 2023, Vol. 44
2023, Vol. 44

2. 中国科学院南京地理与湖泊研究所湖泊与环境国家重点实验室, 南京 210008;
3. 无锡市河湖治理和水资源管理中心, 无锡 214031
 , XU Hai2 , ZHAN Xu1
, XU Hai2 , ZHAN Xu1 , ZHANG Zheng-hui3 , ZHU Meng-yuan2 , ZOU Wei2 , XIAO Man2 , NI Zi-yi1 , ZHU Hui1
, ZHANG Zheng-hui3 , ZHU Meng-yuan2 , ZOU Wei2 , XIAO Man2 , NI Zi-yi1 , ZHU Hui1
2. State Key Laboratory of Lakes and Environment, Nanjing Institute of Geography and Limnology, Chinese Academy of Sciences, Nanjing 210008, China;
3. Wuxi River and Lake Management and Water Resources Management Center, Wuxi 214031, China
蓝藻水华已成为全球性最严重的水环境问题之一, 严重威胁全球水生生态系统和公共健康[1]. 20世纪80年代以来, 全球68%的湖泊都曾出现过有害藻类水华的现象[2]. 藻类水华不仅危害鱼类等水生生物的生存, 更严重的可能导致大型动物甚至人类产生疾病甚至死亡[3]. 在全球气候变化的影响下, 湖泊蓝藻水华呈现明显的范围扩张、持续时间延长、发生频率增加和危害性加剧的趋势, 其防控面临着巨大挑战[4].
氮和磷是引起水体富营养化和蓝藻水华暴发的关键营养元素. 传统观点认为淡水生态系统初级生产力主要受到磷的控制, 然而对于许多浅水湖泊而言, 由于长期积蓄在底泥中的磷极易通过释放进入水相, 被藻类吸收利用, 导致湖泊富营养化治理对外源磷的控制响应滞后, 而对氮的控制响应迅速[5, 6]. 因此, 近十几年来关于水环境中活性氮去除过程的相关研究备受国内外学者广泛关注[7]. 反硝化作用是在微生物的驱动下, 硝态氮被逐步还原为气态氮的过程[8], 被认为是浅水湖泊氮素去除的主要途径[9]. 有研究表明, 全球约20%的氮去除由水体中的反硝化过程来完成[10]. 反硝化作用能够导致生态系统向氮限制状态转变, 在一定程度上能够有效地缓解水体富营养化的发生[6]. 然而, 气候变化能够通过影响湖泊生物地球化学过程, 改变蓝藻对营养盐的输移转化和补给模式, 促使蓝藻水华日益严重.
反硝化作用主要受温度、溶解氧、有机碳和硝态氮浓度等多种环境因素的影响, 其中温度和硝态氮浓度常被作为影响反硝化速率的主要变量[11]. 温度主要影响酶的活性以及反硝化微生物生长繁殖的速率[8], 在反应底物充足的情况下温度控制着能够达到的最大反硝化速率, 即反硝化潜力. 硝态氮作为反硝化作用的底物, 一直被认为是影响反硝化速率的首要因素[12]. 一般而言, 反硝化速率会随着水体硝酸盐浓度的增加而升高[13]. 只有在适宜的温度以及足够的硝态氮浓度下, 反硝化速率才会达到最高[11]. 全球气候变化背景下, 气候变暖导致湖泊温度在春季提前升高, 这会导致浮游植物的群落结构发生变化, 喜好高温的蓝藻可能提前占据优势, 形成藻类水华[14]. 同时湖泊内的反硝化反应也会受到温度的影响, 使氮素脱离水体. 因此在气候变暖的背景下, 反硝化作用与蓝藻水华之间可能存在对氮素的竞争, 这个过程取决于水体温度和氮素浓度的动态变化. 目前关于气候变化背景下反硝化脱氮的动态变化对蓝藻水华发生态势的影响研究相对较少.
太湖是一个大型浅水湖泊, 是我国第三大淡水湖. 近年来, 由于城市化的发展以及农业肥料的使用等, 大量活性氮进入湖体, 使得太湖长期处于富营养化的状态, 蓝藻水华频发. 自2007年无锡饮用水危机事件以来, 太湖富营养化问题引起了国内外学者的广泛关注. 太湖具有很强的氮素自净能力, 反硝化脱氮可以去除50%的外源氮负荷[6], 国家政府也针对太湖富营养化问题进行了高强度的治理[15], 两者共同作用使得太湖氮浓度近年呈现显著的降低趋势[16], 但是蓝藻水华发生情况仍不容乐观, 2017年甚至发生了2007年以来最严重的蓝藻水华[15]. 有研究认为是气候变暖抵消了外源治理的效果[17]. 然而, 气候变暖促进蓝藻水华暴发强度增加的具体机制仍不清楚.
