 2023, Vol. 44
2023, Vol. 44


植被净初级生产力(net primary productivity, NPP), 是指植被在单位时间和单位面积内所储存的有机质总量, 可反映植被在自然环境中受气候、地形和人文因子综合影响下的生产能力[1~3].植被NPP兼具时间上的易变性和空间上的异质性, 是解释区域陆地植被生态系统生产能力和生态环境动态演变的重要指标[4~6].植被NPP在植被固碳[7]、碳平衡调节[8]、生态系统环境质量评价[6]和生态环境保护[9]等领域的研究中有重要意义.
已有研究表明, 植被NPP变化主要受气候变化和人类活动的共同影响, 但影响程度和范围表现出明显的时空差异[10~13].随着研究的逐步深入, 如何定量剥离气候变化和人类活动对植被NPP变化的影响成为研究的热点.已有研究大多采用多元线性回归分析和残差分析实现气候变化和人类活动对植被NPP影响的定量分离.例如, Ge等[14]、Zhang等[15]、张心茹等[16]、刘婧等[17]和Peng等[18]基于植被NPP、降水、气温和太阳辐射等建立多元回归分析和残差分析模型, 分别在中国全境和区域(洞庭湖流域、黄河三角洲、滦河流域和祁连山国家公园)尺度上定量剥离了气候变化和人类活动对植被NPP的影响.以上研究表明, 植被NPP变化受气候变化和人类活动的贡献呈现出明显的时空差异.例如, 全国范围内气候变化和人类活动对2001~2016年植被生长均具有促进作用, 且气候变化的促进作用强于人类活动[14]; 区域尺度上, 气候变化和人类活动对植被生长均具有促进作用, 气候变化对2000~2019年洞庭湖流域和祁连山国家公园植被生长的作用强于人类活动[15, 18], 而人类活动对2000~2017年黄河三角洲和2000~2015年滦河流域植被生长的作用强于气候变化[16, 17].综上可知, 植被NPP时空演变主要受区域气候条件和人类活动强度的影响, 而多元线性回归分析和残差分析可以有效探明气候变化和人类活动对植被生长的定量影响.以上研究多采用气温、降水和太阳辐射等气象因子作为多元回归分析模型的输入变量, 定量剥离气候变化和人类活动对植被生长的影响, 但区域植被生长往往还受日照时数、相对湿度和生物干湿度指数等气象因子的影响[4, 19~21], 且植被生长对气象因子的变化存在明显的滞后期和地域差异[20~22].因此, 在定量剥离气候变化和人类活动的影响时应该充分考虑区域气候条件和植被生长对气象因子的滞后效应.
西南地区是我国喀斯特地貌的主要分布区, 部分地区地表崎岖, 基岩裸露, 水土流失严重, 为典型的生态环境脆弱区[23].该地区植被生长对气候变化和人类活动响应敏感, 生态环境问题已成为限制该地区发展的主要原因之一.在全球气候变暖[24]、林业生态工程实施[25]和“双碳”战略[26]背景下, 西南地区不同植被生态系统NPP时空变化及其与气候变化和人类活动之间的关系亟待厘清.因此, 本文以西南地区为研究区, 首先, 采用相关分析和主成分分析, 在顾及滞后效应的基础上筛选影响植被NPP变化的主导气象因子; 然后, 基于植被NPP和最大相关系数对应滞后期的主导气象因子时间序列数据, 建立多元线性回归分析和残差分析模型, 探明气候变化和人类活动对西南地区植被NPP的影响; 最后, 基于偏相关分析和转移矩阵, 揭示植被NPP对气候变化和土地利用类型变化的响应特征, 以期为西南地区环境保护、生态修复和生态林业工程的实施等提供理论依据.
1 研究区概况研究区位于我国西南部, 位于97°21′~112°04′E、20°54′~34°19′N之间, 包括云南省、贵州省、四川省、广西壮族自治区和重庆市, 总面积为137.63万km2, 约占我国陆表总面积的14.33%.区域内植被类型丰富, 植被覆盖度较高, 根据2020年土地利用类型数据统计可知, 西南地区陆地植被生态系统占比为96.41%, 其中农田、森林和草地生态系统分别占23.80%、48.80%、23.80%, 其余生态系统仅占3.59%(图 1).气候主要为亚热带季风气候和热带季风气候, 大部分地区雨热同期, 降水充沛, 气温和降水均呈现由东南往西北逐渐递减的特征, 年均气温为13.12~14.09℃, 年均降水量为954.33~2 306.93 mm.研究区内地貌类型复杂多样, 包含丘陵、盆地、高原和山地, 是我国喀斯特地貌的主要分布区, 也是生态环境脆弱区之一.

