 2023, Vol. 44
2023, Vol. 44



2. 东北农业大学资源与环境学院, 哈尔滨 150030
2. College of Resources and Environment, Northeast Agricultural University, Harbin 150030, China
近年来, 我国Cd污染状况日益严重, 与1990年相比, 土壤Cd元素含量超过土壤背景值140.21%[1].通过对中国农田和城市土壤重金属污染的Meta分析表明, Cd是我国土壤中污染最严重的重金属元素, 分别占农田和城市土壤污染的33.54%和44.65%[2], Cd的生物富集因子较高[3], 土壤中的Cd被植物吸收后不仅会破坏植物的氧化还原平衡、减少植物营养吸收、损伤植物基因和降低作物产量[4], 还会通过食物链进入人体危及人类健康.
纳米氧化铜(CuO NPs)是较为常见的NPs之一, 具有巨磁阻效应、量子相干效应和库仑堵塞效应等性能[5], 2014年CuO NPs的使用量为570 t, 预计2025年使用量将增长至1 600 t[6].巨大的使用量增加了CuO NPs进入农田土壤环境的可能性.有研究证明, 低剂量下NPs会对植物产生刺激效应, 促进植物的生长发育, 提高植物对环境胁迫的耐受性[7], 当CuO NPs剂量过高时, 会抑制植物光合和生长、产生氧化损伤和降低营养价值, 甚至使植物DNA受损[8].
NPs进入农田后可能与原有的重金属污染产生协同或者拮抗效果.叶兴银等[9]研究表明添加纳米二氧化钛(TiO2 NPs)能显著降低土壤中Cd的生物有效性, 降幅与NPs的施加浓度成正比.与之相反, 王苗苗等[10]研究表明TiO2 NPs会增加小白菜体内Cd积累, 增强氧化应激, 抑制植物生长发育.Hussain等[11]发现叶面施用纳米硒和纳米硅会提高水稻的产量和品质, 降低糙米中的Cd和Pb含量.低剂量下纳米氧化锌(ZnO NPs)对植物无毒, 与Cd表现为拮抗作用, 当其含量增长至250 mg·kg-1和500 mg·kg-1时, ZnO NPs与Cd会产生协同抑制效果, 降低高粱的生物量[12].土壤处理和叶面喷施纳米氧化铁有效降低了Cd胁迫小麦的电解渗漏和籽粒积累, 同时提高了超氧化物歧化酶(SOD)、植物脯氨酸氧化酶(POX)活性和生长参数[13].叶面喷施纳米铁颗粒提高了Cd胁迫下植物的愈创木酚过氧化物酶(GP)、APX和SOD活性, 植物叶绿素含量显著增加[14].方清等[15]研究发现低浓度的CuO NPs可以缓解砷对水稻幼苗的毒害作用, 提高发芽率和生物量.这表明NPs、植物和重金属的种类、NPs的浓度变化、暴露培养条件、NPs的表面电荷和粒径等不同等都会导致NPs和重金属产生不同的联合毒性效果.目前有关纳米材料与重金属相互作用的研究较少, CuO NPs与Cd对植物的联合效果尚不明确.
小油菜(Brassica chinensis L.)在我国种植面积和种植范围广泛, 且对重金属Cd具有一定富集能力.本实验选用夏绿2号小油菜为研究对象, 研究CuO NPs与Cd单一和联合作用下小油菜的鲜重、光合色素含量、丙二醛含量、抗氧化酶活性和小油菜对Cd和Cu的吸收情况, 探讨CuO NPs对Cd胁迫下小油菜生理生化特性, 以期为评估CuO NPs的潜在环境风险提供一定的理论依据.
1 材料与方法 1.1 供试材料根据赵孟良等[16]的品种分析, 选定实验小油菜品种为夏绿2号, 供试小油菜种子购自京研益农种业科技有限公司.CuO NPs购于浙江艾绿化工科技有限公司, 粒径为40 nm, 球形, 纯度≥99.5%, 比表面积为2.84 m2·g-1.
1.2 实验设计采用水培实验的方式, 于农业农村部环境保护科研监测所温室内培育, 选取颗粒均匀且饱满的小油菜种子播种在育苗盘中, 在底部托盘中加入超纯水为种子萌发及生长提供充足水分, 每2 d向盘中加入600 mL超纯水, 保持培养基质湿润, 添加600 mL 1/4 Hoagland营养液、1/2 Hoagland营养液、全营养液分别培养4 d, 营养液配比为∶130.7 mg·L-1 K2SO4、7.5 mg·L-1 KCl、34.0 mg·L-1 KH2PO4、160.2 mg·L-1 MgSO4·H2O、472.3 mg·L-1 Ca(NO3)2·4H2O、0.6 mg·L-1 H3BO3、2.5×10-2mg·L-1 CuSO4·5H2O、0.3 mg·L-1 ZnSO4·7H2O、0.2 mg·L-1 MnSO4·H2O、6.2×10-3mg·L-1 (NH4)6Mo7O24·4H2O、27.8 mg·L-1 FeSO4·7H2O和37.2 mg·L-1 Na2EDTA·2H2O.本实验设置T1(CK)、T2(10 mg·L-1 CuO NPs)、T3(20 mg·L-1 CuO NPs)、T4(50 mg·L-1 CuO NPs)、T5(1 μmol·L-1 Cd)、T6(1 μmol·L-1 Cd+10 mg·L-1 CuO NPs)、T7(1 μmol·L-1 Cd+20 mg·L-1 CuO NPs)、T8(1 μmol·L-1 Cd+50 mg·L-1 CuO NPs)、T9(5 μmol·L-1 Cd)、T10(5 μmol·L-1 Cd+10 mg·L-1 CuO NPs)、T11(5 μmol·L-1 Cd+20 mg·L-1 CuO NPs)、T12(5 μmol·L-1 Cd+50 mg·L-1 CuO NPs)共12组处理, 每组处理设置3个平行.挑选5颗长势均匀的小油菜幼苗移至体积为1.5 L水培箱中, 每4 d更换一次营养液, 每天通气4 h.培养20 d后收取全部小油菜植株进行指标测定分析.
