 2024, Vol. 45
2024, Vol. 45



2. 国家海洋环境监测中心国家环境保护海洋生态环境整治修复重点实验室, 大连 116023
2. State Environmental Protection Key Laboratory of Marine Ecosystem Restoration, National Marine Environmental Monitoring Center, Dalian 116023, China
滨海湿地的蓝碳功能和增汇潜力已成为缓解全球气候变化的长期解决方案之一, 也是我国基于自然实现“双碳”目标的重要解决方案.有研究表明, 滨海湿地单位面积碳埋藏速率是陆地生态系统的几十到上千倍[1], 具有强大的固碳能力.然而, 因滨海湿地处于陆地和海洋的交错过渡地带[2], 是气候变化的敏感区和人类活动的频繁区, 气温升高、降水波动和土地利用(如围垦、水产养殖和工业生产)等扰动决定着滨海湿地的蓝碳功能及其变化趋势.其中, 土地利用方式的改变会直接导致滨海湿地面积的快速减损, 进而引起湿地固碳能力的下降.我国海岸带蓝碳生态系统中, 盐沼湿地是总储碳量和年碳汇量的主要贡献者[3].但在自然活动和人类活动的影响下, 20世纪80年代至21世纪初我国滨海盐沼湿地面积减少了近60%[4].如在1987 ~ 2007年, 盐城市盐沼湿地面积缩小了53%, 而表层(0 ~ 0.2 m)土壤有机碳储量减少了46.3%[5].因此, 为实现固碳减排和避免蓝碳功能的快速丧失, 亟需对受损滨海湿地生态系统开展生态修复.“退塘还湿”是我国滨海湿地的保护修复手段之一, 也是近几年开展较多的滨海湿地修复形式.然而, “退塘还湿”修复区碳汇功能的修复效果评估研究当下较为匮乏.
目前, 涡度相关法在全球碳水循环研究中广泛应用.近些年, 国内外学者在美国特拉华州[6]、爱尔兰西部[7]等以及我国的辽河口[8]、长江口[9, 10]和黄河口[11]等地均开展了关于滨海湿地CO2通量的研究.目前, 相关研究多数针对自然滨海湿地和受损滨海湿地, 研究内容集中在CO2交换特征和动态变化、潮汐过程中潮汐淹水和干湿交替对盐沼湿地碳交换及碳汇形成机制的影响等[11 ~ 13].但已有研究缺乏对受损滨海湿地开展生态修复后CO2交换动态特征及环境因子影响机制、碳源/汇功能的现状与潜力的研究.
20世纪80年代以来, 辽河口滨海湿地区域围塘养殖面积曾高达1.7×104 hm2, 导致生物栖息地丧失、生物多样性下降, 生态系统碳汇功能退化[14].为保护与修复我国滨海湿地, 自2015年起在辽河口区域开展了大规模的“退塘还湿”生态修复工程.截至2020年, “退塘还湿”生态工程修复面积达5.73×103 hm2, 修复自然海岸线17.6 km[15].辽河口“退塘还湿”修复工程的开展为探讨生态修复对生态系统CO2交换及其调控机制提供了绝佳的研究背景, 针对该区域的CO2交换过程变化及影响机制的研究, 将弥补自然与人类活动共同作用下的滨海湿地碳交换研究.本文基于涡度相关法, 依托辽河口“退塘还湿”生态修复区, 探究不同时间尺度CO2交换规律, 分析影响CO2交换的主控因子及其响应机制, 评估修复区盐沼湿地的碳源/汇能力, 以期为评价生态修复工程效果提供数据支持和理论依据.
1 材料与方法 1.1 研究区域概况辽宁辽河口滨海湿地位于辽河平原南部, 辽东湾顶部, 是由大凌河、辽河和大辽河入海交汇冲刷形成, 是我国北方最大的河口湿地, 湿地面积约8.0×104 hm2.该区域气候类型为暖温带大陆性半湿润季风气候, 年平均气温8.4℃, 年平均降水量623.2 mm, 降雨集中在夏季, 雨热同期.区域内地势平缓, 主要分布有苇田、沼泽草地和滩涂等生境类型, 优势植被主要为芦苇(Phragmites australis)和盐地碱蓬(Suaeda salsa).土壤类型有盐土、草甸土和潮滩土等, 土壤盐分含量由沿海向内陆呈现递减趋势[16, 17].该研究依托国家海洋环境监测中心盘锦滨海湿地生态站, 样地位于辽河入海口西侧的“退塘还湿”修复区内(40°56′15.26″N, 121°46′42.55″E).修复初期, 盐地碱蓬作为先锋物种大面积分布.随着潮沟不断冲刷演变, 部分高地盐地碱蓬逐渐被芦苇替代.开路式涡度相关观测系统于2015年安装在距地面8.5 m高的塔上, 各方向风浪区长度超过300 m, 满足涡度相关技术有效应用的基本假设.
