


养鸡场空气微生物污染及工人呼吸暴露风险
白渔樵,
孙兴滨,
仇天雷,
郭雅杰,
高敏,
王旭明
环境科学  2024, Vol. 45 Issue (3): 1840-1848 2024, Vol. 45 Issue (3): 1840-1848 |
|


近些年, 我国居民对肉蛋需求量逐渐增加, 其中每年对鸡蛋单项需求量高达3 000多万t, 常年稳居世界第一[1].集约化养殖模式因其成本低廉和生产效率高等优点被广泛应用于动物养殖业.该模式具有养殖密度大、空间狭小、且动物粪便日生成量大等特点.粪便中的微生物可随动物活动和粪便清理过程发生气溶胶化, 逸散进入空气[2].气载微生物可借助大气运动进行长距离传播, 对周边居民的健康造成潜在危害[3].Gao等[4]的研究显示, 空气是动物源微生物向人类环境扩散的重要介质.养殖场已经成为空气环境中微生物, 特别是致病微生物的重要来源[5].
养殖场空气环境中微生物浓度较高.有研究结果表明, 某些养殖场空气细菌浓度甚至超过了污水处理厂和垃圾填埋场[6].动物粪便被认为是养殖场空气微生物的重要来源[7], 1g动物粪便每小时可向空气释放1 000个可培养菌落, 其中80%为可通过呼吸进入肺部[8].利用分子生物学方法, Yang等[9]和Luiken等[10]证实养殖场动物粪便与空气颗粒物负载微生物的群落结构具有相关性.Song等[11]对养猪场的研究结果显示, 粪便对空气微生物OTU的贡献率为19.9%~59.9%.尽管众多学者对不同种类动物养殖场空气微生物的污染种类及可能来源做了大量分析, 但大部分研究都基于短期调查[10~12], 而随着养殖动物的生长发育以及季节变化, 其肠道菌群也发生改变, 导致其粪便的微生物群落结构在不同的养殖时间下也不尽相同[13, 14], 这使得该环境下空气微生物的污染现状无法系统评估.
养殖场空气中含有大量且多种类的微生物, 甚至是含有致敏和致病微生物的菌属[7].沙云菲等[15]的研究结果显示, 其所调查的4种动物养殖场的空气环境均检测到抗生素耐药菌, 其中包括对抗生素耐药的人类致病菌.已有研究证实了鸡泄殖腔-气溶胶-工人呼吸道微生物群落结构的相关性[16].长期暴露在该环境下, 导致养殖场工人的呼吸系统、眼部和皮肤疾病的发病率显著高于其他工作场所[17].为了对细菌可能引起的健康风险进行有针对性的预防, 研究人员对细菌的气溶胶化行为进行量化, 评估更易逸散入空气进而沉降在呼吸道的细菌[18, 19].但目前针对气溶胶化行为的研究主要集中于污水厂[18, 20]和堆肥厂[19, 21], 养殖场气溶胶化行为的研究鲜有报道[22], 而养殖场时间维度的气溶胶化行为研究更是未见报道.同时, 为了评估职业暴露对工人健康的影响, 近些年研究人员开始关注污染场所空气微生物的呼吸暴露量[23, 24].然而, 目前关于养殖场呼吸暴露量的相关研究较少, 仅有的一篇关于4种动物养殖场的短期研究, 结果表明养殖场工人每日摄入微生物量约107 copies·d-1, 其中致病菌属摄入量约104 copies·d-1[25].目前, 关于养殖场空气微生物的职业呼吸暴露的长期研究并无相关报道.
本研究以蛋鸡场为例, 对其中动物粪便和空气微生物展开80余周的研究.主要研究目的为:①分析养殖场舍内空气与动物粪便中微生物的生物多样性和群落结构的长期时间变化规律, 以及两种介质中微生物的相互关系;②阐明不同细菌菌属以及含有人类致病菌菌属的气溶胶化行为及其随时间的变化趋势;③在此基础上, 分析养殖场空气微生物的呼吸暴露量及其季节性规律.本研究结果将为评估养殖场空气污染现状以及制定相应的行业标准, 以降低工人因职业暴露引起的健康风险提供科学依据.
