 2023, Vol. 44
2023, Vol. 44



由于工业高速发展和城市化稳步推进, 农田土壤重金属污染日益严重.例如, 污水灌溉和农药化肥施用导致土壤Pb和Cd的污染[1].Pb和Cd是非必需元素, 通过土壤-植物吸收转运途径影响农作物的生物量, 阻碍植物从土壤中吸收必需养分, 影响农作物的正常生长[2~4], 最终富集于作物中进一步通过食物链威胁人类健康[5].且多种重金属间相互作用可改变重金属在土壤中的存在形态, 进而影响其植物可利用性[6, 7].
现有研究认为土壤pH、CEC、有机质和黏土矿物等土壤性质是影响重金属有效态的重要因素[8].我国现行农田重金属风险管控标准(GB 15618-2018)按土壤pH划分了单一重金属的标准限值, 但未考虑多种重金属共存可能产生的效应.而不同金属离子对土壤各活性组分亲和力的差异, 影响金属离子在土壤固-液相中的分配[9].例如, 黏土矿物对Cd的亲和力更强, 而铁锰氧化物则对Pb的亲和力更强[10], 多种金属阳离子共存会竞争土壤表面活性位点, 因而不同金属间会产生交互效应[11].因此, 即使处于同等污染水平, 可被农作物吸收利用的重金属含量在不同土壤中也可能存在显著差异.因此, 人们越来越关注复合重金属污染的交互作用行为及其对作物的联合毒性.
目前, 普遍认为重金属被植物吸收和转运是利用必需元素的膜转运蛋白进行[12].大麦和水稻吸收Cd均利用了锰的膜转运蛋白NRAMP5[13, 14], 而拟南芥根利用钙通道吸收Pb[15].与在土壤中类似, 多种重金属离子在通过植物根系质膜时也存在竞争作用, 钙离子和镁离子同时抑制植物根系吸收Pb和Cd[16].小麦将重金属从共质体转运至木质部的转运基因HMA5对Pb和Cd均有响应, 而位于质膜和细胞器膜上的其他转运基因只对Cd有响应[17].Cd易从细胞壁上交换下来并游离于植物细胞中, Pb则常固定于植物根系和细胞壁上[18, 19].然而, Pb和Cd在植物体中转运迁移过程的交互作用行为还不清楚.
因此, 本研究以我国东北、中部和西北地区的3种土壤(海伦黑土、咸宁棕红壤和长武黑垆土)为例, 通过种植小麦研究土壤-小麦(Triticum aestivum L.)体系Pb和Cd的差异, 评估不同土壤-小麦体系中Pb和Cd交互作用及其对小麦富集转运重金属的影响, 以期为复合污染农田土壤的粮食安全风险管控提供科学依据.
1 材料与方法 1.1 供试土壤本研究选取我国3种代表性农田土壤黑土、棕红壤和黑垆土开展试验, 黑土采集自黑龙江省海伦市(N47°39′45.49″, E126°50′43.47″), 棕红壤采集自湖北省咸宁市(N29°50′38.09″, E114°19′1.37″), 黑垆土采集自陕西省长武县(N35°12′24.12″, E107°47′42.36″).采集0~20 cm表层土壤, 风干后过2 mm筛储存于聚乙烯容器中.表 1为3种土壤的基本理化性质[20].黑土的有机质含量最高(54.71g·kg-1), 棕红壤的pH(5.61)和CEC(0.04 cmol·kg-1)最低, 黑垆土的pH(8.25)和碳酸钙含量(73.88 g·kg-1)最高.黑土、棕红壤、黑垆土的Cd背景值分别为0.09、0.14和0.15 mg·kg-1, Pb背景值分别为17.70、21.00和17.46 mg·kg-1.
|
|
表 1 土壤理化性质 Table 1 Soil physical and chemical properties |
1.2 盆栽试验
设置1个空白对照处理和4个(Cd, Pb)复合处理(单位为mg·kg-1), 即(0.3, 125)、(0.3, 250)、(0.6, 125)和(0.6, 250).每个处理设置4个重复.每个处理取过2 mm筛的风干土30 kg, 将适量的硝酸镉[Cd(NO3)2·4H2O]和硝酸铅[Pb(NO3)2]溶液均匀喷入土壤并充分混匀, 补充超纯水使土壤含水量为土壤田间持水量的80%, 根据总质量变化补充超纯水, 老化持续时间共4个月, 使Cd和Pb在土壤颗粒表面达到吸附-解吸平衡.
取老化后的土5 kg装盆, 施加尿素[CO(NH2)2] 1.61 g、磷酸钙[Ca3(PO4)2)] 0.55 g和硫酸钾(K2SO4) 0.93 g作为基肥与老化土壤混合均匀, 使ω(N)为0.15 g·kg-1, ω(P2O5)为0.05 g·kg-1和ω(K2O)为0.1 g·kg-1, 以保证小麦生长.盆栽作物为冬小麦“郑麦9023号”, 此品种为农业部推荐长江中下游小麦主导品种, 种植面积居我国当前优质小麦品种种植面积首位.每盆播种20粒小麦种子, 待出苗4周后, 按每盆8株定苗.每3~5 d采用称重法为植物补水, 维持土壤含水量为田间最大持水量的70%.在小麦分蘖期与拔节期各追肥一次, 将200 mL 0.5%的磷酸二氢钾施于根部附近.培育5个月收获成熟期小麦.
