 2023, Vol. 44
2023, Vol. 44

 , HUA Gui-li , LI Xiang-yu , FENG Liu-xu , SUI Kang-xin , GENG Li-ping , XUE Pei-ying , LIU Wen-ju
, HUA Gui-li , LI Xiang-yu , FENG Liu-xu , SUI Kang-xin , GENG Li-ping , XUE Pei-ying , LIU Wen-ju
近年来, 随着我国工业化、城市化和农业的快速发展, 农田土壤重金属污染已成为国家重点关注的环境问题之一[1, 2]. 2014年《全国土壤污染状况调查公报》显示, 我国耕地土壤点位超标率为19.4%, 重金属为主要一类污染物, 尤其是污水灌溉区超标点位占26.4%, 主要重金属污染物为镉(Cd)和砷(As), 农田Cd和As污染已被列为重点研究对象[3].土壤中累积的Cd和As可被农作物根系吸收并转移至可食部位, 通过食物链威胁人体健康[4~7].华北平原是我国重要的农产品基地之一, 但是长期的污水灌溉、冶炼、农药和化肥的盲目施用等人为活动使土壤中重金属累积, 造成农田土壤重金属污染[8, 9], 且有研究表明土壤中的类金属As和重金属Cd易向农作物体内转移[10~12].小麦是我国第二大主粮作物, 品种繁多且种植面积广, 小麦清洁生产对保障国家粮食安全至关重要[13, 14].然而, 已有的研究发现小麦籽粒对土壤重金属Cd的积累能力较强, 并可通过食物链对人体健康构成威胁[15~17], 因此, 非常有必要采取有效措施来减少小麦根系对土壤中镉的吸收及其向籽粒中转运[18~20].一般而言, 土壤中Cd和As主要通过两个途径向籽粒中转运, 其中木质部介导的Cd和As从根系直接向地上部转运是决定籽粒中Cd和As累积的关键过程之一, 即土壤溶液中的Cd和As经根系吸收后进入小麦体内, 通过质外体或共质体途径横向运输至木质部, 进而直接经木质部向小麦的地上部转移并积累于籽粒中; 同时, 韧皮部运输是决定小麦籽粒Cd和As运输的另一个重要途径, Cd和As经植物根系吸收转运至地上部后, 其在茎节处由木质部转移至韧皮部, 进而由韧皮部重新分配和向上运输, 最终进入籽粒[21, 22].小麦开花后进入灌浆期, 这是小麦籽粒形成的关键生育期[23], 在该时期Cd和As主要通过木质部还是韧皮部的途径向籽粒转运, 是揭示体内Cd和As转运控制籽粒中Cd和As富集的关键所在.此外, Cd由根系向地上部运输并最终在小麦籽粒中积累, 其由节向穗部和穗轴向籽粒的转运过程在调控小麦籽粒Cd积累及其品种差异方面发挥着重要作用[24].然而, 对于在镉砷复合污染区种植的小麦而言, 究竟地上部的哪个或哪些部位在开花后进入灌浆期-成熟期控制着小麦籽粒Cd和As累积, 以及开花后Cd和As在小麦不同部位的迁移特征仍不清晰.基于此, 本研究拟在镉砷复合污染的小麦种植区, 采用田间小区试验, 探索两个小麦品种SN28和JM22在灌浆前期、灌浆后期和成熟期, 地上不同部位Cd和As的分布及迁移特征, 以期为研发阻控镉砷进入小麦可食部位的关键技术和保障粮食安全提供理论和技术支撑.
1 材料与方法 1.1 研究区概况研究区域位于华北平原区某一因污灌和冶炼造成的镉砷复合污染农田, 当地以冬小麦-夏玉米一年两熟为主要种植制度.该区域是受周边有色金属冶炼和污灌共同影响的典型农田, 有色金属铜冶炼厂生产期间废水排入旁边的河道, 对地表水体和地下水造成重金属镉砷污染, 污染物随灌溉水进入农田[25~27]; 此外, 冶炼厂废气中的颗粒物负载的重金属也会随干湿沉降进入农田, 本试验开展时期该冶炼厂已关停6 a.土壤类型为中壤质潮土, 其表层土壤的基本理化性质如下:pH 7.64, ω(碱解氮)87.8 mg·kg-1, ω(有效磷)31.4 mg·kg-1, ω(速效钾)341 mg·kg-1, ω(有机质)21.1 g·kg-1, CEC 23.1 cmol·kg-1, ω(Cd)2.49 mg·kg-1, ω(As)106 mg·kg-1.
1.2 供试材料供试品种:小麦品种为山农28(SN28)和济麦22(JM22), 其中济麦22为笔者课题组筛选出的籽粒镉砷低积累的品种[28].
1.3 试验设计本试验于2020年10月24日至2021年6月20日在镉砷复合污染小麦种植区进行.在种植两个小麦品种JM22和SN28的试验区(每个品种种植面积为5.00 hm2), 分别随机划出3个小区(每个小区面积为10 m×10 m=100 m2)作为3个重复.施肥、灌水和病虫害防治均按照当地农民的管理方式进行.整个生育期小麦的灌溉方式为喷灌, 每次灌水量为555 m3·hm-2, 分别在小麦的越冬期、返青期、拔节期、抽穗开花期和灌浆期进行.