1 材料与方法 1.1 采样信息选取北京周边某蛋鸡养殖场, 对其空气和粪便样品分别进行长期采集, 样品编号为采样时蛋鸡周龄.采样日期与样品对照如表 1所示.空气样品的采集使用中流量TSP采样器(崂应, 中国), 在禽舍内距地面1.5 m处, 采样时间48 h, 气体流量100 L·min-1..粪便采用五点法采样, 之后将5份样品混合均匀成一份样品.每次采集的空气和粪便样品置于-80℃冰箱冷冻, 用于后续DNA提取.
|
|
表 1 采样日期与蛋鸡周龄对照1) Table 1 Comparison between sampling date and week age of layers |
采用MP FastDNA Spin Kit for Soil试剂盒(MP biomedicals, 美国)对空气及粪便样品进行DNA提取, 具体步骤根据试剂盒说明书.本研究中, 使用338F(5′-ACTCCTACGGGAGGCAGCAG-3′)和806R(5′-GGACTACHVGGGTWTCTAAT-3′)引物对16S rRNA基因的V3-V4可变区进行PCR扩增.PCR反应体系为5×FastPfu Buffer(4 µL)、2.5 mmol·L-1 dNTPs(2 µL)、正向Primer(5 µm)(0.8 µL)、反向Primer(5 µm)(0.8 µL)、BSA(0.2 µL)和模板DNA(10 ng), 最后补充ddH2O至20 µL的反应体系.使用的PCR扩增条件如下:预变性温度为95℃, 时间3 min;然后95℃变性30 s, 55℃退火30 s, 72℃延伸45 s, 循环40次;72℃保持10 min;最后10℃保温.用1%的琼脂糖凝胶进行电泳验证.所得DNA置于-20℃冰箱保存.扩增产物利用Illumina公司的MiSeq PE300平台进行测序分析.
1.3 气溶胶化指数(bioaerosolization index, BI)微生物的气溶胶化行为用气溶胶化指数(BI)来进行定义, 计算过程如公式(1)所示[19, 26]:
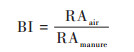
|
(1) |
式中, RAair为微生物在空气中的相对丰度, RAmanure为微生物在粪便中的相对丰度.本实验中, 先按最小样本丰度进行抽平, 抽平后某一分类单元的丰度与样品总丰度的比值即为该分类单元的相对丰度.
1.4 工人每日呼吸暴露量(daily intake, DI)利用工人每日吸入的微生物的总量来表示工人职业暴露量, 公式及呼吸量参照文献[23], 具体计算方法如公式(2)所示:
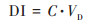
|
(2) |
式中, C为空气中微生物丰度, 单位为copies·m-3, VD为每人每天吸入空气的量, 单位为m3·d-1, 公式得出的DI单位为copies·d-1.吸入空气按每人20 m3·d-1计.
1.5 数据分析使用R 4.1.1的‘vegan’包进行Procrustes分析和Mantel test分析, Procrustes分析和Mantel test分析均基于Bray-Curtis距离, Procrustes分析通过最小二乘法最小化点坐标之间的偏差平方和(表示为M2), 小的偏差平方和代表了两数据集具有更高的一致性.Mantel test分析是将两种环境的微生物群落分别作为一个距离矩阵, 再检验两个矩阵之间的相关性.使用‘pheatmap’包绘制Heatmap图, ‘PerformanceAnalytics’包计算并绘制Pearson系数图, ‘ggplot2’包绘制气泡图矩阵.使用Origin 2018绘制堆积图和折线图.使用IBM SPSS 25进行显著性分析.
2 结果与讨论首先, 对养殖场空气和粪便中微生物含量、丰度和多样性进行检测, 并对其随养殖时间的变化趋势进行分析.如图 1所示, 空气和粪便的16S rRNA含量范围分别在6.08×105~4.90×106 copies·m-3和4.27×108~1.15×1010 copies·g-1内.空气和粪便样品16S rRNA含量的最大值均出现在第45周, 最小值则分别在第19周和第71周检出.总体而言, 两种介质中微生物含量呈现养殖中期较高, 初期和末期较低的趋势.对比分析粪便和空气中微生物含量的变化趋势, 发现空气相对于粪便有一个约为5d左右的滞后期.由此推断粪便对空气中微生物的贡献可能具有滞后性.