1.3 测定项目和方法 1.3.1 生物量测定分地上部和地下部收取小油菜样品, 超纯水清洗3次后用吸水纸吸干.使用分析天平称量并记录鲜样重量.将新鲜样品置于-80℃冷冻保存.小油菜样品于烘箱105℃杀青30 min后在80℃干燥72 h至恒重, 使用磨样机磨碎后于自封袋保存备用.
1.3.2 叶绿素含量测定以2∶1的比例混合丙酮和无水乙醇作为提取剂, 将提取剂和0.2 g小油菜叶片样品与少量CaCO3混合研磨后转移至25 mL容量瓶中, 加入提取剂至25 mL, 使用超声清洗器(50 Hz, 200 W, 25℃)清洗10 min, 静置后使用可见分光光度计测量665 nm和649 nm处的上清液吸光度, 通过公式计算光合色素的含量. Ca、Cb和Ct分别为叶绿素a、叶绿素b和总叶绿素含量(mg·g-1), D649和D665分别为649 nm和665 nm处测定的上清液吸光度.
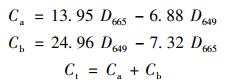
|
使用购自苏州格锐思生物科技有限公司的试剂盒测定小油菜丙二醛(MDA)含量以及过氧化氢酶(CAT)、过氧化物酶(POD)、超氧化物歧化酶(SOD)和谷胱甘肽还原酶(GR)活性.
1.3.4 小白菜Cu和Cd含量测定称取0.25 g小油菜干样加入8 mL硝酸放入消解炉(DigiBlock ED54, LabTech, 中国)中消解, 采用电感耦合等离子体质谱仪(ICP-MS, 美国)测定小白菜Cu和Cd含量.
1.3.5 小白菜亚细胞Cd和Cu含量测定亚细胞Cu和Cd含量测定参照Li等[17]的方法, 分别称取3 g小油菜鲜样于消解管和50 mL离心管中, 在离心管中加入30 mL预冷提取液, 提取液组成为0.25 mmol·L-1蔗糖、50 mmol·L-1(Tris-HCl)缓冲液和1 mmol·L-1二硫赤藓糖醇, pH为7.8.离心管置于涡旋机上以4℃ 2 000 r·min-1涡旋5 min, 滤布过滤, 底层滤渣为细胞壁组分, 将滤液以4℃ 11 900 r·min-1离心45 min, 上层清液为细胞基质组分, 底层固体为细胞器组分.3组组分烘干冷却后, 亚细胞Cu和Cd含量测定方法同1.3.4节.
1.4 数据处理与分析使用Excel对实验数据进行数据整理.使用IBM SPSS Statistics 26进行单因素方差分析(One-way ANOVA), 采用邓肯多重比较检验对实验数据进行差异显著性分析(P < 0.05), 用Origin 2021软件绘图.
2 结果与分析 2.1 CuO NPs对Cd胁迫下小油菜鲜重的影响从图 1可知, 不同Cd处理条件下CuO NPs对小油菜的鲜重影响不同.无Cd处理中, CuO NPs表现出对植物生长的抑制作用, 20 mg·L-1和50 mg·L-1 CuO NPs处理下油菜鲜重显著低于对照处理, 降幅分别为25.39%和28.62%. Cd浓度为1 μmol·L-1下施加20 mg·L-1 CuO NPs为小油菜生长的最适浓度, 小油菜叶片和根系鲜重分达到21.44 g和2.34 g.与对照相比, 50 mg·L-1 CuO NPs下仅根部鲜重显著上升, 增幅为35.73%, 表明在较低的Cd水平下, CuO NPs可以促进Cd胁迫下的植物生长发育.当Cd浓度达到5 μmol·L-1时, CuO NPs对小油菜的地上部生物量无显著影响.与对照相比, 20 mg·L-1 CuO NPs处理组小油菜根系鲜重显著提高14.11%. 与低Cd处理组相反, 5 μmol·L-1 Cd与50 mg·L-1 CuO NPs表现出协同毒性作用, 小油菜叶部和根部鲜重均达到最低值, 分别为12.36 g和1.56 g.这表明Cd与CuO NPs任一浓度发生变化时, 对植物产生的效果也不同.

