 2023, Vol. 44
2023, Vol. 44

2. 浙江农林大学省部共建亚热带森林培育国家重点实验室, 杭州 311300;
3. 浙江农林大学现代农学院, 杭州 311300
 , JIANG Wen-ting1,2 , LIU Xun-yue3 , ZHU Gao-di1,2
, JIANG Wen-ting1,2 , LIU Xun-yue3 , ZHU Gao-di1,2 , TANG Rong-gui1,2 , ZHANG Hai-bo2 , CAI Yan-jiang1,2
, TANG Rong-gui1,2 , ZHANG Hai-bo2 , CAI Yan-jiang1,2
2. State Key Laboratory of Subtropical Silviculture, Zhejiang A&F University, Hangzhou 311300, China;
3. College of Advanced Agricultural Sciences, Zhejiang A&F University, Hangzhou 311300, China
微塑料(microplastics, MPs)通常是指直径小于5 mm的塑料颗粒[1], 因其难降解和易迁移等特点, 被认为是一种全球性新型污染物, 普遍存在于大气、水和土壤等环境介质中[2].微塑料粒径小且降解周期长, 极易被生物体吸收并随之在食物链中转移和富集, 从而危及生态环境安全及人类健康[3].鉴于此, 联合国环境规划署(UNPE)已于2014年将微塑料污染列为全球十大环境问题之一[4].当前关于微塑料的研究大多集中于海岸[5]、海洋[6]、河口[7]和湖泊[8]等水生生态系统.相比于水生生态系统, 陆地生态系统尤其是农业生态系统中微塑料污染的研究仍相对较少.事实上, 农业生产过程中因大量使用塑料薄膜、地膜和含有微塑料成分的农用化学品, 会通过污水灌溉、大气沉降等途径而导致农田土壤中微塑料的累积量远高于海洋生态系统[9].较高质量分数的MPs累积于农田土壤中, 会威胁农业生产活动.例如, MPs可被生菜(Lactuca sativa)和小麦(Triticum aestivum L.)等农作物吸收和富集, 并从根部迁移到地上部, 分布于可直接被食用的茎叶之中[10, 11], 最终对人类生命健康构成威胁.
MPs进入农业土壤后可引起土壤结构和理化性质的变化, 进而扰动土壤养分原有的循环过程, 最终影响植物种子萌发和后续生长[12].种子萌发是植物生命周期的关键阶段之一, 发芽率、发芽势、发芽指数和活力指数等均是决定种子发芽能力的重要指标, 能够直接反映种子的生长趋势[13].有研究表明, MPs胁迫能够延缓种子发芽和抑制植物生长[14, 15].如叶子琪等[16]发现500 mg ·kg-1的聚乙烯微塑料(polyethylene microplastics, PE MPs)可显著降低蚕豆的总生物量, 抑制光合作用, 这与Pignattelli等[17]的研究结果类似.然而, 有研究则发现MPs能够增加植物叶片中叶绿素含量, 促进植物的生长发育[18~20].此外, 新近研究指出, MPs还可诱导植物产生过多活性氧(ROS), 导致细胞器的致命受损[21].通常, 植物体内会产生一系列的抗氧化酶, 如超氧化物歧化酶(SOD)、过氧化物酶(POD)和过氧化氢酶(CAT)以清除ROS带来的毒性[22, 23].然而, 在不同质量分数MPs胁迫下, 植物体内的抗氧化酶活性均呈现出不同的变化趋势[24, 25].丙二醛(MDA)是氧化应激引起的脂质过氧化的副产物, 是细胞膜损伤程度的生物学指标[26], 其含量的变化可以表征植物在MPs胁迫下的过氧化和脂质损伤程度.Dong等[27]发现MPs可通过损坏水稻抗氧化酶结构, 诱导水稻组织脂质过氧化, 提高其MDA含量.Abduro等[28]则指出MPs可降低植物叶片的MDA含量.MPs还可通过影响植物体内可溶性蛋白和可溶性糖等渗透调节物质的含量来缓解外界胁迫[29, 30].如Dong等[31]研究发现浓度为1 g ·L-1的聚丙乙烯微塑料可导致生菜可溶性蛋白和可溶性糖含量显著下降, 但Li等[32]指出浓度为10 g ·L-1的聚丙乙烯微塑料可显著提高黄瓜(Cucumis sativus L.)果实中可溶性蛋白含量.
综上可见, 当前MPs对农作物生长和生理的影响尚有争议, 可能与MPs的质量分数、种类和植物对象等相关, 仍有待进一步研究.低密度聚乙烯(low-density polyethylene, LDPE)是农用塑料薄膜的主要组成成分, 因农用塑料薄膜的大量使用, 其已成为土壤微塑料污染物的主要类型之一[33, 34].空心菜是一种广泛种植的食用蔬菜, 为促进空心菜种植过程中的保温高产, 覆膜栽培技术成为空心菜种植过程中的常用措施之一.然而, 有关低密度聚乙烯微塑料(low-density polyethylene microplastics, LDPE MPs)对于空心菜生长生理的效应, 目前尚不清楚且未见报道.因此, 本研究选用柳叶空心菜(Ipomoea aquatica Forsk)为供试材料, 以粒径 < 200 μm的LDPE MPs颗粒作为胁迫物质, 探讨不同质量分数的LDPE MPs对空心菜生长和生理的影响, 以期为后续评估MPs对蔬菜生产的影响提供参考依据.