1.2 研究方法研究采用开路式涡度相关观测系统与微气象观测系统进行长期连续的定位监测.涡度相关观测系统包括开路红外CO2/H2O分析仪(LI-7500A, LI-COR, USA)和三维超声风速仪(WindMaster Pro, Gill, UK), 原始数据采集频率为10 Hz.微气象观测系统包括4 m高处的风向/风速(02C, Campbell, USA);空气温度/湿度(HMP155, Vaisala, Finland);光合有效辐射(LI-190, LI-COR, USA);降雨量(TR-525USW, Texas Electronics, USA).土壤含水量(Teros12)采样深度为10、20和30 cm, 土壤热通量板(HFP01)深度为地下8 cm.所有数据通过数据采集器(Model 9210, XLite)在线采集, 采集后以30 min为间隔计算平均值后储存.
1.3 数据来源与处理开路式涡度相关观测系统会因天气突变、电力供应、仪器标定和维护等原因产生数据测量野点和出现数据缺失现象.另外, 研究区域的空气中苇絮较多、盐分较高, 也会影响仪器测量精度.由于上述情况, 涡度相关系统获得的CO2通量并非全部有效.因而, 需要对获得的原始数据进行质量控制和数据插补, 数据质量控制及缺失数据的插补过程参考相关文献[18].经过质量控制后, 本研究获得的白天CO2通量有效数据量为75.4%, 夜间CO2通量有效数据量为26.5%.
2 结果与分析 2.1 环境因子变化2021年辽河口修复区空气温度(Ta)呈单峰变化趋势且振幅较大.1 ~ 7月Ta呈上升趋势, 7 ~ 12月Ta呈下降趋势;全年平均气温9.96℃, 月平均最低气温和最高气温分别出现在1月(-9.19℃)和7月(25.7℃).光合有效辐射(photosynthetically active radiation, PAR)全年呈现单峰变化趋势, 但与Ta不同的是, PAR在1 ~ 5月逐渐升高, 而后逐渐下降, 其中7、8月持平, 11、12月持平.全年平均PAR为267.1 μmol·(m2·s)-1, 月平均最小值和最大值分别出现在1月[148.23 μmol·(m2·s)-1]和5月[411.73 μmol·(m2·s)-1, 见图 1(a).2021年, 辽河口修复区生长季土壤含水量(soil water content, SWC)、饱和水汽压差(vapor pressure deficit, VPD)、土壤温度(soil temperature, Ts)和相对湿度(relative humidity, RH)月均值范围分别为29.41% ~ 45.2%、4.52 ~ 6.86 hPa、5.16 ~ 22.38℃和49.10% ~ 82.83%[图 1(b)和图 1(c)].

|
(a)2021年Ta和PAR各月平均值;(b)生长季SWC、VPD各月平均值;(c)生长季Ts和RH各月平均值 图 1 辽河口修复区2021年气象因子和生长季环境因子月均值 Fig. 1 Monthly averages of meteorological factors and environmental factors in the growing season of 2021 in the restored area of Liaohe River estuary |
将2021年修复区每月净生态系统CO2交换(net ecosystem CO2 exchange, NEE)的每天半小时数据计算平均值, 得到每月48个时刻NEE数据, 绘制修复区四季NEE平均日变化特征曲线(图 2).修复区的春季和秋季NEE日变化呈“U”型, 夏季呈“V”型, 而在冬季呈“一”型.“U”型曲线特征表现为:日出后NEE由正转负, 增加至第一个吸收峰, 小段时间内吸收略有减少, 再次增加至第二个吸收峰后逐渐减小, 日落后NEE由负转正, 吸收峰出现在10:00 ~ 14:00.“V”型曲线特征表现为:日出后NEE由正转负, 增加至第一个吸收峰前吸收略有减少, 达到吸收峰后逐渐减小, 日落后NEE由负转正, 吸收峰出现在10:00 ~ 12:30.NEE日变化幅度表现为:夏季 > 春季 > 秋季 > 冬季.春季(3 ~ 5月)修复区各月CO2吸收量逐渐增加, 季节效率(以C计, 下同)为-40.06 g·m-2;夏季(6 ~ 8月)修复区各月吸收CO2状态可观, 季节效率为-63.62 g·m-2;秋季(9 ~ 11月)修复区各月CO2吸收量逐渐减少, 在10月出现急剧下降, 季节效率为2.33 g·m-2;冬季(1、2和12月)修复区白天几乎不吸收CO2, 整体表现为向外释放CO2, 季节效率为34.43 g·m-2.