1.4 样品采集与测定 1.4.1 植物样品的采集小麦开花后分别于灌浆前期(5月23日)、灌浆后期(5月30日)以及成熟期(6月19日)按照S形在每个小区采集植株样品, 为了避免道路扬尘的影响, 选择采集样品区域为远离道路40 m的区域, 每个小区采集3个样点, 每个样点采集5~10株小麦.将整株小麦样品用不锈钢铁锹挖出后, 将土壤轻轻抖落后放入自封袋, 尽快运回实验室.将植株分为地上部、根部分别用自来水、蒸馏水和超纯水洗净, 使用吸水纸拭干表面水分.然后将样品小麦地上部分为穗(spike)、倒一叶、倒二叶和倒三叶及其相关部位, 其中穗包括穗轴(rachis, RH)、颖壳(glume, GM)和籽粒(grain, GA); 倒一叶(旗叶)及其相关部位包括叶片(leaf 1, L1)、叶鞘1(sheath 1, SH1)、节1(node 1, N1)和倒一叶到穗之间的节间1(internode 1, IN1), 依次往下分为倒二叶及其相关部位, 倒三叶及其相关部位(图 1)[29].在烘箱中85℃杀青15 min, 然后于65℃烘干至恒重[25].各个部位的样品用研钵或不锈钢微型粉碎机粉碎后, 用于植株各部位中Cd和As含量的测定.

|
图 1 小麦地上不同部位示意 Fig. 1 Different positions of wheat shoot |
植株样品均采用HNO3-H2O2(5∶2, 体积比)消解, 消解后使用电感耦合等离子体质谱仪(ICP-MS)(PerkinElmer, NexION350X, USA)测定植株不同部位Cd和As的含量[30, 31].测定小麦籽粒中Cd和As含量, 以国家一级标准物质(GBW 10011, 小麦粉)进行准确度和精密度控制, 并同步分析空白样品以去除试剂干扰, 其中Cd回收率为95%~100%, As回收率为90%~95%, 均符合质量控制要求.同时测定小麦植株地上部各部位Cd和As含量, 以国家一级标准绿茶粉(GBW 10052a)进行准确度和精密度控制, Cd回收率为90%~95%, As回收率为90%~100%.
1.5 数据统计分析 1.5.1 转运系数小麦中重金属Cd和As的转运系数按照如下的公式进行计算:
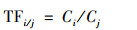
|
式中, TFi/j表示重金属由小麦j部位向i部位的转移系数, Ci为小麦i部位重金属含量(mg·kg-1), Cj为小麦j部位重金属含量(mg·kg-1).
1.5.2 数据统计方法采用Microsoft Excel 2016制作图表, 使用SPSS 18.0进行数据分析, 采用单因素方差分析法(one-way ANOVA)进行显著性检验, 采用最小显著差异法(LSD)进行小麦不同部位之间重金属含量和转运系数的差异显著性比较.小麦籽粒与叶片和节重金属含量之间采用Pearson进行相关性分析, 独立样本T检验方法进行两个品种间Cd或As含量差异的显著性比较.
2 结果与分析 2.1 开花后不同生育时期小麦中Cd和As的分布特征 2.1.1 Cd的分布特征在小麦开花后采集灌浆前期、灌浆后期和成熟期的小麦样品, Cd在小麦地上部各部位的含量分布情况如图 2所示.对JM22而言, 随着小麦叶片位置的降低, 3个生育期叶片中Cd含量呈逐渐上升的趋势, 倒三叶中Cd含量最高且显著高于倒二叶和旗叶, 平均增幅为63.4%.同时, 在灌浆前期和成熟期, JM22的叶鞘3的ω(Cd)平均值为0.895 mg·kg-1, 显著高于其他位置的叶鞘中Cd含量, 平均增幅为56.5% [图 2(a)和2(c)]; 小麦品种SN28不同部位叶片和叶鞘Cd含量也呈现与JM22相似的趋势, 但是仅在灌浆前期达到显著水平[图 2(a)], 这是由于灌浆前期小麦根系吸收的Cd优先向其基部叶片和叶鞘中转运所致.此外, 成熟期SN28和JM22叶片Cd含量相对于灌浆前期显著增加, 增幅分别为343%和141%, 并且JM22旗叶及其叶鞘中Cd含量显著低于SN28 [图 2(c)].

|
(a)、(b)和(c) 分别为小麦灌浆前期、灌浆后期和成熟期不同部位Cd含量; 不同大写或小写字母分别表示SN28或JM22不同部位Cd含量存在显著性差异(P < 0.05); *表示同一部位不同小麦品种间Cd含量差异显著(P < 0.05) 图 2 小麦开花后不同时期两个品种小麦不同部位Cd含量 Fig. 2 Cd concentrations in different parts of wheat shoot for two cultivars at different growing stages after flowering |
节是植物体内Cd转运的关键部位, 一方面Cd被根系吸收后经节和节间向叶片转运, 而叶片中累积的Cd再分配进入韧皮部经节向穗部迁移, 另一方面, Cd被根系吸收后横向运输至木质部, 通过木质部经节和节间直接转移至穗部.结果表明, 灌浆前期和后期小麦节1中Cd含量显著高于其他部位[图 2(a)和2(b)], 其中Cd含量存在节>节间的趋势.对JM22而言, 在灌浆前期、灌浆后期和成熟期, 节和节间中Cd含量随着位置降低呈逐渐下降的趋势, 其中旗叶对应的节1与节间1中Cd含量最高, ω(Cd)平均值分别为1.76 mg·kg-1和0.56 mg·kg-1; 在灌浆前期和后期, 节1和节间1中Cd含量分别显著高于节3和节间3, 平均增幅分别为179%和79.0%; 对于SN28而言, 只有灌浆前期和后期节1中ω(Cd)平均值为3.02 mg·kg-1, 显著高于节3达180% [图 2(a)和2(b)].此外, 随着小麦植株生长发育阶段的延长, 节中Cd含量逐渐下降, 成熟期SN28和JM22节中Cd含量比灌浆期分别降低53.0%和57.7% [图 2(c)].穗中Cd含量在灌浆前期和后期显著低于其他部位, 在成熟期SN28穗中Cd含量显著高于JM22, 增幅为29.7%.