1 材料与方法 1.1 试验材料空心菜种子购于长沙市银田蔬菜种子实业有限公司.LDPE MPs颗粒购于上海冠步机电科技有限公司, 粒径 < 200 μm, 其形貌如图 1所示.

|
图 1 扫描电镜下LDPE MPs颗粒的形貌 Fig. 1 Scanning electron micrograph of LDPE MPs particle surface characteristics |
土壤采自浙江农林大学东湖校区官塘附近的蔬菜地土壤(30°15′ N, 119°43′ E), 其基本理化性质如下:pH 5.47, ω(有机质)17.53 g ·kg-1, ω(全氮)0.33 g ·kg-1, ω(有效磷)30.39 mg ·kg-1, ω(速效钾)65.33 mg ·kg-1.试验前用不锈钢铲在0~20 cm土壤深度范围内采集供试土壤, 过1 cm筛, 以去除植物的根、茎、有机碎屑和石子等杂物.盆栽试验盆子直径15 cm, 高13 cm, 盆底垫纱网以防土漏出, 每盆装风干土2 kg.本试验在学校温室大棚内进行, 将LDPE MPs颗粒与过筛土壤多次搅拌充分混合均匀, 用自来水浇灌, 采用称重法使土壤水分含量达到田间最大持水量(WHC)的40%.稳定15 d后, 以溶液的形式一次性施入尿素、磷酸二氢钠和硫酸钾作为基肥, 控制土壤中氮磷钾的质量浓度比例为1 ∶1 ∶1.
本试验设置不添加LDPE MPs的对照组(CK), 添加0.2%(质量分数, 下同)LDPE MPs(PE1), 添加5% LDPE MPs(PE2)和添加10% LDPE MPs(PE3)这4个处理, 添加质量分数参考了目前已有报道的微塑料含量以及以往报道试验中的含量[35, 36].已有研究表明, 土壤中的MPs质量分数范围为0.03% ~6.75%[37], 考虑到农用薄膜的使用量大、回收率低的情况, 且微塑料会在土壤中不断积累[38], 本试验将最高质量分数设置为10%.每个处理均以溶液的形式施加等量肥料.每个处理设置4个重复, 盆栽随机摆放于温室内.
空心菜种子放置在30~40℃之间的温水中, 浸泡24 h进行催芽处理, 播种至放有营养土的穴盘中育苗, 待幼苗长出两片真叶后, 挑选长势相似的幼苗移栽至盆内, 每盆定植3株.用称重法将含水量调至60% WHC.定植后及时用喷壶灌溉补充水分, 保证土壤水分充足.除虫、除草等管理与当地田间管理方式保持一致.空心菜生长45 d后收割第一茬, 75 d后收割第二茬, 共收割两茬.
1.2.2 发芽试验发芽试验在穴盘内进行, 试验设计、微塑料和土壤处理、施肥量及方法与盆栽试验相同, 催芽后在每个穴内播种1粒空心菜种子, 每盘播种20粒, 共4盘.
播种后7 d内, 每天于固定时间记录空心菜种子的发芽个数, 并用刻度尺测量第7 d的幼苗长度.按式(1)~(4)分别计算种子的发芽势(germination energy, GE)、发芽指数(germination index, GI)、活力指数(vigor index, VI)和发芽率(germination rate, GR)[15].
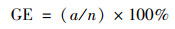
|
(1) |
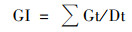
|
(2) |
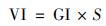
|
(3) |
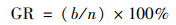
|
(4) |
式中, a为3 d内发芽种子数; b为7 d内发芽种子数; n为供试种子总数; Dt为发芽日数; Gt为与Dt相对应的每日发芽种子数; S为一定时期内的幼苗长度(cm ·株-1).
1.3 植株样品采集与测定空心菜植株收获前, 先用剪刀采集约3 g叶片置于自封袋内, 经液氮粉碎后于-80℃冰箱保存, 用于后续指标的测定.第一茬收获时, 采集基部两节以上的地上部分, 第二茬收获时采集整颗植株.植株用自来水连续冲洗干净, 再以去离子水冲洗, 用滤纸擦干后置于信封内备用.主要指标测定方法如下.
1.3.1 叶绿素含量的测定采用分光光度法, 用丙酮-乙醇混合溶液(V丙酮 ∶V乙醇=1 ∶1)提取植物叶片中的叶绿素, 用紫外可见光分光光度计(岛津UV-2600)分别测量在470、645和663 nm波长下的吸光度值.按式(5)~(8)分别计算叶片叶绿素a、叶绿素b、总叶绿素和类胡萝卜素含量[39].