基于此, 本研究于2021年9月采集太湖竺山湾蓝藻样品进行不同温度下的培养实验, 并于2022年4月及6月采集竺山湾沉积物样品, 利用15N同位素示踪技术, 采用泥浆法对沉积物进行反硝化培养, 阐明温度和硝态氮浓度对反硝化速率的影响. 结合2017~2021年为期5 a的北太湖水质监测数据对蓝藻水华与反硝化作用之间的联系进行分析, 深入探讨太湖水体反硝化脱氮过程对蓝藻水华情势的可能影响, 以期为蓝藻水华发生机制及氮素管理提供理论依据和科技支撑.
1 材料与方法 1.1 研究区域概况太湖是中国第三大淡水湖, 位于长江三角洲, 气候温和湿润, 属亚热带季风气候, 水域面积为2 338.1 km2, 平均水深1.9 m[18]. 该地区人口众多, 具有高水平的工业化和城市化规模, 提供了旅游、水产养殖、饮用水供应等多种生态服务功能, 是中国经济最为发达的地区之一. 近几十年来, 由于人类活动的过度干扰, 加速了太湖富营养化的进程, 加之气候日趋变暖, 使得蓝藻水华暴发频发.
1.2 采样点位太湖地区夏季盛行东南风, 蓝藻水华常在风力的作用下迁移至北部湖区造成大量聚集, 其中竺山湾、大浦口与梅梁湾尤为严重, 而竺山湾又与多条入湖河道相连, 承担着较高的入湖氮负荷.
2017~2021年太湖氮磷浓度和藻类生物量数据来自太湖湖泊生态系统研究站, 在太湖北部湖区的竺山湾、梅梁湾、大浦口、贡湖湾和湖心区共设立14个监测点位, 分别为梅梁湾7个(1~6号、14号)、竺山湾2个(12号和13号)、贡湖湾2个(10号和11号)、大浦口1个(9号)、湖心区1个(8号)和梅梁湾与湖心区的中间位置1个(7号).

|
图 1 采样点位空间分布示意 Fig. 1 Spatial distribution of sampling sites |
使用采水器采集上、中和下3层水样, 水样采集混匀后冷藏带回实验室并立即过滤一部分, 滤膜为Whatman公司的GF/F膜, 孔径0.7 μm. 使用碱性过硫酸钾氧化和紫外分光光度法对原水进行总氮(TN)和总磷(TP)的测定, 对滤后水使用Skalar连续流动分析仪(荷兰Skalar公司, SAN++型)测定硝态氮(NO3--N)、亚硝态氮(NO2--N)、氨氮(NH4+-N)和磷酸盐(PO43--P), 所有样品在测定前均置于-20℃冰箱保存. 滤膜经冷冻48 h后使用热乙醇提取和分光光度法测定Chla浓度.
于2021年9月采集竺山湾13号点的原位湖水, 立即带回实验室进行不同温度蓝藻培养实验. 分别于2022年4月和6月采集竺山湾13号点的沉积物以及上覆水. 使用彼得森采泥器对表层5 cm沉积物进行采集, 采集3次, 混匀后装袋, 冷藏带回实验室, 一部分用于测定理化性质, 一部分用于泥浆实验.
1.4 蓝藻温度阈值培养实验于2021年9月进行不同温度下的蓝藻培养实验, 设立3个温度梯度, 分别为10、20和30℃. 将采集的原位湖水分别分装1.5 L至12个底部被橡胶塞塞紧密封的有机玻璃柱中, 并向每个柱子中加入KNO3溶液与KH2PO4溶液使柱内水体ρ(N)增加2 mg·L-1, ρ(P)增加0.02 mg·L-1. 随机抽取3个柱子充分混匀后取水样立即过滤, 滤膜测定Chla, 作为初始值. 剩余柱子分别随机抽取3个置于10、20和30℃的恒温水浴锅中, 使用日光灯(2 000 lx)12 h循环光照培养4 d后, 取水样立即过滤, 滤膜测定Chla, 作为结束值.
1.5 反硝化适宜温度与底物浓度阈值培养实验于2022年4月进行不同温度的培养实验, 设立7个温度梯度, 分别为5、10、15、20、25、30和35℃, 每个温度设立3个平行. 将采集的沉积物与水样以湿重1∶5的比例混合装入血清瓶, 充分搅匀并曝氦气20 min以使其处于厌氧状态, 包裹锡纸放入水浴锅中设置温度25℃, 恒温避光预培养48 h, 将本底硝态氮消耗完全. 预培养结束后, 向血清瓶中加入Na15NO3进行示踪, 并使其ρ(NO3--N)为4 mg·L-1, 将充分混合均匀的泥浆分装到24个气密性硼硅酸盐小瓶(12 mL, Labco Exetainer)中, 随机抽取3个小瓶立即加入0.1~0.2 mL质量分数为50%的饱和ZnCl2溶液以终止微生物反应作为初始, 并分别随机抽取3个小瓶放入7个不同温度的水浴锅中恒温避光培养4 h, 取出后立即加入ZnCl2溶液.