综上所述, 小麦旗叶、旗叶的叶鞘及节1(穗下第1个节)对调控Cd在籽粒中的累积起了重要的作用.
2.1.2 As的分布特征As在小麦地上部各部位的含量分布情况如图 3所示, 比较灌浆前期、灌浆后期和成熟期小麦各部位的As含量发现, 两个品种小麦叶片中As含量显著高于其他部位.对于JM22而言, 在灌浆前期和后期随着叶片位置降低, 其As含量均呈逐渐上升趋势, 叶3中As含量显著高于旗叶, 增幅达154% [图 3(a)和3(b)], 叶片中As含量大小表现为:灌浆前期>灌浆后期>成熟期; 此外, 叶鞘中As含量显著低于叶片, 平均含量仅为叶片的20.2%.对于SN28而言, 不同部位叶片As含量也呈现与JM22相似的趋势.仅在灌浆前期叶片3和叶鞘3中As含量均表现为JM22显著高于SN28, 增幅分别为168%和53.3% [图 3(a)].
进一步分析节和节间中As含量发现, JM22在灌浆前期节中As含量随着位置降低呈现着逐渐上升的趋势[图 3(a)], 节3中ω(As)为6.34 mg·kg-1, 显著高于其他位置的节, 平均增幅为81.7%; 同时, 在灌浆期每个节中As含量显著高于对应的节间.成熟期不同部位的节和节间中As含量均没有显著变化; 对于SN28而言, 在小麦开花后节中As含量随着位置的降低则呈现着逐渐下降的趋势, 节1中As含量最高, 在灌浆后期和成熟期显著高于节2和节3, 平均增幅分别为189%和240%.此外, 在3个时期, SN28节1中As含量均显著高于JM22.综上所述, 小麦叶3、叶鞘3和节1主要参与了As在籽粒的累积.

|
(a)、(b)和(c)分别为小麦灌浆前期、灌浆后期和成熟期不同部位As含量; 不同大写或小写字母分别表示SN28或JM22不同部位As含量存在显著性差异(P < 0.05); *表示同一部位不同小麦品种间As含量差异显著(P < 0.05) 图 3 小麦开花后不同时期两个品种小麦不同部位As含量 Fig. 3 Arsenic concentrations in different parts of wheat shoot for two cultivars at different growing stages after flowering |
在成熟期分析了小麦穗的穗轴、颖壳和籽粒中Cd和As含量(图 4).穗轴是与籽粒直接相连的部位, 也是Cd和As向籽粒运输的主要途径之一; 颖壳与小麦籽粒也直接相连, 对籽粒中Cd和As累积具有一定的贡献.从穗子3个部位的Cd含量来看[图 4(a)], JM22和SN28籽粒中ω(Cd)分别为0.20 mg·kg-1和0.36 mg·kg-1, 前者比后者显著降低了44.4%; 颖壳ω(Cd)分别为0.74 mg·kg-1和1.25 mg·kg-1, 前者比后者显著降低了40.8%; 穗轴Cd含量分别为1.39 mg·kg-1和1.79 mg·kg-1, 前者比后者显著降低了22.3%; 从穗子3个部位As含量来看[图 4(b)], JM22和SN28籽粒ω(As)分别为0.26 mg·kg-1和0.38 mg·kg-1, JM22籽粒As含量相对于SN28显著降低了33.3%; 进一步分析发现, 小麦颖壳与籽粒呈现相似的As累积趋势, 其中SN28和JM22颖壳ω(As)分别为3.83 mg·kg-1和2.84 mg·kg-1, 后者比前者显著降低了25.8%; 两个品种穗轴中As含量没有显著差异.穗子不同部位中Cd和As含量的变化趋势为:穗轴>颖壳>籽粒, 且具有显著性差异, 因此推断穗轴是调控穗部Cd和As向籽粒中累积的重要部位; 此外, SN28颖壳和籽粒中Cd和As含量显著高于JM22.

