 2023, Vol. 44
2023, Vol. 44

2. 中国科学院雄安创新研究院, 雄安 071899;
3. 广州大学大湾区环境研究院, 珠江三角州水质安全与保护教育部重点实验室, 广州 510006;
4. 澳大利亚联邦科学与工业组织, 肯辛顿 6151
 , MA Chao1,2 , YAN Jia-chen3 , LI Shu-yuan1,2 , LI Qing1 , HU Zi-ru1 , HU Qing2,4 , LIU Shuo1
, MA Chao1,2 , YAN Jia-chen3 , LI Shu-yuan1,2 , LI Qing1 , HU Zi-ru1 , HU Qing2,4 , LIU Shuo1 , WAN Jing-min1
, WAN Jing-min1
2. Xiong'an Innovation Institute, Chinese Academy of Sciences, Xiong'an 071899, China;
3. Key Laboratory for Water Quality and Conservation of the Pearl River Delta, Ministry of Education, Institute of Environmental Research at Greater Bay Area, Guangzhou University, Guangzhou 510006, China;
4. Commonwealth Scientific and Industrial Research Organisation (CSIRO), Kensington 6151, Australia
我国的北方干旱少雨, 多数地表河流补水主要以城市尾水为主[1].尾水中氮类指标的含量较高, 常会引起地表水体富营养化甚至黑臭现象[2].微藻是光合利用度高的初级生产者, 可以利用空气中的CO2为碳源进行光合作用, 在生长过程中吸收污水中的氮磷等物质来合成自身所需物质[3].微藻处理废水技术被认为是经济实用, 有发展潜力的城市尾水深度处理技术[4]. Shi等[5]研究发现小球藻(Chlorella vulgaris)去除污水中无机N的效率达86%.有研究利用微藻在光生物反应器中进行尾水处理, 培养14 d后发现, 硝酸盐的去除率最高可达79.63%[6], 微藻具有对氨氮(NH4+)、亚硝态氮(NO2-)和硝态氮(NO3-)还有总氮(TN)的去除功能[7].但是藻类过量繁殖, 并形成聚集体, 漂浮在水面, 降低水体透明度, 使深层水生植物光合作用受到限制, 导致水中溶解氧来源减少[8].
常用的物理除藻方法有直接打捞、过滤除藻、气浮除藻和超声除藻等, 这类方法见效快, 除藻过程中不易产生二次污染, 可此类方法工作量大, 成本高, 仅适用于藻浓度较高的小片水域作为应急手段[9]. 化学控藻通过向水中投加矿物质或化学药品来沉淀、抑制或杀死藻细胞, 这些化学方法简便易行, 效果显著[10], 但容易产生二次污染.生物控藻是通过合理调节水生食物链, 利用竞争、捕食关系和生物间的相互作用来降低水华生物密度, 从而防止水华的发生或消除水华[11].微生物是一切食物链的组成成分, 在水华水体中与有害藻类的存亡密切相关.有些微生物既能够分解有机物, 为藻类的生长提供物质基础, 又可以抑制藻类的生长, 溶解和杀死藻细胞[12].芽孢杆菌是能形成芽孢的杆菌, 在多种介质中有能有效抵抗或消除某些有害因子的功能[13].芽孢杆菌还具有一定的生物修复功能, 可以减少水质中的含氮污染物, 有效改善环境与水质[14].枯草芽孢杆菌是一种分布广泛非致病性的革兰氏阳性细菌, 其在水体中可抑制某些有害病原菌的生长与繁殖, 还具有还原硝酸盐等功能, 可作为微生物调节剂[15].芽孢杆菌在水华蓝藻治理上存在巨大潜力, 已经引起广大研究者的关注, 张睿[16]研究发现枯草芽孢杆菌对铜绿微囊藻有很强的抑制作用, 同时能够吸附水体中的重金属等污染物, 净化水质.芽孢杆菌用于控藻的优越性首先体现“生物安全”, 芽孢杆菌是一种非致命性的益生菌, 并在水产养殖和植物病害防治方面已取得许多成功案例, 其次能够分泌抗菌物质, 短芽孢杆菌能够产生短杆菌肽和几丁质酶等活性物质, 从而拮抗多种细菌和真菌[17].