于2022年6月进行不同浓度的培养实验, 设立5个浓度梯度, 分别为1、2、3、4和5 mg·L-1, 每个浓度设立6个平行. 将采集的沉积物与水样以湿重1∶5的比例混合装入5个血清瓶, 将每个血清瓶都充分搅匀并曝氦气20 min以使其处于厌氧状态, 以相同的方式对这5个血清瓶进行预培养. 预培养结束后, 向血清瓶中加入Na15NO3, 并使5个血清瓶中ρ(NO3--N)分别为1、2、3、4和5 mg·L-1, 将每个血清瓶中充分混合均匀的泥浆分别分装到6个硼硅酸盐小瓶中, 分别随机抽取3个小瓶立即加入ZnCl2溶液作为初始, 将其余小瓶放入25℃水浴锅中恒温避光培养4 h, 取出后立即加入ZnCl2溶液.
培养结束, 使用膜进样质谱仪测定样品的28N2、29N2和30N2.
1.6 反硝化速率计算反硝化速率根据以下公式计算[19]:
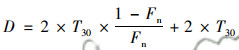
|
式中, D为反硝化速率[μmol·(kg·h)-1]; T30为30N2产生速率[μmol·(kg·h)-1]; Fn为15N丰度(%).
1.7 数据处理与分析使用SPSS 26软件对数据进行Shapiro-Wilk正态性检验和Levene方差齐性检验, 对不符合方差齐性的数据进行对数转换, 利用单因素方差分析的方法分析数据间的显著性差异. 图形绘制使用ArcGIS 10.2和Origin 2018软件.
2 结果与分析 2.1 太湖氮磷浓度与藻类生物量季节变化2017~2021年北太湖逐月水温(WT)与氮磷浓度和藻类生物量的变化如图 2所示. 太湖水体温度在5~35℃范围内变化, 表现出明显的季节变化, 春夏秋冬平均水温分别为17.74、28.57、20.76和7.04℃. 2017年4月水温升高较早, 提前达到了20℃以上, 而其他年份4月水温仅在15℃左右. ρ(TN)季节变化较大, 在1.36~3.36 mg·L-1之间, 但每年的均值相差不大, 分别为2.79、2.36、2.46、2.32和2.05 mg·L-1, 2017、2018、2019和2021年ρ(TN)最高的月份均为3月, 分别为3.30、3.36、3.21和2.49 mg·L-1, 2020年5月ρ(TN)最高, 为3.01 mg·L-1. 整体而言太湖北部湖区TN浓度在冬春季节要显著高于夏秋季节.各年NO3--N浓度最高的月份与TN浓度最高的月份相同, 但夏秋季节的NO3--N浓度极低. 太湖中溶解态无机氮主要以NO3--N为主, NH4+-N浓度普遍低于NO3--N浓度, NO2--N浓度极低. ρ(TP)在冬春季保持在0.1 mg·L-1左右, 夏秋季显著升高并达到0.2 mg·L-1左右, 2017年6月和2020年9月达到了最高值0.26 mg·L-1, 而PO43--P浓度相对比较稳定. 以Chla形式表示的藻类生物量呈现出与NO3--N完全相反的规律, 表现为夏秋季节较高, 冬春季节较低, 但2017年春季生物量显著高于夏季.

|
图 2 2017~2021年太湖北部湖区总氮(TN)、硝态氮(NO3--N)、亚硝态氮(NO2--N)、氨氮(NH4+-N)、总磷(TP)、磷酸盐(PO43--P)、Chla和水体温度(WT)变化 Fig. 2 Variations in total nitrogen (TN), nitrate (NO3--N), nitrite (NO2--N), ammonia (NH4+-N), total phosphorus (TP), phosphate (PO43--P), Chla and water temperature (WT) in northern Lake Taihu from 2017 to 2021 |
在不同温度下蓝藻的生长状况如表 1所示, ρ(Chla)的初始值为(14.58±2.40) μg·L-1, 10℃时ρ(Chla)降低较为显著, 为(4.84±1.16) μg·L-1; 20℃时ρ(Chla)没有出现显著地降低, 为(12.22±6.20) μg·L-1; 在30℃时ρ(Chla)相比于初始增加了72.6%, 为(25.17±7.84) μg·L-1. 以上结果表明, 蓝藻生长的喜好温度较高.
|
|
表 1 培养前及培养4 h后不同温度下Chla浓度/μg·L-1 Table 1 Chla concentrations at different temperatures before and after 4 h incubation/μg·L-1 |
2.3 温度和硝态氮浓度对反硝化速率的影响
温度对反硝化潜力的影响如图 3(a)所示, 在5~35℃都观察到了反硝化作用的发生, 在25℃时反硝化潜力最高, 为(62.98±21.36) μmol·(kg·h)-1, 超过25℃后反硝化潜力不再增加, 30℃和35℃时的反硝化潜力虽然相比25℃时略低, 但没有显著差异, 分别为(56.21±3.81) μmol·(kg·h)-1和(58.29±14.36) μmol·(kg·h)-1, 5℃时反硝化潜力最低, 为(3.64±0.98) μmol·(kg·h)-1. 如图 3(b)所示, 在10~25℃时反硝化潜力与温度存在显著的线性关系(R2=0.99).