|
(a)和(b)分别为小麦穗不同部位Cd和As含量; 不同大写字母或小写字母分别表示SN28或JM22不同部位Cd或As含量存在显著性差异(P < 0.05); *表示同一部位不同小麦品种间Cd或As含量差异显著(P < 0.05) 图 4 小麦两个品种穗的不同部位Cd和As含量 Fig. 4 Concentrations of Cd and As in different parts of wheat spikes for two cultivars |
转运系数(TF)表示重金属在植物不同部位之间的转运能力, 不同时期小麦不同部位之间Cd转运系数如图 5所示.开花后Cd由下部叶片和节向穗部转运, 主要包括:Cd从下部叶位的节经木质部进入相应的叶鞘和叶片, 并在叶片中累积, 叶片中的Cd可经韧皮部随同化产物的小分子一起运输至叶鞘和节, 节中的Cd可继续通过木质部或韧皮部运至该位置的节间和上一个叶位的节, 然后进行下一轮的运转, 最终进入穗部.本研究发现, 在灌浆期和成熟期两个品种小麦中的Cd经不同位置叶鞘进入叶片的能力较强, 显著高于Cd由节到叶鞘的转运, 这说明根系吸收的Cd主要通过节和节间的维管束系统进入叶片中累积; 从叶片中Cd再分配的外运结果来看, 在灌浆前期和灌浆后期, Cd由小麦叶鞘到节的转运系数显著高于叶到叶鞘和节到节间, 平均增幅分别为326%和849%, 尤其是接近穗部的叶鞘1到节1的Cd转运能力最强, 比倒二叶和倒三叶的叶鞘到节转运系数分别显著提高104%和470% [图 5(a)和5(b)]; 此外, Cd在节与节间之间转运的主要特征为:下部叶位的节间到上一个叶位的节Cd转运系数(IN3-N2和IN2-N1)显著高于同一叶位的节向节间的Cd转运系数(N2-IN2和N1-IN1), 这说明Cd由节间向节的转运能力显著强于由节向外的转运能力; 同时, 比较两个小麦品种对Cd的转运能力发现, 在灌浆前期和灌浆后期, 尽管JM22中Cd由节2到节间2的转运系数显著高于SN28, 但是Cd由节间2到节1和叶鞘到节的转运系数JM22远低于SN28[图 5(a)和5(b)].综上分析, 在灌浆期Cd由叶鞘到节与节间到节的高转运和节到叶鞘与节到节间的低转运是调控籽粒中Cd累积的关键步骤, 可见节是Cd向籽粒中转运的重要“中转站”, 尤其是节1. 从成熟期各部位之间Cd的转运来看, SN28穗轴到颖壳和穗轴到籽粒的Cd转运系数显著高于JM22, 说明穗轴在Cd进入籽粒的过程中起着重要作用.

|
(a)、(b)和(c)分别为灌浆前期、灌浆后期和成熟期小麦不同部位之间Cd转运系数; 不同大写字母或小写字母分别表示SN28或JM22不同部位之间Cd存在显著性差异(P < 0.05); *表示相同部位不同小麦品种间Cd转运系数差异显著(P < 0.05) 图 5 开花后不同生育期在两个小麦品种不同部位之间Cd的转运系数 Fig. 5 Translocation factors of Cd in different parts of wheat shoots for two cultivars at different growing stages after flowering |
As在开花后不同生育时期各个部位之间的转运系数如图 6所示, 其在小麦体内的运输途径与Cd类似, 且在灌浆期和成熟期两个品种小麦中As经不同位置叶鞘进入叶片的能力显著高于节到叶鞘的转运; 在灌浆前期、灌浆后期和成熟期, As由小麦叶鞘到节的转运系数显著高于叶到叶鞘和节到节间的转运系数, 平均增幅为438%和190%; 此外, 比较两个小麦品种As转运的差异发现, SN28叶鞘向节的As转运系数显著高于JM22, 说明在小麦开花后As主要通过叶鞘向节中运输并积累于节中; 然而, 在这3个时期As由节1到节间1的转运能力JM22显著强于SN28, 而后节间1中的As继续向穗轴转运, 基于开花后两个品种体内As由叶鞘1到节1和节1到节间1转运能力存在相反的差异, 导致两个品种穗轴中As含量差异不显著; 同时, SN28的节间2到节1的As转运系数显著高于JM22, 造成SN28节1中As含量显著高于JM22, 说明节1是负责As向籽粒运输的一个关键部位.进一步分析小麦穗部各器官之间的转运系数发现, As由穗轴到颖壳、颖壳到籽粒和穗轴直接到籽粒的转运系数均表现为SN28显著高于JM22, 说明穗轴和颖壳在As的转运中起着重要作用, 尤其是JM22穗轴到籽粒的As转运系数比SN28显著降低了44.4%, 而颖壳到籽粒As转运系数则比SN28显著降低了40.6%.

|
(a)、(b)和(c)分别为灌浆前期、灌浆后期和成熟期小麦不同部位之间As转运系数; 不同大写字母或小写字母分别表示SN28或JM22不同部位之间As存在显著性差异(P < 0.05); * 表示相同部位之间不同小麦品种As转运系数差异(P < 0.05) 图 6 开花后不同生育期As在两个小麦品种不同部位之间的转运系数 Fig. 6 Translocation factors of As in different parts of two wheat cultivars at different growing stages after flowering |
进一步分析成熟期两个品种小麦的节和叶片分别与籽粒之间Cd含量的相关性发现, 小麦节与籽粒的Cd含量之间、叶片与籽粒的Cd含量之间均呈显著正相关关系(P < 0.01, n=18)[图 7(a)和7(b)], 这说明籽粒对Cd累积能力的高低与叶片和节中Cd含量密不可分; 同时分析小麦节和叶片与籽粒之间As含量相关性, 发现小麦节与籽粒之间As含量呈现显著正相关关系(P < 0.01, n=18)[图 7(c)], 然而小麦叶片与籽粒之间As含量有负相关的趋势, 但相关性不显著[图 7(d)].这说明As易在叶片中累积的品种, As向籽粒中转运较少, 而节则对籽粒中As累积起着重要作用.因此, 进一步验证了小麦节是Cd和As向籽粒转运的重要部位.