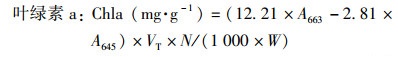
|
(5) |
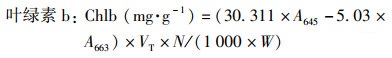
|
(6) |
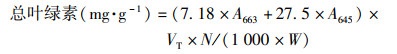
|
(7) |
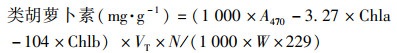
|
(8) |
式中, VT为提取液体积; W为样品鲜重; N为稀释倍数.
1.3.2 生物量的测定植株鲜样在105℃烘箱内杀青30 min, 后60℃烘至恒重, 测定植株干重.
1.3.3 叶片SOD、POD和CAT活性、MDA、可溶性蛋白和可溶性糖含量的测定取经液氮粉碎后的新鲜空心菜叶片, 用PBS缓冲液(0.05 mol ·L-1 pH 7.4)按重量(g) ∶体积(mL)=1 ∶6的比例制备组织匀浆, 离心(4 000 r ·min-1, 15 min), 取上清液待测, 所有步骤均在冰浴上进行.按南京建成生物工程研究所的试剂盒说明测定并计算空心菜叶片超氧化物歧化酶(SOD, 黄嘌呤氧化酶法)、过氧化氢酶(CAT, 紫外吸收法)、过氧化物酶(POD, 愈创木酚法)活力、丙二醛(MDA, 硫代巴比妥酸法)和可溶性蛋白含量(考马斯亮蓝法).
取经液氮粉碎后的新鲜空心菜叶片, 用超纯水按重量(g) ∶体积(mL)=1 ∶10的比例制备组织匀浆, 沸水浴10 min, 冷却后离心(4 000 r ·min-1, 10 min), 取上清液待测.按南京建成生物工程研究所的试剂盒说明测定并计算空心菜叶片可溶性糖含量(蒽酮比色法).
1.4 数据处理与分析所有数据均用Microsoft Excel 2016进行处理与计算, 采用SPSS 26.0软件进行数据分析, 图表中数据均以平均值±标准偏差表示.所有数据均通过方差齐性检验, 符合正态分布, 运用单因素方差分析法(ANOVA)进行差异显著性分析.采用Origin 2018软件作图.
2 结果与分析 2.1 种子发芽率及生长特性、光合色素含量和生物量LDPE MPs各处理对种子发芽率无显著影响(P>0.05), 但对于发芽势、发芽指数和活力指数等指标而言, 随LDPE MPs质量分数的增加, 抑制程度越高(表 1).PE3处理中空心菜的发芽势、发芽指数和活力指数均显著低于CK处理(P < 0.05), 分别降低了42.31%、28.68%和21.47%.与之不同, PE1和PE2处理对发芽势和活力指数均表现出微弱的抑制作用, 但差异不显著(P>0.05).
|
|
表 1 空心菜种子活力1) Table 1 Seed vigor of water spinach |
就第一茬而言, 添加LDPE MPs的各处理对叶绿素a[图 2(a)]、叶绿素b[图 2(b)]、总叶绿素[图 2(c)]和类胡萝卜素[图 2(d)]的含量均无显著影响(P>0.05).但在第二茬中PE2和PE3处理则显著抑制了叶绿素b和类胡萝卜素含量(P < 0.05), 且都在PE3处理中出现最低值.相反, PE1处理则对各光合色素含量表现为微弱的促进作用, 但差异不显著(P>0.05).

|
不同小写字母表示不同处理之间差异显著(P < 0.05), 下同 图 2 空心菜叶片光合色素含量 Fig. 2 Photosynthetic pigment content in leaves of water spinach |
从总体上来看, LDPE MPs各处理对空心菜地上生物量和地下生物量均呈现先升后降的趋势, 且PE2处理的第一茬、第二茬地上和地下生物量达到最大值, 总地上生物量较CK处理增加了39.41%(表 2), 显著促进了空心菜的生长(P < 0.05).PE3处理时, LDPE MPs对空心菜的生长无显著影响(P>0.05).
|
|
表 2 空心菜生物量 Table 2 Water spinach biomass |
2.2 SOD、POD、CAT活性和MDA含量
随着LDPE MPs质量分数的升高, SOD活性先升后降, 在PE1处理中达到最大值, 较CK处理显著提高了8.68%[P < 0.05, 图 3(a)].LDPE MPs添加对POD活性无显著影响, 但PE3处理的POD活性略高于其余处理[P>0.05, 图 3(b)].PE2和PE3处理显著提高第一茬空心菜叶片CAT活性[P < 0.05, 图 3(c)], 第二茬CAT活性在PE3中达到最大值.第一茬中, LDPE MPs的添加显著降低了MDA含量(P < 0.05), 且随着质量分数的增加, MDA含量呈现阶梯式下降, PE1、PE2和PE3处理较CK处理分别降低了15.53%、26.81%和27.39%[图 3(d)].但在第二茬中, PE2处理显著增加了叶片的MDA含量(P < 0.05), 增加幅度为33.58%.