|
图 1 养殖场空气和粪便中微生物丰度及多样性随养殖时间分布规律 Fig. 1 Distribution pattern of microbial abundance and diversity in farm air and feces with growing time |
本研究利用Shannon指数来指示空气和粪便微生物的α多样性随养殖时间的变化趋势.空气微生物的Shannon指数(3.966~5.93)在所调查的养殖时间内均高于粪便, 且该差异性显著(P < 0.001).空气样本检出的OTU个数(655~1 751)同样显著大于粪便(P < 0.001).这与之前关于粪便和空气微生物的丰度和多样性相关研究的结论相同[27].从图 1可知, 空气和粪便样本中的微生物含量和多样性随养殖时间的变化呈现波动趋势, 这可能受到了季节因素的影响.Kumari等[28]对猪场的研究也发现了生物污染物具有季节性趋势.本研究采样时间长达80周, 北京不同季节空气温度波动较大, 动物养殖舍内保暖措施和通风频率均存在季节差异, 导致空气中微生物生存条件和影响因素随之发生相应变化.其他学者在对城市空气微生物的研究中也发现了其群落结构的季节性波动[29, 30].
为考察季节因素对粪便及空气微生物含量的影响, 本研究将样品采样时间按3~5月为春季、6~8月为夏季、9~11月为秋季和12月至次年2月为冬季进行划分, 对4个季节样本的微生物进行组间差异性分析, 结果如图 2所示.空气样本中细菌16S rRNA浓度平均值在冬季最高, 夏季最低, 且冬夏两季差异性显著(P < 0.05).有关猪场的研究中也发现其空气微生物具有季节性差异(冬季 > 秋季 > 夏季)[31], 这与本研究的结果一致.而空气微生物Shannon指数和OTU数的趋势相反, 冬季相应数值均为最低(P < 0.05).同样, 粪便样本的16S rRNA含量最高值也在冬季检出, 显著高于其他季节, 但Shannon指数和OTU数在各季节间均无显著差异.北京地区冬季气温低, 为保证鸡舍温度, 冬季鸡舍会将门窗关闭, 舍内空气不流通, 舍内空气难以向舍外逸散;而夏季鸡舍则会开窗通风, 利于舍内微生物向舍外流通, 导致空气中相应生物浓度较低.Song等[11]的研究也证实了由于冬季禽舍不利空气流通, 粪便对空气的OTU贡献率明显高于夏季.

|
P < 0.05为差异显著 图 2 养殖场空气和粪便微生物丰度及多样性的季节性分布规律 Fig. 2 Seasonal distribution patterns of air and feces microbial abundance and diversity on farm |
本研究首先对养殖场空气和粪便中细菌菌门进行分析, 空气和粪便样品中前5个优势菌门种类相同.其中, 相对丰度最高的菌门均为厚壁菌门(Firmicutes), 剩余4个菌门的相对丰度有所差异, 分别为拟杆菌门(Bacteroidetes)、放线菌门(Actinobacteria)、变形菌门(Proteobacteria)和梭杆菌门(Fusobacteria)(结果未展示).图 3展示了养殖场空气和粪便中丰度前30的优势菌属和包含致病菌的菌属.整体上, 空气细菌种类丰富度大于粪便, 这与图 1结果一致.养殖场空气中丰度最高前3个菌属较为稳定, 依次为乳杆菌属(Lactobacillus, 17%~0.98%), 拟杆菌属(Bacteroides, 17%~2%)和栖粪杆菌属(Faecalibacterium, 14%~1.7%).而动物粪便中优势菌属则随养殖时间的增加而呈现变化.在养殖前中后期, 乳杆菌属(Lactobacillus)和苏黎世杆菌属(Turicibacter)为优势菌属, 真杆菌属(Blautia)和瘤胃菌科UCG-014属(Ruminococcaceae_UCG-014)为第30~50周的优势菌属.在第50~64周, 梭杆菌属(Fusobacterium)为优势菌, 而考克氏菌属(Kocuria)则在第70周后相对丰度较高.相关研究指出, 禽类粪便微生物群落结构与其生长环境及饲料均有很大关系[14].同时, 随着养殖时间的增加, 禽类肠道微生物菌群也发生变化, 表现为相应粪便微生物群落结构的波动[32].

