAcle 900T, 美国)测定样品中的Cd含量. 为保证试验数据的可靠性, 植株Cd含量测定过程中插入国家标准植物样品GBW07604(GSV-1)进行质量控制, 加标回收率均在87%~113%之间.
1.5 数据分析为研究小麦各表型性状及生理生化指标对Cd积累量的影响程度, 采用偏最小二乘回归法(partial least square, PLS)分别建立地上部Cd积累量、地下部Cd积累量及总Cd积累量的回归方程, 并根据PLS回归方程中的无量纲变量重要性投影值指标(variable importance for projection, VIP)来确定影响Cd积累量的关键因子指标. PLS模型集相关分析、多元线性回归分析和主成分分析为一体, 不仅可有效解决自变量间的多重共线性问题, 还可有效降低因子分析维度, 从多维自变量中寻求影响因变量的关键主控因子[24, 25]. VIP值可直观、定量地反映各自变量在预测因变量时的重要程度, 通常其临界值为1.0, 值越高, 表明该因子的敏感度和影响力越强[25, 26]. 耐受指数反映植株在逆境环境中生长的抑制程度[27].

|
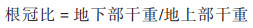
|
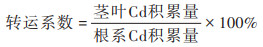
|
采用Microsoft Excel 2016和SPSS statistics 27. 0对数据进行统计分析;采用最小显著极差法(least significant difference, LSD, P < 0.05)进行方差分析(analysis of variance, ANOVA);利用SIMCA-P 11.5进行偏最小二乘回归(partial least square, PLS)分析和选择关键因子;并采用Origin Pro 2018和Adobe Illustrator 2019软件绘图.
2 结果与分析 2.1 外源壳聚糖对镉胁迫下冬小麦幼苗生物量和耐受指数的影响由表 1可知, 与对照(CK)相比, Cd10处理下冬小麦幼苗根长、茎叶长、根系干重和茎叶干重分别降低了46.47%、21.10%、49.84%和63.52%, 差异显著;Cd25处理下冬小麦幼苗根长、茎叶长、根系干重和茎叶干重分别降低了64.75%、35.58%、59.17%和68.22%, 差异也显著. 这些结果表明Cd胁迫显著抑制了冬小麦幼苗的正常生长, 且Cd25处理对冬小麦幼苗的抑制作用强于Cd10处理. 同时, 与对照CK相比, Cd胁迫下幼苗根冠比有不同程度的增加, 表明Cd对冬小麦幼苗地上部的抑制作用大于根部. 与幼苗根系相比, 茎叶的耐受指数更低, 说明茎叶对Cd胁迫的变化更敏感;且Cd25处理下根系和茎叶的耐受指数均低于Cd10处理, 表明幼苗根系和茎叶对高浓度Cd胁迫的变化更敏感.
|
|
表 1 外源壳聚糖对镉胁迫下冬小麦幼苗生物量和耐受指数的影响1) Table 1 Effect of exogenous chitosan on the biomass and tolerance index of winter wheat seedlings under Cd stress |
由表 1可知, 外源壳聚糖的添加可不同程度地促进Cd胁迫下冬小麦幼苗的生长. 当Cd10与壳聚糖同时添加时, 与单一的Cd10处理相比, 冬小麦幼苗的根长、茎叶长、根系干重和茎叶干重均增加, 但未达到显著水平, 且仍低于对照(CK). 当Cd25与壳聚糖同时添加时, 与单一的Cd25处理相比, 冬小麦幼苗的根长、茎叶长、根系干重和茎叶干重均增加, 且除茎叶干重未达到显著水平外, 其余均达到显著水平, 但仍低于对照(CK). 从耐受指数来看, Cd10 + CTS处理对根系和茎叶的抑制率(与CK相比)分别为41.33%和60.66%, 低于Cd10处理(49.84%和63.52%);Cd25 + CTS处理对根系和茎叶的抑制率分别为39.52%和59.89%, 低于Cd25处理(59.17%和68.22%). 以上结果表明外源壳聚糖可在一定程度上缓解Cd胁迫对小麦幼苗生长的抑制作用.
2.2 外源壳聚糖对镉胁迫下冬小麦幼苗叶片光合参数和叶绿素含量的影响如表 2所示, 与对照CK相比, Cd10和Cd25处理下幼苗叶片各光合参数指标均显著降低, 其中Cd10处理下冬小麦幼苗叶片Pn、Gs、Ci和Tr分别降低了36.46%、68.17%、15.93%和55.77%, Cd25处理下幼苗叶片Pn、Gs、Ci和Tr分别降低了44.83%、70.21%、41.99%和62.34%. 这些结果表明Cd胁迫下小麦幼苗叶片的光合作用受到一定程度的抑制, 且高浓度Cd胁迫对小麦幼苗叶片的光合抑制作用更明显.
|
|
表 2 外源壳聚糖对镉胁迫下冬小麦幼苗光合参数和叶绿素含量的影响 Table 2 Effect of exogenous chitosan on the photosynthetic parameters and chlorophyll content of winter wheat seedlings under Cd stress |
光合参数通常可用来反映光合作用的强弱[13]. 由表 2可知, 当Cd10与壳聚糖同时添加时, 与单一的Cd10处理相比, 冬小麦幼苗叶片的Pn、Gs、Ci和Tr分别提高26.03%、37.83%、14.81%和31.44%, 但仍低于对照CK, 且除了Ci达到显著水平外, 其余差异均不显著. 与单一的Cd25处理相比, Cd25+CTS处理下冬小麦幼苗叶片Pn、Gs、Ci和Tr分别提高了39.30%、39.55%、58.07%和42.59%, 但均低于对照CK. 该结果说明外源壳聚糖处理可提高Cd胁迫下冬小麦幼苗的光合作用, 可在一定程度上缓解Cd胁迫对冬小麦幼苗的光合抑制作用.