在藻菌共生体系中, 微藻可以产生细菌吸收利用的有机质, 然而微藻和细菌之间的关系非常复杂.本研究通过分析芽孢杆菌和微藻系统微生物菌群特征, 以期为城市尾水及富营养化水体深度脱氮提供数据支持.
1 材料与方法 1.1 实验材料 1.1.1 实验菌株微藻培养液、芽孢杆菌为实验筛选保存.芽孢杆菌培养液(菌活数)2.21×108 CFU·mL-1, 微藻培养至对数期备用.
1.1.2 实验用水本文模拟城市尾水进行实验, 模拟尾水指标参照《城镇污水处理厂污染物排放标准》(GB 18918-2002)中一级标准的B标准配置, 水质指标如表 1所示.6月在某城镇污水处理厂排放口取回3 t尾水, 在室外非封闭水桶中放置2个月, 尾水出现富营养化现象, 为城市尾水实验用水, 水质指标如表 2所示.
|
|
表 1 模拟尾水水质指标/mg·L-1 Table 1 Water quality index of simulated tail water/mg·L-1 |
|
|
表 2 城市尾水水质指标/mg·L-1 Table 2 Water quality index of black and odorous water/mg·L-1 |
1.2 实验方法
本实验设置微藻组(Z)、芽孢杆菌组(Y)和添加微藻培养液和芽孢杆菌藻芽组(ZY).水样中NH4+指标的测定方法采用《水质NH4+的测定纳氏试剂分光光度法》(HJ 535-2009)标准, 实验选用20 mm比色皿, 其测定检出限为0.025mg·L-1, 测定范围为0.10~2.0 mg·L-1.本实验纳氏试剂的配制选择的是碘化汞-碘化钾-氢氧化钠溶液[18].水样中NO2-指标的测定方法采用《水质亚硝酸盐氮的测定分光光度法》(GB 7493-87)标准, 实验选用10 mm比色皿, 最低检出限0.003mg·L-1, 最高可测定浓度可以高达0.20 mg·L-1[19].水样中NO3-指标的测定方法采用《水质硝酸盐氮的测定紫外分光光度法(试行)》(HJ/T 346-2007)标准, 利用硝酸根离子在220 nm波长处的吸收而定量测定硝酸盐氮, 在275 nm处做另一次测量, 以校正硝酸盐氮值, 方法最低检出质量浓度为0.08mg·L-1, 测定下限为0.32 mg·L-1, 测定上限为4 mg·L-1[20].水样中TN指标的测定方法采用《水质TN的测定碱性过硫酸钾消解紫外分光光度法》(HJ 636-2012)标准, 采用紫外分光光度法于波长220 nm和275 nm处, 分别测定吸光度A220和A275, 按公式(1)计算校正吸光度A, TN(以N计)浓度与校正吸光度A成正比[21].本方法的检出限为0.05 mg·L-1, 测定范围为0.20~7.00 mg·L-1.
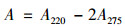
|
(1) |
将微藻培养液与芽孢杆菌培养液等比例以2%投加量加入实验城市尾水中, 由于培养基中含有部分氮素, 实验过程中会导致氮素值高于未添加培养液的进水.
2 结果与分析 2.1 模拟尾水中氮代谢分析Z、Y和ZY在模拟尾水中氮代谢特征如图 1所示.

|
图 1 模拟尾水中氮代谢分析 Fig. 1 Analysis of nitrogen metabolism in simulated tail water |
在模拟尾水中, Z中NH4+浓度在实验期间持续下降, Y和ZY在模拟尾水中NH4+浓度在反应初期升高较快, 第7 d开始, ZY中NH4+浓度开始下降明显, NH4+去除率达到62.3%, 第11 d开始Y中NH4+浓度开始下降, 第17 d时浓度降低较快[图 1(a)].由图 1(b)可明显看出, Z开始几天NO2-浓度出现上升随后快速下降去除率达到99.2%, Y中NO2-浓度变化较小, 去除率仅为18.9%, ZY和Y变化趋势相同, NO2-去除率为50.1%[图 1(b)].Z中NO3-浓度开始3 d呈现下降趋势, 然后浓度迅速增加, Y和ZY中NO3-浓度第1~9 d快速降低, 后期浓度变化不明显, 2组样品中NO3-降解率均在96%以上, 去除效果较好[图 1(c)].图 1(d)显示, 除Z中TN浓度下降明显外, Y和ZY中TN的去除效果不明显.