|
(a)空气中前30个优势菌属;(b)粪便中前30个优势菌属;(c)空气中致病菌属;(d)粪便中致病菌属 图 3 养殖场空气和粪便中优势菌属与致病菌属相对丰度随养殖时间分布趋势 Fig. 3 Trend of relative abundance of dominant and pathogenic genera in farm air and feces with growing time |
通过与已有致病菌名录的对比, 本研究在空气和粪便分别检测出包含致病菌的菌属46个和28个.乳杆菌属在空气中整体上丰度最高, 其次是拟杆菌属(Bacteroides), 这与先前的研究结果相似, 证实了这两个菌属是蛋鸡场致病菌中的优势菌[25].在养殖前期的第20~30周, 空气致病菌检测出的种类最多, 其丰富度随着养殖时间的增加有所降低.在所调查的养殖时间内, 动物粪便中优势致病菌属的种类和丰度均呈现波动趋势.乳杆菌属在所调查的80周内一直处于优势状态, 棒状杆菌属(Corynebacterium_1)在第58周和第64周有很高的检出.不动杆菌属(Acinetobacter)和拟杆菌属(Bacteroides)则在养殖中后期的粪便样本中处于优势, 第35周前检出很少.不动杆菌属广泛存在于自然界中, 其中的约翰逊不动杆菌(Acinetobocter johnsonii)是条件致病菌, 可引起菌血症、脑膜炎以及泌尿系统感染, 特别对于免疫力较低人群, 尤其是新生儿具有严重危害[33].棒状杆菌在山东省某鸭舍中也有较高的检出, 其在PM2.5和PM10中的相对丰度均大于30%, 其含有的纹带棒状杆菌(Corynebacterium striatum)可导致呼吸道和肺部感染, 引起白喉等疾病[12, 34].需要注意的是, 该菌属较易获得多重抗生素耐药性, 因此其对重症监护室患者、呼吸感染群体及新生儿的生命安全可构成重大威胁[35].
为探究空气与粪便样本中细菌的相关性, 本研究对两种介质中微生物结构和丰度的相互关系分别展开研究, 结果如图 4所示.图 4(a)和4(b)中的点分别表示采样时间和菌属, 空气和粪便按照蛋周龄一一对应, 分别表示细菌群落结构在时间维度的相关性和不同菌属在时间维度变化趋势的相关性.在80周的养殖时间内, 空气与粪便微生物的群落结构整体相关性不显著[P > 0.05, 图 4(a)].这可能是由于粪便和空气微生物随时间变化趋势不一致导致的.两种介质中的微生物通常受到不同因素的影响, 例如, 相对于粪便样本, 空气微生物群落结构的季节性差异更加明显(图 2).除此之外, 养殖场空气可能更易受到垫料、羽毛和当地空气环境的影响[11, 25].而动物粪便微生物的主要因素影响包括蛋鸡肠道菌群结构[14]、投喂的饲料和舍内卫生环境等[36].在所调查生长周期内, 空气和粪便中具体细菌菌属的变化趋势具有显著相关性[P < 0.001, 图 4(b)].进一步对空气与粪便样本中细菌含量的相互关系进行分析, 图 4(c)和4(d)分别展示了细菌和致病菌含量在空气和粪便中的数据分布、Pearson相关性及拟合曲线.Pearson相关性结果显示两种介质中细菌总含量[图 4(c)]和致病菌[图 4(d)]总含量均呈现正相关(r=0.70和r=0.68), 且显著性较强(P < 0.001).

|
(a)空气与粪便样本中微生物群落结构相关性分析(Procustes分析和Mantel test分析);(b)空气与粪便样本中细菌菌属相关性分析(Procustes分析和Mantel分析);(c)空气与粪便样本16S rRNA含量Pearson相关性;(d)空气与粪便样本中致病菌16S rRNA含量Pearson相关性;***表明P < 0.001 图 4 空气与粪便微生物群落结构及丰度的相关性分析 Fig. 4 Correlation analysis of the structure and abundance of air and feces microbial communities |
本研究进一步量化了动物粪便微生物进入空气的气溶胶化行为特征, 对气溶胶化指数随养殖时间的变化规律展开分析, 结果如图 5(a)和图 5(b)所示.整体上, 细菌气溶胶化指数随养殖时间的增加而呈上升趋势(P > 0.05), 而致病菌和养殖时间呈负相关(P > 0.05).具体优势细菌和致病菌属的气溶胶化趋势分别如图 5(c)和图 5(d)所示.结果显示, 不同菌属的气溶胶化指数差异较大, 同一种细菌菌属在不同的养殖时间变化趋势也并不一致.其中, 瘤胃菌属([Ruminococcus]_torques_group), 拟杆菌属(Bacteroides), 栖粪杆菌属(Faecalibacterium), 欧鲁森氏菌属(Olsenella), 萨特氏菌属(Sutterella)和罕见小球菌属(Subdoligranulum)在绝大部分养殖时间的BI值都大于0, 具有优先气溶胶化特征.对含有致病菌属的气溶胶化指数的分析结果表明[图 5(d)], 只有拟杆菌属(Bacteroides)明显比其他致病菌属更易气溶胶化.以上结果表明, 相比于养殖时间差异, 不同细菌之间的特性差异是影响微生物气溶胶化行为的主要因素.目前, 养殖场粪便中细菌的气溶胶化行为并无相关报道, Perrott等[37]在实验室对于细菌体外优先气溶胶化行的研究结果显示, 气溶胶化行为主要受细菌自身生物特性影响, 该结论与本研究的一致.鉴于近些年动物抗生素耐药性对人类健康造成的威胁, 将来的研究应着重于具有抗生素耐药性致病菌的气溶胶化行为, 以便深入理解其通过呼吸道沉降引发的职业风险[38, 39].

