 2024, Vol. 45
2024, Vol. 45



我国蔬菜种植面积约2.2×107 hm2, 占农作物总种植面积的13.0%[1]. 与农作物相比, 菜地过度施用氮肥更为常见[2], 主要露地蔬菜氮肥施用总量是推荐量的2.7倍[3]. 长期大量的水肥投入使得菜地土壤C/N快速下降, 强烈地影响着土壤反硝化过程和硝态氮的累积和淋溶[4, 5]. 反硝化作用是菜地土壤氮素流失的重要途径, 也是农业生态系统中氮素循环的关键环节, 决定了矿质氮的含量和形态[6~8]. 据估计, 我国菜地土壤反硝化年损失量(以N计)可达45.8 kg·hm-2[3]. 反硝化过程是由具有不同功能基因和代谢途径的一系列反硝化细菌参与完成[5], NO3-还原为NO2-是反硝化作用的第一步和限速步骤之一[9], 该过程由narG基因编码的硝酸盐还原酶(NaR)催化;NO2-还原为NO是主要限速步骤, 由nirK和nirS基因编码的两种亚硝酸盐还原酶(Cu-NiR和cd1-NiR)催化[10];编码氧化亚氮还原酶(NoS)的nosZ基因控制着反硝化的最后一步, 负责将N2O还原为N2, 其丰度是N2O转化的主要驱动因素[11]. 不同的肥料种类和环境因子(如含水量、pH、NO3--N和NH4+-N等)对反硝化菌的生态位选择起决定性作用[12, 13], 影响着反硝化菌群落组成和多样性, 进而影响硝酸盐还原和N2O转化过程. 因此, 深入了解反硝化菌的群落组成和多样性对于揭示土壤氮素转化机制和调控因素具有重要意义.
生物炭作为一种可以降低土壤氮损失和提高氮利用效率的土壤改良剂[14, 15], 在农业生产中受到了广泛关注. 已有大量研究表明, 生物炭能够直接或间接地调控反硝化细菌的群落组成和多样性. 一方面, 生物炭可以作为碳源和微生境, 来促进反硝化细菌的增殖和多样化, 从而提高土壤反硝化过程的效率和稳定性. 另一方面, 生物炭通过提高土壤pH、SOC、C/N、氮素有效性和阳离子交换量[16~18], 增加反硝化微生物群落的多样性, 调控反硝化微生物群落组成[19]. 此外, 生物炭与不同类型氮肥配施对反硝化功能基因的影响也不同. Li等[20]研究发现, 生物炭与硝态氮肥配施通过降低无机氮含量增加nosZ基因丰度, 从而降低N2O排放量. Zhao等[21]研究发现, 生物炭与尿素配施通过增加土壤中有效C和有效N的含量, 增加nirS、nirK和nosZ基因的丰度. Li等[22]研究发现, 生物炭与添加了二甲基吡唑磷酸盐的氮肥配施, 通过影响生态位分化, 降低了nirS型和nirK型反硝化菌基因的丰度, 并提高了nosZ型反硝化菌基因的丰度. 因此, 反硝化功能基因的类型和丰度对生物炭与不同类型氮肥配施的响应存在着很大的差异.
综上所述, 生物炭对土壤反硝化细菌群落组成和多样性的影响尚不明确, 主要影响因素也具有很大的不确定性, 生物炭与不同类型氮肥配施对菜地土壤反硝化菌群落组成的研究更为鲜见. 因此, 本文以北京顺义菜地土壤为研究对象, 设置不施肥(CK)、单施(NH4)2SO4(N1)、(NH4)2SO4+生物炭(N1B)、单施KNO3(N2)和KNO3 +生物炭(N2B)这5个处理, 明确生物炭与(NH4)2SO4和KNO3配施对土壤理化性质的影响, 分析生物炭与两种类型氮肥配施对土壤反硝化菌群落组成和多样性的影响, 分别探究影响narG型、nirK型、nirS型和nosZ型反硝化菌群落组成的主要环境因子, 以期为理解生物炭对农业生态系统中氮循环的影响提供新的视角和数据支持, 并为优化农业肥料管理措施提供科学依据.
1 材料与方法 1.1 研究区概况试验地点位于中国农业科学院农业环境与可持续发展研究所北京顺义试验基地(40°15' N, 116°55' E, 海拔30.0 m), 处于华北平原北部, 属于潮白河冲积扇下段. 气候属于典型的暖温带半湿润大陆性季风气候, 其特点是夏季高温多雨, 冬季寒冷干燥. 多年平均降水量623.5 mm, 降雨主要集中于夏季7~8月, 多年平均气温12.5℃, 年均日照2 684 h, 有效积温4 500℃, 无霜期195 d左右. 耕层土壤质地主要为潮褐土. 供试生物炭为玉米秸秆生物炭, 由辽宁金和福农业科技股份有限公司提供, 以玉米秸秆为原料, 在600℃和厌氧条件下热解制成. 土壤和生物炭的基本性质见表 1.
|
|
表 1 试验区土壤和生物炭的基本性质1) Table 1 Basic properties of soil and biochar in the experimental area |
1.2 试验设计
田间试验开始于2019年, 每年5~8月种植黄瓜, 8~11月种植白菜. 试验设置5个处理:①对照(CK);②(NH4)2SO4(N1);③(NH4)2SO4 +生物炭[30 t·(hm2·a)-1](N1B);④KNO3(N2);⑤KNO3 +生物炭[30 t·(hm2·a)-1](N2B). 田间各处理施用等量的氮、磷、钾肥, 氮肥为(NH4)2SO4和KNO3, N用量为300 kg·(hm2·a)-1;磷肥为过磷酸钙, P2O5用量为300 kg·(hm2·a)-1;钾肥为硫酸钾, K2O用量为1 050 kg·(hm2·a)-1. 生物炭和氮磷钾肥年用量平均分配给黄瓜季和白菜季, 每季肥料均作为基肥在播种前一次性施用. 其它管理措施与当地传统方法一致. 每个处理3次重复, 各处理随机区组排列, 小区面积为4 m×5 m = 20 m2.