|
图 2 2021年辽河口修复区四季NEE平均日变化 Fig. 2 Average daily variation in NEE in the restored area of Liaohe River estuary in four seasons of 2021 |
2021年修复区NEE日累积年内变化整体呈“V”型[图 3(a)], 湿地生态系统在第20日(1月20日)净释放CO2能力最强, 为1.43 g·(m2·d)-1;在第123日(5月3日)净吸收CO2的能力最强, 为-2.42 g·(m2·d)-1.在第63日(3月4日)前日NEE波动幅度整体较小, 从第63日起湿地NEE波动幅度突然变大, 且一直持续到第313日(11月9日), 此后湿地NEE波动幅度整体逐渐变小.2021年修复区NEE、生态系统呼吸(ecosystem respiration, Reco)和总的初级生产力(gross primary productivity, GPP)月累积变化明显.NEE月累积范围为-30.80 ~ 15.88 g·(m2·month)-1[图 3(b)], 最大值和最小值分别出现在5月和1月.NEE月累积在1 ~ 5月逐渐增加, 6月减少, 7月再次增加, 8 ~ 11月逐渐减少, 12月与11月基本持平.Reco月累积范围为7.55 ~ 27.35 g·(m2·month)-1, 最大值和最小值分别出现在6月和3月.Reco月累积值在1 ~ 3月逐渐减少, 4 ~ 6月逐渐增加, 6、7和8月基本持平, 8 ~ 9月逐渐减少.GPP月累积范围为0 ~ 58.14 g·(m2·month)-1, 最大值出现在7月.GPP月累积值在4月由0转正, 4 ~ 7月逐渐增加至峰值, 8 ~ 10月逐渐减少, 11月降至0.

|
图 3 2021年辽河口修复区NEE日累积年内变化及NEE、Reco和GPP月累积变化 Fig. 3 Cumulative intra-annual variation in daily NEE and monthly cumulative changes in NEE, Reco, and GPP in the restored area of Liaohe River estuary in 2021 |
基于Michaelis-Menten模型, 对2021年修复区盐沼湿地生态系统白天PAR与NEE进行拟合, PAR与NEE呈直角双曲线关系(图 4).NEE随PAR增强而增大, 达到光饱和点时NEE不再增加.初始PAR增强时NEE的增量较大, 曲线弯曲程度大, 后续随PAR的增加NEE的增量逐渐减小, 曲线弯曲程度也趋于平缓.通过上述模型对盐沼湿地生态系统的光合拟合参数进行计算得最大光合速率(Amax)、表观初始光能利用率(α, 以CO2/photons计)和白天生态系统呼吸(Reco,day, 以CO2计)分别为(6.87±0.29)μmol·(m2·s)-1、(0.007±0.001)μmol·μmol-1和(0.48±0.01)μmol·(m2·s)-1.曲线R2 = 0.53, 因此PAR可以解释白天NEE变化的53%, 即PAR是白天光合作用大小的重要调控因子.

|
图 4 2021年辽河口修复区白天PAR与NEE的拟合曲线 Fig. 4 Fitting curve of PAR and daytime NEE in the restored area of Liaohe River estuary in 2021 |
Ta是Reco,night的主控因子.本研究选取5.5 ℃为分界点, 分别讨论Ta与Reco,night的关系, 是由于Ta较高时, Reco,night的温度敏感性较低;Ta较低时, Reco,night的温度敏感性较高[19], 而本研究修复区全年Ta范围为-22.52 ~ 32.36℃, 即振幅较大.因此, 根据实际情况及已有研究[19], 分Ta < 5.5℃和Ta ≥ 5.5℃两种情况分别拟合分析Ta与Reco,night关系(图 5), 表达式如下:
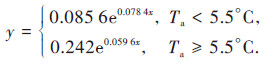
|
(1) |

|
图 5 2021年辽河口修复区Ta与Reco, night的拟合曲线 Fig. 5 Fitting curves of Ta and Reco,night in the restored area of Liaohe River estuary in 2021 |
Ta < 5.5℃时, Q10 = 2.19, R2 = 0.42, 即Ta可以解释Reco,night变化的42%;Ta ≥ 5.5℃时, Q10 = 1.81, R2 = 0.51, 即Ta可以解释Reco,night变化的51%.直观上, Ta < 5.5℃时的曲线斜率明显大于Ta ≥ 5.5时曲线的斜率;数值上, Ta < 5.5℃时的Q10大于Ta ≥ 5.5℃时的Q10, 即不同的温度范围Reco,night对Ta的敏感性不同.
2.6 生长季NEE对SWC、VPD、Ts和RH的响应修复区盐沼湿地生长季NEE与SWC、VPD、Ts和RH通过正态性检验后, 进行相关性分析(表 1), 结果呈显著相关的参数进行线性拟合(图 6).结果表明, NEE与SWC和VPD均呈显著的线性负相关(P < 0.05), 相关系数为-0.914和-0.811.NEE与SWC回归方程R2 = 0.84, 即认为SWC能够解释生长季NEE变化的84%;NEE与VPD回归方程R2 = 0.66, 即认为VPD能够解释生长季NEE变化的66%.修复区生长季NEE随SWC、VPD的增大而不断减小, 即生态系统的碳汇能力增强.而NEE与Ts和RH无显著相关(P > 0.05), 本研究认为Ts与RH的变化不会引起NEE的显著变化.