光合色素是在植物光合作用中参与吸收和传递光能的重要色素, 植物叶片光合色素含量高低是反映外界胁迫程度的重要指标之一[13]. 如表 2所示, 与对照CK相比, Cd10和Cd25处理下冬小麦幼苗叶片的叶绿素a、叶绿素b、类胡萝卜素及总叶绿素含量均显著降低(P < 0.05), 分别降低了44.14%、34.58%、52.16%、41.67%和50.85%、51.65%、53.17%、51.06%. 外源壳聚糖添加后, 幼苗叶片光合色素含量较单一Cd胁迫均显著提高(P < 0.05), 但仍显著低于对照CK. 其中Cd10+CTS处理冬小麦幼苗叶片叶绿素a、叶绿素b、类胡萝卜素及总叶绿素含量分别比Cd10单一胁迫下升高了53.01%、43.98%、59.06%和50.39%;Cd25+CTS处理较Cd25处理冬小麦幼苗叶片叶绿素a、叶绿素b、类胡萝卜素及总叶绿素含量分别升高了22.14%、77.02%、0.08%和36.15%. 以上结果表明, Cd胁迫对小麦幼苗叶片叶绿素的合成与积累具有抑制和破坏作用, 导致幼苗叶片叶绿素含量有不同程度地降低, 且Cd胁迫浓度越高, 其叶绿素含量的降低就越明显. 而外源壳聚糖能够通过提高小麦幼苗叶片光合色素含量的方式来缓解Cd胁迫对幼苗叶片的损伤, 进而缓解Cd胁迫对小麦幼苗的光合抑制作用.
2.3 外源壳聚糖对镉胁迫下冬小麦幼苗根系形态的影响根系具有吸收与运输水分、营养元素和矿质元素的能力[28]. 本研究中冬小麦幼苗根系形态指标结果如表 3所示. 从中可知, 与对照CK相比, Cd10处理下冬小麦幼苗根长、根表面积、根体积和总根尖数分别降低了69.55%、65.22%、60.24%和70.87%, 平均直径增大了13.96%, 差异显著;Cd25处理下冬小麦幼苗根长、根表面积、根体积和总根尖数分别降低了82.83%、78.15%、72.19%和87.12%, 平均直径增大了27.29%, 差异也显著. 以上结果表明Cd胁迫可抑制冬小麦幼苗根系的正常生长, 使得幼苗根长变短、侧根减少和根系变粗, 且Cd25处理对冬小麦幼苗根系生长的抑制作用强于Cd10处理.
|
|
表 3 外源壳聚糖对镉胁迫下冬小麦根系形态的影响 Table 3 Effect of exogenous chitosan on the root morphological indexes of winter wheat seedlings under Cd stress |
由表 3可知, Cd25+CTS处理下小麦幼苗根长、根表面积、根体积和总根尖数较Cd25处理分别提高了49.48%、45.67%、42.23%和35.19%, 但均未达到显著水平, 且仍低于对照CK. Cd10 + CTS处理下小麦幼苗根长、根表面积和根体积较Cd10处理分别提高了13.31%、31.14%和51.81%, 其中幼苗根体积达到显著水平, 且仍低于对照CK. 以上结果表明壳聚糖主要通过扩大根表面积和增加根体积来促进幼苗根系对水分和养分的吸收, 从而提高小麦幼苗抗Cd胁迫的能力.
2.4 外源壳聚糖对镉胁迫下冬小麦幼苗H2O2和MDA含量的影响为明确外源壳聚糖对Cd胁迫下小麦幼苗细胞氧化损伤的缓解作用, 测定了幼苗地上部和地下部的H2O2和MDA含量. 由图 1可知, 与对照CK相比, Cd10和Cd25单一胁迫处理下小麦幼苗根系和叶片中H2O2和MDA含量均显著提高(P < 0.05), 且增幅随着Cd胁迫浓度的增加而增大. Cd10处理的小麦幼苗根系中H2O2和MDA含量分别是对照CK处理的2.37倍和10.34倍, 叶片中H2O2和MDA含量分别是对照CK的4.48倍和2.26倍;但是与Cd10处理相比, Cd10 + CTS处理的小麦幼苗根系和叶片中H2O2和MDA含量均显著降低(P < 0.05), 其中根系H2O2和MDA含量分别降低了47.46%和48.12%, 叶片H2O2和MDA含量分别降低了45.81%和29.87%. 高浓度Cd胁迫(Cd25)下小麦幼苗根系中H2O2和MDA含量分别是对照CK处理的3.20倍和11.52倍, 叶片中H2O2和MDA含量分别是对照CK的7.79倍和3.25倍;但是与Cd25处理相比, Cd25 + CTS处理的小麦幼苗根系和叶片中H2O2和MDA含量均显著降低(P < 0.05), 根系中H2O2和MDA含量分别降低了38.21%和37.65%, 叶片中H2O2和MDA含量分别降低了55.73%和32.51%. 以上结果表明Cd胁迫可诱导小麦幼苗细胞氧化损伤, 且高浓度Cd胁迫导致的氧化损伤更明显;而壳聚糖有助于缓解Cd诱导产生的小麦幼苗细胞氧化损伤, 缓解Cd毒害.

|
不同小写字母表明5种处理之间差异显著(P < 0.05) 图 1 外源壳聚糖对Cd胁迫下小麦幼苗H2O2和MDA含量的影响 Fig. 1 Effect of exogenous chitosan on the contents of H2O2 and MDA of wheat seedlings under Cd stress |
图 2为壳聚糖对Cd胁迫下小麦幼苗根系和叶片中抗氧化酶(SOD、POD)活性的影响. 与对照CK相比, Cd10和Cd25单一胁迫处理下小麦幼苗叶片的SOD和POD均显著升高、幼苗根系中POD均显著降低(P < 0.05), 而Cd胁迫对幼苗根系中SOD无显著影响. Cd10处理的小麦幼苗叶片中SOD和POD分别较对照CK升高24.05%和50.23%, 根系中SOD和POD分别较对照CK降低4.10%和19.61%;与Cd10处理相比, Cd10+CTS处理的小麦幼苗根系和叶片中SOD和POD均无显著差异, 其中根系和叶片SOD分别升高2.78%和4.11%, 叶片POD降低1.46%, 而根系POD升高18.33%. Cd25处理的小麦幼苗叶片SOD和POD分别较对照CK升高21.08%和89.24%, 根系中POD较对照CK降低43.65%, 而根系SOD与对照CK差异不显著;与Cd25处理相比, Cd25 + CTS处理中除幼苗茎叶POD显著降低外, 幼苗根系和茎叶SOD及根系POD变化均不显著, 其中幼苗叶片SOD和POD分别降低2.24%和11.52%, 而幼苗根系SOD和POD分别升高5.61%和13.81%.