2.2 城市尾水中氮代谢分析Z、Y和ZY在城市尾水中氮的代谢特征如图 2所示.

|
图 2 城市尾水中氮代谢分析 Fig. 2 Analysis of nitrogen metabolism in urban tailwater |
Y和ZY在反应的前7 d阶段两组中NH4+浓度几乎保持一致, 3组在前5 d的NH4+浓度持续升高, 第5 d后浓度快速下降[图 2(a)], 主要是反应系统中硝化反应或反硝化反应环节缓慢且培养液中含氮底物的分解, 使NH4+浓度短暂升高, Z、Y和ZY最终降解率分别为91.6%、71.3%和97.1%.表明微藻和芽孢杆菌同时加入提高了城市尾水中NH4+去除率.由图 2(b)可以看出, 除Z中NO2-浓度逐渐降低外, Y和ZY中NO2-浓度出现上升趋势, 表明Y和ZY中芽孢杆菌在NO2-转化为NO3-过程中具有一定的抑制作用, 使系统中NO2-浓度出现增加现象.在图 2(c)中, Z中NO3-浓度不断增加, Y和ZY中NO3-浓度在第1 d迅速降低, 在3 d之后, Y和ZY系统中NO3-浓度较低, 变化趋势不明显, Y和ZY最终去除率分别为80.3%和99%, 降解效果较好.表明芽孢杆菌可以有效去除NO3-, 并可以提高微藻对NO3-的去除效率.图 2(d)中Y和ZY在反应的第3~7 d TN浓度显著增加, 原因是开始阶段加入的培养液中含氮有机物分解较快, 同时系统中菌株内部关系没有完全协调.第17 d时, ZY中TN的去除率不到40%, 系统显示一定的降解效果, 但降解效果一般.Z中TN浓度在反应前期不太稳定, 在反应的第7 d之后才开始缓慢稳定地下降, 但反应结束时系统中的TN去除率只在20%左右, Y中TN去除率较低, ZY中TN去除率高于Z和Y, 表明系统中同时添加微藻和芽孢杆菌可以提供系统中TN去除效率.
2.3 菌株高通量测序分析 2.3.1 物种组成分析稀释曲线(rarefaction curve)主要利用各样本在不同测序深度时的微生物α多样性指数构建曲线, 以此反映各样本在不同测序数量时的微生物多样性, 当曲线趋向平坦时, 说明测序数据量合理[22], 由图 3(a)可以看出测序reads数合理.OTU(operational taxonomic units)是在系统发生学中, 为了便于进行分析, 通过聚类, 将序列按照彼此的相似性分归为许多小组, 一个小组就是一个OTU.可根据不同的相似度水平, 对所有序列进行OTU划分, 通常对97%相似水平下的OTU进行生物信息统计分析[16].Venn图可用于统计多组或多个样本中所共有和独有的物种(如OTU)数目, 可以比较直观地表现环境样本的物种(如OTU)组成相似性及重叠情况.通常情况下, 分析时选用相似水平为97%的OTU样本[23].图 3(b)中2组共有OTU数目为216, Z和ZY中独有的OTU数目分别占14.6%和13.9%, 结果表明Z和ZY系统中OTU总数与各OTU所占比例变化不明显.

|
图 3 不同样本中细菌OTU的稀释曲线和分布 Fig. 3 Rarefaction curves and distribution of bacteria OTU in different samples |
群落组成分析是使用统计学分析方法, 检测样本在不同分类水平上的群落组成, 2组属水平群落组成结果如图 4所示.

|
图 4 属水平群落组成 Fig. 4 Community composition at the genus level |
由图 4可以看出, 2组样品物种组成发生了变化, Z中拟色球蓝细菌(Chroococcidiopsis)、norank_f_A4b、微小杆菌(Exiguobacterium)和瘦鞘丝藻属(Leptolyngbya)所占比例在ZY中明显减少, ZY中短芽孢杆菌(Brevibacillus)、梭状芽孢杆菌(Clostridium)和芽孢杆菌(Bacillus)成为样本数占比较高的菌群.图 4中显示Chroococcidiopsis和norank_ f_A4b这2个菌群相似度较高, 在ZY中减少数量较大, 占比分别减少为原来的29.6%和15.6%. Exiguobacterium和Leptolyngbya处于同一进化分支, 表明亲缘关系更加接近, 在ZY中群落数量减少明显, 占比分别为原来的59.7%和16.3%.ZY中Brevibacillus和Clostridium聚类在一起, 共同属于芽孢杆菌属, 数量比Z中增加明显, 分别增加了1 160倍和4 436倍.同时ZY中菌群结构也发生了明显变化, 在Z中的占比较少的菌群在ZY中出现增加趋势, 如Paracoccus、Limnobacter、Mycobacterium和Kapabacteriales 等.以上结果表明芽孢杆菌的加入明显改变了Z中菌群结构, 抑制了微藻的生长.