|
不同大写字母表示同一Cd浓度处理下添加不同浓度CuO NPs之间差异显著(P < 0.05), 不同小写字母表示施加同一浓度CuO NPs添加不同Cd处理之间差异显著(P < 0.05), 下同 图 1 CuO NPs对Cd污染下小油菜鲜重的影响 Fig. 1 Effect of CuO NPs on fresh weight of Brassica chinensis L. under Cd stress |
本实验中CuO NPs对不同Cd浓度下的小油菜叶绿素含量均有促进作用(图 2), 无Cd处理中, CuO NPs浓度为20 mg·L-1下叶绿素含量达到最高值, 分别为1.03、0.61和1.61 mg·g-1, 当浓度升高至50 mg·L-1, 小油菜的叶绿素含量下降, 表明当浓度持续升高时, CuO NPs对小油菜叶绿素的促进作用开始减弱. 1 μmol·L-1 Cd+10 mg·L-1 CuO NPs处理组中叶绿素b显著升高23.38%. 5 μmol·L-1 Cd处理组中, 小油菜的叶绿素含量均显著上升, 叶绿素a含量涨幅为22.80% ~36.36%, 叶绿素b含量涨幅为25.95% ~30.15%, 总叶绿素含量涨幅为23.98% ~34.04%. 这说明CuO NPs对植物生长的负面作用并非源于NPs可能造成的植物光合系统损伤.

|
图 2 CuO NPs对Cd污染下小油菜叶绿素的影响 Fig. 2 Effect of CuO NPs on chlorophyll of Brassica chinensis L. under Cd stress |
图 3表明, 小油菜的叶部和根部MDA含量均随加入CuO NPs的剂量增加而上升, 无Cd处理下, CuO NPs浓度为50 mg·L-1时, 小油菜叶部和根部MDA含量均显著升高, 分别增加24.92%和47.62%, 显示出CuO NPs诱导植物氧化应激, 增加植物的细胞膜损伤.1 μmol·L-1 Cd处理下, CuO NPs均提高了小油菜的MDA含量, 小油菜地上部MDA含量分别增加36.27%、21.38%和7.78%, 根部丙二醛含量增幅分别是对照的1.49、1.69和2.31倍.5 μmol·L-1 Cd+20 mg·L-1 CuO NPs处理下叶部MDA含量降低7.84%, 但影响不显著, 根部下降24.12%, 其他处理MDA含量同样呈现增长趋势.从总体上看, 除5 μmol·L-1 Cd+20 mg·L-1 CuO NPs处理MDA含量出现小幅降低, CuO NPs均提高了Cd胁迫下小油菜的细胞膜脂质过氧化程度, 增大了小油菜氧化损伤.

|
图 3 CuO NPs对Cd污染下小油菜MDA含量的影响 Fig. 3 Effect of CuO NPs on MDA content of Brassica chinensis L. under Cd stress |
仅CuO NPs暴露处理中,小油菜叶部的CAT和GR活性均显著降低, POD活性在CuO NPs浓度为50 mg·L-1时显著下降(P < 0.05).油菜叶部SOD活性总体呈现先升高后降低的趋势(图 4), 与对照相比分别上升51.16%、22.56%和8.29%, 说明CuO NPs对植物产生氧化损伤, 小油菜SOD活性升高以清除活性氧(ROS). 1 μmol·L-1 Cd处理下, CuO NPs显著抑制了CAT活性, CAT活性分别下降12.16%、32.89%和11.47%; 10 mg·L-1和20 mg·L-1CuO NPs处理下小油菜叶部GR活性显著降低24.70%和29.09%. 1 μmol·L-1 Cd+50 mg·L-1 CuO NPs处理组POD活性显著提高20.71%, 说明二者产生的联合胁迫对大多数抗氧化酶均有抑制效果.5 μmol·L-1 Cd处理下, CuO NPs显著抑制SOD和GR活性, SOD和GR降幅较对照分别为11.66% ~34.25%和16.40% ~19.55%; 相反, 叶部POD活性则显著上升4.87% ~20.71%; CuO NPs浓度为10 mg·L-1和20 mg·L-1时CAT活性显著提高, 分别为对照组的1.9倍和9.09倍, 50 mg·L-1 CuO NPs处理下CAT活性显著降低44.15%. Cd浓度升高后酶活变化幅度远高于1 μmol·L-1 Cd处理组, 说明产生了更多的ROS, 而此时CAT和POD为清除小油菜体内ROS的主要酶.总体来说, CuO NPs激活了小油菜的抗氧化酶防御体系, 但没有缓解小油菜的氧化胁迫.