|
图 7 小麦籽粒与节和叶片中镉砷含量之间的相关关系 Fig. 7 Relationships between Cd or As concentrations in grain and node or in grain and leaf |
已有研究表明水稻中Cd的韧皮部运输是决定稻米中Cd积累的重要环节, Cd在茎节处可由木质部转移至韧皮部进行自下而上或自上而下的转运, 叶片中累积的Cd, 尤其是旗叶中的Cd可通过韧皮部重新分配, 经叶鞘、节1、穗下节间进入穗轴乃至颖壳, 最终进入水稻籽粒.因此, 水稻中节是木质部中Cd向韧皮部转运的重要部位, 在Cd向上转运至籽粒起着重要作用[32, 33].本研究发现, 与水稻同为禾本科单子叶植物的小麦, 开花后地上部积累的Cd和As通过节、节间和叶片等器官的再分配后进入籽粒, 随着在小麦植株地上部位置的升高, 叶片和叶鞘中Cd和As含量呈现逐渐下降, 而其在节和节间呈现逐渐升高的趋势, 这表明土壤中的Cd和As被小麦根部吸收后更易于向基部的叶片和叶鞘以及顶部的节和节间输送.
灌浆期是小麦籽粒形成的关键生育时期, 叶片中合成的光合产物通过韧皮部运输到籽粒, 此过程同时伴随着营养元素以及重金属的迁移, 最后进入籽粒积累[34, 35].通过分析相关研究发现, 成熟期小麦中Cd由节间向相邻节转运系数显著高于节向相邻节间的转运系数[36], 而本研究结果进一步表明在小麦灌浆前期、灌浆后期和成熟期这3个生育期, Cd由节间运输至上一个叶位节的转运能力显著高于节向其同叶位节间的转运能力[图 5(a)和5(b)], 但是成熟期仅在Cd低积累品种JM22中呈现出相似的Cd转运特征, 这说明灌浆期旗叶的生命力及其转运光合产物的能力较强, 可以将叶片中的Cd与光合产物一起通过节转运至穗部.同时, 节1中Cd含量达到最高, 进一步说明节是限制Cd向小麦穗部转运的主要部位, 尤其是节1.这可能是和节中转运蛋白的显著表达有关, 相关研究表明水稻中OsZIP7转运体在茎节处参与调控Cd从木质部向韧皮部运输[37].已有的研究表明Cd在水稻地上部的转运受到节的显著控制, 尤其是穗下的第一节, 这一点与本研究小麦的结果一致[38, 39].研究发现节1的扩散维管束(DVB)和转运维管束(TVB)的面积最小[29], 放大维管束(EVB)与叶片的木质部相连, 如果节DVB太小则会导致节中更多的Cd通过EVB转移到叶片中[40].由此可见, 不同部位的节调控Cd运输的能力存在差异, 可能是由于不同部位节中DVB和TVB面积大小不同所致, 较大的维管束面积为Cd的运输提供更大的通道, 导致更多的Cd向小麦籽粒中累积, 反之亦然.本研究中成熟期小麦叶片Cd含量高于其他部位, 可能是由于SN28和JM22两个品种节中DVB的面积较小, 导致根部吸收的Cd向上运输的过程中一部分积累于节, 其中也有一部分Cd经节流向叶片.通过对比两个品种发现Cd低累积品种JM22叶片Cd含量高于SN28, 因此推测可能是由于更多的Cd转运至叶片并累积下来, 导致小麦籽粒中Cd含量下降所致.同位素示踪试验发现, 小麦籽粒中40%~45%的Cd来源于叶片中Cd的活化再利用[41].通过分析两品种小麦不同部位之间的转运系数, 发现在灌浆前期和灌浆后期Cd由叶鞘到节的转运能力显著强于叶到叶鞘和节到节间的转运, 并且SN28叶到叶鞘和叶鞘到节的转运系数显著高于JM22, 这表明在灌浆期小麦叶片向叶鞘再到节这一途径是Cd向穗部运输的主要途径, 叶鞘到节的高转运与节到节间的低转运影响着Cd向小麦籽粒中运输.进一步分析小麦穗各部位之间的Cd转运, 虽然JM22中Cd由节间到穗轴的转运系数显著高于SN28, 然而其从穗轴到颖壳以及穗轴直接到籽粒的转运规律正好相反, 前人的研究也发现, 相对于高累积品种来说灌浆期低累积品种Cd由节间1到穗轴的转运能力更强[42].这说明穗轴对Cd向籽粒运输起到了重要的抑制作用; 刘畅等[36]也发现低积累品种籽粒、颖壳和穗轴的Cd富集能力显著低于高累积品种, 且相关的转运也显著低于高累积品种, 表明穗部累积少且向籽粒转运少是低累积品种籽粒Cd含量低的重要原因.但是, 冯亚娟等[43]对小麦各器官Cd积累分布规律研究发现, 低累积品种的颖壳对Cd具有很好的滞留作用, 低累积品种颖壳中Cd含量显著高于高累积品种, 这说明对于该试验的小麦品种而言, 颖壳调控着籽粒中Cd的累积.