1.3 测定指标与方法 1.3.1 小麦生长指标及重金属含量记录每株小麦株高, 并计算各处理中小麦的平均株高.分别保存小麦植株的根、茎、叶和籽粒这4个部分, 然后用超纯水冲洗干净并吸干水分, 将鲜样在105℃杀青30 min后, 于65℃烘至恒重, 称量干重.用10 mL HNO3-HClO4(体积比为4∶1)溶液于电热板上加热消解小麦植株各部位, 消解液定容至40 mL后, 4℃冰箱储存待测.用石墨炉原子吸收光谱仪(AA 240Z, Agilent America)分析Pb和Cd含量, 小麦各部位消解待测液的Cd测定范围为0.1~5 μg·L-1; 小麦根Pb、茎叶Pb和籽粒消解待测液Pb的测定范围分别为20~200、5~50和0.1~5 μg·L-1, 检出限为0.01 μg·L-1.同时分析标准物质(圆白菜GBW 10014, 国家标准物质中心), 为质量控制提供参考, 回收率在88.9%~107.5%之间.
1.3.2 土壤性质及重金属形态通过土水比1∶2.5电位法(NYT 1377-2007)测定小麦收获后土壤pH.水溶性有机碳含量(dissolved organic carbon, DOC)采用去离子水按土水比1∶10(质量浓度)浸提, 总有机碳分析仪(Enviro TOC, Elementar Genrmany)测定.
土壤Cd和Pb全量采用10 mL HNO3-HCl-HF(3∶1∶1, 体积比)溶液通过微波消解仪(Mars, America)消解过100目筛的土壤样品, 消解液定容至50 mL后, 于4℃冰箱储存待测.同时分析标准物质(黄棕壤GBW 07403, 国家标准物质中心), 为质量控制提供参考, 回收率在82%~117%之间.黑土和棕红壤中有效态Pb和Cd含量采用0.43 mol·L-1 HNO3溶液、黑垆土采用0.05 mol·L-1 EDTA-2Na溶液提取[21~24], 按固液比1∶5(质量浓度)于25℃下以180 r·min-1速度振荡2 h后, 再以转速5 000 r·min-1离心8 min获取浸提液.用石墨炉原子吸收光谱仪(AA 240Z, Agilent America)分析消解液和浸提液的Cd浓度, 测定范围为0.2~5 μg·L-1; 用火焰原子吸收光谱仪(AA 240 FS, Agilent America)测定Pb浓度, 测定范围为0.5~20 mg·L-1.
3种土壤中水溶态Cd和Pb含量采用去离子水按土水比1∶10(质量浓度)于25℃以180 r·min-1转速振荡2 h后, 再以5 000 r·min-1离心8 min获取浸提液待测.使用石墨炉原子吸收光谱仪(AA 240Z, Agilent America)分析浸提液的Cd和Pb浓度, 浸提液Cd测定范围为0.1~5 μg·L-1, 浸提液Pb的测定范围为1~20 μg·L-1, 浸提液Cd的检出限为0.09 μg·L-1, 浸提液Pb的检出限为0.73μg·L-1.
本研究所用试剂均购自国药集团化学试剂有限公司, 纯度为优级纯.
1.4 数据分析小麦转运系数(translocation factor, TF)反映重金属在植物体各部位间迁移转运的能力, 及其分布规律.各部位转运系数计算公式如下:
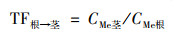
|
(1) |
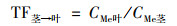
|
(2) |
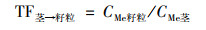
|
(3) |
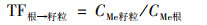
|
(4) |
式中CMe表示对应小麦部位的Cd或Pb含量.
相对转运系数(relative translocation factor, RTF)用于分析Pb和Cd复合处理对小麦重金属转运能力的影响, 其计算公式为:

|
(5) |
当RTF < 0时, 表明Pb和Cd复合处理减弱了小麦转运重金属能力; RTF>0时, 表明Pb和Cd复合处理增强了小麦转运重金属能力, 且RTF绝对值越大, 说明Pb和Cd交互作用对小麦转运影响越强.
采用Origin 2018软件绘图, 使用SPSS 22.0的配对样本T检验(paired-samples T test)对不同处理测定结果进行显著性分析及单因素方差分析(one-way ANOVA), 对每种土壤的各测定结果进行显著性分析, 以P < 0.05判定差异显著.