|
|
表 1 2021年辽河口修复区生长季NEE与SWC、VPD、Ts和RH相关性分析1) Table 1 Correlation analysis of NEE with SWC, VPD, Ts and RH in the restored area of Liaohe River estuary during the growing season of 2021 |

|
图 6 2021年辽河口修复区生长季NEE与SWC、VPD、Ts和RH月均值的线性回归关系 Fig. 6 Linear regression relationship between NEE and monthly averages of SWC, Ts, VPD, and RH in the restored area of Liaohe River estuary during the growing season of 2021 |
辽河口“退塘还湿”修复区NEE平均日变化曲线在春、夏和秋季表现为白天是负值(CO2汇), 夜晚是正值(CO2弱源), 在冬季昼夜源/汇功能变化不显著, 本研究NEE日变化模式与已有研究一致[20].此外本研究发现5月NEE平均日变化曲线峰值[-3.5 μmol·(m2·s)-1, 以CO2计, 下同]较大, 且与植被生长旺盛的7月日变化峰值[-3.64 μmol·(m2·s)-1]相近.这与多数盐沼湿地NEE平均日变化曲线峰值仅出现在夏季的现象不同[12, 21, 22], 原因可能是修复区5月的PAR月平均值最高, 增强了植物的光合作用[23], 且5月盐地碱蓬正处于营养生长期[12], 植物快速生长对环境因子的需求与PAR的增强相符合.修复区湿地四季NEE平均日变化曲线波动幅度不同, 在植物生长旺盛的夏季NEE日变化波动幅度最大, 在植物枯萎死亡的冬季NEE日变化波动稳定在0 ~ 1 μmol·(m2·s)-1, 以上结果主要是湿地季节水热条件差异引起的[24].现有研究对生长季NEE日变化曲线的描述有两种:“U”型[24, 25]和“V”型[21].两种类型曲线不同点为:“U”型曲线底部平缓, 且大多有两个相似的峰值;“V”型曲线底部陡峭, 大多只有一个峰值.相同点为:NEE在12点前后均表现为碳汇能力减弱, 这可能是由于正午气温较高, 植物为减少蒸腾作用流失过多水分而关闭了部分气孔, 导致光合作用强度减弱, 生态系统净吸收CO2的量减小[26], 即碳汇能力减弱.
3.2 NEE的环境调控机制2021年全年, 辽河口“退塘还湿”修复区白天NEE和夜间NEE(Reco,night)最主要的调控因子分别为PAR与Ta.在白天, PAR与NEE的拟合曲线呈直角双曲线关系, 变化趋势与以往研究结果一致[18, 21, 24, 25].在夜间, Ta < 5.5℃和Ta ≥ 5.5℃时Ta与Reco,night的拟合曲线均呈指数关系[27, 28], 且Ta-Reco,night关系模型侧重于0 ~ 30℃的有限温度范围[29], 但多数研究未对Ta分层处理.本研究以5.5℃为分界点分析全年Reco,night与Ta的关系发现:低温范围内Q10较高, 高温范围内Q10较低, 两者均在湿地生态系统Q10范围(1.75 ~ 4.99)内[30].在较低Ta下, 温度是影响土壤中微生物的新陈代谢活动的主控因子, 体现为低气温时Reco,night对Ta更敏感;在较高Ta下, 生态系统的新陈代谢则主要受水、养分和生物群落变化的影响, 体现为高气温时Reco,night对Ta不敏感[19, 31 ~ 33].此外, 本研究中非生长季Ta月均值小于5.5℃, 生长季Ta月均值大于5.5℃, 非生长季时的土壤可能经历了多次冻结-融化过程.因此猜测在非生长季冻融作用对土壤呼吸产生“激发效应”, 即土壤结冻时表面形成的冰膜阻碍CO2向外排放而聚集在土壤内部, 土壤融化时聚集的CO2短时间内向外排放形成高峰, 体现为低气温时Reco,night变化量更大[34, 35].
生长季内, NEE与SWC、VPD均呈显著负相关(P < 0.05), 即SWC和VPD越大NEE值越小, 且SWC比VPD可以更好地解释NEE的变化.与SWC对陆地生态系统呼吸的影响不同, SWC过低通常不会影响湿地生态系统土壤呼吸, 而SWC过高会减少生态系统呼吸排放的CO2 [36], 最终表现为系统净吸收CO2的量增加.已有研究表明[37, 38], 高VPD会诱导气孔关闭, 导致植物可利用CO2浓度下降, 光合作用强度减弱, 即NEE与VPD呈正相关.但本研究中湿地净吸收CO2的量并未因气孔关闭而明显减少, 原因可能是区域内优势植被盐地碱蓬属于C4植物, C4植物的叶肉细胞可通过“CO2泵”的方式利用叶片内细胞间隙含量较低的CO2, 以此实现在低CO2浓度条件下保持较高的光合作用能力[39, 40].因此, 本研究生长季NEE与VPD没有表现为正相关, 但二者呈负相关的具体机制仍需进一步探究.