不同品种之间籽粒和叶片As的积累能力存在很大的差异, 有研究发现在As污染环境下种植水稻, 低累积品种叶片中As含量显著高于高累积品种, 这表明As低累积品种会优先将As转移至叶片当中, 进而影响籽粒中As的累积[44], 这一点与本研究的结果相似, JM22叶片中As含量在灌浆期高于SN28, 但是成熟期籽粒中As含量却显著低于SN28.此外, 土壤施肥可有效降低水稻籽粒和秸秆中As含量[45], 且在灌浆期向小麦叶片喷施硅能够有效降低小麦籽粒中的As含量[46], 这也表明叶片是控制As向籽粒转运的关键部位之一.本研究发现在灌浆前期、灌浆后期和成熟期小麦叶片As含量显著高于其他部位, 是As累积的主要部位.同时, 灌浆前期JM22倒三叶中As含量高于SN28, 且前者As由叶片向叶鞘的转运能力显著低于后者, 因此, JM22籽粒中As较低是由于As在旗叶中的滞留作用较强所致.有研究通过对比水稻中不同部位的As含量发现, 倒三叶的As含量显著高于其他部位叶片As含量[47], 这一点与本研究结果一致.然而节与之呈现相反的变化规律, 且节的As含量显著高于对应的节间, 同时开花后3个时期高累积品种SN28节1中As含量, 以及As由节间2向节1的转运能力均显著高于JM22, 这表明节1是抑制As向上运输的关键位置; 此外, 颖壳也是阻控As向籽粒迁移的重要部位, 这与颖壳对Cd的滞留作用相似, 总之, 旗叶、节1和颖壳综合阻控了As向小麦籽粒的转运.
4 结论(1) 开花后小麦中Cd主要分布在节, 且节1含量最高, 旗叶以及其叶鞘和节是控制小麦籽粒Cd累积的关键部位; 而小麦中As主要分布于叶片, 且叶片3含量最高, 小麦叶片3、叶鞘3以及节1对As在小麦籽粒累积起了重要的作用.
(2) 节是控制Cd、As向各部位转运的重要枢纽; 旗叶的叶鞘到节1、穗轴向颖壳、穗轴向籽粒的转运过程是限制Cd向小麦籽粒转运的关键过程; 旗叶和节1是负责Cd向籽粒转运的关键部位, 穗轴向籽粒的转运主要控制As向小麦籽粒中的转运.
| [1] | Zhao F J, Tang Z, Song J J, et al. Toxic metals and metalloids: Uptake, transport, detoxification, phytoremediation, and crop improvement for safer food[J]. Molecular Plant, 2022, 15(1): 27-44. DOI:10.1016/j.molp.2021.09.016 |
| [2] |
骆永明, 滕应. 中国土壤污染与修复科技研究进展和展望[J]. 土壤学报, 2020, 57(5): 1137-1142. Luo Y M, Teng Y. Research progresses and prospects on soil pollution and remediation in China[J]. Acta Pedologica Sinica, 2020, 57(5): 1137-1142. |
| [3] |
徐长春, 郑戈, 林友华. "十三五"国家重点研发计划农田镉砷污染防治领域资助情况概述[J]. 农业环境科学学报, 2018, 37(7): 1321-1325. Xu C C, Zheng G, Lin Y H. Brief introduction to research projects on prevention and control of cadmium and arsenic pollution in croplands supported by National Key R&D Program of China in 13th Five-Year Period[J]. Journal of Agro-Environment Science, 2018, 37(7): 1321-1325. |
| [4] | Rezapour S, Atashpaz B, Moghaddam S S, et al. Heavy metal bioavailability and accumulation in winter wheat(Triticum aestivum L.) irrigated with treated wastewater in calcareous soils[J]. Science of the Total Environment, 2019, 656: 261-269. DOI:10.1016/j.scitotenv.2018.11.288 |
| [5] | Sui F Q, Zhao D K, Zhu H T, et al. Map-based cloning of a new total loss-of-function allele of OsHMA3 causes high cadmium accumulation in rice grain[J]. Journal of Experimental Botany, 2019, 70(10): 2857-2871. DOI:10.1093/jxb/erz093 |
| [6] | Wang P, Chen H P, Kopittke P M, et al. Cadmium contamination in agricultural soils of China and the impact on food safety[J]. Environmental Pollution, 2019, 249: 1038-1048. DOI:10.1016/j.envpol.2019.03.063 |
| [7] | Liu N, Huang X M, Sun L M, et al. Screening stably low cadmium and moderately high micronutrients wheat cultivars under three different agricultural environments of China[J]. Chemosphere, 2020, 241. DOI:10.1016/j.chemosphere.2019.125065 |
| [8] | Wei L, Ding Q, Guo H M, et al. Forms and mobility of heavy metals/metalloids in sewage-irrigated soils in the North China Plain[J]. Journal of Soils and Sediments, 2021, 21(1): 215-234. DOI:10.1007/s11368-020-02744-7 |
| [9] | Ma S J, Hu Y H, Zeng Q H, et al. Temporal changes of calcareous soil properties and their effects on cadmium uptake by wheat under wastewater irrigation for over 50 years[J]. Chemosphere, 2021, 263. DOI:10.1016/j.chemosphere.2020.127971 |
| [10] | Clemens S. Safer food through plant science: reducing toxic element accumulation in crops[J]. Journal of Experimental Botany, 2019, 70(20): 5537-5557. DOI:10.1093/jxb/erz366 |
| [11] | Zhao F J, Wang P. Arsenic and cadmium accumulation in rice and mitigation strategies[J]. Plant and Soil, 2020, 446(1-2): 1-21. DOI:10.1007/s11104-019-04374-6 |
| [12] | Du B Y, Zhou J, Lu B X, et al. Environmental and human health risks from cadmium exposure near an active lead-zinc mine and a copper smelter, China[J]. Science of the Total Environment, 2020, 720. DOI:10.1016/j.scitotenv.2020.137585 |
| [13] | Liu W T, Liang L C, Zhang X, et al. Cultivar variations in cadmium and lead accumulation and distribution among 30 wheat(Triticum aestivum L.) cultivars[J]. Environmental Science and Pollution Research, 2015, 22(11): 8432-8441. DOI:10.1007/s11356-014-4017-y |