2 结果与分析 2.1 种植后土壤pH和DOC含量表 2是Pb和Cd复合处理下3种土壤种植小麦后的土壤pH和DOC含量.棕红壤、黑土和黑垆土的pH范围分别为4.37~4.91、5.10~5.39和7.57~7.89; 黑土的ω(DOC)最高, 棕红壤次之, 黑垆土最低, 分别为90.31~132.28、40.81~48.55和20.66~41.68 mg·kg-1.不同土壤的pH和DOC差异极显著.
|
|
表 2 小麦种植后土壤pH和DOC含量1) Table 2 Soil pH and DOC after wheat planting |
与3种土壤空白处理pH相比, Pb和Cd复合处理下黑土pH显著降低0.23~0.29个单位; 棕红壤pH仅在0.6, 125处理(Cd, Pb处理, 单位为mg·kg-1)下显著降低0.54个单位, 其余复合处理下变化不显著; 黑垆土pH除0.6, 250处理(Cd, Pb处理, 单位为mg·kg-1)下升高0.14个单位, 其它复合处理降低0.08~0.18个单位.Pb和Cd复合处理对棕红壤的DOC含量影响不显著; 而黑土ω(DOC)随Cd, Pb复合处理(单位为mg·kg-1)从0.3, 125增加至0.6, 250而显著降低41.97 mg·kg-1; 0.6, 250处理(Cd, Pb处理, 单位为mg·kg-1)下黑垆土的ω(DOC)较空白处理显著降低了21.02 mg·kg-1.
2.2 小麦的生长特性Pb和Cd复合处理下3种土壤种植小麦的株高和地上部干重如表 3所示.棕红壤、黑土和黑垆土的小麦株高依次降低, 分别为57.69~64.20、52.83~56.48和43.32~51.81 cm; 而小麦地上部干重随黑土、棕红壤和黑垆土依次降低, 其值分别为26.04~32.93、16.76~24.00和12.09~15.33 g·pot-1. 3种土壤间小麦地上部干重和株高均存在极显著差异.
|
|
表 3 3种Pb和Cd复合土壤种植小麦的株高和干重1) Table 3 Plant height and dry weight of wheat grown in three Pb-Cd composite soils |
与3种土壤空白处理种植小麦的株高相比, 黑土0.3, 250处理(Cd, Pb处理, 单位为mg·kg-1)种植小麦株高比空白处理显著增高3.1 cm, 其它处理下株高均无显著变化; 棕红壤中, 除0.3, 125处理(Cd, Pb处理, 单位为mg·kg-1)小麦株高无显著变化外, 其它复合处理种植小麦株高比空白处理显著降低2.73~5.42 cm; 而黑垆土复合处理种植小麦株高比空白处理显著增高2.68~8.49 cm.与3种土壤空白处理地上部干重相比, 复合处理对黑土种植小麦干重均无显著影响; 棕红壤中除0.3, 250处理(Cd, Pb处理, 单位为mg·kg-1)干重无显著变化外, 其他处理种植小麦干重比空白处理显著降低0.55~6.78 g; 黑垆土0.3, 125处理(Cd, Pb处理, 单位为mg·kg-1)种植小麦干重比空白处理显著降低2.57 g·pot-1, 其它处理下均无显著变化.
2.3 3种复合污染土壤中Cd和Pb形态图 1是黑土、棕红壤和黑垆土中Cd、Pb全量、有效态和水溶态这3种形态含量.除0.3, 125处理(Cd, Pb处理, 单位为mg·kg-1)外, 各复合处理中棕红壤的Cd全量显著低于黑土和黑垆土; 除0.6, 250处理(Cd, Pb处理, 单位为mg·kg-1)外, 各复合处理中黑垆土的Cd全量显著高于黑土和棕红壤[图 1(a)].复合处理下3种土壤的有效态Cd含量依次为:CCd-棕红壤>CCd-黑土>CCd-黑垆土[图 1(b)].除0.3, 125处理(Cd, Pb处理, 单位为mg·kg-1)外, 其他复合处理中3种土壤的Pb全量均为棕红壤最高, 黑垆土次之, 黑土最低[图 1(d)].且3种土壤中有效态Pb含量依次为:CPb-棕红壤>CPb-黑垆土>CPb-黑土[图 1(e)].棕红壤的水溶态Cd和Pb含量均显著高于黑土和黑垆土的[图 1(c)和1(f)].3种复合污染土壤中, 棕红壤的Cd和Pb活性最高, 黑垆土Cd和Pb活性最低.

|
小写字母表示不同种土壤的不同复合处理之间金属含量的差异(P < 0.05) 图 1 3种土壤中3种形态的Cd和Pb含量 Fig. 1 Three forms of Cd and Pb content in three types of soils |
图 2是黑土、黑垆土和棕红壤种植小麦各部位的Cd含量.黑垆土中小麦根的ω(Cd)可达0.69~8.78 mg·kg-1, 显著高于棕红壤和黑土, 分别为0.59~4.68 mg·kg-1和0.18~2.71 mg·kg-1[图 2(a)].棕红壤中小麦茎和叶Cd含量显著高于黑土的及黑垆土的, 其中茎ω(Cd)分别为0.30~2.84、0.16~1.18和0.07~1.23 mg·kg-1, 叶ω(Cd)分别为0.69~7.09、0.22~1.82和0.11~1.55 mg·kg-1[图 2(b)和2(c)].除空白外, 3种土壤种植的小麦籽粒ω(Cd)均超过食品质量安全标准限值[0.1 mg·kg-1, GB 2762-2022, 图 2(d)中虚线], 棕红壤中小麦超标约4.4~9.7倍, 黑土中小麦超标约3.0~6.2倍, 黑垆土中小麦超标约1.4~2.9倍[图 2(d)].