|
(a)共有细菌菌属气溶胶化指数整体变化趋势, (b)共有致病菌菌属气溶胶化指数整体变化趋势, (c)前30共有细菌菌属气溶胶化指数气泡图矩阵, (d)共有致病菌属气溶胶化指数气泡图矩阵;(a)和(b)中粗曲线为均值拟合曲线, 上下两条细曲线之间为标准误差带, 虚线为均值的线性拟合;(c)和(d)中点的大小表示BI值的大小关系, 以1为划分区别点的颜色(区别不同的气溶胶化行为), 大于1为红色, 表示其易逸散进入空气, 小于1为蓝色表示其不易逸散进入空气 图 5 养殖场微生物气溶胶化指数随养殖时间变化规律 Fig. 5 Variation pattern of microbial bioaerosolization index with growing time in the farm |
基于空气中细菌和致病菌的含量, 本研究计算了暴露于养殖场空气中, 工人每日通过呼吸系统对空气微生物的吸入总量, 结果如图 6所示.整体上, 细菌暴露量值在夏季最低[(1.76×107±7.36×106)copies·d-1], 冬季最高[(2.55×107±9.66×106)copies·d-1], 二者差异显著(one-way ANOVA, P < 0.05).工人对致病菌的日暴露量在不同季节性之间差异不显著.Xin等[25]对猪、牛、蛋鸡和肉鸡养殖场工人的呼吸暴露进行研究, 发现4种动物养殖环境的呼吸暴露量均在106~107 copies·d-1之间.其中, 蛋鸡最高, 暴露量高达107 copies·d-1, 这与本研究结果的整体趋势一致[25].对已发表研究中不同污染场所中空气微生物含量进行整理, 利用公式(2)对相应工作场所工人的细菌呼吸暴露量进行计算.结果显示整体上养殖场工人的日细菌呼吸暴露量低于堆肥厂[40], 但高于城市、郊区、实验室[41]、污水处理厂和垃圾填埋场[6].在所调查的80周内, 工人吸入主要致病菌属为乳杆菌属(Lactobacillus)和拟杆菌属(Bacteroides), 约占总沉降量的一半以上(76%±20%), 某些时间点(第16、45、47、49、61、77和83周)甚至超过90%.之前的研究表明, 养殖场生物气溶胶主要沉积在人体的咽喉和支气管部位[42], 同时报道显示, 集约化动物养殖场工人长时间暴露在空气微生物中, 其鼻窦炎、支气管炎等疾病的发病率较高, 且呼吸道及肺功能也受到相应的损害[43, 44].目前, 一些发达国家, 如英国、德国和荷兰等, 关于工人在动物养殖场的职业暴露风险具有成熟的评价标准[45], 而我国该方面并无相关政策和法规.本研究结果首次系统揭示了养鸡场空气中细菌呼吸暴露的长期结果, 需要引起相关部门关注.

|
(a)总细菌日暴露量;(b)总致病菌日暴露量;(c)致病菌属日暴露量(图例只显示丰度前5菌属) 图 6 养鸡场空气中细菌和致病菌的呼吸暴露季节性分布规律 Fig. 6 Seasonal distribution pattern of respiratory exposure of airborne bacteria and pathogenic bacteria in chicken farm |
本研究结果表明, 空气样本中细菌浓度平均值在冬季显著高于夏季, 而生物多样性趋势相反.蛋鸡养殖场空气与粪便中的优势细菌门整体上种类相同, 而两种介质中优势细菌菌属随养殖时间的增加变化趋势存在差异.虽然粪便和空气微生物的群落结构相关性不显著, 但二者的总细菌和致病菌含量随养殖时间呈现相似趋势.粪便中细菌的气溶胶化指数随养殖时间的增加而增加, 致病菌属趋势则相反.养鸡场工人的细菌和致病菌属的平均呼吸暴露量分别为2.54×107 copies·d-1和2.87×105 copies·d-1.本研究结果将为评估养殖场空气污染现状以及制定相应的行业标准, 以降低工人因职业暴露引起的健康风险提供科学依据.