1.3 土壤样品的采集与测定土壤样品分别在冬季(2021年11月)、春季(2022年3月)和夏季(2022年6月)采集, 采集时避开降雨与灌溉之后. 在每个小区内采用五点法采集土壤样品, 采样深度为100 cm, 分5个层次(0~20、20~40、40~60、60~80和80~100 cm). 各层次样品分别混匀后过2 mm筛, 分3份装入自封袋, 一份在室温条件下自然风干, 用于测定土壤pH, 一份保存在4℃冰箱中, 用于测定SWC、NO3--N、NH4+-N、微生物量碳(MBC)和微生物量氮(MBN), 另一份保存在-80℃冰箱中, 用于测定土壤微生物群落组成.
土壤pH的测定:调节土壤和水的比例至1∶2.5, 用pH计测定;土壤含水量的测定:采用105℃烘干法测定;土壤NO3--N和土壤NH4+-N的测定:用0.01 mol·L-1的CaCl2溶液浸提, 滤液采用流动注射自动分析仪(Braun and Lübbe, Germany)测定;MBC和MBN的测定:采用氯仿熏蒸-K2SO4提取法进行提取, 浸提液过滤后使用multi N/C 2100S总有机碳分析仪(Jena, Germany)测定. MBC(MBN)计算公式为:
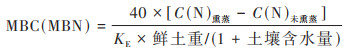
|
式中, C(N)熏蒸和C(N)未熏蒸分别表示氯仿熏蒸和未熏蒸土壤提取液中的全碳(全氮)含量(μg·g-1);KE表示转换系数, 取值为0.45. 计算结果为每单位干土所含微生物量碳(微生物量氮)的量(μg·g-1).
1.4 16S rRNA测序使用E.Z.N.A. Soil DNA Kit(Omega Bio-tek, Inc., USA)试剂盒提取土壤样本的基因组DNA, 在1.0%琼脂糖凝胶上进行电泳检测后用Nanodrop 2000分光光度计(ThermoFisher Scientific, Inc., USA)测定DNA质量和浓度. 利用通用引物338F(5′-ACTCCT ACGGGAGGCAGCAG-3′)和806R(5′-GGACTACHV GGGTWTCTAAT-3′)扩增细菌16S rRNA基因的V3-V4区. 总扩增体积为25 μL, 包括2 μL DNA模板、1 μL Forward Primer、1 μL Reverse Primer、3 μL BSA(2 ng·μL-1)、12.5 μL 2xTaq Plus Master Mix和5.5 μL ddH2O. 用Agencourt AMPure XP(Beckman Coulter, Inc., USA)核酸纯化试剂盒对PCR产物进行自动化纯化, 然后使用NEB Next Ultra Ⅱ DNA Library Prep Kit(New England Biolabs, Inc., USA)建库试剂盒进行文库构建. 高通量测序在北京奥维森基因科技有限公司的Illumina MiSeq(Illumina, Inc., USA)平台上进行.
1.5 数据分析所有数据均用3次重复的平均值和标准差表示. 用Microsoft Office Excel 2019软件进行数据整理, Origin 2023软件进行图表绘制. 通过IBM SPSS Statistics 26软件, 在0.05的显著性水平下, 进行单因素方差分析(ANOVA)和Turkey HSD事后检验, 来确定不同处理之间土壤理化性质的差异. 将OTU代表序列使用BLAST算法与Silva138数据库进行比对[23, 24], e-value阈值设置为1e-5, 得到每个OTU对应的物种分类信息. 使用QIIME(v1.8.0)软件计算Alpha多样性指数. 利用Canoco 5对微生物数据进行冗余分析(RDA), 确定影响土壤细菌群落结构(属水平)的主要因子, 使用R(4.2.2版)中“ggplot2”包绘制属水平物种组成堆积柱状图;利用galaxy线上平台进行LEfSe分析并绘图.
2 结果与分析 2.1 土壤理化性质不同处理土壤含水量如图 1(a)~1(c)所示. 冬季, 各处理土壤平均含水量变化范围在23.2%~26.3%之间. 在土壤垂直剖面上, 0~20 cm和20~40 cm土层不同处理SWC存在显著差异, 其余土层不同处理之间无显著差异. 在0~20 cm土层中, N2B处理SWC比CK和N2处理分别增加了39.3%和30.2%(P<0.05);在20~40 cm土层中, N1B处理比CK和N1处理分别增加了32.8%和20.7%(P<0.05)[图 1(a)]. 春季, 土壤平均含水量变化范围在22.8%~25.7%之间. 20~40 cm土层中不同处理SWC存在显著差异, N1B处理比CK和N1处理分别增加了54.5%和52.6%(P<0.05);其余土层中不同处理之间无显著差异[图 1(b)]. 夏季, 土壤平均含水量变化范围在16.8%~18.9%之间, 整个土壤剖面同一土层的不同处理间差异不显著[图 1(c)].

|
红色水平线表示同一处理所有土层含水量或pH的平均值, 不同大写字母表示不同处理间所有土层含水量或pH的平均值差异显著(P<0.05), 不同小写字母表示同一土层中不同处理间含水量或pH差异显著(P<0.05) 图 1 不同处理土壤含水量和土壤pH的变化 Fig. 1 Variations in soil water content and soil pH under different treatments |
图 1(d)~1(f)为不同处理土壤pH的变化. 冬季和夏季, 在土壤垂直剖面上, 0~20 cm和20~40 cm土壤中不同处理pH存在显著差异, 其余土层中不同处理之间无显著差异. 冬季, 与CK相比, N1、N1B、N2和N2B处理pH分别降低了0.26~0.27、0.17~0.19、0.31~0.36和0.27~0.31个单位(P<0.05), N1B处理比N1处理升高了0.05~0.08个单位, N2B处理比N2处理升高了0.04~0.10个单位[图 1(d)]. 夏季, 与CK相比, N1处理的pH减少了0.15~0.33个单位(P<0.05), N1B处理比N1处理增加了0.04~0.08个单位, N2B处理比N2处理增加了0.06~0.07个单位[图 1(f)]. 春季, 与CK处理相比, 各土层不同处理土壤pH均无显著差异[图 1(e)].