3.3 “退塘还湿”生态修复区碳汇功能滨海盐沼湿地一般表现为碳汇, 不同滨海盐沼湿地碳汇能力见表 2.但修复盐沼湿地碳源/汇功能会因不同的修复年限和形式存在显著差异[41 ~ 43], 如美国新泽西州修复湿地在2014和2015年均表现为碳源, 在2016年表现为碳汇[41].本研究中, 2021年辽河口“退塘还湿”生态修复区湿地固碳量为-66.89 g·(m2·a)-1, 其碳汇能力接近2005年盘锦芦苇湿地[44][-65.00 g·(m2·a)-1], 原因可能是2005年盘锦芦苇湿地Reco/GPP值为0.95, 本研究修复区湿地Reco/GPP为0.7, 即2005年植物和土壤呼吸消耗了大量的植物吸收的碳[45], 导致其年固碳量偏小.我国其他河口盐沼湿地碳汇能力多数大于本研究结果, 如2018年上海九段沙芦苇湿地[10]和2011年黄河三角洲芦苇湿地[24]固碳量分别为-869.00 g·(m2·a)-1和-599.05 g·(m2·a)-1, 两区域的优势植被为芦苇, 生物量显著大于辽河口修复区, 且年平均Ta也高于本研究, 利于植物进行光合作用, 进而增加了生态系统净CO2吸收量.就日均固碳量而言, 本研究2021年生长季辽河口“退塘还湿”生态修复区湿地日均固碳量[-0.55 g·(m2·d)-1]高于2011 ~ 2015年生长季黄河口盐地碱蓬湿地[12]日均固碳量[-0.17 ~ -0.10 g·(m2·d)-1], 主要是由于黄河口的盐地碱蓬株高(20 ~ 40 cm)低于本研究的盐地碱蓬(50 ~ 65 cm).以互花米草(Spartina alterniflor)为优势植被的美国新泽西州修复湿地在2014 ~ 2016年生长季日均固碳量[-2.34 ~ -0.78 g·(m2·d)-1]高于本研究结果, 原因可能是互花米草的光合作用活性强, 湿地净吸收CO2的量大[41], 以及修复年限(15 a)远大于本研究修复区年限(6 a), 湿地生态系统功能趋于完善.综上, 2021年辽河口“退塘还湿”生态修复区盐沼湿地碳汇能力虽不及同区域内的自然湿地[-161.92 g·(m2·a)-1][18], 但其具有长期的碳增汇潜力.
|
|
表 2 不同滨海盐沼湿地的碳汇能力比较 Table 2 Comparison of carbon sink capacities of different coastal salt marshes |
4 结论
(1)2021年辽河口“退塘还湿”生态修复区盐沼湿地NEE平均日变化曲线在春季和秋季呈“U”型, 在夏季呈“V”型, 在冬季呈“一”型.2021年辽河口“退塘还湿”生态修复区四季NEE平均日变化曲线的整体振幅由大到小为:夏季 > 春季 > 秋季 > 冬季, 春季效率为-40.06 g·m-2, 夏季效率为-63.62 g·m-2, 秋季效率为2.33 g·m-2, 冬季效率为34.43 g·m-2.
(2)2021年修复区NEE日累积年内变化整体呈“V”型, NEE、Reco和GPP月累积差异明显.
(3)2021年全年辽河口“退塘还湿”生态修复区白天NEE与PAR呈直角双曲线关系, PAR可以解释白天NEE变化的53%;夜晚NEE与Ta呈指数关系, Ta < 5.5℃时Q10为2.19, Ta可以解释Reco,night变化的42%, Ta ≥ 5.5℃时Q10为1.81, Ta可以解释Reco,night变化的51%.
(4)2021年生长季辽河口“退塘还湿”生态修复区NEE与SWC和VPD均呈显著的线性负相关, 而NEE与Ts和RH无显著相关.
(5)2021年辽河口“退塘还湿”生态修复区盐沼湿地总净固碳量为-66.89 g·m-2, 表现为碳汇.