|
图 4 CuO NPs对Cd污染下小油菜叶部抗氧化酶活性的影响 Fig. 4 Effect of CuO NPs on the activities of antioxidant enzymes in Brassica chinensis L. leaves under Cd stress |
小油菜叶部和根部中Cu含量均随CuO NPs施加水平的升高而增加[图 5(a)和5(b)], 无Cd处理中小油菜叶部Cu含量分别是对照组的4.19、6.33和11.21倍.1 μmol·L-1 Cd处理下, 与对照组相比, 分别增加56.04%、196.80%和335.88%. 5 μmol·L-1 Cd处理下, 分别是对照组的2.49、4.55和7.08倍.对比相同CuO NPs浓度的处理组发现, Cd浓度升高, 小油菜吸收的Cu元素含量降低, 说明Cd对小油菜中Cu的吸收可能存在一定的拮抗作用.小油菜根部Cu元素积累量远高于叶部, 无Cd处理中Cu元素增加28.42、29.68和67.67倍, 低Cd处理组Cu含量增加6.99、7.91和16.82倍, 5 μmol·L-1 Cd处理下增加了8.75、9.35和20.56倍.说明小油菜吸收的Cu元素多滞留在根系中, 仅有少量转运至地上部.

|
(a)叶部Cu含量, (b)根部Cu含量, (c)不同处理下叶部亚细胞各组分Cu含量, (d)不同处理下叶部亚细胞中Cu质量分数 图 5 CuO NPs对Cd污染下小油菜体内Cu含量的影响 Fig. 5 Effect of CuO NPs on Cu content in Brassica chinensis L.under Cd stress |
CuO NPs施加后增加了小油菜亚细胞的Cu元素含量[图 5(c)].同一CuO NPs浓度下溶液Cd含量越高, 亚细胞组分中Cu元素含量越高, 显示Cd添加增加了细胞损伤, 致使更多的Cu元素进入小油菜亚细胞.图 5(d)显示对照处理中Cu多分布在细胞壁中, CuO NPs加入后, 小油菜亚细胞可溶性组分中Cu质量分数上升, 细胞器中质量分数下降, 说明NPs对细胞膜结构造成破坏, Cu元素进入小油菜细胞后主要被储存在可溶性组分中.
2.3.2 器官和亚细胞中Cd含量CuO NPs抑制了小油菜对Cd的吸收[图 6(a)和6(b)].Cd浓度为1 μmol·L-1下施加10 mg·L-1和20 mg·L-1 CuO NPs后小油菜叶部Cd含量分别较空白处理显著降低了45.64%和25.49%, 当CuO NPs浓度增至50 mg·L-1, 小油菜Cd含量上升6.26%, 但根部Cd含量均显著降低(P < 0.05), 与单一Cd处理相比分别下降32.42%、20.03%和33.39%. 这表明, CuO NPs在较低浓度下可以抑制小油菜的Cd吸收, 但当浓度升高时, 二者可能表现出联合效果, 增加小油菜的Cd吸收.5 μmol·L-1 Cd处理下, 添加CuO NPs后小油菜的Cd含量均有不同程度下降, 地上部降幅分别为7.26%、18.25%和10.03%, 根部降幅分别为25.35%、11.80%和3.66%..

|
(a)叶部Cd含量, (b)根部Cd含量, (c)不同处理下叶部亚细胞各组分Cd含量, (d)不同处理下叶部亚细胞Cd质量分数 图 6 CuO NPs对Cd污染下小油菜体内Cd含量的影响 Fig. 6 Effect of CuO NPs on Cd content in Brassica chinensis L. under Cd stress |
小油菜叶部亚细胞中Cd含量见图 6(c).5 μmol·L-1Cd处理下施加10 mg·L-1和20 mg·L-1 CuO NPs后小油菜叶片细胞壁内的Cd含量降幅为26.32%和29.89%. 与之相反, CuO NPs浓度为50 mg·L-1时小油菜细胞壁Cd含量分别升高89.10%和1.19%. 施加CuO NPs后, 细胞可溶性组分中Cd质量分数大幅升高, 细胞器中则相应降低[图 6(d)].总的来说, 低浓度CuO NPs处理降低了小油菜亚细胞中的Cd含量, 当浓度达到50 mg·L-1时促进小油菜亚细胞Cd吸收.
3 讨论鲜重是衡量植物生长情况的基本指标, CuO NPs处理降低了小油菜鲜重, 这与Nair等[18]研究的结果一致. CuO NPs以剂量依赖性方式抑制印度芥菜根和芽的生长, Trujillo-Reyes等[19]研究表明10 mg·L-1、20 mg·L-1 CuO NPs使莴苣的根系和叶片生物量分别减少66%、69%和54%、52%. Cu基NPs对植物的胁迫分为两种方式, 一种是铜离子或铜以颗粒形式或纳米形式释放, 被植物吸收造成毒性影响, 另一种是通过诱导ROS等方式造成植物细胞损伤, 干扰植物的生长代谢[20].CuO NPs在营养液中释放出的铜离子会导致激素失衡, 对植物根系结构造成影响, 生长素浓度的变化或根系细胞的过度木质化会导致植物根系生长迟缓, 减少植物矿质营养元素的可用性[21, 22], 降低小油菜的生物量.低Cd浓度下, CuO NPs对植物的生长表现出促进作用, 20 mg·L-1CuO NPs处理为小油菜生长的最适浓度.有研究显示在低浓度比例下, TiO2 NPs与Cd表现出拮抗联合毒性效应, 而在较高比例共同暴露时表现出联合毒性效应[23].本实验中, 二者共同暴露下CuO NPs降低了小油菜地上部和地下部的Cd含量, 这与小油菜生物量变化趋势相符, 一方面, NPs可以通过物理和化学在表面上吸附金属离子, 降低营养液中游离的Cd2+浓度, 缓解Cd对植物的毒性.在CuO-NP和Cd2+的二元体系中, pH为7时CuO NPs对Cd离子的去除效率约为20%[24].Subpiramaniyam等[25]认为Cu和Cd元素之间存在拮抗效应, 竞争会降低它们的生态毒性.不同的金属可能在细胞膜上共享相同的转运蛋白, 植物不能清楚区分重要的微量营养素和非必需金属[26], 而植物吸收微量营养素的速度远快于非必需重金属[27].Cd和Cu是具有相同电子价的金属元素, 因此存在竞争位点, Cu对Cd元素的吸收和转移具有一定的抑制作用, 这会降低植物的Cd含量.随CuO NPs浓度升高, 与Cd的拮抗作用转变为协同作用, 植物中的重金属含量越高, 对生物分子的氧化损伤越严重, 当受胁迫植物中两种金属元素含量较高时, 植物对这两种元素的吸收将表现出一定的加性效应.