|
图 1 2020年西南地区植被生态系统空间分布示意 Fig. 1 Spatial distribution of vegetation ecosystems in southwest China in 2020 |
本文使用美国国家航空航天局网站(https://urs.earthdata.nasa.gov/)提供的2000~2021年的MOD17A3 NPP数据, 数据原始格式为HDF, 时间分辨率为每年, 空间分辨率为500 m, 该数据利用MODIS/TERRA卫星遥感参数, 通过BIOME-BGC生态过程模型模拟得到全球植被净初级生产力数据集, 该数据集已在全球和区域植被生产力研究中得到了广泛应用.首先, 使用MRT工具对HDF格式的数据产品进行了批量拼接、重采样和重投影; 然后, 乘以比例因子0.1, 并将少量异常值赋值为空值; 最后, 基于研究区矢量数据批量裁剪得到2000~2021年西南地区植被NPP时间序列.
2.1.2 气象数据气象数据来源于国家气象科学数据中心网站(https://data.cma.cn/), 该平台提供1999~2021年中国2 416个气象站点的日观测数据.首先, 对各气象因子进行异常值处理, 对各站点年缺失值少于5%的数据使用线性内插法进行插补, 否则对该站点该年数据进行舍弃; 然后, 基于插补后的数据集计算得到月平均地表温度、月累积降水、月平均大气压强、月累积日照时数、月平均相对湿度、月平均最高气温、月平均气温和月平均最低气温, 并使用Anusplin气象插值模型, 选用三变量薄盘光滑样条函数, 在顾及经度、纬度和高程的基础上插值得到1999~2021年各气象因子月时间序列; 接下来, 基于月平均气温和月累积降水时间序列, 计算得到可能蒸散率、可能蒸散量、生物干湿度指数和徐文铎湿度指数时间序列[27]; 最后, 基于研究区矢量数据, 裁剪得到1999~2021年西南地区各气象因子时间序列.以上气象数据空间分辨率均为500 m.
2.1.3 土地利用数据土地利用数据和行政区矢量数据来源于中国科学院资源环境科学数据中心网站(https://www.resdc.cn/).采用2000、2005、2010、2015和2020年的中国土地利用现状遥感监测数据, 空间分辨率为1 km.为保证数据空间分辨率一致, 本文将土地利用类型数据重采样到500 m.由于土地利用数据每5 a为一期, 本文使用2000年土地利用数据代表2000、2001和2002年, 2005年代表2003、2004、2005、2006和2007年, 依次类推, 得到2000~2021年各年份土地利用类型的代表年份.该产品将土地利用类型划分为6个一级类型, 分别为耕地, 林地, 草地, 水体, 城乡、工矿、居民地和未利用土地, 本文根据以上土地利用类型提取出农田、森林和草地生态系统数据.
2.2 研究方法 2.2.1 主导气象因子筛选考虑到植被生长是一个相对缓慢的过程, 植被生长对各气象因子响应程度不一且具有一定的滞后效应[20~22].因此, 本文考虑影响植被生长的主导气象因子及其滞后效应.影响植被NPP变化的主导气象因子筛选步骤如下[21]:首先, 采用相关分析法计算西南地区植被NPP与前0~12月各气象因子时间序列的相关系数; 然后, 利用一阶偏导法求得最大相关系数; 最后, 根据最大相关系数得到植被NPP对各气象因子变化最大响应的滞后期, 以此得到西南地区植被NPP与各气象因子的最大相关系数及其对应的滞后期.植被NPP与气象因子最大相关系数及滞后期计算结果如表 1所示.
|
|
表 1 植被NPP与气象因子最大相关系数及滞后期 Table 1 Maximum correlation coefficient and its lag time between vegetation NPP and climate factors |
植被动态变化受多种气象因子的共同影响[20, 21, 27], 但数据过多会造成信息冗余, 也会给统计分析带来一定的难度.因此, 本文通过主成分分析实现主导气象因子的进一步筛选[27].基于植被NPP和各气象因子最大相关系数对应滞后期时间序列提取出的各主成分特征值、方差贡献率和累积贡献率, 结果如表 2所示.本文共提取影响植被NPP变化的4个主成分, 其特征值均大于1, 贡献率分别为47.46%、23.14%、11.25%和8.77%, 累计贡献率达到90.62%, 可有效反映影响植被NPP时空演变的主要气象因子信息.在第1、2、3和4主成分中, 信息负荷值最大的分别为平均气温(0.984)、降水(0.988)、日照时数(-0.784)和生物干湿度指数(-0.876).因此, 本文选取以上4个气象因子作为影响西南地区植被NPP变化的主导气象因子.
|
|
表 2 植被NPP与气象因子主成分分析 Table 2 Principal component analysis between vegetation NPP and climate factors |
2.2.2 多元线性回归分析和残差分析
本文使用多元线性回归分析和残差分析实现气候变化和人类活动对植被NPP变化的定量厘定[20, 21, 28].以平均气温(Tem)、降水(Pre)、日照时数(SD)和生物干湿度指数(BK)这4个主导气象因子为多元线性回归模型的输入因子, 基于矩阵式(2)求得的多元线性回归模型回归系数, 得到植被NPP预测值(NPPPre), 即气候变化影响下的植被NPP[式(1)].在不考虑其他因素影响的情况下, 植被NPP残差值(NPPRes)为植被NPP与植被NPP预测值之差, 视为人类活动影响所产生的植被NPP[式(3)].