|
图 2 3种土壤中小麦4个部位Cd含量 Fig. 2 Amount of Cd in the four parts of wheat planted in three types of soil |
3种Pb和Cd复合污染土壤中, Pb对小麦各部位Cd累积产生了不同程度的影响.在0.3 mg·kg-1和0.6 mg·kg-1 Cd处理下, 随Pb添加量增大, 黑土中小麦根Cd含量分别下降了49.1%和36.2%; 棕红壤中小麦根Cd含量分别下降了13.4%和24.9%; 而黑垆土中小麦根Cd含量在0.3 mg·kg-1处理下, 随Pb增加显著上升了61.7%[图 2(a)].Pb对3种土壤的小麦茎、对棕红壤和黑垆土小麦叶的Cd含量均无显著影响[图 2(b)和2(c)].黑土中小麦叶的Cd含量在0.3 mg·kg-1处理下, 随Pb添加量增大而下降了44.2%, 而0.6 mg·kg-1 Cd处理下无显著差异[图 2(c)].在0.3 mg·kg-1和0.6 mg·kg-1 Cd处理下, 黑土中小麦籽粒Cd的含量分别随Pb添加量增大而显著下降了51%和33%; 棕红壤中小麦籽粒Cd含量在0.3 mg·kg-1 Cd处理下随Pb添加量增大而显著减少17.9%, 在0.6 mg·kg-1 Cd处理下无显著差异; 同时, 黑垆土中小麦籽粒Cd含量随Pb添加量增加无显著差异[图 2(d)].
2.5 Cd对小麦Pb累积分配的影响图 3是黑土、黑垆土和棕红壤种植小麦各部位Pb的含量.复合处理下, 棕红壤中小麦根和茎Pb的含量显著高于黑土的和黑垆土的, 根ω(Pb)分别在82.41~267.6、47.79~117.28和30.64~76.47 mg·kg-1范围内; 茎ω(Pb)分别为8.62~23.44、3.89~6.00和4.01~9.19 mg·kg-1[图 3(a)和3(b)].0.3, 125处理(Cd, Pb处理, 单位为mg·kg-1)下, 棕红壤中小麦叶ω(Pb)为21.30 mg·kg-1, 显著高于黑土和黑垆土; 0.6, 250处理(Cd, Pb处理, 单位为mg·kg-1)下, 黑垆土中小麦叶的ω(Pb)高达73.66 mg·kg-1, 显著高于棕红壤和黑土[图 3(c)].黑土中小麦籽粒ω(Pb)未超过食品质量安全标准限值[0.2 mg·kg-1, 图 3(d)虚线]; 黑垆土中除0.6, 250处理(Cd, Pb处理, 单位为mg·kg-1)的籽粒Pb含量超标1.1倍, 其它处理下籽粒Pb含量均未超标; 而棕红壤的复合处理中籽粒Pb含量超标约3~15倍[图 3(d)].

|
图 3 3种土壤中小麦4个部位Pb含量 Fig. 3 Amount of Pb in the four parts of wheat planted in three types of soil |
同时, 在3种Pb和Cd复合污染土壤中, Cd对小麦各部位的Pb累积也产生了不同程度的影响.随Cd添加量增加, 125 mg·kg-1 Pb处理下黑土中小麦根部Pb含量显著减少26.5%, 而棕红壤中显著增加26.4%, 黑垆土中则无显著差异; 250 mg·kg-1 Pb处理下黑土和黑垆土中小麦根部Pb含量分别显著降低了10.5%和20.7%, 而棕红壤中显著增加20.2%[图 3(a)].125 mg·kg-1 Pb处理下棕红壤中小麦茎部Pb含量随Cd添加量升高而增加12.2%, 其它处理下小麦茎部Pb的含量随Cd添加量升高均显著下降约17.9~34.6%[图 3(b)].250 mg·kg-1 Pb处理下黑垆土中小麦叶的Pb含量随Cd添加量升高显著增加约92.6%, 其它土壤复合处理下小麦叶的Pb含量随Cd变化无显著差异[图 3(c)].棕红壤中麦粒Pb含量在125 mg·kg-1和250 mg·kg-1 Pb处理下随Cd添加量升高分别显著增加73.2%和19.1%, 而黑土和黑垆土的小麦Pb含量随Cd变化均无显著差异[图 3(d)].