|
图 3 空心菜叶片抗氧化酶活性和丙二醛含量 Fig. 3 Antioxidant enzyme activity and malondialdehyde content in leaves of water spinach |
特别地, PE2处理显著降低空心菜叶片第一茬可溶性蛋白含量, 较CK降低了29.63%, PE3处理可溶性蛋白含量较CK显著提高了27.80%[P < 0.05, 图 4(a)], 但在第二茬中各处理间无显著差异(P>0.05).同时, LDPE MPs在两茬收获中均显著提高了可溶性糖含量(P < 0.05), 但不同质量分数处理间无显著差异[P>0.05, 图 4(b)].此外, 第二茬的可溶性蛋白和可溶性糖含量均显著高于第一茬(P < 0.05).

|
图 4 空心菜叶片可溶性蛋白和可溶性糖含量 Fig. 4 Soluble protein and soluble sugar content in leaves of water spinach |
本研究发现, LDPE MPs显著抑制空心菜种子的活力, 这与Pignattelli等[17]的研究结果一致.种子发芽阶段对水分的需求量极大, 而MPs可以吸附在种子、胚根和胚毛的表面, 堵塞种子囊中空隙[14], 这导致种子无法及时吸收水分供给自身发育, 进而抑制种子萌发.本研究还发现, LDPE MPs对空心菜种子发芽的抑制效果会随着质量分数的升高而增强, 这可能是由于MPs颗粒中化合物浸出导致, 随着MPs质量分数的升高, MPs释放出更多的有毒物质[40], 从而降低种子活力.然而, 本研究却并未发现LDPE MPs对种子的发芽率产生显著影响, 这可能是由于随着MPs暴露时间增加, 种子自身已吸收的充足水分抵消了MPs对于种子萌发的负效应[41].
此外, PE2和PE3显著降低空心菜叶片叶绿素a、叶绿素b、总叶绿素和类胡萝卜素的含量.这可能是因为MPs被空心菜吸收后在体内积累, 阻隔了部分叶绿素的合成途径, 与叶绿素合成有关的蛋白酶活性受到抑制, 进而使叶绿素含量下降[42].此外, MPs的氧化胁迫使ROS在植物细胞中积累, 破坏叶绿体结构[43], 进而使叶绿素合成受阻, 最终导致光合色素含量的下降.
特别地, PE2可显著增加空心菜地上和地下生物量, 而其他处理则无显著影响, 这与文献[19]的研究结果类似, 造成这一现象的原因可能是MPs的添加降低了土壤容重[44, 45].本研究结果发现, PE2的土壤容重较CK显著降低[(1.09±0.03)g ·cm-3和(1.21±0.04)g ·cm-3, P < 0.05].土壤容重的降低可以减小植物根系的渗透阻力, 改善土壤通气性, 从而促进根系生长, 增加根系总长度、直径、体积和质量等[46, 47].以上差异还可能通过改变土壤团聚体直接或间接影响土壤结构, 进而促进植株生长[48].然而, PE2和PE3在第二茬中对空心菜叶片叶绿素a、叶绿素b、总叶绿素和类胡萝卜素的含量表现出显著的抑制作用, 这与生物量的结果相反.由此可以推测, 土壤容重降低对植物生长的促进作用缓解了因光合色素含量的降低而带来的负面效应.此外, 本研究还发现, PE3的生物量显著低于PE2, 生物量有下降趋势, 这与本研究中LDPE MPs对空心菜光合色素含量影响结果一致, 由此本研究推测MPs可通过影响植株光合作用, 进而对植株生长构成威胁.
3.2 LDPE对空心菜生理特性的影响本研究发现, 随着LDPE MPs质量分数的升高, 空心菜体内的SOD活性呈先升后降的规律, SOD活性的短暂升高是机体为免受外界胁迫和毒害而做出的调节反应, 而之后活性下降是由于产生的过量自由基已超过酶作用阈值[49].这与大豆在受到聚氯乙烯微塑料胁迫时激发体内抗氧化系统, 从而减少氧化损伤的研究结果一致[50].SOD可以催化超氧阴离子生成H2 O2和O2, 是机体抗氧化防御的第一道防线[51].CAT和POD均是普遍存在的氧化还原酶, 可催化分解过量H2 O2以防止脂质过氧化[51], 因此CAT和POD在活力变化上较SOD有滞后性.在本研究中, MPs的添加虽没有使POD活性出现显著差异, 但可看到PE3处理下CAT活性出现了显著提高.CAT和POD活性的上升可能是空心菜为防止ROS增加的一种保护策略, 也是空心菜在LDPE MPs暴露下避免氧化应激的一种适应性反应.类似的, Zhou等[49]发现在50 mg ·L-1和100 mg ·L-1浓度下, 聚苯乙烯微塑料显著提高了水稻CAT和POD活性.