由图 5看出, 在Z中样本数占比较高的菌群分别为Chroococcidiopsis_PCC_7203 (24.38%)、bacterium-g_norank_ f_A4b (23.65%)、Exiguobacterium (7.09%)、Leptolyngbya_PCC-6306 (9.41%)和Bacillus (1.99%).在ZY中样本数占比较高的菌群分别为Brevibacillus (22.94%)、Clostridium_sensu_stricto13 (8.78%)和Bacillus (4.88%), Chroococcidiopsis_PCC_7203 样本数占比仅为7.84%, 远远低于Z中样本数占比, Leptolyngbya在ZY中样本数占比远远降低.

|
1.others, 2.Chroococcidiopsis_PCC_7203, 3. Bacterium-g_norank_ f__A4b, 4. Brevibacillus, 5. Exiguobacterium, 6. Leptolyngbya_PCC-6306, 7. Clostridium_sensu_stricto13, 8. Bacillus_PB19, 9. norank_ f__Blastocatellaceae, 10. Pseudomonas, 11. unclassified_ f__Rhodobacteraceae, 12. SWB02, 13. Allorhizobium-Neorhizobium, 14. Pararhizobium-Rhizobium, 15. Reyranella, 16. unclassified_ f__Rhizobiaceae, 17. norank_ f__Caldilineaceae, 18. Lysobacter, 19. Bosea, 20. norank_ f__Saprospiraceae, 21. norank_ f__Sandaracinaceae, 22. Brevundimonas, 23. Ancylobacter, 24. Devosia 图 5 属水平优势菌比例 Fig. 5 Proportions of dominant bacteria at the genus level |
同源蛋白簇(cluster of grthologous groups of protein, COG), 该数据库是对细菌、藻类和真核生物的21个完整基因组的编码蛋白根据系统进化关系分类构建而成, 蛋白所占的量反映对应环境下代谢特征[24].Z和ZY中COG功能预测如图 6所示.

|
图 6 样品中编码蛋白种类分布和数目变化 Fig. 6 Distribution and number of encoded proteins in the samples |
图 6(a)显示了不同功能蛋白在2组样品中的组成比例, 从图 6(b)可以看出, Z中芽孢杆菌的加入, 在ZY中表现出编码蛋白数目发生了变化, 其中转录蛋白(transcription, K) 和脂质运输和代谢蛋白(lipid transport and metabolism, I)数目下调明显; 能源生产和转化(energy production and conversion, C)、辅酶转运和代谢(Coenzyme transport and metabolism, H)、翻译、核糖体结构和生物转化(translation, ribosomal structure and biogenesis, J)、复制、重组和修复(replication, recombination and repair, L)、细胞壁/膜/包膜生物发生(replication, recombination and repair, M)、位置修饰、蛋白质周转、伴侣(posttranslational modification, protein turnover, chaperones, O)、无机离子转运和代谢(inorganic ion transport and metabolism, P)、一般功能预测(general function prediction only, R)、功能未知(function unknown, S)及信号转导机制(signal transduction mechanisms, T)这些功能蛋白数目上调明显.结果表明在Z中K和I的功能优于ZY, 其中C、H、J、L、M、O、P、R、S和T功能优于Z.图 6(c)和6(d)显示, 2组样品中的前5个最优势功能蛋白的顺序不同, Z和ZY分别为S、R、T、氨基酸运输代谢功能(amino acid transport and metabolism, E)和M蛋白与S、E、R、T和C蛋白, 说明芽孢杆菌的加入对菌群代谢差生了影响.以上结果表明, 在ZY中E成为较优势功能蛋白, 芽孢杆菌影响了菌群的代谢功能.