3 讨论 3.1 土壤性质对Pb和Cd活性的影响土壤理化性质直接影响土壤重金属迁移性、有效性和活性[25].本研究中pH最低的棕红壤中Pb和Cd活性最高, 且各处理间水溶态Pb和Cd含量变化与pH变化呈相反趋势.低pH和低CEC等使土壤固持重金属能力较弱, 有利于增强重金属活性[26], 从而使土壤颗粒表面的重金属交互作用更显著.0.6 mg·kg-1 Cd处理的棕红壤pH随外源Pb的增加而显著升高, 水溶态Cd含量显著降低; 250 mg·kg-1 Pb处理的棕红壤的pH随外源Cd的增加而升高, 水溶态Pb含量显著降低[表 2, 图 1(c)和1(f)].棕红壤中高活性重金属胁迫植物产生应激反应, 根系释放小分子有机酸等分泌物[27].在正常的植物胞质pH下, 有机酸以阴离子的形式存在, 释放时可与质子结合, 导致根际碱化[28, 29].此外, 金属胁迫会降低植物中H+-ATP酶的泵送活性, 从而导致质子流出减少[30].上述过程使土壤pH上升, 从而降低有效态重金属含量.
在各Pb和Cd复合处理间, 黑土和黑垆土的pH和水溶态Pb和Cd含量均无显著变化.黑土中大量有机质可以通过吸附和螯合作用来固定重金属, 降低Pb和Cd的活性[31].黑垆土的高pH、CEC和碳酸钙含量表明铁锰氧化物和黏土矿物等活性组分表面的负电荷数量更多, 对重金属离子的吸附固持能力更强[32]; 且高pH环境会使得碳酸盐结合态和铁锰氧化态重金属含量更高[8].因此黑垆土中Pb和Cd的有效态和水溶态含量在3种土壤中最低(图 1).
3.2 土壤-小麦转运体系中的Pb和Cd交互作用重金属在进入植物体的过程中会通过竞争转运通道而发生交互作用[16, 33].而植物体对重金属的转运能力受多重因素影响, 包括植物品种、培育土壤性质和生长特性等.本研究中3种土壤种植的同种小麦对重金属的富集转运机制有显著差异(图 4).黑土中Pb和Cd复合处理抑制了小麦Cd和Pb从根向茎的转运[图 4(a)和4(d)], 这是因为高有机质土壤中羧基、氨基及羟基等官能团会螯合重金属, 有机结合态重金属被固定在小麦根部, 使其向茎中转运受阻[31], 其它形态的Cd从韧皮部进入木质部, 并通过钙、锌等营养元素的转运通道进一步向上运输到籽粒[34, 35].黑土-小麦的株高和干重随外源Pb的增加而增加(表 3), 且转运重金属能力均随着外源Pb或Cd含量的增加而减弱[图 4(a)和4(d)], Pb和Cd间存在拮抗作用.例如, 随着Cd添加量从0.3 mg·kg-1增加至0.6 mg·kg-1, 125 mg·kg-1 Pb处理下黑土种植小麦Pb从根向籽粒的相对转运系数降低了15%, 拮抗作用增强[图 4(d)].这可能是由于金属离子会利用与水分子配位的氢键作用于DNA, 导致碱基对氢键断裂[36, 37], 从而进一步使小麦转运基因表达减弱.因此, 黑土中小麦根和籽粒的Cd和Pb含量随着外源Pb和Cd的增加而减少(图 2和图 3).

|
图 4 Pb和Cd复合污染土壤种植小麦Cd及Pb的相对转移系数 Fig. 4 Cd and Pb relative translocation factor of wheat in Pb-Cd composite soil |
本研究棕红壤-小麦体系中Pb和Cd活性和转运能力最强.一方面棕红壤的土壤性质导致重金属活性高, 另一方面棕红壤中小麦生长状态最佳, 特别是低含量0.3, 125处理(Cd, Pb处理, 单位为mg·kg-1)下小麦株高最高(表 3).低剂量Pb和Cd会增强作物的光合、蒸腾速率和气孔导度[38, 39], 使得与光合和呼吸作用相关的叶部能量和物质代谢加快, 进一步增强Cd的转运, 导致叶Cd含量高于根Cd含量(图 2).复合处理使棕红壤-小麦的Cd和Pb转运能力增强[图 4(b)和4(e)].棕红壤中小麦转运机制中, Cd对小麦Pb含量的影响强于Pb对小麦Cd含量的影响(图 2和图 3).棕红壤-小麦株高随Cd添加量增加而显著降低(表 3), 且Pb从茎向籽粒转运能力随Cd添加量增加而增强, 存在协同作用.其中随着Cd添加量从0.3 mg·kg-1增加至0.6 mg·kg-1, 250 mg·kg-1 Pb处理下黑土种植小麦Pb从茎向籽粒的相对转运系数升高了3.15倍, 协同作用增强[图 4(e)].Cd通常与烟草胺或植物螯合蛋白结合后在韧皮部运输[40], 但进入木质部后向上运输的金属需剥离植物螯合素等大分子[41, 42].与Pb相比, 植物螯合素对Cd有更强的亲和力, 更易将Pb交换下来, 从而增强了Pb在木质部中的运移[43].