| [1] |
韩太鑫. 我国蛋鸡行业发展趋势及模式创新思考[J]. 中国禽业导刊, 2022, 39(5): 18-21. Han T X. The development trend and model innovation of domestic egg chicken sector[J]. Guide to Chinese Poultry, 2022, 39(5): 18-21. |
| [2] | Chen M, Qiu T L, Sun Y M, et al. Diversity of tetracycline- and erythromycin-resistant bacteria in aerosols and manures from four types of animal farms in China[J]. Environmental Science and Pollution Research, 2019, 26(23): 24213-24222. DOI:10.1007/s11356-019-05672-3 |
| [3] | Bai H, He L Y, Wu D L, et al. Spread of airborne antibiotic resistance from animal farms to the environment: dispersal pattern and exposure risk[J]. Environment International, 2022, 158. DOI:10.1016/j.envint.2021.106927 |
| [4] | Gao M, Zhang X L, Yue Y, et al. Air path of antimicrobial resistance related genes from layer farms: emission inventory, atmospheric transport, and human exposure[J]. Journal of Hazardous Materials, 2022, 430. DOI:10.1016/j.jhazmat.2022.128417 |
| [5] | Cambra-López M, Torres A G, Aarnink A J A, et al. Source analysis of fine and coarse particulate matter from livestock houses[J]. Atmospheric Environment, 2011, 45(3): 694-707. DOI:10.1016/j.atmosenv.2010.10.018 |
| [6] |
于学政, 韩云平, 曹英楠, 等. 人为源微生物气溶胶的分布特征及风险研究进展[J]. 微生物学通报, 2023, 50(2): 667-686. Yu X Z, Han Y P, Cao Y N, et al. Progress in the distribution characteristics and risks of bioaerosols from anthropogenic sources[J]. Microbiology China, 2023, 50(2): 667-686. |
| [7] |
高敏, 仇天雷, 秦玉成, 等. 养鸡场空气中抗性基因和条件致病菌污染特征[J]. 环境科学, 2017, 38(2): 510-516. Gao M, Qiu T L, Qin Y C, et al. Sources and pollution characteristics of antibiotic resistance genes and conditional pathogenic bacteria in concentrated poultry feeding operations[J]. Environmental Science, 2017, 38(2): 510-516. |
| [8] | Chien Y C, Chen C J, Lin T H, et al. Characteristics of microbial aerosols released from chicken and swine feces[J]. Journal of the Air & Waste Management Association, 2011, 61(8): 882-889. |
| [9] | Yang Y, Zhou R J, Chen B W, et al. Characterization of airborne antibiotic resistance genes from typical bioaerosol emission sources in the urban environment using metagenomic approach[J]. Chemosphere, 2018, 213: 463-471. DOI:10.1016/j.chemosphere.2018.09.066 |
| [10] | Luiken R E C, Van Gompel L, Bossers A, et al. Farm dust resistomes and bacterial microbiomes in European poultry and pig farms[J]. Environment International, 2020, 143. DOI:10.1016/j.envint.2020.105971 |
| [11] | Song L, Wang C, Jiang G Y, et al. Bioaerosol is an important transmission route of antibiotic resistance genes in pig farms[J]. Environment International, 2021, 154. DOI:10.1016/j.envint.2021.106559 |
| [12] | Wu B, Qin L W, Wang M, et al. The composition of microbial aerosols, PM2.5, and PM10 in a duck house in Shandong province, China[J]. Poultry Science, 2019, 98(11): 5913-5924. DOI:10.3382/ps/pez365 |
| [13] |
陈东虹, 袁岩聪, 马雪惠, 等. 家禽肠道微生物的组成、分布及功能研究进展[J]. 中国家禽, 2022, 44(11): 121-127. Chen D H, Yuan Y C, Ma X H, et al. Research progress on composition, distribution and function of poultry intestinal microorganisms[J]. China Poultry, 2022, 44(11): 121-127. |
| [14] | 宋启明, 王瑞祥, 马宁. 鸡肠道菌群组成及主要影响因素[J]. 吉林畜牧兽医, 2022, 43(12): 73-74. |
| [15] |
沙云菲, 孙兴滨, 辛文鹏, 等. 4种动物养殖场空气中抗生素耐药菌的生物多样性及群落结构[J]. 环境科学, 2020, 41(2): 656-664. Sha Y F, Sun X B, Xin W P, et al. Bacterial diversity and community structure antibiotic-resistant bacteria in bioaerosol of animal farms[J]. Environmental Science, 2020, 41(2): 656-664. |