图 2为不同处理土壤NH4+-N和NO3--N含量的变化. 在整个土壤剖面中, 各处理的土壤NH4+-N和NO3--N含量均随着土层深度的增加而逐渐降低. 各土层N2和N2B处理的NH4+-N含量与CK相比无显著差异. 冬季, 各土层N1和N1B处理的土壤NH4+-N含量均显著高于CK处理, 分别增加了184.7%~287.9%和181.1%~245.8%(P<0.05), 在0~20 cm土层中, N1处理比N1B处理减少了17.7%(P<0.05), 在20~40 cm土层中, 两种处理无显著差异, 在40~100 cm各土层中, N1处理比N1B处理分别增加了37.2%、34.8%和34.7%(P<0.05). 春季, 与CK处理相比, 各土层N1和N1B处理的土壤NH4+-N含量分别增加了113.4%~151.7%和99.3%~195.5%(P<0.05). 在0~60 cm各土层中, N1B处理的土壤NH4+-N含量比N1处理分别增加了17.4%、14.1%和14.5%(P<0.05), 其它土层两种处理无显著差异. 夏季, 与CK处理相比, 各土层N1和N1B处理的土壤NH4+-N含量分别增加了338.2%~412.2%和422.9%~568.0%(P<0.05). 除80~100 cm土层外, 其余土层N1B处理的土壤NH4+-N含量均显著高于N1处理, 增幅在27.1%~33.6%之间.

|
红色水平线表示同一处理所有土层NH4+-N或NO3--N含量的平均值, 不同大写字母表示不同处理间所有土层NH4+-N或NO3--N含量的平均值差异显著(P<0.05), 不同小写字母表示同一土层中不同处理间NH4+-N或NO3--N含量差异显著(P<0.05) 图 2 不同处理土壤NH4+-N和NO3--N含量的变化 Fig. 2 Variations in soil NH4+-N and NO3--N content under different treatments |
本试验结果表明, 生物炭与氮肥配施显著影响了NO3--N含量及其空间分布[图 2(d)~2(f)]. 与CK处理相比, 各季节不同土层N2处理和N2B处理的土壤NO3--N含量均显著增加, 增幅在167.5%~785.4%和231.0%~896.7%. 冬季, 在整个土壤剖面中, 各土层N2B处理的土壤NO3--N含量均显著高于N2处理, 增幅在12.6%~52.5%之间. 除80~100 cm土层无显著差异外, 其余土层N1B处理的土壤NO3--N含量均显著高于N1处理, 增幅在12.2%~30.6%之间. 春季, 在整个土壤剖面中, 各土层N2B处理的土壤NO3--N含量均显著高于N2处理, 增幅在8.9%~74.7%之间. 在40~80 cm土层中, N1B处理的土壤NO3--N含量比N1处理分别减少了19.0%和22.2%(P<0.05), 其余土层N1B处理均显著高于N1处理, 增幅在21.9%~88.4%之间. 夏季, 在0~40 cm土层中, N2B处理的土壤NO3--N含量比N2处理分别增加了5.5%和20.5%(P<0.05), 在40~100 cm土层中, N2B处理比N2处理分别减少了20.9%、38.5%和43.5%(P<0.05). 在0~40 cm土层中, N1B处理比N1处理分别增加了17.9%和3.6%(P<0.05), 在40~80 cm土层中, N1B处理比N1处理分别减少了17.6%和23.3%(P<0.05), 80~100 cm土层两种处理无显著差异. 与N1处理相比, N1B处理土壤NH4+-N含量平均值增加了14.6%, NO3--N含量平均值增加了9.4%;与N2处理相比, N2B处理土壤NH4+-N含量平均值增加了16.0%, NO3--N含量平均值增加了15.9%.
2.2 土壤MBC和MBN含量如图 3所示, 土壤MBC和MBN含量随着土壤深度的增加而逐渐降低. 在整个土壤剖面中, 除80~100 cm土层外, 其它土层各处理MBC含量均表现为:N2B>N1B>N2>N1>CK. 与CK处理相比, 各季节不同土层N1B和N2B处理的土壤MBC含量均显著增加, 增幅在20.1%~51.5%和30.7%~94.2%. 冬季, 各土层N1B处理的土壤MBC含量均显著高于N1处理, 分别提高了13.9%~36.3%;各土层N2B处理均显著高于N2处理, 分别提高了14.7%~30.5%. 春季和夏季, 各土层N1B处理的土壤MBC含量均显著高于N1处理, 分别提高了14.3%~33.9%和15.8%~34.9%;除80~100 cm土层无显著差异外, 其余土层N2B处理均显著高于N2处理, 增幅在12.6%~27.7%和10.5%~24.9%.

|
红色水平线表示同一处理所有土层MBC或MBN含量的平均值, 不同大写字母表示不同处理间所有土层MBC或MBN含量的平均值差异显著(P<0.05), 不同小写字母表示同一土层中不同处理间MBC或MBN含量差异显著(P<0.05) 图 3 不同处理土壤MBC和MBN含量的变化 Fig. 3 Variations in soil MBC and MBN content under different treatments |
图 3(d)~3(f)为土壤MBN含量的变化. 在整个土壤剖面中, 各土层不同处理MBN含量均表现为:N2B>N1B>N2>N1>CK. 冬季和春季的0~60 cm土层中, 与CK处理相比, N1B处理的土壤MBN含量分别增加了58.7%~189.7%和73.0%~227.4%, N2B处理分别增加了123.8%~229.6%和161.4%~292.3%;N1B比N1分别增加了42.9%~171.3%和47.7%~181.3%;N2B比N2分别增加了27.6%~151.8%和54.3%~123.8%;其余土层处理之间无显著差异. 夏季, 各土层N1B处理的土壤MBN含量均显著高于CK和N1处理, 分别增加了64.4%~287.9%和48.5%~252.1%;各土层N2B处理均显著高于CK和N2处理, 分别增加了119.9%~401.4%和46.9%~97.0%.