| [14] | Sui F Q, Chang J D, Tang Z, et al. Nramp5 expression and functionality likely explain higher cadmium uptake in rice than in wheat and maize[J]. Plant and Soil, 2018, 433(1-2): 377-389. DOI:10.1007/s11104-018-3849-5 |
| [15] | Ma J F, Shen R F, Shao J F. Transport of cadmium from soil to grain in cereal crops: a review[J]. Pedosphere, 2021, 31(1): 3-10. DOI:10.1016/S1002-0160(20)60015-7 |
| [16] | Li X F, Zhou D M. A meta-analysis on phenotypic variation in cadmium accumulation of rice and wheat: implications for food cadmium risk control[J]. Pedosphere, 2019, 29(5): 545-553. DOI:10.1016/S1002-0160(19)60828-3 |
| [17] | Zhuang Z, Niño-Savala A G, Mi Z D, et al. Cadmium accumulation in wheat and maize grains from China: interaction of soil properties, novel enrichment models and soil thresholds[J]. Environmental Pollution, 2021, 275. DOI:10.1016/j.envpol.2021.116623 |
| [18] |
孙丽娟, 秦秦, 宋科, 等. 镉污染农田土壤修复技术及安全利用方法研究进展[J]. 生态环境学报, 2018, 27(7): 1377-1386. Sun L J, Qin Q, Song K, et al. The remediation and safety utilization techniques for Cd contaminated farmland soil: a review[J]. Ecology and Environmental Sciences, 2018, 27(7): 1377-1386. |
| [19] | Wu J W, Mock H P, Giehl R F H, et al. Silicon decreases cadmium concentrations by modulating root endodermal suberin development in wheat plants[J]. Journal of Hazardous Materials, 2019, 364: 581-590. DOI:10.1016/j.jhazmat.2018.10.052 |
| [20] | Chen H P, Zhang W W, Yang X P, et al. Effective methods to reduce cadmium accumulation in rice grain[J]. Chemosphere, 2018, 207: 699-707. DOI:10.1016/j.chemosphere.2018.05.143 |
| [21] | Liu Y K, Lu M, Tao Q, et al. A comparative study of root cadmium radial transport in seedlings of two wheat(Triticum aestivum L.) genotypes differing in grain cadmium accumulation[J]. Environmental Pollution, 2020, 266. DOI:10.1016/j.envpol.2020.115235 |
| [22] | Liu C X, Guttieri M J, Waters B M, et al. Cadmium concentration in terminal tissues as tools to select low-cadmium wheat[J]. Plant and Soil, 2018, 430(1-2): 127-138. DOI:10.1007/s11104-018-3712-8 |
| [23] |
潘建清, 陆敏, 杨肖娥. 不同小麦品种灌浆期生长和镉积累的差异研究[J]. 农业环境科学学报, 2021, 40(4): 756-765. Pan J Q, Lu M, Yang X E. Variations in growth and cadmium accumulation among 30 wheat(Triticum aestivum L.) cultivars during the grain filling stage[J]. Journal of Agro-Environment Science, 2021, 40(4): 756-765. |
| [24] | Qiao K, Wang F H, Liang S, et al. New biofortification tool: wheat TaCNR5 enhances zinc and manganese tolerance and increases zinc and manganese accumulation in rice grains[J]. Journal of Agricultural and Food Chemistry, 2019, 67(35): 9877-9884. DOI:10.1021/acs.jafc.9b04210 |
| [25] |
肖冰, 薛培英, 韦亮, 等. 基于田块尺度的农田土壤和小麦籽粒镉砷铅污染特征及健康风险评价[J]. 环境科学, 2020, 41(6): 2869-2877. Xiao B, Xue P Y, Wei L, et al. Characteristics of Cd, As, and Pb in soil and wheat grains and health risk assessment of grain-Cd/As/Pb on the field scale[J]. Environmental Science, 2020, 41(6): 2869-2877. |
| [26] |
张素荣, 王昌宇, 刘继红, 等. 雄安新区西南部土壤重金属污染特征及生态风险评价[J]. 地学前缘, 2021, 28(4): 238-249. Zhang S R, Wang C Y, Liu J H, et al. Assessments of heavy metal pollution in soils of the southwestern Xiong'an District and its ecological risk[J]. Earth Science Frontiers, 2021, 28(4): 238-249. |
| [27] | Zhang X Y, Geng L P, Gao P P, et al. Bioimaging of Pb by LA-ICP-MS and Pb isotopic compositions reveal distributions and origins of Pb in wheat grain[J]. Science of the Total Environment, 2022, 802. DOI:10.1016/j.scitotenv.2021.149729 |
| [28] |
孙洪欣, 薛培英, 赵全利, 等. 镉、铅积累与转运在冬小麦品种间的差异[J]. 麦类作物学报, 2015, 35(8): 1161-1167. Sun H X, Xue P Y, Zhao Q L, et al. Differences of cadmium and lead accumulation and transportion among winter wheat varieties[J]. Journal of Triticeae Crops, 2015, 35(8): 1161-1167. |
| [29] | Zhang L G, Zhang C, Du B Y, et al. Effects of node restriction on cadmium accumulation in eight Chinese wheat(Triticum turgidum) cultivars[J]. Science of the Total Environment, 2020, 725. DOI:10.1016/j.scitotenv.2020.138358 |
| [30] | Gao P P, Xue P Y, Dong J W, et al. Contribution of PM2.5-Pb in atmospheric fallout to Pb accumulation in Chinese cabbage leaves via stomata[J]. Journal of Hazardous Materials, 2021, 407. DOI:10.1016/j.jhazmat.2020.124356 |