|
不同小写字母表明5种处理之间差异显著(P < 0.05) 图 2 外源壳聚糖对Cd胁迫下小麦幼苗抗氧化酶(SOD和POD)活性的影响 Fig. 2 Effect of exogenous chitosan on the antioxidant enzyme activities (SOD and POD) of wheat seedlings under Cd stress |
由表 4可知, 不同处理下小麦幼苗不同组织中Cd含量均表现为:地下部 > 地上部, 且Cd25处理下小麦幼苗地下部和地上部Cd含量均显著高于Cd10处理, 表明小麦幼苗对Cd的富集吸收会随着Cd胁迫浓度的增大而增强. 由表 4可知, Cd10和Cd25处理下小麦幼苗根系的Cd含量分别是茎叶的11.18倍和11.47倍, 而根系的Cd积累量分别是茎叶的4.72倍和4.52倍, 表明小麦幼苗吸收的Cd大部分积累到根系中. 当Cd10与壳聚糖同时添加时, 与单一的Cd10处理相比, 小麦幼苗根系和茎叶中Cd含量均显著降低(分别降低17.92%和13.22%), 而幼苗根系和茎叶中Cd积累量分别降低4.09%和6.71%, 转运系数降低了3.69%, 差异均不显著. Cd25+CTS处理下小麦幼苗根系和茎叶中Cd含量较单一的Cd25处理均显著降低(分别降低28.32%和21.63%), 而幼苗根系和茎叶中Cd积累量分别降低1.91%和5.37%, 转运系数降低2.76%, 差异均不显著. 以上结果表明外源壳聚糖可在一定程度上缓解小麦幼苗对Cd的吸收, 也能降低Cd从小麦根系向茎叶的转运.
|
|
表 4 外源壳聚糖对镉胁迫下小麦幼苗镉含量、积累量和转运系数的影响1) Table 4 Effect of exogenous chitosan on the Cd content, accumulation, and translocation factor of wheat seedlings under Cd stress |
2.7 影响冬小麦幼苗镉积累的主控因子分析
利用PLS模型的变量重要性投影值(VIP)分析方法, 定量分析了小麦幼苗各表型性状及生理生化指标对小麦幼苗地上部Cd积累量、地下部Cd积累量及总Cd积累量的影响力VIP值(图 3). VIP值越大, 表明该指标在预测因变量变化时的影响力越强, 紧密度越高. 鉴于此, 本研究幼苗地上部各表型性状及生理生化指标中, VIP值大于1.0的指标分别为:地上部Cd含量、茎叶干重、茎叶耐受系数、类胡萝卜素和地上部POD活性, 确定地上部Cd含量为地上部Cd积累量变化响应的最关键因子, 茎叶干重和茎叶耐受指数次之. 幼苗地下部各表型性状及生理生化指标中, VIP值大于1.0的指标分别为:总根尖数、总根长、根表面积、地下部Cd含量、根平均直径、根长、根体积和根冠比, 确定总根尖数为地下部Cd积累量变化响应的最关键因子, 总根长和根表面积次之. 幼苗各表型性状及生理生化指标中, VIP值大于1.0的指标分别为:根平均直径、总根尖数、总根长、地上部Cd含量、茎叶耐受系数、茎叶干重、根表面积、地下部Cd含量、根长、地上部SOD活性、类胡萝卜素和地上部POD活性, 表明根平均直径为幼苗总Cd积累量变化响应的最关键因子, 总根尖数和总根长次之.

|
a1, c2.茎叶长;a2, c4.茎叶干重;a3, c7.茎叶耐受指数;a4, c8.净光合速率;a5, c9.气孔导度;a6, c10.胞间CO2浓度;a7, c11.蒸腾速率;a8, c12.叶绿素a;a9, c13.叶绿素b;a10, c14.类胡萝卜素;a11, c15.叶绿素(a+b);a12, c21.地上部MDA含量;a13, c23.地上部H2O2含量;a14, c25.地上部SOD活性;a15, c27.地上部POD活性;a16, c29.地上部Cd含量;b1, c1.根长;b2, c3.根系干重;b3, c5.根冠比;b4, c6.根系耐受指数;b5, c16.总根长;b6, c17.根表面积;b7, c18.根体积;b8, c19.根平均直径;b9, c20.总根尖数;b10, c22.地下部MDA含量;b11, c24.地下部H2O2含量;b12, c26.地下部SOD活性;b13, c28.地下部POD活性;b14, c30.地下部Cd含量 图 3 基于PLS回归模型的小麦幼苗镉积累主控因子分析 Fig. 3 Identification of the main controlling factors of Cd accumulation in wheat seedlings based on a PLS regression model |
相关性分析结果表明(图 4), 小麦幼苗的根长、茎叶长、根系干重、茎叶干重、Pn、Gs、Tr、叶绿素a、类胡萝卜素、总叶绿素、根表面积、根体积和总根尖数与地上部及地下部Cd含量和Cd积累量呈极显著负相关(P < 0.01), 而根平均直径、幼苗的MDA和H2O2含量与地上部及地下部Cd含量和Cd积累量呈极显著正相关(P < 0.01).