2.3 土壤反硝化菌的Alpha多样性图 4为不同处理表层土壤反硝化菌的Alpha多样性指数, Chao1指数反映微生物群落的丰富度, Shannon指数反映微生物群落的多样性. 在narG型反硝化菌中, 与CK处理相比, N1处理的Chao1指数增加, 但并不显著;N1处理的Shannon指数显著高于CK和N1B处理(P<0.05). 在nirK型反硝化菌中, N1处理的Chao1指数显著低于CK处理(P<0.05), 不同处理间Shannon指数无显著差异. 在nirS型反硝化菌中, 与CK处理相比, N1处理的Chao1指数显著降低(P<0.05);N2处理的Shannon指数显著高于CK和N2B处理(P<0.05). 在nosZ型反硝化菌中, 不同处理间Chao1指数和Shannon指数均无显著差异.

|
不同小写字母表示不同处理间Alpha多样性指数差异显著(P<0.05) 图 4 不同处理下土壤narG型、nirK型、nirS型和nosZ型反硝化菌的Alpha多样性指数 Fig. 4 Alpha diversity index of soil denitrification bacteria of narG, nirK, nirS, and nosZ types under different treatments |
图 5为不同处理中相对丰度大于1%的反硝化菌属. 结果表明, 不同处理中4种反硝化菌所有优势属均属于变形菌门, 分布于α-、β-和γ-变形菌纲. narG型反硝化菌群落的优势菌属主要包括贪噬菌属(Variovorax)和氢噬胞菌属(Hydrogenophaga). 与CK处理相比, 其余处理Variovorax的相对丰度均增加;N1B(8.8%)和N2B处理(8.2%)的相对丰度小于N1(9.8%)和N2处理(10.9%). Hydrogenophaga相对丰度范围为8.1%~11.5%, 最大值和最小值分别出现在N2B和N1B处理中[图 5(a)]. nirK型反硝化菌群落的优势菌属主要包括根瘤菌属(Rhizobium)、中华根瘤菌属(Sinorhizobium)和慢生根瘤菌属(Bradyrhizobium). Rhizobium相对丰度范围为8.5%~25.5%, 最大值和最小值分别出现在N1和N2B处理中. N1B(14.4%)和N2B处理(5.7%)的Sinorhizobium相对丰度小于N1(17.2%)和N2处理(19.8%). Bradyrhizobium相对丰度范围为6.6%~16.3%, 最大值和最小值分别出现在N2B和N1处理中[图 5(b)]. nirS型反硝化菌群落的优势菌属主要包括Noviherbaspirillum和红长命菌属(Rubrivivax). 与CK处理相比, 其余处理Noviherbaspirillum的相对丰度均增加, 其中N1B(6.6%)和N2B处理(6.9%)的相对丰度小于N1(9.0%)和N2处理(7.5%). 与CK处理相比, 其余处理Rubrivivax的相对丰度均减少;N2B处理(6.1%)高于N2处理(5.2%), N1B处理(3.3%)低于N1处理(3.4%)[图 5(c)]. nosZ型反硝化菌群落的优势菌属主要包括沙壤土杆菌属(Ramlibacter)、微枝形杆菌属(Microvirga)和马赛菌属(Massilia). 与CK处理相比, 其余处理Ramlibacter的相对丰度均减少;N2B处理(21.9%)高于N2处理(9.3%), N1B处理(5.4%)低于N1处理(8.3%). Microvirga相对丰度范围为3.5%~6.6%, 最大值和最小值分别出现在N2B和N1B处理中. Massilia相对丰度范围为2.3%~10.4%, 最大值和最小值分别出现在N1和N2B处理中[图 5(d)].

|
(a)narG, (b)nirK, (c)nirS, (d)nosZ 图 5 不同处理下土壤narG型、nirK型、nirS型和nosZ型反硝化菌属水平的群落组成 Fig. 5 Community composition of soil denitrification bacteria of narG, nirK, nirS, and nosZ types at the genus level under different treatments |
图 6为不同处理中丰度有显著差异的菌属. 为了进一步研究narG型、nirS型、nirK型和nosZ型反硝化菌各属水平的相对丰度差异, 采用线性回归分析(LDA)来估算每个属丰度对差异效果影响的大小, 发现了各处理中的生物标记物. 在narG型反硝化菌中, Acidovorax(1.5%)和Pseudomonas(1.4%)分别是CK和N1处理的生物标记物;N1B处理中, Burkholderia(1.0%)、Klebsiella(2.4%)和Acidiphilium(5.2%)是生物标记物. 在nirK型反硝化菌中, Mesorhizobium(10.8%)、Bosea(8.7%)和Alcaligenes(1.8%)分别是CK、N1和N1B处理的生物标记物;N2B处理中, Chelatococcus(3.5%)、Methylobacterium(5.9%)和Rhodopseudomonas(17.2%)是生物标记物. nirS型反硝化菌的生物标记物包括CK处理中的Rubrivivax(10.5%)、N1处理中的Sulfuritalea(1.2%)、N1B处理中Sulfuricaulis(2.8%)和N2B处理中Microvirgula(3.8%). nosZ型反硝化菌的生物标记物包括CK处理中的Azoarcus(2.2%)、Hydrogenophaga(2.5%)和Ramlibacter(30.8%);N1处理中Burkholderia(1.0%)和Mesorhizobium(3.9%);N2B处理中Leptothrix(1.5%)、Oligotropha(2.4%)和Microvirga(6.6%).