叶绿素含量会反映出胁迫环境对植物光合作用的影响.金属纳米粒子可以促进植物的光合作用, El-Shazoly等[28]发现50 mg·L-1 CuO NPs提高了红花的叶绿素b和类胡萝卜素含量.本实验中, 小油菜叶绿素含量随CuO NPs浓度增加呈现出上涨的趋势, 这表明CuO NPs的毒性效应可能并非来自植物吸收转运至地上部的铜离子, 更多的可能来自于其尺寸效应带来的氧化损伤.Wang等[29]研究证明在pH为7.5时, 蒸馏水中100 mg·L-1 CuO NPs仅释放出了约0.17 mg·L-1 Cu2+.Shahane等[30]研究认为来自蔬菜可食用部分的CuO NPs溶解部分产生的风险比未溶解部分的CuO NPs小93倍, 植物光合作用对Cd高度敏感, 低浓度Cd处理会对叶绿素合成起促进作用, 高浓度Cd会使植物叶绿素合成受阻, 叶绿体结构受损, 影响植物光合, CuO NPs降低小油菜的Cd吸收同样会使小油菜的叶绿素含量增加.
ROS是氧化代谢的有害产物, DNA损伤、脂质过氧化、蛋白质降解和膜损伤是活性氧诱导的氧化应激的常见特征.MDA含量可以反映出植物脂质过氧化和植物细胞损伤的程度.暴露在CuO NPs的植物叶片和根部MDA含量呈浓度依赖性增加, 说明CuO NPs浓度越大, 植物氧化损伤越严重, 已有多篇研究报道过类似现象[18, 31].重金属胁迫与NPs都会造成植物损伤, 增加植物的MDA含量, 除5 μmol·L-1 Cd+20 mg·L-1 CuO NPs处理由于小油菜中总Cd含量下降最多, MDA含量小幅降低以外, Cd胁迫下CuO NPs同样增加了植物的MDA含量, 表明尽管其他处理中Cd含量同样降低, 但不足以抵消CuO NPs带来的氧化胁迫.CAT、POD、SOD和GR等抗氧化酶可以帮助植物抵御外来胁迫带来的ROS积累.CuO NPs增强了小油菜叶部氧化胁迫, SOD活性增高, 将超氧自由基转化为O2和H2O2, 1 μmol·L-1和5 μmol·L-1 Cd处理下POD活性均有增高, 表明活性氧产生量增加, 植物抵抗氧化应激损伤的防御机制快速激活; 5 μmol·L-1 Cd处理下, CAT呈现先升高后降低的趋势, CAT用于催化H2O2分解为氧气和水[32], 表明植物体内H2O2快速升高, CAT活性提高以抵御CuO NPs带来的ROS损伤, 而浓度上升后超过了抗氧化酶的催化能力, 抗氧化酶活性受到抑制.部分抗氧化酶活性随CuO NPs浓度升高而降低, 一方面可能由于CuO NPs的氧化状态远比其他NPs毒性更大, 胁迫植物产生的H2O2浓度过高, 抑制了抗氧化酶活性, 另一方面可能由于CuO NPs破坏了植物结构, 抑制植物对养分的吸收, 植物吸收的营养不足以支持植物的正常代谢, 无法顺利合成蛋白质, 从而抑制了抗氧化酶的合成.