| [16] | Yang F, Gao Y L, Zhao H C, et al. Revealing the distribution characteristics of antibiotic resistance genes and bacterial communities in animal-aerosol-human in a chicken farm: from one-health perspective[J]. Ecotoxicology and Environmental Safety, 2021, 224. DOI:10.1016/j.ecoenv.2021.112687 |
| [17] | Just N, Kirychuk S, Gilbert Y, et al. Bacterial diversity characterization of bioaerosols from cage-housed and floor-housed poultry operations[J]. Environmental Research, 2011, 111(4): 492-498. DOI:10.1016/j.envres.2011.01.009 |
| [18] | Wang R N, Li X, Yan C. Seasonal fluctuation of aerosolization ratio of bioaerosols and quantitative microbial risk assessment in a wastewater treatment plant[J]. Environmental Science and Pollution Research, 2021, 28(48): 68615-68632. DOI:10.1007/s11356-021-15462-5 |
| [19] | He P J, Wei S Y, Shao L M, et al. Aerosolization behavior of prokaryotes and fungi during composting of vegetable waste[J]. Waste Management, 2019, 89: 103-113. DOI:10.1016/j.wasman.2019.04.008 |
| [20] | Lu F, Hu T Y, Wei S Y, et al. Bioaerosolization behavior along sewage sludge biostabilization[J]. Frontiers of Environmental Science & Engineering, 2021, 15(3). DOI:10.1007/s11783-020-1339-5 |
| [21] | Wang R N, Yu A Y, Qiu T L, et al. Aerosolization behaviour of fungi and its potential health effects during the composting of animal manure[J]. International Journal of Environmental Research and Public Health, 2022, 19(9). DOI:10.3390/ijerph19095644 |
| [22] | Xin H B, Qiu T L, Guo Y J, et al. Aerosolization behavior of antimicrobial resistance in animal farms: a field study from feces to fine particulate matter[J]. Frontiers in Microbiology, 2023, 14. DOI:10.3389/fmicb.2023.1175265 |
| [23] | Li L Y, Wang Q, Bi W J, et al. Municipal solid waste treatment system increases ambient airborne bacteria and antibiotic resistance genes[J]. Environmental Science & Technology, 2020, 54(7): 3900-3908. |
| [24] | Gao M, Yu A Y, Chen M, et al. Airborne fungi and human exposure in different areas of composting facilities[J]. Ecotoxicology and Environmental Safety, 2022, 243. DOI:10.1016/j.ecoenv.2022.113991 |
| [25] | Xin H B, Gao M, Wang X M, et al. Animal farms are hot spots for airborne antimicrobial resistance[J]. Science of the Total Environment, 2022, 851. DOI:10.1016/j.scitotenv.2022.158050 |
| [26] | Moletta-Denat M, Bru-Adan V, Delgenes J P, et al. Selective microbial aerosolization in biogas demonstrated by quantitative PCR[J]. Bioresource Technology, 2010, 101(19): 7252-7257. DOI:10.1016/j.biortech.2010.04.035 |
| [27] | Scott H M, Acuff G, Bergeron G, et al. Antimicrobial resistance in a One Health context: exploring complexities, seeking solutions, and communicating risks[J]. Annals of the New York Academy of Sciences, 2019, 1441(1): 3-7. DOI:10.1111/nyas.14057 |
| [28] | Kumari P, Choi H L. Seasonal variability in airborne biotic contaminants in swine confinement buildings[J]. PLoS One, 2014, 9(11). DOI:10.1371/journal.pone.0112897 |
| [29] |
雒月云, 毛怡心, 庄思琪, 等. 北京市不同季节室外细菌气溶胶分布特征及其环境影响因素分析[J]. 环境科学研究, 2022, 35(2): 556-565. Luo Y Y, Mao Y X, Zhuang S Q, et al. Seasonal distribution characteristics of bacterial aerosols and their correlations with environmental factors in Beijing, China[J]. Research of Environmental Sciences, 2022, 35(2): 556-565. |
| [30] | 杜鹏瑞, 杜睿, 任伟珊. 北京大气PM2.5中致病菌的变化特征[A]. 见: 第十三届全国气溶胶会议摘要集[C]. 石家庄: 中国颗粒学会气溶胶专业委员会, 2017. 17. |
| [31] |
程晓亮, 陈剑波, 王树华, 等. 不同季节猪舍内气溶胶中的细菌多样性分析[J]. 家畜生态学报, 2022, 43(6): 55-62. Cheng X L, Chen J B, Wang S H, et al. Diversity of bacteria in air of pig houses in different seasons[J]. Acta Ecologae Animalis Domastici, 2022, 43(6): 55-62. |