|
图 6 不同处理下土壤narG型、nirK型、nirS型和nosZ型反硝化菌的属水平LEfSe分析 Fig. 6 Genus level LEfSe analysis of narG, nirK, nirS, and nosZ denitrification bacteria in soil under different treatments |
土壤反硝化细菌群落组成的主要影响因子如图 7所示. 土壤环境因子共能解释narG型反硝化菌群落组成变化的72.6%, 其中pH(19.7%)和NH4+-N(16.3%)对narG型反硝化菌群落组成变化有极显著影响(P<0.01), SWC(13.1%)对narG型反硝化菌群落组成变化有显著影响(P<0.05). 土壤环境因子共能解释nirK型反硝化菌群落组成变化的67.2%, 其中pH(23.5%)和MBN(15.1%)对nirK型反硝化菌群落组成变化有极显著影响(P<0.01). 土壤环境因子共能解释nirS型反硝化菌群落组成变化的64.8%, 其中pH(23.2%)和NO3--N(15.9%)对nirS型反硝化菌群落组成变化有显著影响(P<0.05). 土壤环境因子共能解释nosZ型反硝化菌群落组成变化的67.9%, 其中pH(26.2%)对nosZ型反硝化菌群落组成变化有极显著影响(P<0.01), MBC/MBN(11.3%)和NO3--N(9.4%)对nosZ型反硝化菌群落组成变化有显著影响(P<0.05).

|
红色箭头表示环境因子在排序轴上的投影, 蓝色箭头表示反硝化菌属在排序轴上的投影 图 7 土壤环境因子与土壤narG型、nirK型、nirS型和nosZ型反硝化菌在属水平的冗余分析 Fig. 7 Redundancy analysis of soil environmental factors and soil denitrification bacteria of narG, nirK, nirS, and nosZ at the genus level |
微生物是土壤生态系统的重要组成部分, 对土壤环境变化的响应十分敏感. 本研究发现, 不同处理间反硝化菌的群落组成存在显著差异, 与CK处理相比, N1和N1B处理均降低了nirK型和nirS型反硝化菌的Chao1指数[图 4(b)和4(c)], 但N1B处理的Chao1指数显著高于单施氮肥处理. 这说明施加(NH4)2SO4降低了nirK型和nirS型反硝化菌的Chao1指数, 添加生物炭提高了Chao1指数. 这是因为N1B处理的NO3--N含量比N1处理增加了9.4%[图 2(d)~2(f)], 添加(NH4)2SO4会导致土壤中的硫氧化细菌增加[25], 从而与反硝化菌竞争硝酸盐, 降低nirK型和nirS型反硝化菌的Chao1指数, 生物炭本身含有可溶性有机碳[26], 土壤有机碳是土壤反硝化过程中非常重要的电子供体[27], 因此添加生物炭有助于nirK型和nirS型反硝化菌群落丰富度的增加. 在本研究中, 单施KNO3显著提升了nirS型反硝化菌的Shannon指数, 但各处理nirK型反硝化菌的Shannon指数并无显著差异[图 4(b)和4(c)], 这说明nirS型反硝化菌群落的多样性比nirK型反硝化菌对KNO3更敏感[28, 29]. 此外, nirS型反硝化菌的物种丰富度(Chao1指数)高于nirK型反硝化菌, 这与大多数研究结果一致[10, 30, 31].
土壤pH值和MBN是影响nirK型反硝化菌群落组成变化的主要环境因子. pH值的变化会导致nirK型反硝化菌群落组成和优势菌的变化[29]. 本研究发现, 单施氮肥会导致土壤pH降低[图 1(d)~1(f)], 从而抑制nirK型反硝化菌的生长和活性[13], 同时也会影响土壤中氮素形态和可利用性[32]. 与单施氮肥相比生物炭配施氮肥能有效提高土壤pH[图 1(d)~1(f)], 能促进反硝化菌的生长, 提高反硝化速率[12, 33]. 同时, 生物炭配施氮肥显著增加MBN含量[图 3(d)~3(f)], 较高的MBN意味着较高的氮源供应和反硝化潜力, 也会促进nirK型反硝化菌的丰度和多样性[34]. 各处理中占优势的nirK型反硝化菌均属于变形菌门, 除属于γ-变形菌纲的Ensifer外, 其它菌属均属于α-变形菌纲. 这与Hou等[35]研究结果一致, 大多数nirK型反硝化菌属于α-变形菌纲, 超过50%的nirK型反硝化菌属于根瘤菌目. 本研究结果发现, 与nirK型反硝化菌相比, nirS型反硝化菌受土壤pH影响较小, 这可能与nirK基因的分子组成和编码的亚硝酸盐还原酶的特性有关. 相关研究也发现, nirK基因的丰度和多样性比nirS基因对pH反应更为敏感[12]. 除土壤pH值外, 土壤NO3--N含量也是影响nirS型反硝化菌群落组成的重要因素之一. NO3--N作为反硝化过程的电子受体和反应底物, nirS型反硝化菌群落受NO3--N浓度的影响更为明显. Zhang等[36]研究表明, 中等水平(10~20 mg·L-1)和吸附态的NO3--N对其活性和丰度更有利. 此外, 配施生物炭还可以通过改善土壤理化性质(如:土壤组成、土壤持水力和土壤pH等)来抑制硝化作用, 促进反硝化作用[37], 从而影响土壤氮素供应水平.
3.2 生物炭和不同类型氮肥配施对narG型和nosZ型反硝化菌的影响narG型和nosZ型反硝化菌是参与土壤反硝化过程的关键微生物. 本研究表明, 与CK和N1B处理相比, N1处理增加了narG型反硝化菌的Chao1指数和Shannon指数(图 4), 这是因为N1处理的ω(NH4+-N)为27.8 mg·kg-1, 相比CK处理(7.95 mg·kg-1)显著增加, 相比N1B处理(31.8 mg·kg-1)显著减少[图 2(a)~2(c)], narG型反硝化菌在高氮处理下显著减少[38]. pH、NH4+-N和SWC是影响narG型反硝化菌群落组成变化的主要环境因子. NH4+-N可为微生物提供氮源和能源, 增加反硝化菌的基因丰度[39]. 本研究中生物炭与氮肥配施比单施氮肥土壤NH4+-N含量增加了14.8%[图 2(a)~2(c)], 其含量可以影响反硝化菌的生长和竞争力, 导致群落组成的变化[40], 适量的NH4+-N含量可以增加反硝化菌的多样性和丰度. 此外生物炭与氮肥配施还可以提高土壤有机质、全氮和碱解氮的含量, 促进土壤微生物的活性和数量[41]. 本研究发现, 与单施氮肥相比, 生物炭与氮肥配施土壤保水能力更强[图 1(a)~1(c)], 这与已有研究的结果一致[42~44]. 土壤水分会影响土壤中的氧含量和有机质的可利用性[34], SWC增加可以促进土壤厌氧环境的形成, 促进narG型反硝化菌将硝酸盐还原为亚硝酸盐的过程[45]. 不同类型的反硝化细菌对SWC的适应性可能有所不同[46]. 本研究中的narG型反硝化菌的优势属分布于α-变形菌纲、β-变形菌纲和γ-变形菌纲. N1处理中, Pseudomonas的数量高于其它组且相对丰度大于1%, 说明在本研究中NH4+-N的施加主要影响Pseudomonas. 本研究发现, pH、MBC / MBN和NO3--N是影响nosZ型反硝化菌群落组成变化的主要环境因子. 与MBN相同, MBC / MBN也是反映土壤微生物活性的重要指标, 影响着氮素利用效率(NUE)和碳利用效率(CUE)[47], 间接作用于nosZ型反硝化菌的群落变化. 本研究中的nosZ型反硝化菌的优势属主要分布于变形菌门, 这与已有研究的结果一致[48, 49]. N2B处理中Leptothrix、Oligotropha和Microvirga的丰度高于其它处理, 这说明NO3--N和生物炭的添加主要影响了上述3种菌属.