| [1] |
王法明, 唐剑武, 叶思源, 等. 中国滨海湿地的蓝色碳汇功能及碳中和对策[J]. 中国科学院院刊, 2021, 36(3): 241-251. Wang F M, Tang J W, Ye S Y, et al. Blue carbon sink function of Chinese coastal wetlands and carbon neutrality strategy[J]. Bulletin of Chinese Academy of Sciences, 2021, 36(3): 241-251. |
| [2] |
段云莹, 裴绍峰, 廖名稳, 等. 辽河三角洲滨海湿地水域初级生产力研究进展[J]. 海洋地质前沿, 2022, 38(6): 16-24. Duan Y Y, Pei S F, Liao M W, et al. Research progress on primary productivity of coastal wetland waters in Liao River Delta[J]. Marine Geology Frontiers, 2022, 38(6): 16-24. |
| [3] | Gu J L, Luo M, Zhang X J, et al. Losses of salt marsh in China: trends, threats and management[J]. Estuarine, Coastal and Shelf Science, 2018, 214: 98-109. DOI:10.1016/j.ecss.2018.09.015 |
| [4] |
李捷, 刘译蔓, 孙辉, 等. 中国海岸带蓝碳现状分析[J]. 环境科学与技术, 2019, 42(10): 207-216. Li J, Liu Z M, Sun H, et al. Analysis of blue carbon in China' s coastal zone[J]. Environmental Science & Technology, 2019, 42(10): 207-216. |
| [5] |
许振, 左平, 王俊杰, 等. 土地利用变化对盐城滨海湿地土壤有机碳库的影响[J]. 海洋通报, 2014, 33(4): 444-450. Xu Z, Zuo P, Wang J J, et al. Effects of land use changes on the organic carbon storage of the soil in Yancheng coastal wetlands[J]. Marine Science Bulletin, 2014, 33(4): 444-450. |
| [6] | Vázquez-Lule A, Vargas R. Biophysical drivers of net ecosystem and methane exchange across phenological phases in a tidal salt marsh[J]. Agricultural and Forest Meteorology, 2021, 300. DOI:10.1016/j.agrformet.2020.108309 |
| [7] | McVeigh P, Sottocornola M, Foley N, et al. Meteorological and functional response partitioning to explain interannual variability of CO2 exchange at an Irish Atlantic blanket bog[J]. Agricultural and Forest Meteorology, 2014, 194: 8-19. DOI:10.1016/j.agrformet.2014.01.017 |
| [8] |
刘凯, 杨继松, 袁晓敏, 等. 辽河口盐沼和光滩CO2排放通量及其影响因素[J]. 湿地科学, 2017, 15(6): 855-861. Liu K, Yang J S, Yuan X M, et al. CO2 emission flux and its influence factors in salt marsh and mudflat in the Liaohe estuary[J]. Wetland Science, 2017, 15(6): 855-861. |
| [9] |
贺贵书. 下垫面农林利用对崇明东滩围垦湿地CO2通量的影响[D]. 上海: 华东师范大学, 2020. He G S. The impact of agroforestry use and undering surface changes on CO2 fluxes of the reclamation wetland at Dongtan of Chongming Island[D]. Shanghai: East China Normal University, 2020. |
| [10] |
陈梓涵. 九段沙潮汐盐沼湿地CO2通量及影响机制研究[D]. 上海: 华东师范大学, 2020. Chen Z H. Ecosystem carbon flux and its influencing mechanisms over a salt marsh in the Jiuduansha Shoals[D]. Shanghai: East China Normal University, 2020. |
| [11] |
贺文君, 韩广轩, 许延宁, 等. 潮汐作用下干湿交替对黄河三角洲盐沼湿地净生态系统CO2交换的影响[J]. 应用生态学报, 2018, 29(1): 269-277. He W J, Han G X, Xu Y N, et al. Effects of drying and wetting cycles induced by tides on net ecosystem exchange of CO2 over a salt marsh in the Yellow River Delta, China[J]. Chinese Journal of Applied Ecology, 2018, 29(1): 269-277. |
| [12] |
邢庆会. 黄河三角洲潮汐盐沼湿地净生态系统CO2交换及影响机制[D]. 烟台: 中国科学院大学(中国科学院烟台海岸带研究所), 2018. Xing Q H. Net ecosystem CO2 exchange and its influencing mechanisms in a tidal salt marsh in the Yellow River Delta, China [D]. Yantai: University of Chinese Academy of Sciences(Yantai Institute of Coastal Zone Research, Chinses Academy of Sciences), 2018. |
| [13] |
赵明亮. 淹水梯度对黄河三角洲芦苇湿地生态系统碳交换的影响[D]. 重庆: 西南大学, 2021. Zhao M L. Effect of inundation depth on ecosystem carbon exchange in a reed wetland in the Yellow River Delta[D]. Chongqing: Southwest University, 2021. |
| [14] |
索安宁, 于永海, 韩富伟. 辽河三角洲盘锦湿地景观格局变化的生态系统服务价值响应[J]. 生态经济, 2011(6): 147-151. Suo A N, Yu Y H, Han F W. Response of ecosystem service value to wetland landscape pattern change in Panjin region of Liaohe Delta[J]. Ecological Economy, 2011(6): 147-151. |
| [15] | 胡婧怡. 辽河口北国海滨, 绿苇红滩[N]. 人民日报, 2023-02-21 (08). |
| [16] |