| [31] |
董俊文, 高培培, 孙洪欣, 等. 设施叶菜类蔬菜重金属镉、铅和砷累积特征及健康风险评价[J]. 环境科学, 2022, 43(1): 481-489. Dong J W, Gao P P, Sun H X, et al. Characteristics and health risk assessment of cadmium, lead, and arsenic accumulation in leafy vegetables planted in a greenhouse[J]. Environmental Science, 2022, 43(1): 481-489. |
| [32] | Tan L T, Zhu Y X, Fan T, et al. OsZIP7 functions in xylem loading in roots and inter-vascular transfer in nodes to deliver Zn/Cd to grain in rice[J]. Biochemical and Biophysical Research Communications, 2019, 512(1): 112-118. |
| [33] | Hao X H, Zeng M, Wang J, et al. A node-expressed transporter OsCCX2 is involved in grain cadmium accumulation of rice[J]. Frontiers in Plant Science, 2018, 9. DOI:10.3389/fpls.2018.00476 |
| [34] | Song Y, Wang Y B N, Mao W F, et al. Dietary cadmium exposure assessment among the Chinese population[J]. PLoS One, 2017, 12(5). DOI:10.1371/journal.pone.0177978 |
| [35] |
熊孜, 李菊梅, 赵会薇, 等. 不同小麦品种对大田中低量镉富集及转运研究[J]. 农业环境科学学报, 2018, 37(1): 36-44. Xiong Z, Li J M, Zhao H W, et al. Accumulation and translocation of cadmium in different wheat cultivars in farmland[J]. Journal of Agro-Environment Science, 2018, 37(1): 36-44. |
| [36] |
刘畅, 徐应明, 黄青青, 等. 不同冬小麦品种镉富集转运及离子组特征差异[J]. 环境科学, 2022, 43(3): 1596-1605. Liu C, Xu Y M, Huang Q Q, et al. Variations in cadmium accumulation and transport and ionomic traits among different winter wheat varieties[J]. Environmental Science, 2022, 43(3): 1596-1605. |
| [37] | Tian S Q, Liang S, Qiao K, et al. Co-expression of multiple heavy metal transporters changes the translocation, accumulation, and potential oxidative stress of Cd and Zn in rice(Oryza sativa)[J]. Journal of Hazardous Materials, 2019, 380. DOI:10.1016/j.jhazmat.2019.120853 |
| [38] | Huang G X, Ding C F, Guo F Y, et al. The role of node restriction on cadmium accumulation in the brown rice of 12 Chinese rice(Oryza sativa L.) cultivars[J]. Journal of Agricultural and Food Chemistry, 2017, 65(47): 10157-10164. |
| [39] | Feng X M, Han L, Chao D Y, et al. Ionomic and transcriptomic analysis provides new insight into the distribution and transport of cadmium and arsenic in rice[J]. Journal of Hazardous Materials, 2017, 331: 246-256. |
| [40] | Yamaji N, Ma J F. Node-controlled allocation of mineral elements in Poaceae[J]. Current Opinion in Plant Biology, 2017, 39: 18-24. |
| [41] | Yan B F, Nguyen C, Pokrovsky O S, et al. Cadmium allocation to grains in durum wheat exposed to low Cd concentrations in hydroponics[J]. Ecotoxicology and Environmental Safety, 2019, 184. DOI:10.1016/j.ecoenv.2019.109592 |
| [42] | Shi G L, Li D J, Wang Y F, et al. Accumulation and distribution of arsenic and cadmium in winter wheat(Triticum aestivum L.) at different developmental stages[J]. Science of the Total Environment, 2019, 667: 532-539. |
| [43] |
冯亚娟, 李廷轩, 蒲勇, 等. 不同镉积累类型小麦各器官镉积累分布规律及机理分析[J]. 作物学报, 2022, 48(7): 1761-1770. Feng Y J, Li T X, Pu Y, et al. Characteristics of cadmium accumulation and distribution in different organs of wheat with different cadmium-accumulating type[J]. Acta Agronomica sinica, 2022, 48(7): 1761-1770. |
| [44] | Chen Y, Moore K L, Miller A J, et al. The role of nodes in arsenic storage and distribution in rice[J]. Journal of Experimental Botany, 2015, 66(13): 3717-3724. |
| [45] | Greger M, Landberg T. Silicon reduces cadmium and arsenic levels in field-grown crops[J]. Silicon, 2019, 11(5): 2371-2375. |
| [46] |
张世杰, 孙洪欣, 薛培英, 等. 叶面施硅时期对冬小麦镉铅砷累积的阻控效应研究[J]. 河北农业大学学报, 2018, 41(3): 1-6, 36. Zhang S J, Sun H X, Xue P Y, et al. Study on the effect of foliar application of silicon fertilizer on lead cadmium and arsenic accumulation in winter wheat(Triticum aestivum L.)[J]. Journal of Hebei Agricultural University, 2018, 41(3): 1-6, 36. |
| [47] |
李道军. 砷和镉在小麦(Triticum aestivum L.)体内积累和分布及磷硅对小麦砷吸收和转运的影响[D]. 南京: 南京农业大学, 2016. Li D J. Arsenic(As) and cadmium(Cd) accumulation and distribution in wheat(Triticum aestivum L.) and the influce of phosphate and silicate[D]. Nanjing: Nanjing Agricultural University, 2016. |