第一茬中, LDPE MPs胁迫使空心菜叶片的MDA含量显著降低, 结合上述微塑料胁迫下空心菜叶片抗氧化酶活性变化可推测, 空心菜对MPs胁迫所作出的氧化应激反应减轻了其细胞膜脂质的过氧化程度.另有研究表明, 在聚苯乙烯微塑料暴露下, 水生植物密刺苦草(Vallisneria denseserrulata)的SOD和POD等抗氧化酶活性显著提高, 而MDA含量显著降低[28], 与本研究表现出相同的结果.但是, 第二茬中, PE2处理的MDA含量显著高于其余处理, 这表明LDPE MPs对空心菜产生了氧化胁迫.Yang等[52]发现在高浓度的PE MPs胁迫下, 绿藻(Chlorella pyrenoidosa)MDA含量显著上升, 与本研究的结果相似.
另外, PE3显著提高空心菜叶片可溶性蛋白含量, 说明空心菜可主动增加和积累可溶性蛋白用于提高细胞的持水能力, 以缓解LDPE MPs的外界胁迫, 进一步保护自身重要物质和生物膜[53].同时, LDPE MPs可提高空心菜叶片可溶性糖含量, 这与本研究中空心菜地上生物量的显著增加结果一致, 表明LDPE MPs可通过提升可溶性糖这一重要的供能及缓冲物质含量, 缓解LDPE MPs带来的氧化胁迫, 进而促进空心菜地上生物量的累积[54].本研究还发现, 经LDPE MPs胁迫后, 第二茬空心菜叶片可溶性蛋白含量无显著差异, 而可溶性糖含量显著高于对照组, Dong等[55]的研究也呈现类似的结果, 这可能是由于可溶性糖含量的增加缓解了LDPE MPs的胁迫作用, 进而使可溶性蛋白质含量并未发生显著变化[18].本研究中空心菜第二茬叶片可溶性蛋白和可溶性糖含量均显著高于第一茬, 这与张亚楠等[56]研究的结果一致, 表明空心菜对连作条件做出抗逆反应.
4 结论(1) 本试验在0% ~10%质量分数范围内, LDPE MPs对空心菜种子活力具有显著的抑制作用, 且抑制效果随着LDPE MPs质量分数的升高而增强.LDPE MPs可能通过抑制空心菜种子活力进而影响种子的发芽, 因而种子发芽势同活力指数的变化趋势具有相似性.
(2) 5%质量分数的LDPE MPs可以降低土壤容重, 促进根系生长, 从而显著提高空心菜的生物量.
(3) 随着LDPE MPs质量分数(0% ~10%)的增加, 空心菜叶片抗氧化酶活性呈SOD活性先升后降, POD、CAT活性升高的趋势, 且可溶性蛋白和可溶性糖含量均有提升, 而MDA含量显著降低, 表明空心菜对LDPE MPs胁迫做出了氧化应激反应, 减轻了其细胞膜脂质的过氧化程度.
| [1] | Thompson R C, Olsen Y, Mitchell R P, et al. Lost at sea: where is all the plastic?[J]. Science, 2004, 304(5672). DOI:10.1126/science.1094559 |
| [2] |
廉宇航, 刘维涛, 史瑞滢, 等. 聚乙烯和聚乳酸微塑料对大豆生长和生理生化及代谢的影响[J]. 中国环境科学, 2022, 42(6): 2894-2903. Lian Y H, Liu W T, Shi R Y, et al. Impact of polyethylene and polylactic acid microplastics on growth, physio-biochemistry and metabolism in soybean (Glycine max)[J]. China Environmental Science, 2022, 42(6): 2894-2903. DOI:10.3969/j.issn.1000-6923.2022.06.045 |
| [3] | Rillig M C. Microplastic in terrestrial ecosystems and the soil?[J]. Environmental Science & Technology, 2012, 46(12): 6453-6454. |
| [4] | Qi R M, Jones D L, Li Z, et al. Behavior of microplastics and plastic film residues in the soil environment: a critical review[J]. Science of the Total Environment, 2020, 703. DOI:10.1016/j.scitotenv.2019.134722 |
| [5] | Kwon O Y, Kang J H, Hong S H, et al. Spatial distribution of microplastic in the surface waters along the coast of Korea[J]. Marine Pollution Bulletin, 2020, 155. DOI:10.1016/j.marpolbul.2019.110729 |
| [6] | Li J X, Gao F L, Zhang D, et al. Zonal distribution characteristics of microplastics in the southern Indian ocean and the influence of ocean current[J]. Journal of Marine Science and Engineering, 2022, 10(2). DOI:10.3390/jmse10020290 |
| [7] | Ta A T, Babel S. Microplastics pollution with heavy metals in the aquaculture zone of the Chao Phraya River Estuary, Thailand[J]. Marine Pollution Bulletin, 2020, 161. DOI:10.1016/j.marpolbul.2020.111747 |
| [8] | Zhang Q J, Liu T, Liu L, et al. Distribution and sedimentation of microplastics in Taihu Lake[J]. Science of the Total Environment, 2021, 795. DOI:10.1016/j.scitotenv.2021.148745 |
| [9] | Horton A A, Walton A, Spurgeon D J, et al. Microplastics in freshwater and terrestrial environments: evaluating the current understanding to identify the knowledge gaps and future research priorities[J]. Science of the Total Environment, 2017, 586: 127-141. DOI:10.1016/j.scitotenv.2017.01.190 |
| [10] |
李连祯, 周倩, 尹娜, 等. 食用蔬菜能吸收和积累微塑料[J]. 科学通报, 2019, 64(9): 928-934. Li L Z, Zhou Q, Yin N, et al. Uptake and accumulation of microplastics in an edible plant[J]. Chinese Science Bulletin, 2019, 64(9): 928-934. |
| [11] |
李瑞杰, 李连祯, 张云超, 等. 禾本科作物小麦能吸收和积累聚苯乙烯塑料微球[J]. 科学通报, 2020, 65(20): 2120-2127. Li R J, Li L Z, Zhang Y C, et al. Uptake and accumulation of microplastics in a cereal plant wheat[J]. Chinese Science Bulletin, 2020, 65(20): 2120-2127. |