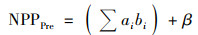
|
(1) |
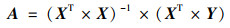
|
(2) |
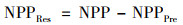
|
(3) |

|
图 2 2000~2021年西南地区植被NPP时间变化特征 Fig. 2 Temporal variation in vegetation NPP in southwest China from 2000 to 2021 |
式中, NPPPre表示植被NPP预测值; NPPRes表示植被NPP残差值; NPP为MODIS NPP, 即植被NPP; bi为线性回归方差的输入因子; ai为方程输入因子所对应的回归系数; i为主导气象因子; β为常数项; A为输出因子NPPPre的合集; X为输入因子的合集; XT为输入因子合集的转置矩阵.本文中, 平均气温(Tem)、降水(Pre)、日照时数(SD)和生物干湿度指数(BK)为方程输入因子.
2.2.3 Theil-Sen Median趋势分析和Mann-Kendall显著性检验Theil-Sen Median趋势分析法是一种非参数趋势估算方法[29], 抗噪性强, 不易受误差值和异常值的影响, 适用于长时间序列数据的趋势分析.β为变化斜率, 当β>0时, 表示植被NPP在时间序列上呈上升趋势; 当β<0时, 表示植被NPP在时间序列上呈下降趋势; 当β=0, 表示植被NPP在时间序列上无变化.Mann-Kendall显著性检验法为广泛使用的一种非参数检验方法, 不对数据采用何种分布方式和规律作要求.本文拟将植被NPP变化趋势分为3个等级, 未通过0.05置信水平检验为不显著变化; 0.05置信水平下为显著变化; 0.01置信水平下为极显著变化.
2.2.4 偏相关分析偏相关分析[20]可以控制其他气象因子的影响, 达到研究植被NPP与单一气象因子相关性的目的.根据计算得到的偏相关系数, 利用t检验法判断其相关性是否显著[21], 分别当P≥0.05、0.01≤P<0.05和P<0.01, 认为植被NPP与气象因子分别呈不显著相关、显著相关和极显著相关.
3 结果与分析 3.1 西南地区植被NPP时间变化特征由图 2(a)可知, 2000~2021年西南地区植被NPP呈波动上升趋势, 上升速率为3.54 g·(m2·a)-1, 植被NPP多年平均值为743.46 g·m-2.植被NPP在2004年和2010年呈下降趋势. 2000~2021年农田、森林和草地生态系统NPP均呈波动上升趋势, 其中农田生态系统NPP上升趋势最显著, 上升速率为6.31g·(m2·a)-1, 草地生态系统上升速率次之, 为3.05g·(m2·a)-1, 森林生态系统上升速率最低, 为2.60g·(m2·a)-1.森林生态系统多年植被NPP平均值最高, 为866.65 g·m-2, 农田生态系统次之, 草地生态系统最低.由图 2(b)可知, 气候变化的影响下, 2000~2021年西南地区植被NPPPre上升速率为1.46 g·(m2·a)-1. 2000~2021年西南地区农田、森林和草地生态系统均呈波动上升趋势, 农田生态系统NPPPre上升斜率高于森林和草地生态系统. 研究时段内西南地区森林生态系统相比于农田和草地植被生态系统更稳定, 植被NPPPre变化区间较小.由图 2(c)可知, 人类活动影响下, 2000~2021年西南地区植被NPPRes上升速率为2.08 g·(m2·a)-1.农田、森林和草地生态系统NPPRes均呈波动上升趋势, 其中, 农田生态系统NPPRes上升趋势最为显著, 为3.66g·(m2·a)-1, 森林和草地生态系统NPPRes上升斜率较为接近, 分别为1.60g·(m2·a)-1和1.63g·(m2·a)-1.

|
图 3 2000~2021年西南地区植被NPP空间变化特征 Fig. 3 Spatial variation in vegetation NPP in southwest China from 2000 to 2021 |
由上文可知, 2000~2021年西南地区及各陆地植被生态系统NPP均呈上升趋势, 其中, 农田生态系统NPP上升趋势最为显著, 但森林生态系统NPP多年均值高于农田和草地生态系统.气候变化和人类活动影响下, 西南地区各陆地植被生态系统NPP均呈上升趋势, 农田生态系统的上升趋势明显大于森林和草地生态系统.研究时段内森林生态系统NPP相比于农田和草地生态系统变化幅度更小, 更为稳定.