2.3.4 样品菌群差异性分析利用卡方检验[25] (chi-square exact test)分析2组样品中菌群结构在属水平的差异显著性, 结果如图 7所示.

|
Y轴表示某一分类水平下的物种名, 物种对应的每个柱子表示该物种在各样本相对丰度, 不同颜色表示不同样本; 中间区域为所设定的置信区间内, 圆点对应的数值表示物种在两样本中相对丰度的差值, 圆点颜色显示为物种丰度占比较大的样本颜色, 圆点上的I型区间为差值的上下限值, *** 表示P≤0.001 图 7 属水平差异性检验 Fig. 7 Difference test at the genus level |
图 7中结果进一步表明, 芽孢杆菌加入, 显著降低了Z中拟色球蓝细菌(Chroococcidiopsis)、uncultured_bacterium_g_norank_ f_A4b和瘦鞘丝藻属(Leptolyngbya) 这3个属的数量; 显著增加了短芽孢杆菌(Brevibacillus)、梭状芽孢杆菌(Clostridium)和芽孢杆菌(Bacillus). norank_ f-norank_o-PB19、unclassified_ f__Rhodobacteraceae和SWB02 在ZY中也是显著增加的菌群, 芽孢杆菌可以明显影响产藻菌的结构组成和种群数量.
3 讨论图 1(a)的Y和ZY在模拟尾水中NH4+浓度在反应初期升高较快, 实验反应开始一周内呈持续且快速升高的趋势, 主要是系统中部分含氮底物被氨化细菌等分解为铵根离子, 在一定时间内没被及时转化造成一定量的累积.图 1(b)的ZY中NO2-浓度在反应前期只有较小的变化浮动, 可降解约50%的NO2-, 反应中没有出现NO2-浓度极不稳定的现象, 表明系统中微生物在参与氮循环过程中可能需要的溶解氧较少.由图 1(c)可知, Z中NO3-浓度在反应的第1~3 d期间呈快速下降的趋势, 之后的反应阶段模拟污水中其含量出现持续上升的现象, 主要是系统中由于藻类的光合作用产生氧气.当系统中溶解氧出现升高时, NO2-易转化为NO3-[26], 而且系统不能形成缺氧环境, 影响了反硝化细菌的活性及反硝化能力, 使其不能及时将中间产物NO3-转化为氮气, 造成了中间产物NO3-长时间出现累积现象.图 2 (a)的Z和ZY对城市尾水中NH4+去除效果较好, 2组都达到了95%以上, Z中NH4+降解率高达99%以上, 几乎完全去除尾水中的NH4+, Li等[27]分别在经过灭菌处理的城市污水原水和原水里培养小球藻, 14 d的批次培养后小球藻对原水中NH4+去除率最高可达93.9%, 结果表明微藻去除NH4+效率较高.由图 2 (b)可知, Z中NO2-去除效果最好, 芽孢杆菌与微藻的共同作用在氮循环反应NO2-转化为NO3-过程中发挥出较强的稳定效果, 可能具有平衡反应系统中溶解氧供应需求的功能.图 2 (c)的ZY中NO3-去除效果最好, 其降解率高达99%以上, 几乎完全去除城市尾水中的NO3-, 微藻与芽孢杆菌的共同作用可提高脱氮菌去除水中的NO3-的降解率及稳定性, Sydney等[6]利用微藻对城市污水二级出水进行处理也得出了79.63%的硝酸盐去除率.微藻的同化过程在将无机氮转化为其有机形式中发挥关键作用, 有研究发现藻类优先利用NH4+而不是NO3-, 并且直到NH4+几乎完全消耗完才开始消耗NO3-, 因此系统中NH4+去除效率较高[28].也有研究发现, NO3-在氧化水生环境中具有较好的稳定性, 但硝酸盐的存在可以诱导硝酸还原酶的活性也是微藻的必需氮源[29], 因此ZY系统对NO3-具有较好的去除效果.