|
不同小写字母间存在显著性差异 图 3 温度和反硝化潜力的关系 Fig. 3 Relationship between temperature and denitrification potential |
硝态氮浓度对反硝化速率的影响如图 4所示, 反硝化速率随着硝态氮浓度的增加而增加, 在ρ(NO3--N)达到4 mg·L-1时反硝化速率达到最高, 为(47.34±7.37) μmol·(kg·h)-1, 5 mg·L-1时反硝化速率不再增加, 其速率与4 mg·L-1时基本一致, 为(47.26±2.14) μmol·(kg·h)-1. 1 mg·L-1时反硝化速率最低, 为(18.68±0.53) μmol·(kg·h)-1. ρ(NO3--N)为2 mg·L-1和3 mg·L-1时反硝化速率分别为(21.52±1.74) μmol·(kg·h)-1和(31.53±0.37) μmol·(kg·h)-1.

|
图 4 不同硝态氮浓度下的反硝化速率 Fig. 4 Denitrification rates at different nitrate concentrations |
野外长期监测数据显示, 太湖不同形态氮浓度呈现出显著的季节变化, 硝态氮和氨氮浓度从春季开始显著降低, 直到夏秋季节一直处于全年较低值, 而这正是蓝藻生长繁殖并大面积暴发形成水华的时期. 关于太湖夏秋季总氮浓度处于全年最低的原因, 很多研究人员认为是藻类吸收的原因, 但藻类吸收只会改变氮素的形态, 不会改变总氮的浓度. Xu等[6]结合水位数据计算的水柱总氮现存量也在夏秋季节明显降低, 表明氮素的绝对量从湖体损失了. 蓝藻的生长繁殖会同化吸收水体中的溶解态无机氮(DIN), 以氨氮优先[20], 其次是硝态氮, 亚硝态氮. 太湖溶解态无机氮主要以硝态氮为主, 氨氮和亚硝态氮浓度普遍较低. 对硝态氮与总氮的比值(NO3--N/TN)和Chla浓度进行相关性分析, 结果显示太湖Chla浓度与NO3--N/TN呈显著负相关关系(r=-0.77, P<0.01), 表明太湖蓝藻的生长繁殖期是硝态氮的低谷期. Xu等[21]研究结果发现春季无机氮浓度的下降并没有完全支持叶绿素的增加, 因此, 太湖夏季无机氮浓度降低, 一方面是藻类生长吸收以藻体有机氮的形式存在, 另一方面也是反硝化脱氮的结果[22]. Xu等[6]通过质量平衡计算了太湖季节反硝化脱氮量, 发现50%的外源氮输入可以通过反硝化作用被脱除.
反硝化是一个酶促反应, 温度的高低对其速率的影响尤为显著[23]. 从本文的研究结果可以看出, 太湖沉积物的反硝化作用对温度的变化非常敏感, 10~25℃范围内反硝化作用与温度之间呈线性增加关系, 25~35℃范围内是反硝化作用的最适温度, 表明从冬季到春季, 随着温度升高, 反硝化速率呈现快速增加趋势, 蓝藻水华暴发期太湖沉积物的反硝化潜力最高. 但赵锋等[24]通过流动培养研究太湖不同湖区春夏两季原位反硝化速率, 发现夏季反硝化速率普遍低于春季. 硝态氮浓度是影响反硝化速率的另一个重要环境因子[25], 甚至比温度的影响更为显著[26].太湖反硝化达到最大潜力的ρ(NO3--N)为4mg·L-1, 在1.0mg·L-1时最低, 太湖夏季氮浓度, 尤其是ρ(NO3--N)处于全年最低值, 在0.07~0.31 mg·L-1范围. 因此, 夏季反硝化作用受到硝酸盐底物的严重限制.
蓝藻水华作为富营养化的产物, 反过来又会影响湖泊的氮磷循环. 蓝藻生长繁殖过程可以同化吸收大气中的二氧化碳和水体中的溶解态氮, 这些藻体在水动力、风速和风向适宜的条件下漂浮至水面, 迁移至下风口经过一段时间堆积死亡后又将氮素重新释放到水体[15, 27]. 蓝藻降解产生的溶解态有机碳(DOC)以及厌氧环境有利于促进泥-水界面反硝化作用的进行. 而且反硝化微生物可以附着在蓝藻上, 形成以藻颗粒为中心的耗氧-厌氧微环境, 使其成为反硝化作用的另一个场所从而提高氮的去除效率[28]. 太湖夏季总磷浓度持续居高不下, 在蓝藻水华暴发期间总磷浓度出现峰值, 相关性分析结果表明总磷浓度与Chla浓度呈现显著的正相关关系(r=0.88, P<0.01). 而溶解态磷酸盐的浓度却始终变化不大, 表明蓝藻水华暴发期, 蓝藻会同化吸收大量的磷[29], 以藻体有机磷的形式贡献总磷浓度. 蓝藻水华暴发一方面提高了水体的pH, 另一方面导致水体出现周期性厌氧环境, 长期积蓄在底泥中的磷极易释放进入水相, 被藻类吸收利用, 导致控磷效果并不理想[30]. 相比于缺少气态脱除方式的磷, 氮可以通过反硝化等作用从湖体脱除, 促使蓝藻水华出现氮限制[5], 因此, 在控磷的同时协同控氮对蓝藻水华的控制将起到更快的效果[5, 6].