3.2 西南地区植被NPP空间变化特征由图 3(a)和3(d)可知, 2000~2021年西南地区植被NPP变化斜率在-67.33~77.27g·(m2·a)-1之间.西南地区植被NPP呈上升和下降趋势的面积占比分别为89.06%和10.94%, 其中, 呈显著上升和极显著上升的面积占比为47.27%, 主要分布在广西南部、四川东部和重庆西部, 以及云南和贵州交界处.农田、森林和草地生态系统NPP呈上升趋势的面积占比分别为97.00%、84.47%和88.10%, 均远大于呈下降趋势的面积占比, 其中, 农田生态系统呈显著上升和极显著上升的面积占比高达87.86%, 而森林和草地生态系统呈显著上升和极显著上升的区域仅分别占其总面积的36.17%和31.43%.
由图 3(b)和3(e)可知, 气候变化影响下, 2000~2021年西南地区植被NPPPre变化斜率为-55.36~63.84g·(m2·a)-1.气候变化导致西南地区植被NPPPre呈上升趋势和下降趋势的区域分别占总面积的82.54%和17.46%, 其中, 呈极显著上升和显著上升趋势的区域占42.82%, 主要分布在四川西北部和东部、贵州西部、云南东北部、重庆西部和北部; 呈极显著下降和显著下降趋势的区域仅占2.14%, 零星分布在四川中部、云南中南部和广西东北部.农田和草地生态系统NPPPre呈上升趋势的区域分别占其总面积的94.13%和81.15%, 高于森林生态系统(75.34%).

|
图 4 西南地区植被NPP与气象因子最大偏相关关系空间分布 Fig. 4 Spatial distribution of maximum partial correlation between vegetation NPP and climate factors in southwest China |
由图 3(c)和3(f)可知, 人类活动影响下, 2000~2021年西南地区植被NPPRes变化斜率为-62.76~81.51g·(m2·a)-1.植被NPPRes呈上升趋势的区域占78.86%, 明显大于呈下降趋势区域的面积占比.植被NPPRes呈极显著上升和显著上升的区域占31.81%, 主要分布在四川东北部、重庆西南部和广西中部.农田和生态系统NPPRes呈上升趋势的区域分别占其总面积的95.29%, 高于森林(75.74%)和草地生态系统(71.64%), 其中, 农田生态系统NPPRes呈极显著上升和显著上升的区域占其总面积的76.16%, 而森林和草地生态系统仅分别占24.09%和10.67%.
综上可知, 西南地区陆地植被生态系统NPP绝大部分区域呈上升趋势, 其中, 农田生态系统NPP呈上升趋势的面积占比和强度均高于森林和草地生态系统.整体上, 气候变化和人类活动对西南地区植被NPP促进作用大于抑制作用.气候变化和人类活动对农田生态系统促进作用面积占比均最大, 说明农田生态系统更易受气候变化和人类活动的影响.
3.3 西南地区植被NPP变化驱动因素分析 3.3.1 植被NPP与气象因子的偏相关关系由图 4(a)~4(d)可知, 西南地区植被NPP与平均气温的最大偏相关系数在-0.943~0.934之间, 总体呈东北高、西南低的空间分布格局; 与降水的最大偏相关系数在-0.919~0.948之间, 高值区主要分布在四川省东部、云南和贵州交界处; 与日照时数的最大偏相关系数在-0.922~0.952之间, 高值区主要分布在四川和云南西部, 低值区主要分布在四川东部、重庆西部, 以及广西、云南和贵州大部分地区; 与生物干湿度指数的最大偏相关系数在-0.948~0.913之间, 高值区主要分布在西南地区东北部.
由图 4(e)~4(h)可知, 区域尺度上, 西南地区植被NPP与平均气温、降水和日照时数最大偏相关系数呈正相关的面积占比大于呈负相关的面积占比, 而与生物干湿度指数呈负相关的面积占比大于呈正相关的面积占比.西南地区植被NPP与平均温度呈正相关的面积占比最高, 为76.27%, 其中, 呈极显著和显著上升的面积占比为18.14%, 高于其他气象因子, 主要分布在四川西北部, 以及云南、贵州、重庆和四川交界处.植被生态系统尺度上, 除农田生态系统NPP与日照时数最大偏相关系数呈负相关的面积占比大于呈正相关的面积占比外, 与区域尺度结果类似, 农田、森林和草地生态系统NPP与平均气温、降水和日照时数最大偏相关系数呈正相关的面积占比大于呈负相关的面积占比, 而与生物干湿度指数呈负相关的面积占比大于呈正相关的面积占比, 且农田、森林和草地生态系统NPP与平均气温呈正相关的面积占比均最高, 分别为75.84%、73.60%和79.36%.此外, 草地生态系统NPP与平均气温、降水和日照时数最大偏相关系数呈正相关的面积占比大于农田和森林生态系统, 分别为79.36%、60.92%和69.16%.农田生态系统NPP与日照时数呈负相关的面积占比为76.98%, 其中, 呈极显著负相关和显著负相关的面积占比高达24.29%, 远高于森林(7.08%)和草地生态系统(2.66%).