|
1.地下部POD活性, 2.胞间CO2浓度, 3.叶绿素b, 4.类胡萝卜素, 5.总根长, 6.根表面积, 7.根长, 8.根体积, 9.总根尖数, 10.茎叶干重, 11.茎叶耐受指数, 12.根系干重, 13.根系耐受指数, 14.叶绿素a, 15.叶绿素(a+b), 16.气孔导度, 17.蒸腾速率, 18.茎叶长, 19.净光合速率, 20.地下部SOD活性, 21.地下部H2O2含量, 22.地上部MDA含量, 23.地上部H2O2含量, 24.地上部Cd积累量, 25.总Cd积累量, 26.地下部Cd积累量, 27.地上部POD活性, 28.地下部MDA含量, 29.地上部Cd含量, 30.地下部Cd含量, 31.根冠比, 32.根平均直径, 33.地上部SOD活性 图 4 Cd胁迫下外源壳聚糖诱导小麦幼苗各指标变化的相关性分析 Fig. 4 Correlation analysis of each index change of wheat seedlings induced by exogenous chitosan under Cd stress |
Cd胁迫会导致植株生长发育异常, 阻碍植株光合能力和叶绿素合成, 干扰其对水分和养分的吸收和运输, 破坏植株细胞内氧化还原稳态, 造成细胞氧化损伤, 扰乱正常的细胞结构和功能, 最终导致植株的生物量、产量和品质下降[29]. 本研究中, 10 µmol·L-1和25 µmol·L-1 Cd胁迫下小麦幼苗的株高、根长及根系和茎叶的干物质量均显著降低, 幼苗生长受到明显抑制, 且Cd25处理对冬小麦幼苗的抑制作用强于Cd10处理, 这与Liu等[7]和Zhou等[30]的研究结果一致;同时, Cd胁迫下幼苗根冠比有不同程度的增加, 这可能是由于小麦幼苗茎叶对Cd胁迫更敏感[13]. 相关性分析结果显示, 小麦幼苗的根长、茎叶长、根系干重、茎叶干重与地上部及地下部Cd含量和Cd积累量呈极显著负相关(P < 0.01), 这也证实了高浓度Cd胁迫会导致小麦Cd毒害, 抑制其正常生长. 有研究表明, 壳聚糖作为一种植物抗逆诱导剂, 可提高植物对重金属胁迫的抵抗能力, 缓解重金属对植物的损伤[31]. 曲丹阳等[14]研究发现, 外源施用适当浓度的壳聚糖能够显著促进Cd胁迫下玉米幼苗的生长, 缓解Cd胁迫对玉米幼苗的抑制作用. 本研究中, Cd10 + CTS处理及Cd25 + CTS处理小麦幼苗的根长、茎叶长、根系干重和茎叶干重及其耐受指数较对应的单一Cd胁迫处理均有不同程度的增加, 表明外源壳聚糖可在一定程度上缓解Cd胁迫对小麦幼苗生长的抑制作用, 这与王云等[4]的研究结果一致. 夏雪姣等[32]研究发现, Cd胁迫浓度达到40 mg·L-1时, 不同品种小麦根的生长均受到强烈抑制, 其中小麦根长下降69%~80%. 本研究中, Cd10及Cd25处理下小麦幼苗根长、根表面积、根体积和总根尖数较对照CK均显著降低, 而根平均直径显著增大, 表明10 µmol·L-1和25 µmol·L-1 Cd胁迫均显著抑制小麦幼苗根系的生长发育, 呈现根长变短、侧根减少、根系变粗的趋势. 这主要是由于小麦根系内蛋白质可螯合固定Cd2+, 且当Cd2+浓度达到某一阈值时会加速根系细胞木质化, 从而导致根系变短变粗[33~35]. Qin等[8]研究表明, Cd主要是通过抑制细根(< 1 mm)伸长和表面积增大来抑制植物的生长. 相关性分析结果显示, 幼苗地上部及地下部Cd含量和Cd积累量与根表面积、根体积、总根尖数呈极显著负相关(P < 0.01), 而与根平均直径呈极显著正相关(P < 0.01), 这也进一步证实高浓度Cd胁迫会导致小麦根系变短变粗且侧根减少. 而本研究中Cd10 + CTS及Cd25 + CTS处理小麦幼苗的根长、根表面积和根体积较对应的单一Cd胁迫处理均有不同程度的提高. 这与他人的研究结果一致, 如有研究表明适宜浓度的壳聚糖可促进玉米[13]和小麦[4]根系的生长.
有研究证实, Cd胁迫下植株光合色素含量呈现不同程度降低, 如Liu等[7]研究表明, 10 µmol·L-1 Cd胁迫下小麦幼苗叶绿素a、叶绿素b和总叶绿素含量分别较对照组下降19.77%、58.21%和26.82%;魏畅等[36]研究发现, 玉米叶片叶绿素a、叶绿素b、类胡萝卜素和总叶绿素含量均随着Cd胁迫浓度的增大而呈现降低趋势. 本研究中, Cd10和Cd25处理下冬小麦幼苗叶片的叶绿素a、叶绿素b、类胡萝卜素及总叶绿素含量均较对照CK显著降低(P < 0.05), 且高Cd胁迫降低得更明显. 这主要是由于:①Cd胁迫会抑制叶绿素生物合成相关酶活性, 导致叶绿素的生物合成受阻[37];②Cd2+会与叶绿素分子中的Mg2+竞争活性位点, 导致脱镁叶绿素含量增加[10, 38];③Cd胁迫造成叶绿体中ROS升高, 直接破坏相关光合色素的结构[39];④Cd2+可取代叶绿素或酶中的Ca2+等, 破坏其功能, 进而影响光合色素水平[36, 40]. 本试验中10 µmol·L-1和25 µmol·L-1 Cd胁迫下小麦幼苗的Pn、Gs、Ci和Tr均显著降低(表 2), 表明Cd胁迫显著抑制幼苗叶片的光合速率, 这与Liu等[7]和魏畅等[36]研究的结果相一致. 相关性分析结果显示, 幼苗地上部及地下部Cd含量和Cd积累量与幼苗叶片的Pn、Gs、Tr、叶绿素a、类胡萝卜素和总叶绿素均呈极显著负相关(P < 0.01). Gs反映植株气孔张开的程度[13];而气孔作为CO2进入植株和植株蒸腾失水的主要通道, 可持续感知外界环境信息[36]. 通常认为, Pn下降主要由气孔限制和非气孔限制导致:若Gs和Ci同时下降, 则Pn下降主要原因是气孔限制;若Ci保持不变或上升, 则是因为非气孔限制[13]. 因此, 气孔限制因素是本研究中Cd胁迫下Pn下降的主要诱因. 本研究中, 外源壳聚糖添加后, 幼苗叶片光合色素含量(叶绿素a、叶绿素b、类胡萝卜素及总叶绿素)较单一Cd胁迫组均显著提高(P < 0.05), 表明外源壳聚糖可缓解Cd胁迫对小麦幼苗叶绿素合成的抑制与破坏作用, 这与他人的研究结果相一致[13, 41]. 有研究表明, 壳聚糖可促进植株叶片对光能的吸收和利用, 提高叶绿体PSⅡ光电子传递速率和光化学效率[42~44].