4 结论(1)生物炭与两种类型氮肥配施均能够增加土壤矿质氮含量, 与单施(NH4)2SO4相比, 生物炭和(NH4)2SO4配施的NH4+-N含量增加了14.6%, NO3--N含量增加了9.4%;与单施KNO3相比, 生物炭与KNO3配施的NH4+-N含量增加了16.0%, NO3--N含量增加了15.9%.
(2)反硝化菌群落的优势门类在不同处理中均为变形菌门, 但其组成和优势菌会受到生物炭与两种类型氮肥配施的影响. 生物炭与(NH4)2SO4配施主要增加了α-、β-和γ-变形菌纲的相对丰度. 在该处理下, 反硝化菌群落中3个变形菌纲分别占比5.2%~11.8%、8.1%~16.9%和0%~2.4%, 占比前3的属分别为Acidiphilium、Sulfuricaulis和Klebsiella. 生物炭与KNO3配施主要增加了α-和β-变形菌纲的相对丰度. 在该处理下, 反硝化菌群落中α-和β-变形菌纲分别占比1.5%~61.7%和14.4%~27.2%, 占比前3的属分别为Rhodopseudomonas、Microvirga和Methylobacterium.
(3)土壤pH、NH4+-N和SWC是影响narG型反硝化菌群落组成变化的主要环境因子, pH、MBC/MBN和NO3--N是影响nosZ型反硝化菌群落组成变化的主要环境因子, pH值和MBN是影响nirK型反硝化菌群落组成变化的主要环境因子, pH值和NO3--N是影响nirS型反硝化菌群落组成变化的主要环境因子. nirS型反硝化菌的物种丰富度高于nirK型反硝化菌;nirK基因的丰度和多样性比nirS基因对pH更敏感, 与nirK型反硝化菌群落相比, nirS型反硝化菌群落对NO3--N更敏感.
| [1] | 国家统计局. 国家数据[EB/OL]. https://data.stats.gov.cn/easyquery.htm?cn=C01, 2023-04-07. |
| [2] | Zhu X Q, Zhou P, Miao P, et al. Nitrogen use and management in orchards and vegetable fields in China: challenges and solutions[J]. Frontiers of Agricultural Science and Engineering, 2022, 9(3): 386-395. |
| [3] |
黄绍文, 唐继伟, 李春花, 等. 我国蔬菜化肥减施潜力与科学施用对策[J]. 植物营养与肥料学报, 2017, 23(6): 1480-1493. Huang S W, Tang J W, Li C H, et al. Reducing potential of chemical fertilizers and scientific fertilization countermeasure in vegetable production in China[J]. Journal of Plant Nutrition and Fertilizer, 2017, 23(6): 1480-1493. |
| [4] | Abdalla M, Hastings A, Cheng K, et al. A critical review of the impacts of cover crops on nitrogen leaching, net greenhouse gas balance and crop productivity[J]. Global Change Biology, 2019, 25(8): 2530-2543. DOI:10.1111/gcb.14644 |
| [5] | Zhang L Y, Jing Y M, Chen C R, et al. Effects of biochar application on soil nitrogen transformation, microbial functional genes, enzyme activity, and plant nitrogen uptake: a meta-analysis of field studies[J]. GCB Bioenergy, 2021, 13(12): 1859-1873. DOI:10.1111/gcbb.12898 |
| [6] |
蔡玉佳, 沈菊培, 张成军, 等. 不同土地利用方式土壤氨氧化微生物和反硝化微生物时空分布特征[J]. 生态学报, 2022, 42(14): 5847-5858. Cai Y J, Shen J P, Zhang C J, et al. Spatial-temporal patterns of soil ammonia oxidizer and denitrifier bacteria under different land-use types[J]. Acta Ecologica Sinica, 2022, 42(14): 5847-5858. |
| [7] | Sun X, Han X G, Ping F, et al. Effect of rice-straw biochar on nitrous oxide emissions from paddy soils under elevated CO2 and temperature[J]. Science of the Total Environment, 2018, 628⁃629: 1009-1016. |
| [8] | Dannenmann M, Butterbach-Bahl K, Gasche R, et al. Dinitrogen emissions and the N2: N2O emission ratio of a Rendzic Leptosol as influenced by pH and forest thinning[J]. Soil Biology and Biochemistry, 2008, 40(9): 2317-2323. DOI:10.1016/j.soilbio.2008.05.009 |
| [9] | Yang J W, Su W H, Yu Q L, et al. The long-term decomposition of wild animal corpses leads to carbon and phosphorus accumulation and disturbs the ecological succession of the denitrification community encoded by narG[J]. Applied Soil Ecology, 2022, 175. DOI:10.1016/j.apsoil.2022.104455 |
| [10] | Sun H S, Jiang S X. A review on nirS-type and nirK-type denitrifiers via a scientometric approach coupled with case studies[J]. Environmental Science: Processes & Impacts, 2022, 24(2): 221-232. |
| [11] | Liu H S, Zheng X Z, Li Y F, et al. Soil moisture determines nitrous oxide emission and uptake[J]. Science of the Total Environment, 2022, 822. DOI:10.1016/j.scitotenv.2022.153566 |
| [12] | Liang Y Q, Wu C F, Wei X M, et al. Characterization of nirS- and nirK-containing communities and potential denitrification activity in paddy soil from eastern China[J]. Agriculture, Ecosystems & Environment, 2021, 319. DOI:10.1016/j.agee.2021.107561 |
| [13] |
谢军, 王子芳, 王蓥燕, 等. 化肥和有机肥配施生物炭对根际土壤反硝化势和反硝化细菌群落的影响[J]. 环境科学, 2023, 44(8): 4565-4574. Xie J, Wang Z F, Wang Y Y, et al. Effect of chemical fertilizer and manure combined with biochar on denitrification potential and denitrifying bacterial community in rhizosphere soil[J]. Environmental Science, 2023, 44(8): 4565-4574. |
| [14] | Pan Y C, She D L, Shi Z Q, et al. Do biochar and polyacrylamide have synergistic effect on net denitrification and ammonia volatilization in saline soils?[J]. Environmental Science and Pollution Research, 2021, 28(42): 59974-59987. DOI:10.1007/s11356-021-14886-3 |