与黑土和棕红壤相比, 黑垆土种植小麦的生长特性表现最差(表 3), 高pH和高碳酸钙的黑垆土使小麦受到碱胁迫.碱胁迫可使小麦中Ca2+、Mg2+、Zn2+和Cu2+析出, 导致营养缺乏, 生长较差[44], 且析出的大量阳离子竞争作用会使Pb和Cd在小麦中的转运受限.相比于空白处理, Pb和Cd的添加进一步抑制了小麦Cd从茎向麦粒和叶转运的能力[图 4(c)].同时, 碱胁迫还会诱导植物分泌大量酚酸和脂肪酸[45], 重金属与大分子酸稳定螯合后难以向麦粒运输, 因此黑垆土中小麦麦粒Cd和Pb含量维持在较低水平[图 2(d)和3(d)].250 mg·kg-1 Pb处理下, 黑垆土中小麦中Pb从茎向叶转运增强使小麦叶Pb含量骤升[图 3(c)和4(f)].植物对Pb的转运能力较弱, 难以进入扩大维管束并向籽粒部位转运, 只能通过弥漫性维管束转移至叶片中储存[10].
4 结论(1) 由于棕红壤的pH最低, Pb和Cd在土壤界面的交互作用最显著, 其Cd和Pb活性最高; 而黑土受有机质影响, 黑垆土受碱化和碳酸钙影响, 二者的Pb和Cd活性较低.
(2) 3种土壤中黑垆土种植小麦生长最差, Pb和Cd复合污染对黑土中小麦生长影响最小, 但抑制了棕红壤中小麦的生长.
(3) pH和有机质含量是影响小麦转运Pb和Cd能力及其交互作用行为的关键因素.Pb和Cd复合污染在高有机质的黑土中存在拮抗作用, 在酸化的棕红壤存在协同作用, 而在碱化的黑垆土中无显著交互作用.
| [1] |
戴翌晗. 典型矿冶城市农田土壤重金属污染来源解析[J]. 环境生态学, 2022, 4(5): 25-31. Dai Y H. Source apportionment of heavy metals in agricultural soil of a typical mining and metallurgy city[J]. Environmental Ecology, 2022, 4(5): 25-31. |
| [2] | Kushwaha A, Hans N, Kumar S, et al. A critical review on speciation, mobilization and toxicity of lead in soil-microbe-plant system and bioremediation strategies[J]. Ecotoxicology and Environmental Safety, 2018, 147: 1035-1045. DOI:10.1016/j.ecoenv.2017.09.049 |
| [3] | Bali S, Jamwal V L, Kohli S K, et al. Jasmonic acid application triggers detoxification of lead(Pb) toxicity in tomato through the modifications of secondary metabolites and gene expression[J]. Chemosphere, 2019, 235: 734-748. DOI:10.1016/j.chemosphere.2019.06.188 |
| [4] | Ismael M A, Elyamine A M, Moussa M G, et al. Cadmium in plants: uptake, toxicity, and its interactions with selenium fertilizers[J]. Metallomics, 2019, 11(2): 255-277. DOI:10.1039/C8MT00247A |
| [5] | Lanier C, Bernard F, Dumez S, et al. Combined effect of Cd and Pb spiked field soils on bioaccumulation, DNA damage, and peroxidase activities in Trifolium repens[J]. Environmental Science and Pollution Research, 2016, 23(2): 1755-1767. DOI:10.1007/s11356-015-5414-6 |
| [6] | Kavvadias V, Paschalidis C, Vavoulidou E, et al. Effects of soil amended with cadmium and lead on growth, yield, and metal accumulation and distribution in parsley[J]. Communications in Soil Science and Plant Analysis, 2012, 43(1-2): 161-175. DOI:10.1080/00103624.2012.634708 |
| [7] | Zhao Z J, Nan Z R, Wang Z W, et al. Interaction between Cd and Pb in the soil-plant system: a case study of an arid oasis soil-cole system[J]. Journal of Arid Land, 2014, 6(1): 59-68. DOI:10.1007/s40333-013-0194-7 |
| [8] |
赵一鸣, 董颖博, 林海, 等. 土壤理化性质对重金属形态的影响[J]. 农业工程, 2018, 8(12): 38-43. Zhao Y M, Dong Y B, Lin H, et al. Influence of soil physical and chemical properties on forms of heavy metals[J]. Agricultural Engineering, 2018, 8(12): 38-43. |
| [9] | Qiu H, Versieren L, Rangel G G, et al. Interactions and toxicity of Cu-Zn mixtures to Hordeum vulgare in different soils can be rationalized with bioavailability-based prediction models[J]. Environmental Science & Technology, 2016, 50(2): 1014-1022. |
| [10] | Li T Q, Jiang H, Yang X E, et al. Competitive sorption and desorption of cadmium and lead in paddy soils of eastern China[J]. Environmental Earth Sciences, 2013, 68(6): 1599-1607. DOI:10.1007/s12665-012-1853-2 |
| [11] | Evangelou V P, Marsi M, Vandiviere M M. Stability of Ca2+-, Cd2+-, Cu2+--[illite-humic] complexes and pH influence[J]. Plant and Soil, 1999, 213(1-2): 63-74. |
| [12] | Zhao F J, Tang Z, Song J J, et al. Toxic metals and metalloids: uptake, transport, detoxification, phytoremediation, and crop improvement for safer food[J]. Molecular Plant, 2022, 15(1): 27-44. DOI:10.1016/j.molp.2021.09.016 |
| [13] | Sasaki A, Yamaji N, Yokosho K, et al. Nramp5 is a major transporter responsible for manganese and cadmium uptake in rice[J]. The Plant Cell, 2012, 24(5): 2155-2167. DOI:10.1105/tpc.112.096925 |