3.2 外源壳聚糖增强冬小麦的抗氧化能力Cd胁迫可诱导植物体内产生并积累大量的活性氧自由基(reactive oxygen species, ROS), 从而导致细胞代谢紊乱, 破坏植物的正常生长和代谢[6, 14]. H2O2是Cd胁迫诱导氧化损伤的主要活性氧自由基, 其在植物细胞内大量产生会加剧膜脂过氧化, 对细胞产生毒害作用[14, 45]. MDA是细胞膜脂质过氧化的产物, 通常用其含量的高低来判断细胞膜过氧化水平和受损程度[28]. 本研究中, Cd10和Cd25胁迫下小麦幼苗根系和叶片中H2O2和MDA含量均显著提高(图 1, P < 0.05), 且增幅随Cd胁迫浓度的增加而增大, 说明小麦幼苗根系和叶片细胞膜均受到严重的氧化损伤, 且高Cd胁迫导致的毒害损伤更显著, 这与Amirjani[46]在小麦上的研究结果类似. 有研究发现蚕豆幼苗根部[47]和土豆块茎[48]中MDA也随着Cd胁迫显著升高. 本研究中外源壳聚糖添加后, 小麦幼苗根系和叶片中H2O2和MDA含量较单一Cd胁迫均显著降低(P < 0.05), 表明外源壳聚糖有助于缓解Cd胁迫对小麦幼苗细胞膜的损伤, 这与曲丹阳等[14]在玉米上的研究结果一致.
抗氧化酶系统是植物体内重要的ROS清除系统, SOD可歧化超氧阴离子生成H2O2和O2, POD能够将H2O2转变为H2O, 进而使活性氧降低, 减轻膜脂过氧化对细胞膜的损伤[13, 28]. 大量研究表明, Cd对不同植物的抗氧化酶活性影响不一致, 如Cd可提高水稻[49]中SOD和POD活性, 却抑制了油菜[50]中SOD的产生. 王振博等[6]研究表明, Cd胁迫下小麦幼苗根系和叶片中SOD和POD均显著升高, 且增幅随Cd胁迫浓度的增加而增大. 本研究中Cd10和Cd25胁迫下小麦幼苗叶片中SOD和POD均显著升高, 而根系中POD显著降低、SOD无显著变化. 这表明:一方面, 高浓度的Cd胁迫使根系内酶活性和代谢途径受抑制, 根系自身清除ROS的能力下降[10, 28];另一方面, 幼苗茎叶通过提高酶活性来减轻Cd胁迫的氧化损伤. 而外源壳聚糖添加后, 小麦幼苗根系和叶片中SOD和POD较单一Cd胁迫均有不同程度的提高, 表明外源壳聚糖可通过提高幼苗体内抗氧化酶活性来缓解Cd胁迫对小麦幼苗的氧化损伤, 这与曲丹阳等[13]在玉米上的研究结果一致.
3.3 外源壳聚糖抑制冬小麦对镉的吸收与积累植物对Cd的吸收和积累受诸多因素的影响, 如植物种类及品种[30, 51]、根际微生物[52]、镉浓度[53]、pH[54]和必需微量元素含量[7]等. 通常情况下, 植物对Cd的吸收和积累主要在根部, 且会随着Cd胁迫浓度的增大而增强[6, 55]. 本研究中, Cd10和Cd25处理下小麦幼苗根系的Cd含量分别是茎叶的11.18倍和11.47倍, 根系的Cd积累量分别是茎叶的4.72倍和4.52倍, 且Cd25处理下小麦幼苗根系和茎叶Cd含量和Cd积累量均不同程度地高于Cd10处理, 这与已有的研究结果相一致[6, 55]. PLS模型分析发现, 影响小麦幼苗地下部Cd积累量的关键主控因子是总根尖数, 影响幼苗总Cd积累量的关键因子是根平均直径. 根系作为植物吸收水分和养分的主要器官, 也最先感受和响应外界胁迫[28]. 相关性分析结果显示, 幼苗地上部及地下部Cd含量和Cd积累量与根表面积、根体积和总根尖数呈极显著负相关(P < 0.01), 而与根平均直径、MDA和H2O2含量呈极显著正相关(P < 0.01). 外源壳聚糖添加后, 小麦幼苗根系和茎叶中Cd含量较Cd单一胁迫组均显著降低, 幼苗Cd积累量和转运系数也降低(表 4), 表明外源壳聚糖可在一定程度上缓解小麦幼苗对Cd的吸收和转运, 这与王云等[4]报道的壳聚糖能缓解Cd对小麦幼苗的毒害作用、降低小麦幼苗对Cd吸收的结果相一致. 植株对Cd的低积累与一系列复杂的生理过程紧密相关, 如根部的高截留[56]、根部向茎叶和籽粒的低转运[57]及高效地阻塞节点上的Cd[58]等. 目前, 壳聚糖还被证实可有效降低小麦植株对钒的吸收和转运[59], 但壳聚糖是如何降低小麦对重金属尤其是Cd的吸收和转运的机制尚不清楚, 还有待于运用植物关键功能基因/蛋白等分子生物学手段进行深入研究.
4 结论(1)外源壳聚糖可通过提高小麦叶片的叶绿素含量和光合作用参数, 促进Cd胁迫下小麦幼苗的生长和生物量的增加, 缓解Cd胁迫对小麦幼苗生长的抑制.
(2)外源壳聚糖可通过调控小麦幼苗的抗氧化酶活性, 减少MDA的积累, 缓解Cd胁迫导致的氧化损伤;降低小麦幼苗对Cd的吸收和积累, 减轻Cd胁迫对小麦幼苗的毒害作用.