CuO NPs的吸附和摄取都会随着CuO NPs暴露浓度的增加而增加[33], 随加入CuO NPs浓度增加, 小油菜叶片和根系Cu含量均大幅升高.植物根系分泌的大量的有机物、代谢物甚至H+, 会降低营养液的pH值[34], 增加营养液中游离铜离子的生物利用度.CuO NPs可以吸附在植物根系表面, 经过晶体溶解、生物转化和生物积累通过植物组织进行分配[35].NPs进入植物的外部保护层后, 可以通过质外体或共质体两种途径运输, 水和溶解矿物质通过细胞壁和质膜外的细胞外区域运动为质外体运输, 而共质体途径则通过相邻细胞的胞间连丝连接的细胞质运输水和溶质[36].由于植物具有排斥机制, 阻止过多的重金属运输到地上部, 以维持组织和器官中矿物质元素的动态平衡, 小油菜根中积累的Cu和Cd元素远比叶片中积累的多.添加Cd后小油菜体内的Cu含量出现明显下降, NPs表面吸附的金属离子可通过压缩核动力源的双电层来增强NPs聚集, 增强的NPs聚集可进一步减少可用表面积, 这可能导致NPs溶解的减少[37], Cu和Cd拮抗作用同样会减少吸收量.小油菜亚细胞中Cu元素主要分布在细胞壁中, 植物细胞壁是重金属进入植物的第一屏障, 在存在团聚的情况下, 直径大于20 nm的纳米颗粒几乎不能穿透细胞壁[38], 亚细胞中细胞壁中的Cu元素质量分数下降, 可溶性组分中质量分数上升, 说明NPs暴露产生ROS或其他影响, 可能减少细胞壁厚度、触发细胞壁松动或改变细胞壁孔径[39], 致使更多NPs进入亚细胞.与宋阿琳等[40]研究的结果一致, Cd元素主要分布在细胞可溶性组分中, 植物会避免将摄入的金属元素储存在较为重要的细胞器中, 而植物的可溶性组分主要由质外体中的水分和液泡细胞器之间的溶胶状基质组成, 液泡内含有有机碱、有机酸和蛋白质等可与重金属结合的物质, 将重金属转移至可溶性组分中, 可以减少细胞损伤, 这是植物抵御重金属中毒并参与重金属解毒的重要机制[41].CuO NPs降低了植物细胞器中的Cu和Cd质量分数, 增加了可溶性组分中的Cu和Cd质量分数, 减少了重金属对植物细胞的破坏作用.
4 结论(1) 低浓度CuO NPs能够缓解Cd对小油菜根系发育的胁迫, 促进Cd污染下小油菜的生长, 提高生物量; 高浓度(50 mg·L-1)下表现出协同毒性作用, 降低了作物生物量.
(2) CuO NPs提高了小油菜的光合色素含量, 但增强了Cd处理后的小油菜的氧化损伤, CuO NPs造成的毒性作用主要来自未溶解部分带来的氧化胁迫.
(3) 添加CuO NPs增加了小油菜的Cu元素含量, 而抑制了Cd吸收; 细胞器中Cu和Cd质量分数降低, 而可溶性组分重金属则有所增加, 缓解了重金属Cd对小油菜的毒性作用.
| [1] | Ren S Y, Song C Q, Ye S J, et al. The spatiotemporal variation in heavy metals in China's farmland soil over the past 20 years: A meta-analysis[J]. Science of the Total Environment, 2022, 806. DOI:10.1016/j.scitotenv.2021.150322 |
| [2] | Yuan X H, Xue N D, Han Z G. A meta-analysis of heavy metals pollution in farmland and urban soils in China over the past 20 years[J]. Journal of Environmental Sciences, 2021, 101: 217-226. DOI:10.1016/j.jes.2020.08.013 |
| [3] | Hu B F, Xue J, Zhou Y, et al. Modelling bioaccumulation of heavy metals in soil-crop ecosystems and identifying its controlling factors using machine learning[J]. Environmental Pollution, 2020, 262. DOI:10.1016/j.envpol.2020.114308 |
| [4] |
徐佳慧, 王萌, 张润, 等. 土壤镉污染的生物毒性研究进展[J]. 生态毒理学报, 2020, 15(5): 82-91. Xu J H, Wang M, Zhang R, et al. Toxicity of cadmium pollution in soil to organisms: A Review[J]. Asian Journal of Ecotoxicology, 2020, 15(5): 82-91. |
| [5] |
常敬先, 吴娜娜, 闫四海. 氧化铜纳米材料制备工艺技术及应用研究进展[J]. 信息记录材料, 2021, 22(11): 8-9. Chang J X, Wu N N, Yan S H, et al. Research progress on preparation technology and application of CuO nanomaterials[J]. Information Recording Materials, 2021, 22(11): 8-9. |
| [6] | Zhao S Y, Su X X, Wang Y Y, et al. Copper oxide nanoparticles inhibited denitrifying enzymes and electron transport system activities to influence soil denitrification and N2O emission[J]. Chemosphere, 2020, 245. DOI:10.1016/j.chemosphere.2019.125394 |
| [7] |
马晓琳, 何恩静, 肖翔. 纳米材料的非毒性环境效应研究进展[J]. 生态毒理学报, 2021, 16(1): 15-27. Ma X L, He E J, Xiao X. Research progress on non-toxic environmental effects of nanomaterials[J]. Asian Journal of Ecotoxicology, 2021, 16(1): 15-27. |
| [8] | Dimkpa C O, McLean J E, Britt D W, et al. Nano-CuO and interaction with nano-ZnO or soil bacterium provide evidence for the interference of nanoparticles in metal nutrition of plants[J]. Ecotoxicology, 2015, 24(1): 119-129. DOI:10.1007/s10646-014-1364-x |
| [9] |
叶兴银, 张卫, 龙精华, 等. 人工纳米颗粒输入对稻田土壤Cd形态转化及生物有效性的影响[J]. 环境工程学报, 2018, 12(12): 3426-3432. Ye X Y, Zhang W, Long J H, et al. Impact of engineered nanoperticles on Cd chemical speciation and bioavailability in paddy soil[J]. Chinese Journal of Environmental Engineering, 2018, 12(12): 3426-3432. DOI:10.12030/j.cjee.201805142 |
| [10] |
王苗苗, 强沥文, 王伟, 等. 纳米二氧化钛对镉胁迫下小白菜毒性效应的影响[J]. 农业环境科学学报, 2020, 39(6): 1185-1195. Wang M M, Qiang L W, Wang W, et al. Effects of Nano titanium dioxide on the toxicity of Chinese cabbage under cadmium stress[J]. Journal of Agro-Environment Science, 2020, 39(6): 1185-1195. |