| [14] | Wu D Z, Yamaji N, Yamane M, et al. The HvNramp5 transporter mediates uptake of cadmium and manganese, but not iron[J]. Plant Physiology, 2016, 172(3): 1899-1910. DOI:10.1104/pp.16.01189 |
| [15] | Moon J Y, Belloeil C, Ianna M L, et al. Arabidopsis CNGC family members contribute to heavy metal ion uptake in plants[J]. International Journal of Molecular Sciences, 2019, 20(2): 413-428. DOI:10.3390/ijms20020413 |
| [16] | Kim Y Y, Yang Y Y, Lee Y. Pb and Cd uptake in rice roots[J]. Physiologia Plantarum, 2002, 116(3): 368-372. DOI:10.1034/j.1399-3054.2002.1160312.x |
| [17] | Aprile A, Sabella E, Francia E, et al. Combined effect of cadmium and lead on durum wheat[J]. International Journal of Molecular Sciences, 2019, 20(23): 5891-5908. DOI:10.3390/ijms20235891 |
| [18] | Pourrut B, Shahid M, Dumat C, et al. Lead uptake, toxicity, and detoxification in plants[J]. Reviews of Environmental Contamination and Toxicology, 2011, 213: 113-136. |
| [19] | Landberg T, Jensén P, Greger M. Strategies of cadmium and zinc resistance in willow by regulation of net accumulation[J]. Biologia Plantarum, 2011, 55(1): 133-140. DOI:10.1007/s10535-011-0018-6 |
| [20] |
樊宇. 几种典型土壤中Pb/Cd交互作用对小麦积累转运的影响[D]. 武汉: 华中农业大学, 2021. Fan Y. The effect of lead and cadmium interaction in several typical soils on the accumulation and transportation of wheat[D]. Wuhan: Huazhong Agricultural University, 2021. |
| [21] |
吕明超, 宋静, 余海波, 等. 不同振荡方式对土壤有效态重金属提取的影响[J]. 农业环境科学学报, 2014, 33(2): 339-344. Lü M C, Song J, Yu H B, et al. Effects of different oscillation modes on extraction of available heavy metals in soils[J]. Journal of Agro-Environment Science, 2014, 33(2): 339-344. |
| [22] |
屈晓泽, 许伟伟, 赵晓鹏, 等. 基于多表面形态模型的土壤镉在小白菜中的富集效应[J]. 环境化学, 2019, 38(9): 1945-1954. Qu X Z, Xu W W, Zhao X P, et al. Accumulation of Cd in soil-cabbage system based on multi-surface speciation model[J]. Environmental Chemistry, 2019, 38(9): 1945-1954. |
| [23] |
李发生, 韩梅, 熊代群, 等. 不同浸提剂对几种典型土壤中重金属有效态的浸提效率研究[J]. 农业环境科学学报, 2003, 22(6): 704-706. Li F S, Han M, Xiong D Q, et al. Efficiency of some extractants for available heavy metals from several typical soils[J]. Journal of Agro-Environment Science, 2003, 22(6): 704-706. |
| [24] |
袁峰, 唐先进, 吴骥子, 等. 两种铁基材料对污染农田土壤砷、铅、镉的钝化修复[J]. 环境科学, 2021, 42(7): 3535-3548. Yuan F, Tang X J, Wu J Z, et al. Simultaneous immobilization of arsenic, lead, and cadmium in paddy soils using two iron-based materials[J]. Environmental Science, 2021, 42(7): 3535-3548. DOI:10.13227/j.hjkx.202010183 |
| [25] | 周启星, 黄国宏. 环境生物地球化学及全球环境变化[M]. 北京: 科学出版社, 2001. |
| [26] | Zeng F R, Ali S, Zhang H T, et al. The influence of pH and organic matter content in paddy soil on heavy metal availability and their uptake by rice plants[J]. Environmental Pollution, 2011, 159(1): 84-91. DOI:10.1016/j.envpol.2010.09.019 |
| [27] | Fu H J, Yu H Y, Li T X, et al. Influence of cadmium stress on root exudates of high cadmium accumulating rice line(Oryza sativa L.)[J]. Ecotoxicology and Environmental Safety, 2018, 150: 168-175. DOI:10.1016/j.ecoenv.2017.12.014 |
| [28] | Javed M T, Stoltz E, Lindberg S, et al. Changes in pH and organic acids in mucilage of Eriophorum angustifolium roots after exposure to elevated concentrations of toxic elements[J]. Environmental Science and Pollution Research, 2013, 20(3): 1876-1880. DOI:10.1007/s11356-012-1413-z |
| [29] | Javed M T, Akram M S, Habib N, et al. Deciphering the growth, organic acid exudations, and ionic homeostasis of Amaranthus viridis L. and Portulaca oleracea L. under lead chloride stress[J]. Environmental Science and Pollution Research, 2018, 25(3): 2958-2971. DOI:10.1007/s11356-017-0735-2 |