(3)PLS模型结果表明, 影响小麦幼苗地下部Cd积累量的关键主控因子为总根尖数, 影响幼苗总Cd积累量的关键因子为根平均直径.
| [1] |
张静静, 朱爽阁, 朱利楠, 等. 不同钝化剂对微碱性土壤镉、镍形态及小麦吸收的影响[J]. 环境科学, 2020, 41(1): 460-468. Zhang J J, Zhu S G, Zhu L N, et al. Effects of different amendments on fractions and uptake by winter wheat in slightly alkaline soil contaminated by cadmium and nickel[J]. Environmental Science, 2020, 41(1): 460-468. |
| [2] | Qu J H, Yuan Y H, Zhang X M, et al. Stabilization of lead and cadmium in soil by sulfur-iron functionalized biochar: performance, mechanisms and microbial community evolution[J]. Journal of Hazardous Materials, 2022, 425. DOI:10.1016/j.jhazmat.2021.127876 |
| [3] | Rai P K, Lee S S, Zhang M, et al. Heavy metals in food crops: health risks, fate, mechanisms, and management[J]. Environment International, 2019, 125: 365-385. DOI:10.1016/j.envint.2019.01.067 |
| [4] |
王云, 蔡汉, 陆任云, 等. 壳聚糖对镉胁迫条件下小麦生长及生理的影响[J]. 生态学杂志, 2007, 26(10): 1671-1673. Wang Y, Cai H, Lu R Y, et al. Effects of chitosan on Triticum aestivum growth and physiology under cadmium stress[J]. Chinese Journal of Ecology, 2007, 26(10): 1671-1673. |
| [5] |
姜瑛, 魏畅, 焦秋娟, 等. 外源硅对镉胁迫下玉米生理参数及根系构型分级的影响[J]. 草业学报, 2022, 31(9): 139-154. Jiang Y, Wei C, Jiao Q J, et al. Effects of exogenous silicon application on physiological parameters, root architecture and diameter distribution of maize under cadmium stress[J]. Acta Prataculturae Sinica, 2022, 31(9): 139-154. |
| [6] |
王振搏, 刘红恩, 许嘉阳, 等. 镉胁迫下施硒对冬小麦幼苗镉吸收、抗氧化能力及植株超微结构的影响[J]. 植物营养与肥料学报, 2023, 29(3): 530-543. Wang Z B, Liu H E, Xu J Y, et al. Effects of selenium application on cadmium uptake, antioxidant activity, and ultrastructure of winter wheat seedlings under cadmium stress[J]. Journal of Plant Nutrition and Fertilizers, 2023, 29(3): 530-543. |
| [7] | Liu H T, Jiao Q J, Fan L N, et al. Integrated physio-biochemical and transcriptomic analysis revealed mechanism underlying of Si-mediated alleviation to cadmium toxicity in wheat[J]. Journal of Hazardous Materials, 2023, 452. DOI:10.1016/j.jhazmat.2023.131366 |
| [8] | Qin S Y, Liu H E, Nie Z J, et al. AsA-GSH cycle and antioxidant enzymes play important roles in Cd tolerance of wheat[J]. Bulletin of Environmental Contamination and Toxicology, 2018, 101(5): 684-690. DOI:10.1007/s00128-018-2471-9 |
| [9] | Abbas T, Rizwan M, Ali S, et al. Effect of biochar on alleviation of cadmium toxicity in wheat (Triticum aestivum L.) grown on Cd-contaminated saline soil[J]. Environmental Science and Pollution Research, 2018, 25(26): 25668-25680. DOI:10.1007/s11356-017-8987-4 |
| [10] |
熊敏先, 吴迪, 许向宁, 等. 土壤重金属镉对高等植物的毒性效应研究进展[J]. 生态毒理学报, 2021, 16(6): 133-149. Xiong M X, Wu D, Xu X N, et al. Advances in toxic effects of soil heavy metal cadmium on higher plants[J]. Asian Journal of Ecotoxicology, 2021, 16(6): 133-149. |
| [11] |
孙爽, 杨晓光, 李克南, 等. 中国冬小麦需水量时空特征分析[J]. 农业工程学报, 2013, 29(15): 72-82. Sun S, Yang X G, Li K N, et al. Analysis of spatial and temporal characteristics of water requirement of winter wheat in China[J]. Transactions of the Chinese Society of Agricultural Engineering, 2013, 29(15): 72-82. DOI:10.3969/j.issn.1002-6819.2013.15.010 |
| [12] | Zhang L X, Gao C, Chen C, et al. Overexpression of rice OsHMA3 in wheat greatly decreases cadmium accumulation in wheat grains[J]. Environmental Science & Technology, 2020, 54(16): 10100-10108. |
| [13] |
曲丹阳, 张立国, 顾万荣, 等. 壳聚糖对镉胁迫下玉米幼苗根系生长及叶片光合的影响[J]. 生态学杂志, 2017, 36(5): 1300-1309. Qu D Y, Zhang L G, Gu W R, et al. Effects of chitosan on root growth and leaf photosynthesis of maize seedlings under cadmium stress[J]. Chinese Journal of Ecology, 2017, 36(5): 1300-1309. |
| [14] |
曲丹阳, 顾万荣, 李丽杰, 等. 壳聚糖对镉胁迫下玉米幼苗叶片AsA-GSH循环的调控效应[J]. 植物科学学报, 2018, 36(2): 291-299. Qu D Y, Gu W R, Li L J, et al. Regulation of chitosan on the ascorbate-glutathione cycle in Zea mays seedling leaves under cadmium stress[J]. Plant Science Journal, 2018, 36(2): 291-299. |
| [15] | Ahmed K B M, Khan M M A, Siddiqui H, et al. Chitosan and its oligosaccharides, a promising option for sustainable crop production-a review[J]. Carbohydrate Polymers, 2020, 227. DOI:10.1016/j.carbpol.2019.115331 |
| [16] | Ahmad B, Khan M M A, Jaleel H, et al. Exogenously sourced γ-irradiated chitosan-mediated regulation of growth, physiology, quality attributes, and yield in Mentha piperita L.[J]. Turkish Journal of Biology, 2017, 41(2): 388-401. |
| [17] | Rahman M, Mukta J A, Sabir A A, et al. Chitosan biopolymer promotes yield and stimulates accumulation of antioxidants in strawberry fruit[J]. PLoS One, 2018, 13(9). DOI:10.1371/journal.pone.0203769 |
| [18] |
王聪, 董永义, 贾俊英, 等. NaCl胁迫下外源壳聚糖对菜用大豆叶绿体抗氧化系统的影响[J]. 植物营养与肥料学报, 2016, 22(5): 1356-1365. Wang C, Dong Y Y, Jia J Y, et al. Effects of exogenous chitosan on antioxidant system in chloroplast of vegetable soybean under NaCl stress[J]. Journal of Plant Nutrition and Fertilizer, 2016, 22(5): 1356-1365. |