3.2 反硝化脱氮对蓝藻水华态势的影响机制蓝藻的生长繁殖和反硝化过程对氮的共同需求, 会使两者产生一个相互的影响. 浮游植物的生长具有周期性, 受温度影响大, 不同种群之间对温度耐受范围不同. 相对于其他藻种来说, 蓝藻的喜好温度相对较高, 通常在25℃以上[14, 31]. 和浮游植物的生长相同, 湖泊各种生源要素的生物地球化学循环也受到温度的调控. 需要指出的是, 虽然在低温时反硝化速率较低, 但是在5~35℃范围内都可以发生, 而偏离适宜温度时水体浮游植物生长会明显受抑, 较低的温度下蓝藻的生长繁殖会受到限制. 因此, 在达到蓝藻可以大量生长繁殖的温度之前, 湖泊里的氮素会被反硝化作用不断脱除[32]. 随着湖泊温度的升高, 蓝藻开始复苏并大量生长繁殖[33], 这时会和反硝化作用共同利用湖泊中的氮, 产生竞争的局面. 但是与反硝化不同的是, 蓝藻吸收的氮并没有将其脱离湖泊, 而是以藻体有机氮的形式短暂储存在湖泊内[34], 蓝藻死亡降解后氮会以有机氮和氨氮的形式重新回到水体[35], 部分氮会通过硝化-反硝化作用被去除[36], 而部分氮又被蓝藻吸收利用[20], 使蓝藻水华持续发生.
由于气候变暖导致湖泊温度提前升高, 促使蓝藻春季物候期提前, 蓝藻水华发生时间提前且持续时间延长[37]. 近年来太湖蓝藻水华由最初的夏季集中暴发向春季扩张[37], 这会导致湖泊里大量的氮素尚未被反硝化作用脱除就被藻类吸收利用, 使蓝藻水华暴发态势更为严重, 并且减少的脱氮量可能会影响后期蓝藻水华态势的发展. 2017年4月太湖春季温度提前升高, 5月中旬时水温达到25℃左右, 在春季出现了大规模的蓝藻水华[38], 并一直持续到夏秋季节, 氮的可利用性可能是主要的控制因素.
3.3 太湖水华防治的措施和建议20世纪70年代, 形成了以控磷为主的湖泊富营养化的治理策略[39], 并在多个深水湖泊取得了效果[40], 但在许多大型富营养化浅水湖泊的治理成效并不明显. 在太湖进行的原位营养富集实验发现, 在控磷的基础上减少氮负荷对控制太湖蓝藻水华至关重要[5]. 外源不断输入的磷会大量蓄积在沉积物中, 与深水湖泊不同的是, 浅水湖泊极易发生沉积物再悬浮并将磷释放至水体中[15], 并且蓝藻生长消耗水体中的溶解态总磷(DTP)也会导致磷的大量释放[41], 与磷不同的是, 氮除了被藻类同化吸收外还会通过反硝化作用脱离水体, 使太湖夏秋季出现氮限制. 因此防治太湖水华在控磷的基础上控氮也是必要的. 并且Xu等[32]指出保持ρ(TN)在0.8 mg·L-1, ρ(TP)在0.05 mg·L-1以下时, 水华暴发的风险将会降低.
气候变暖导致的水温提前升高使太湖内源磷污染日趋加重[15], 并导致蓝藻水华对控制外源磷输入的响应比较缓慢, 因此要加强内源磷的治理. 底泥疏浚对于内源污染有着较好的治理效果, 但内源污染从根本上都是来自外源, 长期的外源输入会使水质出现反弹[42]. 因此, 对外源营养盐输入的管理也至关重要, 在管理污染排放的基础上可以在太湖流域增加沟塘湿地以减少营养盐的入湖负荷[43]. 相比于缺少脱除途径的磷, 对氮的控制在减轻外源的基础上也应重视反硝化脱氮作用, 比如培育水生植物[44]、搭建生态浮床[45]等提高反硝化效率, 在蓝藻水华暴发前, 尽可能减少湖泊内氮的浓度.
4 结论(1) 太湖总氮浓度夏秋季低于冬春季, 溶解态无机氮主要以硝态氮为主, 并在夏秋季处于低谷; 总磷浓度与Chla浓度的变化一致, 夏秋季显著高于冬春季, 表明夏秋季节藻类生长主要受氮浓度的限制.