由图 4可知, ZY中短芽孢杆菌、梭状芽孢杆菌和芽孢杆菌成为样本数占比较高的菌群, 芽孢杆菌的加入抑制了藻类的生长, 使藻细胞逐渐减少.有研究发现细菌与藻之间的相互作用是影响藻类生长的关键性因素, 水华发生时, 藻细胞数目迅速增加, 抑藻细菌能够优先利用藻类分泌的有机物, 数量不断增加, 并占细菌群落中的绝大多数, 抑藻细菌杀死藻细胞, 释放有机物刺激周围群落中抑藻细菌的生长[30].随着抑藻细菌在时间以及空间上的持续作用, 藻细胞数目不断减少, 最终消亡, 伴随着抑藻细菌能够优先用的有机物含量的不断下降, 抑藻细菌不断减少, 回到最初水平[31].原因是部分芽孢杆菌可以通过释放胞外物质对微藻产生一定的抑制作用[32].图 6显示在ZY中, 芽孢杆菌影响了菌群的代谢功能, 氨基酸运输代谢功能蛋白数量增加, 表明系统中氨基酸浓度增加.有些芽孢杆菌能够分泌细胞外物质, 对宿主藻类起抑制或杀灭作用, 有研究认为水华和赤潮的突然消亡可能与溶藻细菌的感染有关[33].Ahn等[34]研究结果显示, 枯草芽孢杆菌所分泌枯草菌环肽在10 mg·L-1的浓度下, 可在2 d内去除85%的微藻; Mu等[35]筛选出的1株芽孢杆菌, 可通过分泌具有耐热性的胞外物质促使微藻裂解, 达到抑藻目的, Zhao等[36]研究也得出部分氨基酸对藻类生长具有抑制作用.因此在城市尾水富营养化修复过程中, 可以施加芽孢杆菌菌剂, 通过直接和间接作用抑制藻类的生长, 防止水体富营养化现象发生.
4 结论(1) 系统中同时添加微藻与芽孢杆菌, 可更大程度地降解水中NO3-和TN.
(2) 微藻脱氮系统中芽孢杆菌的加入, 改变了系统微生物群落结构及代谢功能.
(3) 短芽孢杆菌、梭状芽孢杆菌和芽孢杆菌对拟色球蓝细菌和瘦鞘丝藻具有一定的抑制作用.
| [1] |
王钰涛, 范晨阳, 朱金鑫, 等. 尾水排放对受纳水体底栖生物膜细菌群落和水溶性有机质的影响机制[J]. 环境科学, 2021, 42(12): 5826-5835. Wang Y T, Fan C Y, Zhu J X, et al. Impacts of wastewater effluent discharge on bacteria community and water-soluble organic matter in benthic biofilm in receiving river[J]. Environmental Science, 2021, 42(12): 5826-5835. |
| [2] |
廖雪珂, 严晗璐, 王智源, 等. 低温季节水平潜流和垂直潜流人工湿地尾水深度处理中试[J]. 环境科学, 2020, 41(12): 5509-5517. Liao X K, Yan H L, Wang Z Y, et al. Advanced treatment of tail water using pilot-scale horizontal and vertical subsurface flow constructed wetlands in low-temperature seasons[J]. Environmental Science, 2020, 41(12): 5509-5517. |
| [3] |
李胜男, 陈豪宇, 彭华, 等. 丰水期东洞庭湖超微型浮游藻类时空分布特征及其影响因子[J]. 环境科学, 2020, 41(6): 2679-2687. Li S N, Chen H Y, Peng H, et al. Temporal and spatial variation patterns of picophytoplankton and their correlations with environmental factors during the wet season in east lake Dongting[J]. Environmental Science, 2020, 41(6): 2679-2687. |
| [4] | Olguín E J. Dual purpose microalgae-bacteria-based systems that treat wastewater and produce biodiesel and chemical products within a biorefinery[J]. Biotechnology Advances, 2012, 30(5): 1031-1046. DOI:10.1016/j.biotechadv.2012.05.001 |
| [5] | Shi J, Podola B, Melkonian M. Removal of nitrogen and phosphorus from wastewater using microalgae immobilized on twin layers: an experimental study[J]. Journal of Applied Phycology, 2007, 19(5): 417-423. DOI:10.1007/s10811-006-9148-1 |
| [6] | Sydney E B, Da Silva T E, Tokarski A, et al. Screening of microalgae with potential for biodiesel production and nutrient removal from treated domestic sewage[J]. Applied Energy, 2011, 88(10): 3291-3294. DOI:10.1016/j.apenergy.2010.11.024 |
| [7] |
杨坤, 李静, 卢文轩. 不同初始氮浓度对小球藻生长及氮吸收的影响[J]. 环境科学与技术, 2014, 37(12): 40-46. Yang K, Li J, Lu W X. Effects of different initial nitrogen concentrations on growth and nitrogen absorption of chlorella vulgaris[J]. Environmental Science & Technology, 2014, 37(12): 40-46. |
| [8] | Chen J J, Lee Y R, Huang Y C, et al. Isolation of microalgae with potential for integrated biomass production and nutrient removal from wastewater[J]. Desalination and Water Treatment, 2018, 127: 151-157. DOI:10.5004/dwt.2018.22928 |