| [19] | 王学奎, 黄见良. 植物生理生化实验原理与技术[M]. (第三版). 北京: 高等教育出版社, 2015. |
| [20] | Giannopolitis C N, Ries S K. Superoxide dismutases: I. Occurrence in higher plants[J]. Plant Physiology, 1977, 59(2): 309-314. DOI:10.1104/pp.59.2.309 |
| [21] | Maehly A C, Chance B. The assay of catalases and peroxidases[J]. Methods of Biochemical Analysis, 1954, 1: 357-424. |
| [22] | 刘萍, 李明军. 植物生理学实验[M]. (第二版). 北京: 科学出版社, 2016. |
| [23] | Wu F B, Zhang G P, Dominy P. Four barley genotypes respond differently to cadmium: Lipid peroxidation and activities of antioxidant capacity[J]. Environmental and Experimental Botany, 2003, 50(1): 67-78. DOI:10.1016/S0098-8472(02)00113-2 |
| [24] | Kamruzzaman M, Elmasry G, Sun D W, et al. Prediction of some quality attributes of lamb meat using near-infrared hyperspectral imaging and multivariate analysis[J]. Analytica Chimica Acta, 2012, 714: 57-67. DOI:10.1016/j.aca.2011.11.037 |
| [25] |
李岚涛, 张铎, 盛开, 等. 施氮量对菊芋块茎产量、品质与植株生理特性的影响[J]. 植物营养与肥料学报, 2022, 28(3): 496-506. Li L T, Zhang D, Sheng K, et al. Effects of nitrogen application rates on tuber yield, quality and plant physiological activities of Helianthus tuberosus L.[J]. Journal of Plant Nutrition and Fertilizers, 2022, 28(3): 496-506. |
| [26] |
杨晓江, 李静, 金轲, 等. 干旱胁迫下丛枝菌根真菌对羊草功能性状的影响[J]. 中国草地学报, 2023, 45(5): 20-31. Yang X J, Li J, Jin K, et al. Effects of arbuscular mycorrhizal fungi on functional traits of Leymus chinensis under drought stress[J]. Chinese Journal of Grassland, 2023, 45(5): 20-31. |
| [27] |
李贺勤, 王维华, 江绪文. 抗坏血酸对铜胁迫下广藿香幼苗生长、铜积累和抗氧化酶活性的影响[J]. 环境化学, 2016, 35(7): 1431-1437. Li H Q, Wang W H, Jiang X W. Effects of exogenous ascorbic acid on seedling growth, copper accumulation and antioxidant enzyme activities of pogostemon cablin under copper stress[J]. Environmental Chemistry, 2016, 35(7): 1431-1437. |
| [28] |
王龙, 樊婕, 魏畅, 等. 外源抗坏血酸对铜胁迫菊苣幼苗生长的缓解效应[J]. 草业学报, 2021, 30(4): 150-159. Wang L, Fan J, Wei C, et al. Mitigative effect of exogenous ascorbic acid on the growth of copper-stressed chicory (Cichorium intybus) seedlings[J]. Acta Prataculturae Sinica, 2021, 30(4): 150-159. |
| [29] | Hu J H, Chen G L, Xu K, et al. Cadmium in cereal crops: uptake and transport mechanisms and minimizing strategies[J]. Journal of Agricultural and Food Chemistry, 2022, 70(20): 5961-5974. DOI:10.1021/acs.jafc.1c07896 |
| [30] | Zhou Z, Wei C, Liu H T, et al. Exogenous ascorbic acid application alleviates cadmium toxicity in seedlings of two wheat (Triticum aestivum L.) varieties by reducing cadmium uptake and enhancing antioxidative capacity[J]. Environmental Science and Pollution Research, 2022, 29(15): 21739-21750. DOI:10.1007/s11356-021-17371-z |
| [31] |
余劲聪, 侯昌萍, 何舒雅, 等. 壳聚糖修复重金属污染土壤的研究进展[J]. 土壤通报, 2017, 48(1): 250-256. Yu J C, Hou C P, He S Y, et al. Remediation of heavy metal contaminated soils by chitosan[J]. Chinese Journal of Soil Science, 2017, 48(1): 250-256. |
| [32] |
夏雪姣, 菅明阳, 韩玉翠, 等. 镉胁迫对小麦形态发育及生理代谢的影响[J]. 农业生物技术学报, 2018, 26(9): 1494-1503. Xia X J, Jian M Y, Han Y C, et al. Effects of cadmium stress on morphological development and physiological metabolism in wheat (Triticum aestivum)[J]. Journal of Agricultural Biotechnology, 2018, 26(9): 1494-1503. |
| [33] |
王树凤, 施翔, 孙海菁, 等. 镉胁迫下杞柳对金属元素的吸收及其根系形态构型特征[J]. 生态学报, 2013, 33(19): 6065-6073. Wang S F, Shi X, Sun H J, et al. Metal uptake and root morphological changes for two varieties of Salix integra under cadmium stress[J]. Acta Ecologica Sinica, 2013, 33(19): 6065-6073. |
| [34] | Jones D L, Darah P R, Kochian L V. Critical evaluation of organic acid mediated iron dissolution in the rhizosphere and its potential role in root iron uptake[J]. Plant and Soil, 1996, 180(1): 57-66. DOI:10.1007/BF00015411 |
| [35] |
陈涵, 庄玉婷, 冯嘉仪, 等. 铅镉复合污染对台湾相思生长和元素吸收的影响[J]. 西南林业大学学报, 2022, 42(1): 76-82. Chen H, Zhuang Y T, Feng J Y, et al. Effects of combined pollution of Pb and Cd on the growth and element uptake of Acacia confusa [J]. Journal of Southwest Forestry University, 2022, 42(1): 76-82. |
| [36] |
魏畅, 焦秋娟, 柳海涛, 等. 镉暴露条件下玉米生长及根系构型分级特征研究[J]. 草业学报, 2022, 31(3): 101-113. Wei C, Jiao Q J, Liu H T, et al. Physiological effects of different Cd concentrations on maize root architecture and classification[J]. Acta Prataculturae Sinica, 2022, 31(3): 101-113. |
| [37] | Jariyal A. Effect of cadmium stress on growth and development of Cicer arietinum (Fabaceae)[J]. Environment & We: An International Journal of Science & Technology, 2020, 15: 49-59. |
| [38] | Parmar P, Kumari N, Sharma V. Structural and functional alterations in photosynthetic apparatus of plants under cadmium stress[J]. Botanical Studies, 2013, 54(1). DOI:10.1186/1999-3110-54-45 |
| [39] | Janik E, Maksymiec W, Mazur R, et al. Structural and functional modifications of the major light-harvesting complex Ⅱ in cadmium- or copper-treated Secale cereale [J]. Plant and Cell Physiology, 2010, 51(8): 1330-1340. DOI:10.1093/pcp/pcq093 |