| [11] | Hussain B, Lin Q, Hamid Y, et al. Foliage application of selenium and silicon nanoparticles alleviates Cd and Pb toxicity in rice (Oryza sativa L.)[J]. Science of the Total Environment, 2020, 712. DOI:10.1016/j.scitotenv.2020.136497 |
| [12] | Wang F Y, Adams C A, Shi Z Y, et al. Combined effects of ZnO NPs and Cd on sweet sorghum as influenced by an arbuscular mycorrhizal fungus[J]. Chemosphere, 2018, 209: 421-429. DOI:10.1016/j.chemosphere.2018.06.099 |
| [13] | Hussain A, Ali S, Rizwan M, et al. Responses of wheat (Triticum aestivum) plants grown in a Cd contaminated soil to the application of iron oxide nanoparticles[J]. Ecotoxicology and Environmental Safety, 2019, 173: 156-164. DOI:10.1016/j.ecoenv.2019.01.118 |
| [14] | Akandi Z N, Makarian H, Pirdashti H, et al. Iron nanoparticles-induced improvement of antioxidant enzymes and photosynthetic pigments of purslane (Portulaca oleracea L.) Under Cadmium Stress[J]. Gesunde Pflanzen, 2022, 74(4): 829-838. DOI:10.1007/s10343-022-00680-9 |
| [15] |
方清, 晏士玮, 崔彩云, 等. 纳米氧化铜对砷胁迫下水稻种子发芽及幼苗生长的影响[J]. 安徽农业大学学报, 2020, 47(5): 826-831. Fang Q, Yan S W, Cui C Y, et al. Effects of copper oxide nanoparticles on germination and growth of rice seeds exposed to arsenic stress[J]. Journal of Anhui Agricultural University, 2020, 47(5): 826-831. |
| [16] |
赵孟良, 韩睿, 王紫莹, 等. 20份小白菜品种引种试验农艺性状的多样性分析[J]. 南方农业学报, 2020, 51(8): 1960-1969. Zhao M L, Han R, Wang Z Y, et al. Diversity of agronomic characters of 20 pakchoi varieties in introduction experiment[J]. Journal of Southern Agriculture, 2020, 51(8): 1960-1969. DOI:10.3969/j.issn.2095-1191.2020.08.022 |
| [17] | Li H, Luo N, Zhang L J, et al. Do arbuscular mycorrhizal fungi affect cadmium uptake kinetics, subcellular distribution and chemical forms in rice?[J]. Science of the Total Environment, 2016, 571: 1183-1190. DOI:10.1016/j.scitotenv.2016.07.124 |
| [18] | Nair P M G, Chung I M. Study on the correlation between copper oxide nanoparticles induced growth suppression and enhanced lignification in Indian mustard (Brassica juncea L.)[J]. Ecotoxicology and Environmental Safety, 2015, 113: 302-313. DOI:10.1016/j.ecoenv.2014.12.013 |
| [19] | Trujillo-Reyes J, Majumdar S, Botez C E, et al. Exposure studies of core-shell Fe/Fe3O4 and Cu/CuO NPs to lettuce (Lactuca sativa) plants: Are they a potential physiological and nutritional hazard?[J]. Journal of Hazardous Materials, 2014, 267: 255-263. DOI:10.1016/j.jhazmat.2013.11.067 |
| [20] |
张海, 彭程, 杨建军, 等. 金属型纳米颗粒对植物的生态毒理效应研究进展[J]. 应用生态学报, 2013, 24(3): 885-892. Zhang H, Peng C, Yang J J, et al. Eco-toxicological effect of metal-based nanoparticles on plants: Research progress[J]. Chinese Journal of Applied Ecology, 2013, 24(3): 885-892. |
| [21] | Nair P M G, Kim S H, Chung I M. Copper oxide nanoparticle toxicity in mung bean (Vigna radiata L.) seedlings: physiological and molecular level responses of in vitro grown plants[J]. Acta Physiologiae Plantarum, 2014, 36(11): 2947-2958. DOI:10.1007/s11738-014-1667-9 |
| [22] | Lequeux H, Hermans C, Lutts S, et al. Response to copper excess in Arabidopsis thaliana: Impact on the root system architecture, hormone distribution, lignin accumulation and mineral profile[J]. Plant Physiology and Biochemistry, 2010, 48(8): 673-682. |
| [23] | Wang P, Zhao L H, Huang Y X, et al. Combined toxicity of nano-TiO2 and Cd2+ to Scenedesmus obliquus: Effects at different concentration ratios[J]. Journal of Hazardous Materials, 2021, 418. DOI:10.1016/j.jhazmat.2021.126354 |
| [24] | Xiao Y L, Tang W, Peijnenburg W J G M, et al. Aggregation, solubility and cadmium-adsorption capacity of CuO nanoparticles in aquatic environments: Effects of pH, natural organic matter and component addition sequence[J]. Journal of Environmental Management, 2022, 310. DOI:10.1016/j.jenvman.2022.114770 |