| [9] | Lu Y P, Wang J, Yu Y, et al. Inhibition of Camellia sinensis (L.) O. Kuntze on Microcystis aeruginosa and isolation of the inhibition factors[J]. Biotechnology Letters, 2013, 35(7): 1029-1034. DOI:10.1007/s10529-013-1188-4 |
| [10] | Shi W Q, Bi L, Pan G. Effect of algal flocculation on dissolved organic matters using cationic starch modified soils[J]. Journal of Environmental Sciences, 2016, 45: 177-184. DOI:10.1016/j.jes.2015.12.018 |
| [11] | Dumack K, Müller M E H, Bonkowski M. Description of Lecythium terrestris sp. nov. (chlamydophryidae, cercozoa), a soil dwelling protist feeding on fungi and algae[J]. Protist, 2016, 167(2): 93-105. DOI:10.1016/j.protis.2016.01.001 |
| [12] | Liu J Y, Yang C Y, Chi Y X, et al. Algicidal characterization and mechanism of Bacillus licheniformis Sp34 against Microcystis aeruginosa in Dianchi Lake[J]. Journal of Basic Microbiology, 2019, 59(11): 1112-1124. DOI:10.1002/jobm.201900112 |
| [13] |
韩博远, 张闻, 胡芳雨, 等. 模拟及实际根系分泌物对芘污染土壤微生物群落的影响[J]. 环境科学, 2022, 43(2): 1077-1088. Han B Y, Zhang W, Hu F Y, et al. Influence of artificial root exudates and actual root exudates on the microbial community in pyrene-contaminated soil[J]. Environmental Science, 2022, 43(2): 1077-1088. |
| [14] |
邸琰茗, 王广煊, 黄兴如, 等. 再生水补水对河道底泥细菌群落组成与功能的影响[J]. 环境科学, 2017, 38(2): 743-751. Di Y M, Wang G X, Huang X R, et al. Effect of reclaimed water on bacterial community composition and function in urban river sediment[J]. Environmental Science, 2017, 38(2): 743-751. |
| [15] | 闫杨, 刘月静, 陈芳. 枯草芽孢杆菌的应用现状概述[J]. 生物学教学, 2019, 44(2): 2-3. DOI:10.3969/j.issn.1004-7549.2019.02.001 |
| [16] | 张睿. 枯草芽孢杆菌对铜绿微囊藻抑制效果的研究[D]. 上海: 上海海洋大学, 2015. |
| [17] | Song Z, Liu Q X, Guo H, et al. Tostadin, a novel antibacterial peptide from an antagonistic microorganism Brevibacillus brevis XDH[J]. Bioresource Technology, 2012, 111: 504-506. DOI:10.1016/j.biortech.2012.02.051 |
| [18] | Cui P, Liao H P, Bai Y D, et al. Hyperthermophilic composting reduces nitrogen loss via inhibiting ammonifiers and enhancing nitrogenous humic substance formation[J]. Science of the Total Environment, 2019, 692: 98-106. DOI:10.1016/j.scitotenv.2019.07.239 |
| [19] |
赵志瑞, 刘硕, 李铎, 等. 脱氮菌剂在低溶解氧黑臭水体中氮代谢特征[J]. 环境科学, 2020, 41(1): 304-312. Zhao Z R, Liu S, Li D, et al. Characteristics of nitrogen metabolism by denitrifying bacterial agents in low dissolved oxygen black odor water[J]. Environmental Science, 2020, 41(1): 304-312. |
| [20] |
梁洪儒, 吴艳玲. 水中硝酸盐氮两种检测方法的比对分析[J]. 山东水利, 2021(6): 71-73. Liang H R, Wu Y L. Comparative analysis of two detection methods of nitrate nitrogen in water[J]. Shandong Water Resources, 2021(6): 71-73. |
| [21] |
景胜元. 碱性过硫酸钾消解紫外分光光度法测定水质中总氮实验空白的分析与研究[J]. 山西化工, 2021, 41(4): 131-133, 155. Jing S Y. Analysis and study on the blank of total nitrogen determination in water by UV spectrophotometry with alkaline potassium persulfate digestion[J]. Shanxi Chemical Industry, 2021, 41(4): 131-133, 155. |