| [15] | Su X X, Wang Y Y, He Q, et al. Biochar remediates denitrification process and N2O emission in pesticide chlorothalonil-polluted soil: Role of electron transport chain[J]. Chemical Engineering Journal, 2019, 370: 587-594. DOI:10.1016/j.cej.2019.03.195 |
| [16] | Hailegnaw N S, Mercl F, Pračke K, et al. Mutual relationships of biochar and soil pH, CEC, and exchangeable base cations in a model laboratory experiment[J]. Journal of Soils and Sediments, 2019, 19(5): 2405-2416. DOI:10.1007/s11368-019-02264-z |
| [17] | Zhang Y, Zhao C C, Chen G L, et al. Response of soil microbial communities to additions of straw biochar, iron oxide, and iron oxide-modified straw biochar in an arsenic-contaminated soil[J]. Environmental Science and Pollution Research, 2020, 27(19): 23761-23768. DOI:10.1007/s11356-020-08829-7 |
| [18] | Qin J H, Niu A Y, Li Q W, et al. Effect of soluble calcium on enhancing nitrate retention by biochar[J]. Journal of Environmental Management, 2020, 274. DOI:10.1016/j.jenvman.2020.111133 |
| [19] | Liao J Y, Hu A, Zhao Z W, et al. Biochar with large specific surface area recruits N2O-reducing microbes and mitigate N2O emission[J]. Soil Biology and Biochemistry, 2021, 156. DOI:10.1016/j.soilbio.2021.108212 |
| [20] | Li W, Xie H X, Ren Z H, et al. Response of N2O emissions to N fertilizer reduction combined with biochar application in a rain-fed winter wheat ecosystem[J]. Agriculture, Ecosystem and Environment, 2022, 333. DOI:10.1016/j.agee.2022.107968 |
| [21] | Zhao L, Li Q H, Qian X J, et al. Effects of the combined application of biochar-based fertilizer and urea on N2O emissions, nitrifier, and denitrifier communities in the acidic soil of pomelo orchards[J]. Journal of Soils and Sediments, 2022, 22(12): 3119-3136. DOI:10.1007/s11368-022-03273-1 |
| [22] | Li J, Wang S, Luo J F, et al. Effects of biochar and 3, 4-dimethylpyrazole phosphate (DMPP) on soil ammonia-oxidizing bacteria and nosZ-N2O reducers in the mitigation of N2O emissions from paddy soils[J]. Journal of Soils and Sediments, 2021, 21(2): 1089-1098. DOI:10.1007/s11368-020-02811-z |
| [23] | Ye J, McGinnis S, Madden T L. BLAST: improvements for better sequence analysis[J]. Nucleic Acids Research, 2006, 34(S2): W6-W9. |
| [24] | Pruesse E, Quast C, Knittel K, et al. SILVA: a comprehensive online resource for quality checked and aligned ribosomal RNA sequence data compatible with ARB[J]. Nucleic Acids Research, 2007, 35(21): 7188-7196. DOI:10.1093/nar/gkm864 |
| [25] | Chen X, Wei H, Zhang J E. Nitrogen and sulfur additions improved the diversity of nirK- and nirS-type denitrifying bacterial communities of farmland soil[J]. Biology, 2021, 10(11). DOI:10.3390/biology10111191 |
| [26] |
刘杰云, 邱虎森, 王聪, 等. 生物质炭对双季稻田土壤反硝化功能微生物的影响[J]. 环境科学, 2019, 40(5): 2394-2403. Liu J Y, Qiu H S, Wang C, et al. Influence of biochar amendment on soil denitrifying microorganisms in double rice cropping system[J]. Environmental Science, 2019, 40(5): 2394-2403. |
| [27] | Zhang Y S, Xu B K, Han J P, et al. Effects of drying-rewetting cycles on ferrous iron-involved denitrification in paddy soils[J]. Water, 2021, 13(22). DOI:10.3390/w13223212 |
| [28] | Wei W, Isobe K, Nishizawa T, et al. Higher diversity and abundance of denitrifying microorganisms in environments than considered previously[J]. The ISME Journal, 2015, 9(9): 1954-1965. DOI:10.1038/ismej.2015.9 |
| [29] | Xiao X, Xie G X, Yang Z H, et al. Variation in abundance, diversity, and composition of nirK and nirS containing denitrifying bacterial communities in a red paddy soil as affected by combined organic-chemical fertilization[J]. Applied Soil Ecology, 2021, 166. DOI:10.1016/j.apsoil.2021.104001 |
| [30] | Shi Y L, Zhang Q W, Liu X R, et al. Organic manure input and straw cover improved the community structure of nitrogen cycle function microorganism driven by water erosion[J]. International Soil and Water Conservation Research, 2022, 10(1): 129-142. DOI:10.1016/j.iswcr.2021.03.005 |
| [31] | Li P P, Zhang S Q, Li F, et al. Long term combined fertilization and soil aggregate size on the denitrification and community of denitrifiers[J]. Applied Soil Ecology, 2020, 156. DOI:10.1016/j.apsoil.2020.103718 |
| [32] | Azziz G, Monza J, Etchebehere C, et al. nirS- and nirK-type denitrifier communities are differentially affected by soil type, rice cultivar and water management[J]. European Journal of Soil Biology, 2017, 78: 20-28. DOI:10.1016/j.ejsobi.2016.11.003 |