李延峰, 毛德华, 王宗明, 等. 双台河口国家级自然保护区芦苇叶面积指数遥感反演与空间格局分析[J]. 湿地科学, 2014, 12(2): 163-169. Li Y F, Mao D H, Wang Z M, et al. Remote sensing retrieval and spatial pattern analysis of leaf area index of phragmites australis in Shuangtai Estuary National Nature Reserve[J]. Wetland Science, 2014, 12(2): 163-169. |
| [17] |
齐玥, 孙永光, 马恭博, 等. 辽河口沉积物环境要素时空动态及影响因素分析[J]. 环境科学, 2020, 41(7): 3175-3185. Qi Y, Sun Y G, Ma G B, et al. Temporal and spatial variation patterns of the environmental elements in the sediments of the Liaohe Estuary and the related influencing factors[J]. Environmental Science, 2020, 41(7): 3175-3185. |
| [18] |
邢庆会, 上官魁星, 廖国祥, 等. 辽河口芦苇湿地净生态系统CO2交换及其环境调控[J]. 海洋环境科学, 2021, 40(2): 228-234. Xing Q H, Shangguan K X, Liao G X, et al. Net ecosystem CO2 exchange and its environmental regulation in the Liao river estuarine reed wetland[J]. Marine Environmental Science, 2021, 40(2): 228-234. |
| [19] | Johnston A S A, Meade A, Ardö J, et al. Temperature thresholds of ecosystem respiration at a global scale[J]. Nature Ecology & Evolution, 2021, 5(4): 487-494. |
| [20] |
马安娜, 陆健健. 长江口崇西湿地生态系统的二氧化碳交换及潮汐影响[J]. 环境科学研究, 2011, 24(7): 716-721. Ma A N, Lu J J. Net ecosystem exchange of carbon and tidal effects in Chongxi wetland, Yangtze Estuary[J]. Research of Environmental Sciences, 2011, 24(7): 716-721. |
| [21] | Zhong Q C, Wang K Y, Lai Q F, et al. Carbon dioxide fluxes and their environmental control in a reclaimed coastal wetland in the Yangtze Estuary[J]. Estuaries and Coasts, 2016, 39(2): 344-362. DOI:10.1007/s12237-015-9997-4 |
| [22] |
初小静, 韩广轩, 朱书玉, 等. 环境和生物因子对黄河三角洲滨海湿地净生态系统CO2交换的影响[J]. 应用生态学报, 2016, 27(7): 2091-2100. Chu X J, Han G X, Zhu S Y, et al. Effect of environmental and biotic factors on net ecosystem CO2 exchange over a coastal wetland in the Yellow River Delta[J]. Chinese Journal of Applied Ecology, 2016, 27(7): 2091-2100. |
| [23] |
邢庆会, 韩广轩, 于君宝, 等. 黄河口潮间盐沼湿地生长季净生态系统CO2交换特征及其影响因素[J]. 生态学报, 2014, 34(17): 4966-4979. Xing Q H, Han G X, Yu J B, et al. Net ecosystem CO2 exchange and its controlling factors during the growing season in an inter-tidal salt marsh in the Yellow River Estuary, China[J]. Acta Ecologica Sinica, 2014, 34(17): 4966-4979. |
| [24] |
杨利琼, 韩广轩, 于君宝, 等. 开垦对黄河三角洲湿地净生态系统CO2交换的影响[J]. 植物生态学报, 2013, 37(6): 503-516. Yang L Q, Han G X, Yu J B, et al. Effects of reclamation on net ecosystem CO2 exchange in wetland in the Yellow River Delta, China[J]. Chinese Journal of Plant Ecology, 2013, 37(6): 503-516. |
| [25] |
魏思羽. 黄河三角洲盐沼湿地碳交换过程及其对潮汐淹水的响应[D]. 烟台: 中国科学院大学(中国科学院烟台海岸带研究所), 2020. Wei S Y. Ecosystem carbon exchange and its response to tidal flooding in a salt marsh in the Yellow River Delta[D]. Yantai: University of Chinese Academy of Sciences (Yantai Institute of Coastal Zone Research, Chinses Academy of Sciences), 2020. |
| [26] |
付为国, 李萍萍, 卞新民, 等. 镇江北固山湿地芦苇光合日变化的研究[J]. 西北植物学报, 2006, 26(3): 496-501. Fu W G, Li P P, Pu X M, et al. Diurnal photosynthetic changes of phragmites communis in the wetland lying in Beigushan Mountain of Zhengjian Prefecture[J]. Acta Botanica Boreali-Occidentalia Sinica, 2006, 26(3): 496-501. DOI:10.3321/j.issn:1000-4025.2006.03.011 |
| [27] | Schäfer K V R, Tripathee R, Artigas F, et al. Carbon dioxide fluxes of an urban tidal marsh in the Hudson ‐Raritan estuary[J]. Journal of Geophysical Research: Biogeosciences, 2014, 119(11): 2065-2081. DOI:10.1002/2014JG002703 |
| [28] | Han G X, Yang L Q, Yu J B, et al. Environmental controls on net ecosystem CO2 exchange over a reed (Phragmites australis) wetland in the Yellow River Delta, China[J]. Estuaries and Coasts, 2013, 36(2): 401-413. DOI:10.1007/s12237-012-9572-1 |
| [29] | Buckley L B, Huey R B. Temperature extremes: geographic patterns, recent changes, and implications for organismal vulnerabilities[J]. Global Change Biology, 2016, 22(12): 3829-3842. DOI:10.1111/gcb.13313 |
| [30] |