(2) 太湖反硝化作用的最适温度为25~35℃, 在10~25℃范围内反硝化速率与温度呈显著的线性关系(R2=0.99); 最高底物浓度阈值为4 mg·L-1, ρ(NO3--N)低于4 mg·L-1时反硝化速率随浓度升高而升高.
(3) 蓝藻水华暴发时太湖反硝化潜力最高, 但较低的硝态氮浓度导致反硝化作用受底物限制, 实际反硝化速率较低.
(4) 反硝化作用和蓝藻的生长繁殖会竞争硝态氮, 蓝藻生长的适宜温度在25℃以上, 温度较低时不会大量繁殖, 气候变暖导致蓝藻水华提前暴发, 氮被反硝化脱除前被大量吸收利用, 导致蓝藻水华暴发严重.
致谢: 太湖湖泊生态系统研究站提供了2017~2021年逐月的北太湖水体各形态氮磷和浮游植物叶绿素a浓度数据, 表示感谢.
| [1] | Brooks B W, Lazorchak J M, Howard M D A, et al. Are harmful algal blooms becoming the greatest inland water quality threat to public health and aquatic ecosystems?[J]. Environmental Toxicology and Chemistry, 2016, 35(1): 6-13. DOI:10.1002/etc.3220 |
| [2] | Ho J C, Michalak A M, Pahlevan N. Widespread global increase in intense lake phytoplankton blooms since the 1980s[J]. Nature, 2019, 574(7780): 667-670. DOI:10.1038/s41586-019-1648-7 |
| [3] | Wang H J, Xu C, Liu Y, et al. From unusual suspect to serial killer: cyanotoxins boosted by climate change may jeopardize megafauna[J]. The Innovation, 2021, 2(2). DOI:10.1016/j.xinn.2021.100092 |
| [4] | Hou X J, Feng L, Dai Y H, et al. Global mapping reveals increase in lacustrine algal blooms over the past decade[J]. Nature Geoscience, 2022, 15(2): 130-134. DOI:10.1038/s41561-021-00887-x |
| [5] | Xu H, Paerl H W, Qin B Q, et al. Nitrogen and phosphorus inputs control phytoplankton growth in eutrophic Lake Taihu, China[J]. Limnology and Oceanography, 2010, 55(1): 420-432. DOI:10.4319/lo.2010.55.1.0420 |
| [6] | Xu H, McCarthy M J, Paerl H W, et al. Contributions of external nutrient loading and internal cycling to cyanobacterial bloom dynamics in Lake Taihu, China: implications for nutrient management[J]. Limnology and Oceanography, 2021, 66(4): 1492-1509. DOI:10.1002/lno.11700 |
| [7] | Yu C Q, Huang X, Chen H, et al. Managing nitrogen to restore water quality in China[J]. Nature, 2019, 567(7749): 516-520. DOI:10.1038/s41586-019-1001-1 |
| [8] |
徐硕, 李玉双, 魏建兵, 等. 河岸带土壤反硝化作用研究进展[J]. 生态科学, 2021, 40(4): 229-236. Xu S, Li Y S, Wei J B, et al. Research progress of riparian soil denitrification[J]. Ecological Science, 2021, 40(4): 229-236. DOI:10.14108/j.cnki.1008-8873.2021.04.026 |
| [9] | Roland F A E, Darchambeau F, Borges A V, et al. Denitrification, anaerobic ammonium oxidation, and dissimilatory nitrate reduction to ammonium in an East African Great Lake (Lake Kivu)[J]. Limnology and Oceanography, 2018, 63(2): 687-701. DOI:10.1002/lno.10660 |
| [10] | Seitzinger S, Harrison J A, Böhlke J K, et al. Denitrification across landscapes and waterscapes: a synthesis[J]. Ecological Applications, 2006, 16(6): 2064-2090. DOI:10.1890/1051-0761(2006)016[2064:DALAWA]2.0.CO;2 |
| [11] | Zhong J C, Fan C X, Liu G F, et al. Seasonal variation of potential denitrification rates of surface sediment from Meiliang Bay, Taihu Lake, China[J]. Journal of Environmental Sciences, 2010, 22(7): 961-967. DOI:10.1016/S1001-0742(09)60205-9 |
| [12] | Zhao Y Q, Xia Y Q, Ti C P, et al. Nitrogen removal capacity of the river network in a high nitrogen loading region[J]. Environmental Science & Technology, 2015, 49(3): 1427-1435. |
| [13] |
代军帅, 左小虎, 王明霞, 等. 硝酸盐对土壤反硝化活性及蒽厌氧降解的影响[J]. 环境科学, 2018, 39(1): 422-429. Dai J S, Zuo X H, Wang M X, et al. Effect of nitrate amendment on soil denitrification activity and anthracene anaerobic degradation[J]. Environmental Science, 2018, 39(1): 422-429. DOI:10.13227/j.hjkx.201706267 |
| [14] |
邓建明, 秦伯强. 全球变暖对淡水湖泊浮游植物影响研究进展[J]. 湖泊科学, 2015, 27(1): 1-10. Deng J M, Qin B Q. A review on studies of effects of climate change on phytoplankton in freshwater systems[J]. Journal of Lake Sciences, 2015, 27(1): 1-10. |