综上可知, 气候变化对西南地区植被NPP变化具有双重作用, 且作用强度呈明显地域差异.区域尺度上, 平均气温、降水和日照时数对西南地区植被NPP变化以正向促进作用为主, 而生物干湿度指数则以负向抑制作用为主.植被生态系统尺度上, 日照时数对农田生态系统NPP变化的抑制作用大于促进作用, 而对森林和草地生态系统NPP变化的促进作用大于抑制作用.平均气温、降水和日照时数对草地生态系统NPP变化的促进作用强于农田和森林生态系统.
3.3.2 植被NPP与土地利用变化关系由表 3可知, 2000~2020年西南地区土地利用类型发生变化的面积为473 601 km2, 占西南地区总面积的34.88%, 其中, 耕地和林地的相互转换占西南地区土地利用变化总量的38.34%, 是这期间最主要的土地利用类型变化.西南地区整体上呈现出耕地、草地和水域被占用, 而林地、建设用地和未利用地面积增加的变化特征.西南地区植被NPP呈上升趋势的区域中主要以耕地、林地和草地这3个地类之间的相互转移为主.在耕地转变为林地和草地的区域中植被NPP呈上升趋势的区域占83.7%, 而其他用地转变为建设用地中植被NPP呈下降趋势的区域占43.11%, 高于其他地类转为水域(33.15%)、林地(19.73%)、草地(18.75%)、耕地(15.46%)和未利用地(13.68%)区域中植被NPP呈下降趋势的占比.西南地区其他用地转为建设用地的土地利用变化主要发生在四川中部、云南中东部、贵州中部和广西东部, 与植被NPP呈下降趋势区域的空间分布具有较好的空间耦合性.以上区域快速城市化和工业化导致耕地、林地和草地等向建设用地转化是西南地区植被NPP下降的主要原因之一, 而耕地转为林地和草地的区域中植被NPP多呈上升趋势.
|
|
表 3 2000~2020年西南地区土地利用转移矩阵/km2 Table 3 Land use transition matrix in southwest China from 2000 to 2020/km2 |
4 讨论 4.1 西南地区植被NPP时空变化特征
在时间变化上, 2000~2021年西南地区植被NPP整体上呈波动上升趋势, 这与已有研究的结果一致[30~32].植被NPP在2004年和2010年呈明显下降趋势, 这与2003年为全球自然灾害重灾年和2009年我国西南地区发生的极端干旱气候有关.干旱会加剧土壤水分蒸发, 叶片气孔导度降低, 使植被的生长面临水分胁迫, 从而对植被NPP产生负向影响[33, 34], 加之植被生长对气候变化的响应存在滞后效应[20, 21], 因此, 2004年和2010年西南地区植被NPP呈下降趋势.研究时段内陆地植被生态系统NPP均呈波动上升趋势, 其中, 农田生态系统NPP上升趋势最显著, 其次为草地生态系统和森林生态系统.得益于科学的田间管理方式和集约化农业的进步, 天然林保护、植树造林、退耕还林和退牧还草等生态林业工程的实施, 使得西南地区陆地植被生态NPP均呈上升趋势[20, 21, 25].
在空间变化上, 西南地区植被NPP呈上升趋势和下降趋势的面积占比分别为89.06%和10.94%.植被NPP呈下降趋势的区域主要分布在云南、贵州和广西, 其中, 呈显著下降和极显著下降的区域主要分布于云南西南部和广西东北部.下降的原因主要有以下两个方面:一方面是由于城市化进程加快, 例如, 据广西统计年鉴(2006~2021年), 广西城市化率由2000年的28.15%提高至2020年的54.20%, 而位于广西东北部的桂林市的城市化率更是由2000年的22.95%提高至2020年的52.58%, 城市化进程的加快, 使得大量耕地、林地和草地转化为建设用地, 植被覆盖度下降, 从而导致植被NPP降低; 另一方面, 已有研究表明, 2001~2019年广西、贵州和云南森林面积分别减少了22 802.04、3 145.47和8 896.76 km2, 森林资源的锐减, 导致植被覆盖度下降, 水土流失问题加剧, 生态环境质量恶化, 从而导致植被NPP下降[35, 36].