| [12] |
杨杰, 李连祯, 周倩, 等. 土壤环境中微塑料污染: 来源、过程及风险[J]. 土壤学报, 2021, 58(2): 281-298. Yang J, Li L Z, Zhou Q, et al. Microplastics contamination of soil environment: sources, processes and risks[J]. Acta Pedologica Sinica, 2021, 58(2): 281-298. |
| [13] | Guo M, Zhao F R, Tian L W, et al. Effects of polystyrene microplastics on the seed germination of herbaceous ornamental plants[J]. Science of the Total Environment, 2022, 809. DOI:10.1016/j.scitotenv.2021.151100 |
| [14] | Bosker T, Bouwman L J, Brun N R, et al. Microplastics accumulate on pores in seed capsule and delay germination and root growth of the terrestrial vascular plant Lepidium sativum[J]. Chemosphere, 2019, 226: 774-781. DOI:10.1016/j.chemosphere.2019.03.163 |
| [15] |
张彦, 窦明, 邹磊, 等. 不同微塑料赋存环境对小麦萌发与幼苗生长影响研究[J]. 中国环境科学, 2021, 41(8): 3867-3877. Zhang Y, Dou M, Zou L, et al. Effects of different microplastics occurrence environment on seed germination and seedling growth of wheat (Triticum aestivum L.)[J]. China Environmental Science, 2021, 41(8): 3867-3877. DOI:10.3969/j.issn.1000-6923.2021.08.044 |
| [16] |
叶子琪, 蒋小峰, 汤其阳, 等. 聚乙烯微塑料对蚕豆幼苗的毒性效应[J]. 南京大学学报(自然科学), 2021, 57(3): 385-392. Ye Z Q, Jiang X F, Tang Q Y, et al. Toxic effects of polyethylene microplastics on higher plant Vicia faba[J]. Journal of Nanjing University (Natural Science), 2021, 57(3): 385-392. |
| [17] | Pignattelli S, Broccoli A, Renzi M. Physiological responses of garden cress (L. sativum) to different types of microplastics[J]. Science of the Total Environment, 2020, 727. DOI:10.1016/j.scitotenv.2020.138609 |
| [18] |
邱陈陈, 李国新, 李青松, 等. 聚苯乙烯纳米塑料对大蒜生长生理的影响[J]. 环境科学, 2022, 43(8): 4387-4393. Qiu C C, Li G X, Li Q S, et al. Effects of polystyrene nanoplastics (PS-NPs) on the physiology of Allium sativum L.[J]. Environmental Science, 2022, 43(8): 4387-4393. |
| [19] | Hernández-Arenas R, Beltrán-Sanahuja A, Navarro-Quirant P, et al. The effect of sewage sludge containing microplastics on growth and fruit development of tomato plants[J]. Environmental Pollution, 2021, 268. DOI:10.1016/j.envpol.2020.115779 |
| [20] | Zhang Q G, Zhao M S, Meng F S, et al. Effect of polystyrene microplastics on rice seed germination and antioxidant enzyme activity[J]. Toxics, 2021, 9(8). DOI:10.3390/toxics9080179 |
| [21] | Gao M L, Liu Y, Song Z G. Effects of polyethylene microplastic on the phytotoxicity of di-n-butyl phthalate in lettuce (Lactuca sativa L. var. ramosa Hort)[J]. Chemosphere, 2019, 237. DOI:10.1016/j.chemosphere.2019.124482 |
| [22] | Li J, Zheng X Q, Liu K C, et al. Effect of tetracycline on the growth and nutrient removal capacity of Chlamydomonas reinhardtii in simulated effluent from wastewater treatment plants[J]. Bioresource Technology, 2016, 218. DOI:10.1016/j.biortech.2016.07.080 |
| [23] | Cao D J, Shi X D, Li H, et al. Effects of lead on tolerance, bioaccumulation, and antioxidative defense system of green algae, cladophora[J]. Ecotoxicology and Environmental Safety, 2015, 112: 231-237. DOI:10.1016/j.ecoenv.2014.11.007 |
| [24] |
黄献培, 向垒, 郭静婕, 等. 聚苯乙烯微球对菜心种子及幼苗的毒性效应[J]. 农业环境科学学报, 2021, 40(5): 926-933. Huang X P, Xiang L, Guo J J, et al. Toxicity of polystyrene microplastics on seeds and seedlings of Brassica campestris L.[J]. Journal of Agro-Environment Science, 2021, 40(5): 926-933. |
| [25] | Yu H W, Qi W X, Cao X F, et al. Microplastic residues in wetland ecosystems: do they truly threaten the plant-microbe-soil system?[J]. Environment International, 2021, 156. DOI:10.1016/j.envint.2021.106708 |