| [15] |
秦伯强. 浅水湖泊湖沼学与太湖富营养化控制研究[J]. 湖泊科学, 2020, 32(5): 1229-1243. Qin B Q. Shallow lake limnology and control of eutrophication in Lake Taihu[J]. Journal of Lake Sciences, 2020, 32(5): 1229-1243. |
| [16] | Yang Z, Zhang M, Shi X L, et al. Nutrient reduction magnifies the impact of extreme weather on cyanobacterial bloom formation in large shallow Lake Taihu (China)[J]. Water Research, 2016, 103: 302-310. DOI:10.1016/j.watres.2016.07.047 |
| [17] | Qin B Q, Paerl H W, Brookes J D, et al. Why Lake Taihu continues to be plagued with cyanobacterial blooms through 10 years (2007-2017) efforts[J]. Science Bulletin, 2019, 64(6): 354-356. DOI:10.1016/j.scib.2019.02.008 |
| [18] | Zou W, Zhu G W, Xu H, et al. Temporal dependence of chlorophyll a-nutrient relationships in Lake Taihu: Drivers and management implications[J]. Journal of Environmental Management, 2022, 306. DOI:10.1016/j.jenvman.2022.114476 |
| [19] | Hou L J, Yin G Y, Liu M, et al. Effects of sulfamethazine on denitrification and the associated N2O release in estuarine and coastal sediments[J]. Environmental Science & Technology, 2015, 49(1): 326-333. |
| [20] |
韩菲尔, 赵中华, 李大鹏, 等. 利用稳定同位素(15N)示踪技术研究浮游藻类氮素吸收速率特征[J]. 海洋与湖沼, 2019, 50(4): 811-821. Han F E, Zhao Z H, Li D P, et al. The use of 15N to measure nitrogen uptake of phytoplankton[J]. Oceanologia et Limnologia Sinica, 2019, 50(4): 811-821. |
| [21] | Xu H, Paerl H W, Zhu G W, et al. Long-term nutrient trends and harmful cyanobacterial bloom potential in hypertrophic Lake Taihu, China[J]. Hydrobiologia, 2017, 787(1): 229-242. DOI:10.1007/s10750-016-2967-4 |
| [22] | Shen Y S, Huang Y Y, Hu J, et al. The nitrogen reduction in eutrophic water column driven by Microcystis blooms[J]. Journal of Hazardous Materials, 2020, 385. DOI:10.1016/j.jhazmat.2019.121578 |
| [23] | Shiau Y J, Dham V, Tian G L, et al. Factors influencing removal of sewage nitrogen through denitrification in mangrove soils[J]. Wetlands, 2016, 36(4): 621-630. DOI:10.1007/s13157-016-0770-7 |
| [24] |
赵锋, 许海, 詹旭, 等. 太湖春夏两季反硝化与厌氧氨氧化速率的空间差异及其影响因素[J]. 环境科学, 2021, 42(5): 2296-2302. Zhao F, Xu H, Zhan X, et al. Spatial differences and influencing factors of denitrification and ANAMMOX rates in spring and summer in Lake Taihu[J]. Environmental Science, 2021, 42(5): 2296-2302. |
| [25] | Zhang D D, Li M Y, Yang Y C, et al. Nitrite and nitrate reduction drive sediment microbial nitrogen cycling in a eutrophic lake[J]. Water Research, 2022, 220. DOI:10.1016/j.watres.2022.118637 |
| [26] | Palacin-Lizarbe C, Camarero L, Catalan J. Denitrification temperature dependence in remote, cold, and N-Poor lake sediments[J]. Water Resources Research, 2018, 54(2): 1161-1173. DOI:10.1002/2017WR021680 |
| [27] |
肖琳, 杨佳怡. 水华对沉积物中氮的转化和去除的影响[J]. 环境科学, 2019, 40(9): 4018-4022. Xiao L, Yang J Y. Water bloom modified sediment nitrogen transformation and removal[J]. Environmental Science, 2019, 40(9): 4018-4022. |
| [28] |
刘志迎, 许海, 詹旭, 等. 蓝藻水华对太湖水柱反硝化作用的影响[J]. 环境科学, 2019, 40(3): 1261-1269. Liu Z Y, Xu H, Zhan X, et al. Influence of cyanobacterial blooms on denitrification rate in shallow Lake Taihu, China[J]. Environmental Science, 2019, 40(3): 1261-1269. |
| [29] |
陈洁, 许海, 詹旭, 等. 沉积物参与下氮磷脉冲式输入对太湖水体营养盐浓度和藻类生长的影响[J]. 环境科学, 2020, 41(6): 2671-2678. Chen J, Xu H, Zhan X, et al. Influence of nutrient pulse input on nitrogen and phosphorus concentrations and algal growth in the sediment-water system of Lake Taihu[J]. Environmental Science, 2020, 41(6): 2671-2678. |