| [32] |
胡永飞, 呙于明. 家禽肠道微生物组研究进展[J]. 中国家禽, 2022, 44(11): 1-9. Hu Y F, Guo Y M. Research advances in poultry gut microbiome[J]. China Poultry, 2022, 44(11): 1-9. |
| [33] |
赵醒醒, 蔡燕, 李丽利, 等. 约翰逊不动杆菌致新生儿迟发型败血症2例[J]. 微生物与感染, 2022, 17(4): 235-238. Zhao X X, Cai Y, Li L L, et al. Two cases of late-onset sepsis in newborns caused by Acinetobacter johnsonii[J]. Journal of Microbes and Infections, 2022, 17(4): 235-238. |
| [34] | Saito S, Kawamura I, Tsukahara M, et al. Cellulitis and bacteremia due to Corynebacterium striatum identified by matrix-assisted laser desorption ionization-time of flight mass spectrometry[J]. Internal Medicine, 2016, 55(9): 1203-1205. DOI:10.2169/internalmedicine.55.5484 |
| [35] |
顾珉, 沈海英, 顾炳仁, 等. 药品抽验中检出鲍曼不动杆菌案例的风险评估回顾和策略探讨[J]. 中国药品标准, 2020, 21(6): 487-493. Gu M, Shen H Y, Gu B R, et al. Review and discussion on risk assessment of cases with Acinetobacter baumannii detected in national drug sampling[J]. Drug Standards of China, 2020, 21(6): 487-493. |
| [36] | Grzyb J, Podstawski Z, Bulski K. Bacterial aerosol, particulate matter, and microclimatic parameters in the horse stables in Poland[J]. Environmental Science and Pollution Research, 2022, 29(18): 26992-27006. DOI:10.1007/s11356-021-18142-6 |
| [37] | Perrott P, Turgeon N, Gauthier-Levesque L, et al. Preferential aerosolization of bacteria in bioaerosols generated in vitro[J]. Journal of Applied Microbiology, 2017, 123(3): 688-697. DOI:10.1111/jam.13514 |
| [38] | Lowder B V, Guinane C M, Ben Zakour N L, et al. Recent human-to-poultry host jump, adaptation, and pandemic spread of Staphylococcus aureus[J]. Proceedings of the National Academy of Sciences of the United States of America, 2009, 106(46): 19545-19550. |
| [39] | Brown-Jaque M, Calero-Cáceres W, Muniesa M. Transfer of antibiotic-resistance genes via phage-related mobile elements[J]. Plasmid, 2015, 79: 1-7. DOI:10.1016/j.plasmid.2015.01.001 |
| [40] | Ferguson R M W, Neath C E E, Nasir Z A, et al. Size fractionation of bioaerosol emissions from green-waste composting[J]. Environment International, 2021, 147. DOI:10.1016/j.envint.2020.106327 |
| [41] | Tao Y L, Yue Y, Wang J. Abundance and diversity of antibiotic resistance genes possibly released to ambient air by experiments in biology laboratories[J]. Science of the Total Environment, 2021, 797. DOI:10.1016/j.scitotenv.2021.149147 |
| [42] |
张兰河, 贺雨伟, 陈默, 等. 畜禽养殖场空气中可培养抗生素耐药菌污染特点研究[J]. 环境科学, 2016, 37(12): 4531-4537. Zhang L H, He Y W, Chen M, et al. Pollution characteristics of antibiotic resistant bacteria from atmospheric environment of animal feeding operations[J]. Environmental Science, 2016, 37(12): 4531-4537. |
| [43] | Heederik D, Sigsgaard T, Thorne P S, et al. Health effects of airborne exposures from concentrated animal feeding operations[J]. Environmental Health Perspectives, 2007, 115(2): 298-302. DOI:10.1289/ehp.8835 |
| [44] | Li H, Zhou X Y, Yang X R, et al. Spatial and seasonal variation of the airborne microbiome in a rapidly developing city of China[J]. Science of the Total Environment, 2019, 665: 61-68. DOI:10.1016/j.scitotenv.2019.01.367 |
| [45] | Robertson S, Douglas P, Jarvis D, et al. Bioaerosol exposure from composting facilities and health outcomes in workers and in the community: a systematic review update[J]. International Journal of Hygiene and Environmental Health, 2019, 222(3): 364-386. DOI:10.1016/j.ijheh.2019.02.006 |