| [26] | Yu H W, Zhang X L, Hu J W, et al. Ecotoxicity of polystyrene microplastics to submerged carnivorous Utricularia vulgaris plants in freshwater ecosystems[J]. Environmental Pollution, 2020, 265. DOI:10.1016/j.envpol.2020.114830 |
| [27] | Dong Y M, Gao M L, Song Z G, et al. Microplastic particles increase arsenic toxicity to rice seedlings[J]. Environmental Pollution, 2020, 259. DOI:10.1016/j.envpol.2019.113892 |
| [28] | Abduro Ogo H, Tang N, Li X W, et al. Combined toxicity of microplastic and lead on submerged macrophytes[J]. Chemosphere, 2022, 295. DOI:10.1016/j.chemosphere.2022.133956 |
| [29] | Sudhakar M P, Kumar B R, Mathimani T, et al. A review on bioenergy and bioactive compounds from microalgae and macroalgae-sustainable energy perspective[J]. Journal of Cleaner Production, 2019, 228: 1320-1333. DOI:10.1016/j.jclepro.2019.04.287 |
| [30] | Wang Y P, Zhang Z J, Wang X C, et al. Exogenous dopamine improves apple fruit quality via increasing flavonoids and soluble sugar contents[J]. Scientia Horticulturae, 2021, 280. DOI:10.1016/j.scienta.2021.109903 |
| [31] | Dong Y M, Song Z G, Liu Y, et al. Polystyrene particles combined with di-butyl phthalate cause significant decrease in photosynthesis and red lettuce quality[J]. Environmental Pollution, 2021, 278. DOI:10.1016/j.envpol.2021.116871 |
| [32] | Li Z X, Li Q F, Li R J, et al. The distribution and impact of polystyrene nanoplastics on cucumber plants[J]. Environmental Science and Pollution Research, 2021, 28(13): 16042-16053. DOI:10.1007/s11356-020-11702-2 |
| [33] | Huang Y, Liu Q, Jia W Q, et al. Agricultural plastic mulching as a source of microplastics in the terrestrial environment[J]. Environmental Pollution, 2020, 260. DOI:10.1016/j.envpol.2020.114096 |
| [34] |
杨光蓉, 陈历睿, 林敦梅. 土壤微塑料污染现状、来源、环境命运及生态效应[J]. 中国环境科学, 2021, 41(1): 353-365. Yang G R, Chen L R, Lin D M. Status, sources, environmental fate and ecological consequences of microplastic pollution in soil[J]. China Environmental Science, 2021, 41(1): 353-365. |
| [35] | Ng E L, Lin S Y, Dungan A M, et al. Microplastic pollution alters forest soil microbiome[J]. Journal of Hazardous Materials, 2021, 409. DOI:10.1016/j.jhazmat.2020.124606 |
| [36] |
许学慧, 胡海娜, 陈颖. 聚乙烯微塑料对大豆生长的影响[J]. 中国土壤与肥料, 2021(6): 262-268. Xu X H, Hu H N, Chen Y. Study on the effect of polyethylene microplastics on soybean growth[J]. Soil and Fertilizer Sciences in China, 2021(6): 262-268. |
| [37] | Fuller S, Gautam A. A procedure for measuring microplastics using pressurized fluid extraction[J]. Environmental Science & Technology, 2016, 50(11): 5774-5780. |
| [38] |
李鹏飞, 侯德义, 王刘炜, 等. 农田中的(微)塑料污染: 来源、迁移、环境生态效应及防治措施[J]. 土壤学报, 2021, 58(2): 314-330. Li P F, Hou D Y, Wang L W, et al. (Micro)plastics pollution in agricultural soils: sources, transportation, ecological effects and preventive strategies[J]. Acta Pedologica Sinica, 2021, 58(2): 314-330. |
| [39] |
张宇阳, 于涛, 马文宝, 等. 不同郁闭度对野外回归的梓叶槭幼树形态和生理特征的影响[J]. 生物多样性, 2020, 28(3): 323-332. Zhang Y Y, Yu T, Ma W B, et al. Physiological and morphological effects of different canopy densities on reintroduced Acer catalpifolium[J]. Biodiversity Science, 2020, 28(3): 323-332. |
| [40] | Pflugmacher S, Tallinen S, Kim Y J, et al. Ageing affects microplastic toxicity over time: effects of aged polycarbonate on germination, growth, and oxidative stress of Lepidium sativum[J]. Science of the Total Environment, 2021, 790. DOI:10.1016/j.scitotenv.2021.148166 |
| [41] |