| [40] | Faller P, Kienzler K, Krieger-Liszkay A. Mechanism of Cd2+ toxicity: Cd2+ inhibits photoactivation of photosystemⅡ by competitive binding to the essential Ca2+ site[J]. Biochimica et Biophysica Acta (BBA)-Bioenergetics, 2005, 1706(1-2): 158-164. DOI:10.1016/j.bbabio.2004.10.005 |
| [41] | Mohamed S A, Ahmed H S, El-Baowab A A. Effect of chitosan, putrescine and irrigation levels on the drought tolerance of sour orange seedlings[J]. Egyptian Journal of Horticulture, 2018, 45(2): 257-273. DOI:10.21608/ejoh.2018.3063.1050 |
| [42] | Zong H Y, Liu S, Xing R E, et al. Protective effect of chitosan on photosynthesis and antioxidative defense system in edible rape (Brassica rapa L.) in the presence of cadmium[J]. Ecotoxicology and Environmental Safety, 2017, 138: 271-278. DOI:10.1016/j.ecoenv.2017.01.009 |
| [43] | Zhang X Q, Li K C, Liu S, et al. Size effects of chitooligomers on the growth and photosynthetic characteristics of wheat seedlings[J]. Carbohydrate Polymers, 2016, 138: 27-33. DOI:10.1016/j.carbpol.2015.11.050 |
| [44] | Zhang X Q, Li K C, Xing R E, et al. miRNA and mRNA expression profiles reveal insight into chitosan-mediated regulation of plant growth[J]. Journal of Agricultural and Food Chemistry, 2018, 66(15): 3810-3822. DOI:10.1021/acs.jafc.7b06081 |
| [45] | Wang D F, Liu Y G, Tan X F, et al. Effect of exogenous nitric oxide on antioxidative system and S-nitrosylation in leaves of Boehmeria nivea (L.) Gaud under cadmium stress[J]. Environmental Science and Pollution Research, 2015, 22(5): 3489-3497. DOI:10.1007/s11356-014-3581-5 |
| [46] | Amirjani M. Effects of cadmium on wheat growth and some physiological factors[J]. International Journal of Forest, Soil and Erosion, 2012, 2(1): 50-58. |
| [47] | Nouairi I, Jalali K, Essid S, et al. Alleviation of cadmium-induced genotoxicity and cytotoxicity by calcium chloride in faba bean (Vicia faba L. var. minor) roots[J]. Physiology and Molecular Biology of Plants, 2019, 25(4): 921-931. DOI:10.1007/s12298-019-00681-5 |
| [48] | Xu D Y, Chen Z F, Sun K, et al. Effect of cadmium on the physiological parameters and the subcellular cadmium localization in the potato (Solanum tuberosum L.)[J]. Ecotoxicology and Environmental Safety, 2013, 97: 147-153. DOI:10.1016/j.ecoenv.2013.07.021 |
| [49] | Lin L, Zhou W H, Dai H X, et al. Selenium reduces cadmium uptake and mitigates cadmium toxicity in rice[J]. Journal of Hazardous Materials, 2012, 235-236: 343-351. DOI:10.1016/j.jhazmat.2012.08.012 |
| [50] | Wang C, Zhang S H, Wang P F, et al. Excess Zn alters the nutrient uptake and induces the antioxidative responses in submerged plant Hydrilla verticillata (L. f.) royle[J]. Chemosphere, 2009, 76(7): 938-945. DOI:10.1016/j.chemosphere.2009.04.038 |
| [51] | Wang L, Zhang Q Y, Liao X Y, et al. Phytoexclusion of heavy metals using low heavy metal accumulating cultivars: a green technology[J]. Journal of Hazardous Materials, 2021, 413. DOI:10.1016/j.jhazmat.2021.125427 |
| [52] | Liu H T, Luo L, Jiang G Y, et al. Sulfur enhances cadmium bioaccumulation in Cichorium intybus by altering soil properties, heavy metal availability and microbial community in contaminated alkaline soil[J]. Science of the Total Environment, 2022, 837. DOI:10.1016/j.scitotenv.2022.155879 |
| [53] | Jung M C. Heavy metal concentrations in soils and factors affecting metal uptake by plants in the vicinity of a Korean Cu-W mine[J]. Sensors, 2008, 8(4): 2413-2423. DOI:10.3390/s8042413 |
| [54] | Zhao K L, Liu X M, Xu J M, et al. Heavy metal contaminations in a soil-rice system: identification of spatial dependence in relation to soil properties of paddy fields[J]. Journal of Hazardous Materials, 2010, 181(1-3): 778-787. DOI:10.1016/j.jhazmat.2010.05.081 |
| [55] | Sebastian A, Prasad M N V. Cadmium minimization in rice. A review[J]. Agronomy for Sustainable Development, 2014, 34(1): 155-173. DOI:10.1007/s13593-013-0152-y |
| [56] | Liu W T, Zhou Q X, Sun Y B, et al. Identification of Chinese cabbage genotypes with low cadmium accumulation for food safety[J]. Environmental Pollution, 2009, 157(6): 1961-1967. DOI:10.1016/j.envpol.2009.01.005 |
| [57] | Liu J G, Zhu Q S, Zhang Z J, et al. Variations in cadmium accumulation among rice cultivars and types and the selection of cultivars for reducing cadmium in the diet[J]. Journal of the Science of Food and Agriculture, 2005, 85(1): 147-153. DOI:10.1002/jsfa.1973 |
| [58] | Guo J Y, Li K, Zhang X Z, et al. Genetic properties of cadmium translocation from straw to brown rice in low-grain cadmium rice (Oryza sativa L.) line[J]. Ecotoxicology and Environmental Safety, 2019, 182. DOI:10.1016/j.ecoenv.2019.109422 |
| [59] | Tham L X, Nagasawa N, Matsuhashi S, et al. Effect of radiation-degraded chitosan on plants stressed with vanadium[J]. Radiation Physics and Chemistry, 2001, 61(2): 171-175. DOI:10.1016/S0969-806X(00)00388-1 |