4.2 气候变化和人类活动影响下西南地区植被NPP时空变化特征气候变化影响下, 2000~2021年西南地区陆地植被生态系统NPPPre均呈波动上升趋势, 其中, 森林生态系统NPPPre的上升趋势和波动幅度均最小.农田和草地生态系统NPPPre呈上升趋势的面积占比均高于森林生态系统.这是由于森林生态系统相比于其他植被生态系统抵御气候变化的能力更强, 特别是常绿阔叶林具有较低的植被NPP脆弱性, 光能利用率高于其他类型植被, 自我修复能力强, 较为稳定, 而农田和草地承载力较弱, 抵御气候变化的能力就较低, 易受气候变化影响[14, 37].
人类活动影响下, 西南地区陆地植被NPPRes均呈波动上升趋势, 其中农田生态系统NPPRes上升趋势最为显著, 波动范围最大.农田、森林和草地生态系统NPPRes呈上升趋势的面积占比远大于呈下降趋势的面积占比.农田生态系统NPPRes呈上升趋势的区域主要分布在四川盆地, 这主要是由于该地区农田生态系统以农作物为主, 且近年来由于农田灌溉水利设施的建设、排水工程的完善以及田间管理技术的进步等都在一定程度上对农作物的生长产生影响, 从而影响到植被NPP的时空变化, 这与已有结果高度一致[38, 39].
4.3 西南地区植被NPP对驱动因素的响应特征区域尺度上, 平均气温、降水和日照时数对西南地区植被NPP变化以正向促进作用为主, 其中, 平均气温对西南地区植被NPP呈正向作用的面积占比高于其他气象因子.这是由于西南地区降水丰沛, 日照条件充足, 2000~2021年西南地区区域平均降水量在954.33~2 306.93 mm之间, 日照时数在1 637.00~1 880.00 h之间, 丰沛的降水和充足的日照时数降低了植被NPP对其变化响应的敏感性, 加之西南地区地形起伏较大, 平均气温变化幅度较大且呈地域差异明显, 因此, 平均气温对西南地区植被NPP呈正向作用强于其他气象因子, 这与已有研究的结果一致[31, 40, 41].
植被生态系统尺度上, 平均气温、降水和日照时数对草地生态系统NPP变化的促进作用强于农田和森林生态系统.一方面, 草地主要分布在西南地区西北部, 而降水和温度均呈东南高西北低的空间分布特征, 因此, 西南地区西北部降水资源较东南部少, 且温度较低, 因此, 草地生态系统NPP变化受水、热和光等条件的正向影响更加明显, 这与已有研究结果一致[42]; 另一方面, 由于农田生态系统受高效农业工程和科学田间管理的影响, 森林生态系统由于其生态多样性更加丰富, 植被群落结构更加稳定, 使得农田和森林生态系统对气候变化的响应敏感性低于草地生态系统.因此, 相较于农田和森林生态系统, 草地生态系统对气候变化响应更为敏感[42, 43].草地生态系统NPP与平均气温、降水和日照时数呈正相关的面积占比高于农田和森林生态系统.
西南地区土地利用类型之间的转移对植被NPP变化具有双重作用.通过对2000~2020年西南地区土地利用转移矩阵的定量分析发现, 整体上, 土地利用类型发生变化的区域占研究区总面积的34.88%, 主要表现为耕地和草地被占用, 而林地、建设用地和未利用地面积增加的变化特征.近21 a西南地区土地利用类型转变的原因可能是自2000年以后, 我国高度重视生态环境问题, 启动了一系列重大生态保护和修复工程, 如退耕还林、退耕还草和植树造林等项目[20, 21, 25, 35].随着西南地区经济快速发展, 城市化进程加快, 城镇建设用地的面积不断扩张, 大量的农田和草地被占用, 在一定程度上会对西南地区生态系统格局产生影响[44, 45], 但由于生态保护措施和土地管理方式的改变带来的生态系统功能和生态环境质量的提升, 整体上西南地区的植被NPP呈上升趋势[30~32].
5 结论(1) 时间尺度上, 2000~2021年西南地区植被NPP及各陆地植被生态系统NPP均呈波动上升趋势, 其中, 农田生态系统NPP上升斜率高于草地和森林生态系统.气候变化和人类活动对三大生态系统NPP变化均表现为促进作用, 且对农田生态系统的促进作用最强.
(2) 空间尺度上, 西南地区植被NPP呈上升趋势的面积远高于呈下降趋势的面积, 呈显著上升和极显著上升的区域主要分布在广西南部、四川东部、重庆西部, 以及云南和贵州交界处.气候变化和人类活动对农田生态系统促进作用面积占比均最大, 森林生态系统面积占比均最小.