吴佳妮, 杨天志, 连加攀, 等. 聚苯乙烯纳米塑料(PSNPs)对大豆(Glycine max)种子发芽和幼苗生长的影响[J]. 环境科学学报, 2020, 40(12): 4581-4589. Wu J N, Yang T Z, Lian J P, et al. Effects of polystyrene nanoplastics (PSNPs) on seed germination and seedling growth of soybean (Glycine max)[J]. Acta Scientiae Circumstantiae, 2020, 40(12): 4581-4589. |
| [42] | Baker N R. Chlorophyll fluorescence: a probe of photosynthesis in vivo[J]. Annual Review of Plant Biology, 2008, 59: 89-113. |
| [43] | Moinuddin M, Gulzar S, Hameed A, et al. Differences in photosynthetic syndromes of four halophytic marsh grasses in Pakistan[J]. Photosynthesis Research, 2017, 131(1): 51-64. |
| [44] | de Souza Machado A A, Lau C W, Till J, et al. Impacts of microplastics on the soil biophysical environment[J]. Environmental Science & Technology, 2018, 52(17): 9656-9665. |
| [45] | de Souza Machado A A, Lau C W, Kloas W, et al. Microplastics can change soil properties and affect plant performance[J]. Environmental Science & Technology, 2019, 53(10): 6044-6052. |
| [46] | Li Z X, Li Q F, Li R J, et al. Physiological responses of lettuce (Lactuca sativa L.) to microplastic pollution[J]. Environmental Science and Pollution Research, 2020, 27(24): 30306-30314. |
| [47] | van Weert S, Redondo-Hasselerharm P E, Diepens N J, et al. Effects of nanoplastics and microplastics on the growth of sediment-rooted macrophytes[J]. Science of the Total Environment, 2019, 654: 1040-1047. |
| [48] | Zhang G S, Liu Y F. The distribution of microplastics in soil aggregate fractions in Southwestern China[J]. Science of the Total Environment, 2018, 642: 12-20. |
| [49] | Zhou C Q, Lu C H, Mai L, et al. Response of rice (Oryza sativa L.) roots to nanoplastic treatment at seedling stage[J]. Journal of Hazardous Materials, 2021, 401. DOI:10.1016/j.jhazmat.2020.123412 |
| [50] |
安菁, 刘欢语, 郑艳, 等. 土壤微塑料残留对大豆幼苗生长及生理生化特征的影响[J]. 四川农业大学学报, 2021, 39(1): 41-46, 113. An J, Liu H Y, Zheng Y, et al. Effects of soil microplastics residue on soybean seedlings growth and the physiological and biochemical characteristics[J]. Journal of Sichuan Agricultural University, 2021, 39(1): 41-46, 113. |
| [51] | Gao K, Li B, Chen R Z, et al. A feasibility study of using silkworm larvae as a novel in vivo model to evaluate the biotoxicity of ionic liquids[J]. Ecotoxicology and Environmental Safety, 2021, 209. DOI:10.1016/j.ecoenv.2020.111759 |
| [52] | Yang W F, Gao X X, Wu Y X, et al. The combined toxicity influence of microplastics and nonylphenol on microalgae Chlorella pyrenoidosa[J]. Ecotoxicology and Environmental Safety, 2020, 195. DOI:10.1016/j.ecoenv.2020.110484 |
| [53] | Zhu Z L, Wang S C, Zhao F F, et al. Joint toxicity of microplastics with triclosan to marine microalgae Skeletonema costatum[J]. Environmental Pollution, 2019, 246: 509-517. |
| [54] |
李月灵, 金则新, 罗光宇, 等. 干旱胁迫下接种丛枝菌根真菌对七子花非结构性碳水化合物积累及C、N、P化学计量特征的影响[J]. 应用生态学报, 2022, 33(4): 963-971. Li Y L, Jin Z X, Luo G Y, et al. Effects of arbuscular mycorrhizal fungi inoculation on non-structural carbohydrate contents and C: N: P stoichiometry of Heptacodium miconioides under drought stress[J]. Chinese Journal of Applied Ecology, 2022, 33(4): 963-971. |
| [55] | Dong J W, Li L Q, Liu Q Q, et al. Interactive effects of polymethyl methacrylate (PMMA) microplastics and salinity variation on a marine diatom Phaeodactylum tricornutum[J]. Chemosphere, 2022, 289. DOI:10.1016/j.chemosphere.2021.133240 |
| [56] |
张亚楠, 王兴祥, 李孝刚, 等. 连作对棉花抗枯萎病生理生化特性的影响[J]. 生态学报, 2016, 36(14): 4456-4464. Zhang Y N, Wang X X, Li X G, et al. Effects of continuous cropping on physiological and biochemical resistance of cotton to Fusarium wilt[J]. Acta Ecologica Sinica, 2016, 36(14): 4456-4464. |