王思琦, 李团胜, 付丛生, 等. 生态系统呼吸温度敏感性的分布及影响因素[J]. 环境科学与技术, 2021, 44(9): 167-173. Wang S Q, Li T S, Fu C S, et al. Distribution and influencing factors of the temperature sensitivity of ecosystem respiration[J]. Environmental Science & Technology, 2021, 44(9): 167-173. |
| [31] | Johnston A S A, Sibly R M. The influence of soil communities on the temperature sensitivity of soil respiration[J]. Nature Ecology & Evolution, 2018, 2(10): 1597-1602. |
| [32] | Green J K, Seneviratne S I, Berg A M, et al. Large influence of soil moisture on long-term terrestrial carbon uptake[J]. Nature, 2019, 565(7740): 476-479. DOI:10.1038/s41586-018-0848-x |
| [33] | Zhang Z Y, Zhang R D, Zhou Y, et al. A temperature threshold to identify the driving climate forces of the respiratory process in terrestrial ecosystems[J]. European Journal of Soil Biology, 2018, 89: 1-8. DOI:10.1016/j.ejsobi.2018.08.001 |
| [34] |
陈龙飞, 何志斌, 明姣, 等. 冻融作用对土壤呼吸影响的研究进展[J]. 干旱区研究, 2020, 37(4): 917-924. Chen L F, He Z B, Ming J, et al. Review of the effects of freezethaw processes on soil respiration[J]. Arid Zone Research, 2020, 37(4): 917-924. |
| [35] | Sorensen P O, Templer P H, Finzi A C. Contrasting effects of winter snowpack and soil frost on growing season microbial biomass and enzyme activity in two mixed-hardwood forests[J]. Biogeochemistry, 2016, 128(1-2): 141-154. DOI:10.1007/s10533-016-0199-3 |
| [36] |
谢艳兵, 贾庆宇, 周莉, 等. 盘锦湿地芦苇群落土壤呼吸作用动态及其影响因子分析[J]. 气象与环境学报, 2006, 22(4): 53-58. Xie Y B, Jia Q Y, Zhou L, et al. Soil respiration and its controlling factors at Phragmites communis wetland in Panjin[J]. Journal of Meteorology and Environment, 2006, 22(4): 53-58. |
| [37] | Hsu P K, Takahashi Y, Merilo E, et al. Raf-like kinases and receptor-like (pseudo)kinase GHR1 are required for stomatal vapor pressure difference response[J]. Proceedings of the National Academy of Sciences of the United States of America, 2021, 118(47). DOI:10.1073/PNAS.2107280118 |
| [38] | Goodrich J P, Campbell D I, Clearwater M J, et al. High vapor pressure deficit constrains GPP and the light response of NEE at a Southern Hemisphere bog[J]. Agricultural and Forest Meteorology, 2015, 203: 54-63. DOI:10.1016/j.agrformet.2015.01.001 |
| [39] |
叶子飘, 杨小龙, 康华靖. C3和C4植物光能利用效率和水分利用效率的比较研究[J]. 浙江农业学报, 2016, 28(11): 1867-1873. Ye Z P, Yang X L, Kang H J. Comparison of light-use and wateruse efficiency for C3 and C4 species[J]. Acta Agriculturae Zhejiangensis, 2016, 28(11): 1867-1873. DOI:10.3969/j.issn.1004-1524.2016.11.10 |
| [40] | 张桂芳, 丁在松. C3植物的C4光合途径[J]. 生物学通报, 2015, 50(3): 12-16. |
| [41] | Duman T, Schäfer K V R. Partitioning net ecosystem carbon exchange of native and invasive plant communities by vegetation cover in an urban tidal wetland in the New Jersey Meadowlands (USA)[J]. Ecological Engineering, 2018, 114: 16-24. DOI:10.1016/j.ecoleng.2017.08.031 |
| [42] | He G S, Wang K Y, Zhong Q C, et al. Agroforestry reclamations decreased the CO2 budget of a coastal wetland in the Yangtze estuary[J]. Agricultural and Forest Meteorology, 2021, 296. DOI:10.1016/j.agrformet.2020.108212 |
| [43] | Wang F M, Eagle M, Kroeger K D, et al. Plant biomass and rates of carbon dioxide uptake are enhanced by successful restoration of tidal connectivity in salt marshes[J]. Science of the Total Environment, 2021, 750. DOI:10.1016/j.scitotenv.2020.141566 |
| [44] | Zhou L, Zhou G S, Jia Q Y. Annual cycle of CO2 exchange over a reed (Phragmites australis) wetland in Northeast China[J]. Aquatic Botany, 2009, 91(2): 91-98. DOI:10.1016/j.aquabot.2009.03.002 |
| [45] | Falge E, Baldocchi D, Tenhunen J, et al. Seasonality of ecosystem respiration and gross primary production as derived from FLUXNET measurements[J]. Agricultural and Forest Meteorology, 2002, 113(1-4): 53-74. |