(3) 区域尺度上, 植被NPP与平均气温、降水和日照时数呈正相关的面积大于呈负相关的面积, 而与生物干湿度指数呈负相关的面积大于呈正相关的面积.植被生态系统尺度上, 西南地区植被NPP与平均温度呈正相关的面积占比最高, 而日照时数对农田生态系统NPP变化的抑制作用大于促进作用, 对森林和草地生态系统NPP变化的促进作用大于抑制作用.
| [1] | Yan W B, He Y L, Cai Y, et al. Relationship between extreme climate indices and spatiotemporal changes of vegetation on Yunnan Plateau from 1982 to 2019[J]. Global Ecology and Conservation, 2021, 31. DOI:10.1016/j.gecco.2021.e01813 |
| [2] | Song L Y, Li M Y, Xu H, et al. Spatiotemporal variation and driving factors of vegetation net primary productivity in a typical karst area in China from 2000 to 2010[J]. Ecological Indicators, 2021, 132. DOI:10.1016/j.ecolind.2021.108280 |
| [3] | Wu L Z, Ma X F, Dou X, et al. Impacts of climate change on vegetation phenology and net primary productivity in arid Central Asia[J]. Science of the Total Environment, 2021, 796. DOI:10.1016/j.scitotenv.2021.149055 |
| [4] |
徐勇, 黄雯婷, 郑志威, 等. 基于空间尺度效应的西南地区植被NPP影响因子探测[J]. 环境科学, 2023, 44(2): 900-911. Xu Y, Huang W T, Zheng Z W, et al. Detecting influencing factor of vegetation NPP in Southwest China based on spatial scale effect[J]. Environmental Science, 2023, 44(2): 900-911. |
| [5] | Tum M, Zeidler J N, Günther K P, et al. Global NPP and straw bioenergy trends for 2000-2014[J]. Biomass and Bioenergy, 2016, 90: 230-236. DOI:10.1016/j.biombioe.2016.03.040 |
| [6] |
汲玉河, 周广胜, 王树东, 等. 2000-2019年秦岭地区植被生态质量演变特征及驱动力分析[J]. 植物生态学报, 2021, 45(6): 617-625. Ji Y H, Zhou G S, Wang S D, et al. Evolution characteristics and itsdriving forces analysis of vegetation ecological quality in Qinling Mountains region from 2000 to 2019[J]. Chinese Journal of Plant Ecology, 2021, 45(6): 617-625. |
| [7] | Mao F J, Du H Q, Zhou G M, et al. Simulated net ecosystem productivity of subtropical forests and its response to climate change in Zhejiang Province, China[J]. Science of the Total Environment, 2022, 838. DOI:10.1016/j.scitotenv.2022.155993 |
| [8] | Qin Y W, Xiao X M, Wigneron J P, et al. Large loss and rapid recovery of vegetation cover and aboveground biomass over forest areas in Australia during 2019-2020[J]. Remote Sensing of Environment, 2022, 278. DOI:10.1016/j.rse.2022.113087 |
| [9] | Li W J, Xie S Y, Wang Y, et al. Effects of urban expansion on ecosystem health in Southwest China from a multi-perspective analysis[J]. Journal of Cleaner Production, 2021, 294. DOI:10.1016/j.jclepro.2021.126341 |
| [10] | Gang C C, Zhang Y Z, Wang Z Q, et al. Modeling the dynamics of distribution, extent, and NPP of global terrestrial ecosystems in response to future climate change[J]. Global and Planetary Change, 2017, 148: 153-165. DOI:10.1016/j.gloplacha.2016.12.007 |
| [11] | Beltrán-Sanz N, Raggio J, Gonzalez S, et al. Climate change leads to higher NPP at the end of the century in the Antarctic Tundra: response patterns through the lens of lichens[J]. Science of the Total Environment, 2022, 835. DOI:10.1016/j.scitotenv.2022.155495 |
| [12] | Yan Y C, Wu C J, Wen Y Y. Determining the impacts of climate change and urban expansion on net primary productivity using the spatio-temporal fusion of remote sensing data[J]. Ecological Indicators, 2021, 127. DOI:10.1016/j.ecolind.2021.107737 |
| [13] |
茆杨, 蒋勇军, 张彩云, 等. 近20年来西南地区植被净初级生产力时空变化与影响因素及其对生态工程响应[J]. 生态学报, 2022, 42(7): 2878-2890. Mao Y, Jiang Y J, Zhang C Y, et al. Spatio-temporal changes and influencing factors of vegetation net primary productivity in Southwest China in the past 20 years and its response to ecological engineering[J]. Acta Ecologica Sinica, 2022, 42(7): 2878-2890. |
| [14] | Ge W Y, Deng L Q, Wang F, et al. Quantifying the contributions of human activities and climate change to vegetation net primary productivity dynamics in China from 2001 to 2016[J]. Science of the Total Environment, 2021, 773. DOI:10.1016/j.scitotenv.2021.145648 |