| [33] | Olaya-Abril A, Hidalgo-Carrillo J, Luque-Almagro V M, et al. Effect of pH on the denitrification proteome of the soil bacterium Paracoccus denitrificans PD1222[J]. Scientific Reports, 2021, 11(1). DOI:10.1038/s41598-021-96559-2 |
| [34] | Yang Y J, Liu H X, Lv J L. Response of N2O emission and denitrification genes to different inorganic and organic amendments[J]. Sci Rep, 2022, 12(1). DOI:10.1038/s41598-022-07753-9 |
| [35] | Hou S P, Ai C, Zhou W, et al. Structure and assembly cues for rhizospheric nirK- and nirS-type denitrifier communities in long-term fertilized soils[J]. Soil Biology and Biochemistry, 2018, 119: 32-40. DOI:10.1016/j.soilbio.2018.01.007 |
| [36] | Zhang R J, Xu X M, Jia D T, et al. Sediments alleviate the inhibition effects of antibiotics on denitrification: functional gene, microbial community, and antibiotic resistance gene analysis[J]. Science of the Total Environment, 2022, 804. DOI:10.1016/j.scitotenv.2021.150092 |
| [37] |
屈田华, 李永夫, 张少博, 等. 生物质炭输入影响土壤氮素转化与氧化亚氮排放的研究进展[J]. 浙江农林大学学报, 2021, 38(5): 926-936. Qu T H, Li Y F, Zhang S B, et al. Effects of biochar application on soil nitrogen transformation and N2O emissions: a review[J]. Journal of Zhejiang A&F University, 2021, 38(5): 926-936. |
| [38] | Wang J Q, Shi X Z, Zheng C Y, et al. Inhibitory effect of nitrogen deposition on soil denitrifying activity in a subtropical forest[J]. Plant and Soil, 2023, 485(1). DOI:10.1007/s11104-022-05850-2 |
| [39] |
张培玉, 曲洋, 于德爽, 等. 菌株qy37的异养硝化/好氧反硝化机制比较及氨氮加速降解特性研究[J]. 环境科学, 2010, 31(8): 1819-1826. Zhang P Y, Qu Y, Yu D S, et al. Comparison of heterotrophic nitrification and aerobic denitrification system by strain qy37 and its accelerating removal characteristic of NH4+-N[J]. Environmental Science, 2010, 31(8): 1819-1826. |
| [40] | Cao Q, Li X Z, Xie Z J, et al. Compartmentation of microbial communities in structure and function for methane oxidation coupled to nitrification-denitrification[J]. Bioresource Technology, 2021, 341. DOI:10.1016/j.biortech.2021.125761 |
| [41] |
袁晶晶, 同延安, 卢绍辉, 等. 生物炭与氮肥配施对土壤肥力及红枣产量、品质的影响[J]. 植物营养与肥料学报, 2017, 23(2): 468-475. Yuan J J, Tong Y A, Lu S H, et al. Effects of biochar and nitrogen fertilizer application on soil fertility and jujube yield and quality[J]. Journal of Plant Nutrition and Fertilizer, 2017, 23(2): 468-475. |
| [42] | Fischer B M C, Manzoni S, Morillas L, et al. Improving agricultural water use efficiency with biochar-A synthesis of biochar effects on water storage and fluxes across scales[J]. Science of the Total Environment, 2019, 657: 853-862. |
| [43] | Fischer B M C, Morillas L, Conejo J R, et al. Investigating the impacts of biochar on water fluxes in a rice experiment in the dry corridor of Central America, Costa Rica[J]. Hydrological Processes, 2022, 36(12). DOI:10.1002/hyp.14765 |
| [44] | Aller D, Rathke S, Laird D, et al. Impacts of fresh and aged biochars on plant available water and water use efficiency[J]. Geoderma, 2017, 307: 114-121. |
| [45] | Chon K, Chang J S, Lee E, et al. Abundance of denitrifying genes coding for nitrate (narG), nitrite (nirS), and nitrous oxide (nosZ) reductases in estuarine versus wastewater effluent-fed constructed wetlands[J]. Ecological Engineering, 2011, 37(1): 64-69. |
| [46] | Chen Q R, Fan J F, Ming H X, et al. Effects of environmental factors on denitrifying bacteria and functional genes in sediments of Bohai Sea, China[J]. Marine Pollution Bulletin, 2020, 160. DOI:10.1016/j.marpolbul.2020.111621 |
| [47] | Mooshammer M, Wanek W, Hämmerle I, et al. Adjustment of microbial nitrogen use efficiency to carbon: nitrogen imbalances regulates soil nitrogen cycling[J]. Nature Communications, 2014, 5(1). DOI:10.1038/ncomms4694 |
| [48] |
李冀, 朱莹, 张晓君. 非典型氧化亚氮还原酶基因nosZ Ⅱ研究进展[J]. 微生物学通报, 2017, 44(7): 1714-1719. Li J, Zhu Y, Zhang X J. Researches on the atypical nitrous oxide reductase[J]. Microbiology China, 2017, 44(7): 1714-1719. |
| [49] |
邓文芳, 吴佳鹏, 洪义国, 等. 真菌反硝化过程及其驱动的N2O产生机制研究进展[J]. 微生物学报, 2021, 61(6): 1551-1566. Deng W F, Wu J P, Hong Y G, et al. The diversity distribution and N2O production driven by fungal denitrification in different natural ecosystems[J]. Acta Microbiologica Sinica, 2021, 61(6): 1551-1566. |