| [22] | Ye J, Joseph S D, Ji M K, et al. Chemolithotrophic processes in the bacterial communities on the surface of mineral-enriched biochars[J]. The ISME Journal, 2017, 11(5): 1087-1101. |
| [23] | Ji P, Rhoads W J, Edwards M A, et al. Impact of water heater temperature setting and water use frequency on the building plumbing microbiome[J]. The ISME Journal, 2017, 11(6): 1318-1330. |
| [24] | Wong K, Shaw T I, Oladeinde A, et al. Rapid microbiome changes in freshly deposited cow feces under field conditions[J]. Frontiers in Microbiology, 2016, 7. DOI:10.3389/fmicb.2016.00500 |
| [25] | Zheng J, Xiao X H, Zhang Q, et al. Correlation of placental microbiota with fetal macrosomia and clinical characteristics in mothers and newborns[J]. Oncotarget, 2017, 8(47): 82314-82325. |
| [26] | Seuntjens D, Han M F, Kerckhof F M, et al. Pinpointing wastewater and process parameters controlling the AOB to NOB activity ratio in sewage treatment plants[J]. Water Research, 2018, 138: 37-46. |
| [27] | Li Y C, Chen Y F, Chen P, et al. Characterization of a microalga Chlorella sp. well adapted to highly concentrated municipal wastewater for nutrient removal and biodiesel production[J]. Bioresource Technology, 2011, 102(8): 5138-5144. |
| [28] | He T X, Xie D T, Li Z L, et al. Ammonium stimulates nitrate reduction during simultaneous nitrification and denitrification process by Arthrobacter arilaitensis Y-10[J]. Bioresource Technology, 2017, 239: 66-73. |
| [29] | Barsanti L, Gualtieri P. Algae: anatomy, biochemistry, and biotechnology (2nd ed.)[M]. Boca Raton: CRC Press, 2014. |
| [30] | Doucette G J, McGovern E R, Babinchak J A. Algicidal bacteria active against gymnodinium breve (dinophyceae). i. bacterial isolation and characterization of killing activity[J]. Journal of Phycology, 1999, 35(6): 1447-1454. |
| [31] |
瞿建宏, 刘韶斌. 水体中芽孢杆菌和微囊藻的生长及其资源竞争[J]. 湛江海洋大学学报, 2002, 22(3): 13-18. Qu J H, Liu S B. The growth of Bacillus sp. and Microcystis aeruginosa and their competition for resources[J]. Journal of Zhanjiang Ocean University, 2002, 22(3): 13-18. |
| [32] |
张睿, 王广军, 李志斐, 等. 枯草芽孢杆菌对铜绿微囊藻抑制效果的研究[J]. 中国环境科学, 2015, 35(6): 1814-1821. Zhang R, Wang G J, Li Z F, et al. Inhibition of Microcystis aeruginosa by Bacillus subtilis[J]. China Environmental Science, 2015, 35(6): 1814-1821. |
| [33] | Lee S J, Kim Y, Kim H G, et al. Algalytic activity of α-mannosidase on harmful marine microalgae[J]. Journal of Applied Phycology, 2000, 12(2): 191-193. |
| [34] | Ahn C Y, Joung S H, Jeon J W, et al. Selective control of cyanobacteria by surfactin-containing culture broth of Bacillus subtilis C1[J]. Biotechnology Letters, 2003, 25(14): 1137-1142. |
| [35] | Mu R M, Fan Z Q, Pei H Y, et al. Isolation and algae-lysing characteristics of the algicidal bacterium B5[J]. Journal of Environmental Sciences, 2007, 19(11): 1336-1340. |
| [36] | Zhao L, Chen L N, Yin P H. Algicidal metabolites produced by Bacillus sp. strain B1 against Phaeocystis globosa[J]. Journal of Industrial Microbiology and Biotechnology, 2014, 41(3): 593-599. |