| [25] | Subpiramaniyam S, Hong S C, Yi P I, et al. Influence of sawdust addition on the toxic effects of cadmium and copper oxide nanoparticles on Vigna radiata seeds[J]. Environmental Pollution, 2021, 289. DOI:10.1016/j.envpol.2021.117311 |
| [26] | Verkleij J A C, Golan-Goldhirsh A, Antosiewisz D M, et al. Dualities in plant tolerance to pollutants and their uptake and translocation to the upper plant parts[J]. Environmental and Experimental Botany, 2009, 67(1): 10-22. |
| [27] | Mishra V K, Tripathi B D. Concurrent removal and accumulation of heavy metals by the three aquatic macrophytes[J]. Bioresource Technology, 2008, 99(15): 7091-7097. |
| [28] | El-Shazoly R M. Evaluation of the effectiveness of safflower (Carthamus tinctorius) seeds amendment with silicon or humic acid during germination under CuO (NPs) phytotoxicity[J]. Egyptian Journal of Botany, 2019, 59(2): 357-374. |
| [29] | Wang L Y, Wang M, Peng C S, et al. Toxic effects of Nano-CuO, micro-CuO and Cu2+ on Chlorella sp.[J]. Journal of Environmental Protection, 2013, 4(1B): 86-91. |
| [30] | Shahane S P, Kumar A. Estimation of health risks due to copper-based nanoagrochemicals[J]. Environmental Science and Pollution Research, 2022, 29(17): 25046-25059. |
| [31] | Wang W, Ren Y F, He J Y, et al. Impact of copper oxide nanoparticles on the germination, seedling growth, and physiological responses in Brassica pekinensis L.[J]. Environmental Science and Pollution Research, 2020, 27(25): 31505-31515. |
| [32] | Willekens H, Chamnongpol S, Davey M, et al. Catalase is a sink for H2O2 and is indispensable for stress defence in C3 plants[J]. The EMBO Journal, 1997, 16(16): 4806-4816. |
| [33] | Deng C Y, Wang Y, Cota-Ruiz K, et al. Bok choy (Brassica rapa) grown in copper oxide nanoparticles-amended soils exhibits toxicity in a phenotype-dependent manner: Translocation, biodistribution and nutritional disturbance[J]. Journal of Hazardous Materials, 2020, 398. DOI:10.1016/j.jhazmat.2020.122978 |
| [34] | Dakora F D, Phillips D A. Root exudates as mediators of mineral acquisition in low-nutrient environments[J]. Plant and Soil, 2002, 245(1): 35-47. |
| [35] | Rajput V, Minkina T, Mazarji M, et al. Accumulation of nanoparticles in the soil-plant systems and their effects on human health[J]. Annals of Agricultural Sciences, 2020, 65(2): 137-143. |
| [36] | Murali M, Gowtham H G, Singh S B, et al. Fate, bioaccumulation and toxicity of engineered nanomaterials in plants: Current challenges and future prospects[J]. Science of the Total Environment, 2022, 811. DOI:10.1016/j.scitotenv.2021.152249 |
| [37] | Arenas-Lago D, Monikh F A, Vijver M G, et al. Dissolution and aggregation kinetics of zero valent copper nanoparticles in (simulated) natural surface waters: Simultaneous effects of pH, NOM and ionic strength[J]. Chemosphere, 2019, 226: 841-850. |
| [38] | Ma C X, White J C, Dhankher O P, et al. Metal-based nanotoxicity and detoxification pathways in higher plants[J]. Environmental Science & Technology, 2015, 49(12): 7109-7122. |
| [39] | Kim J H, Lee Y, Kim E J, et al. Exposure of iron nanoparticles to Arabidopsis thaliana enhances root elongation by triggering cell wall loosening[J]. Environmental Science & Technology, 2014, 48(6): 3477-3485. |
| [40] |
宋阿琳, 李萍, 李兆君, 等. 镉胁迫下两种不同小白菜的生长、镉吸收及其亚细胞分布特征[J]. 环境化学, 2011, 30(6): 1075-1080. Song A L, Li P, Li Z J, et al. Characterization of plant growth, cadmium uptake and its subcellular distribution in pakchoi exposed to cadmium stress[J]. Environmental Chemistry, 2011, 30(6): 1075-1080. |
| [41] |
李春烨, 丁国华, 刘保东. 重金属影响植物细胞超微结构和功能的研究进展[J]. 中国农学通报, 2013, 29(18): 114-118. Li C Y, Ding G H, Liu B D. Research progress of heavy metal affecting plant cell ultrastructure and function[J]. Chinese Agricultural Science Bulletin, 2013, 29(18): 114-118. |