| [30] | Howarth R, Paerl H W. Coastal marine eutrophication: control of both nitrogen and phosphorus is necessary[J]. Proceedings of the National Academy of Sciences of the United States of America, 2008, 105(49). DOI:10.1073/pnas.0807266106 |
| [31] | Paerl H W, Huisman J. Blooms like it hot[J]. Science, 2008, 320(5872): 57-58. |
| [32] | Chen X F, Jiang H Y, Sun X, et al. Nitrification and denitrification by algae-attached and free-living microorganisms during a cyanobacterial bloom in Lake Taihu, a shallow Eutrophic Lake in China[J]. Biogeochemistry, 2016, 131(1-2): 135-146. |
| [33] |
吴晓东, 孔繁翔, 张晓峰, 等. 太湖与巢湖水华蓝藻越冬和春季复苏的比较研究[J]. 环境科学, 2008, 29(5): 1313-1318. Wu X D, Kong F X, Zhang X F, et al. Comparison of overwintering and rrecruitment of cyanobacteria in Taihu Lake and Chaohu Lake[J]. Environmental Science, 2008, 29(5): 1313-1318. |
| [34] | Baulch H M, Stanley E H, Bernhardt E S. Can algal uptake stop NO3- pollution?[J]. Nature, 2011, 477(7366). DOI:10.1038/nature10418 |
| [35] |
朱梦圆, 朱广伟, 王永平. 太湖蓝藻水华衰亡对沉积物氮、磷释放的影响[J]. 环境科学, 2011, 32(2): 409-415. Zhu M Y, Zhu G W, Wang Y P. Influence of scum of algal bloom on the release of N and P from sediments of Lake Taihu[J]. Environmental Science, 2011, 32(2): 409-415. |
| [36] |
林伟, 李玉中, 李昱佳, 等. 氮循环过程的微生物驱动机制研究进展[J]. 植物营养与肥料学报, 2020, 26(6): 1146-1155. Lin W, Li Y Z, Li Y J, et al. Advances in the mechanism of microbe-driven nitrogen cycling[J]. Journal of Plant Nutrition and Fertilizers, 2020, 26(6): 1146-1155. |
| [37] | Shi K, Zhang Y L, Zhang Y B, et al. Phenology of phytoplankton blooms in a trophic lake observed from long-term MODIS data[J]. Environmental Science & Technology, 2019, 53(5): 2324-2331. |
| [38] |
朱伟, 陈怀民, 王若辰, 等. 2017年太湖水华面积偏大的原因分析[J]. 湖泊科学, 2019, 31(3): 621-632. Zhu W, Chen H M, Wang R C, et al. Analysis on the reasons for the large bloom area of Lake Taihu in 2017[J]. Journal of Lake Sciences, 2019, 31(3): 621-632. |
| [39] | Schindler D W. Eutrophication and recovery in experimental lakes: implications for lake management[J]. Science, 1974, 184(4139): 897-899. |
| [40] | Schindler D W, Carpenter S R, Chapra S C, et al. Reducing phosphorus to curb lake eutrophication is a success[J]. Environmental Science & Technology, 2016, 50(17): 8923-8929. |
| [41] | Xie L Q, Xie P, Li S X, et al. The low TN: TP ratio, a cause or a result of Microcystis blooms?[J]. Water Research, 2003, 37(9): 2073-2080. |
| [42] |
张建华, 殷鹏, 张雷, 等. 底泥疏浚对太湖内源及底栖生物恢复的影响[J]. 环境科学, 2023, 44(2): 828-838. Zhang J H, Yin P, Zhang L, et al. Effects of sediment dredging on the reduction of sediment internal loading of Lake Taihu and the self-recovery ability of benthic organism[J]. Environmental Science, 2023, 44(2): 828-838. |
| [43] |
王子聪, 许海, 詹旭, 等. 天目湖流域沟塘湿地脱氮速率的时空差异[J]. 环境科学研究, 2022, 35(4): 979-988. Wang Z C, Xu H, Zhan X, et al. Temporal and spatial characteristics of nitrogen removal in ditches and ponds in tianmuhu Lake Basin[J]. Research of Environmental Sciences, 2022, 35(4): 979-988. |
| [44] |
赵琳, 李正魁, 周涛, 等. 伊乐藻-氮循环菌联用对太湖梅梁湾水体脱氮的研究[J]. 环境科学, 2013, 34(8): 3057-3063. Zhao L, Li Z K, Zhou T, et al. Denitrification study of elodea nuttallii-Nitrogen cycling bacteria restoration in Meiliang Bay, Taihu Lake[J]. Environmental Science, 2013, 34(8): 3057-3063. |
| [45] |
唐伟, 许海, 詹旭, 等. 生态浮床对千岛湖水体氮磷净化效果研究[J]. 环境科学研究, 2022, 35(4): 926-935. Tang W, Xu H, Zhan X, et al. Nitrogen and phosphorus purification effects of ecological floating bed in Qiandao Lake[J]. Research of Environmental Sciences, 2022, 35(4): 926-935. |