| [30] | Janicka-Russak M, Kabała K, Burzyński M. Different effect of cadmium and copper on H+-ATPase activity in plasma membrane vesicles from Cucumis sativus roots[J]. Journal of Experimental Botany, 2012, 63(11): 4133-4142. DOI:10.1093/jxb/ers097 |
| [31] | Covelo E F, Vega F A, Andrade M L. Competitive sorption and desorption of heavy metals by individual soil components[J]. Journal of Hazardous Materials, 2007, 140(1-2): 308-315. |
| [32] |
王怡雯, 芮玉奎, 李中阳, 等. 冬小麦吸收重金属特征及与影响因素的定量关系[J]. 环境科学, 2020, 41(3): 1482-1490. Wang Y W, Rui Y K, Li Z Y, et al. Characteristics of heavy metal absorption by winter wheat and its quantitative relationship with influencing factors[J]. Environmental Science, 2020, 41(3): 1482-1490. |
| [33] | Gonçalves J F, Antes F G, Maldaner J, et al. Cadmium and mineral nutrient accumulation in potato plantlets grown under cadmium stress in two different experimental culture conditions[J]. Plant Physiology and Biochemistry, 2009, 47(9): 814-821. |
| [34] | Uraguchi S, Kamiya T, Sakamoto T, et al. Low-affinity cation transporter(OsLCT1) regulates cadmium transport into rice grains[J]. Proceedings of the National Academy of Sciences of the United States of America, 2011, 108(52): 20959-20964. |
| [35] | Ma J F, Shen R F, Shao J F. Transport of cadmium from soil to grain in cereal crops: a review[J]. Pedosphere, 2021, 31(1): 3-10. |
| [36] | Saleh S R, Kandeel M M, Ghareeb D, et al. Wheat biological responses to stress caused by cadmium, nickel and lead[J]. Science of the Total Environment, 2020, 706: 136013-136023. |
| [37] | Theophanides T, Anastassopoulou J. The effects of metal ion contaminants on the double stranded DNA helix and diseases[J]. Journal of Environmental Science and Health, Part A, 2017, 52(10): 1030-1040. |
| [38] | Ban Y H, Xu Z Y, Yang Y R, et al. Effect of dark septate endophytic fungus Gaeumannomyces cylindrosporus on plant growth, photosynthesis and Pb tolerance of maize(Zea mays L.)[J]. Pedosphere, 2017, 27(2): 283-292. |
| [39] | Muszyńska E, Hanus-Fajerska E, Ciarkowska K. Studies on lead and cadmium toxicity in Dianthus carthusianorum calamine ecotype cultivated in vitro[J]. Plant Biology, 2018, 20(3): 474-482. |
| [40] | Hazama K, Nagata S, Fujimori T, et al. Concentrations of metals and potential metal-binding compounds and speciation of Cd, Zn and Cu in phloem and xylem saps from castor bean plants(Ricinus communis) treated with four levels of cadmium[J]. Physiologia Plantarum, 2015, 154(2): 243-255. |
| [41] | Mendoza-Cózatl D G, Butko E, Springer F, et al. Identification of high levels of phytochelatins, glutathione and cadmium in the phloem sap of Brassica napus. A role for thiol-peptides in the long-distance transport of cadmium and the effect of cadmium on iron translocation[J]. The Plant Journal, 2008, 54(2): 249-259. |
| [42] | Li L P, Zhang Y Q, Ippolito J A, et al. Cadmium foliar application affects wheat Cd, Cu, Pb and Zn accumulation[J]. Environmental Pollution, 2020, 262: 114329-114336. |
| [43] | Clemens S, Ma J F. Toxic heavy metal and metalloid accumulation in crop plants and foods[J]. Annual Review of Plant Biology, 2016, 67: 489-512. |
| [44] | Wang H, Ahan J, Wu Z H, et al. Alteration of nitrogen metabolism in rice variety 'Nipponbare' induced by alkali stress[J]. Plant and Soil, 2012, 355(1-2): 131-147. |
| [45] | Li H, Xu C Y, Han L, et al. Extensive secretion of phenolic acids and fatty acids facilitates rhizosphere pH regulation in halophyte Puccinellia tenuiflora under alkali stress[J]. Physiologia Plantarum, 2022, 174(2): 13678-13689. |