| [15] | Zhang M, Yuan N Q, Lin H, et al. Quantitative estimation of the factors impacting spatiotemporal variation in NPP in the Dongting Lake wetlands using Landsat time series data for the last two decades[J]. Ecological Indicators, 2022, 135. DOI:10.1016/j.ecolind.2022.108544 |
| [16] |
张心茹, 曹茜, 季舒平, 等. 气候变化和人类活动对黄河三角洲植被动态变化的影响[J]. 环境科学学报, 2022, 42(1): 56-69. Zhang X R, Cao Q, Ji S P, et al. Quantifying the contributions of climate change and human activities to vegetation dynamic changes in the Yellow River Delta[J]. Acta Scientiae Circumstantiae, 2022, 42(1): 56-69. |
| [17] |
刘婧, 汤峰, 张贵军, 等. 2000—2015年滦河流域植被净初级生产力时空分布特征及其驱动因子分析[J]. 中国生态农业学报(中英文), 2021, 29(4): 659-671. Liu J, Tang F, Zhang G J, et al. Spatio-temporal distribution of net primary productivity and its driving factors in the Luanhe River Basin from 2000 to 2015[J]. Chinese Journal of Eco-Agriculture, 2021, 29(4): 659-671. |
| [18] | Peng Q, Wang R H, Jiang Y L, et al. Contributions of climate change and human activities to vegetation dynamics in Qilian Mountain National Park, Northwest China[J]. Global Ecology and Conservation, 2021, 32. DOI:10.1016/j.gecco.2021.e01947 |
| [19] | Wang Y J, Shen X J, Jiang M, et al. Daytime and nighttime temperatures exert different effects on vegetation net primary productivity of marshes in the western Songnen Plain[J]. Ecological Indicators, 2022, 137. DOI:10.1016/j.ecolind.2022.108789 |
| [20] |
徐勇, 黄雯婷, 窦世卿, 等. 2000~2020年西南地区植被NDVI对气候变化和人类活动响应特征[J]. 环境科学, 2022, 43(6): 3230-3240. Xu Y, Huang W T, Dou S Q, et al. Responding mechanism of vegetation cover to climate change and human activities in Southwest China from 2000 to 2020[J]. Environmental Science, 2022, 43(6): 3230-3240. |
| [21] |
徐勇, 戴强玉, 黄雯婷, 等. 2000~2020年西南地区植被NDVI时空变化及驱动机制探究[J]. 环境科学, 2023, 44(1): 323-335. Xu Y, Dai Q Y, Huang W T, et al. Spatio-temporal variation of vegetation cover and its driving mechanisms exploration in Southwest China from 2000 to 2020[J]. Environmental Science, 2023, 44(1): 323-335. |
| [22] | Kong D X, Miao C Y, Wu J W, et al. Time lag of vegetation growth on the Loess Plateau in response to climate factors: Estimation, distribution, and influence[J]. Science of the Total Environment, 2020, 744. DOI:10.1016/j.scitotenv.2020.140726 |
| [23] |
赵筱青, 石小倩, 李驭豪, 等. 滇东南喀斯特山区生态系统服务时空格局及功能分区[J]. 地理学报, 2022, 77(3): 736-756. Zhao X Q, Shi X Q, Li Y H, et al. Spatio-temporal pattern and functional zoning of ecosystem services in the karst mountainous areas of southeastern Yunnan[J]. Acta Geographica Sinica, 2022, 77(3): 736-756. |
| [24] | Letcher T M. Managing global warming: an interface of technology and human issues[M]. Massachusetts: Academic Press, 2018. |
| [25] | Zhang Y, Yuan J, You C M, et al. Contributions of National Key Forestry Ecology Projects to the forest vegetation carbon storage in China[J]. Forest Ecology and Management, 2020, 462. DOI:10.1016/j.foreco.2020.117981 |
| [26] |
李成, 杨舒慧, 吴芳, 等. "双碳"背景下中国能源消费碳排放与植被固碳的时空分异[J]. 中国环境科学, 2022, 42(4): 1945-1953. Li C, Yang S H, Wu F, et al. Spatio-temporal patterns of CO2 emissions from energy consumption and vegetation carbon sequestration in China under the emission peak and carbon neutrality[J]. China Environmental Science, 2022, 42(4): 1945-1953. |
| [27] |
谢春平, 吴显坤, 薛晓明, 等. 浙江楠适生区与气候环境关系的分析[J]. 四川农业大学学报, 2021, 38(3): 264-271. Xie C P, Wu X K, Xue X M, et al. Analysis on relationship between suitable habitat of Phoebe chekiangensis and climatic environmental factors[J]. Journal of Sichuan Agricultural University, 2021, 38(3): 264-271. |
| [28] | Yang H, Hu J, Zhang S, et al. Climate variations vs. human activities: distinguishing the relative roles on vegetation dynamics in the three Karst Provinces of Southwest China[J]. Frontiers in Earth Science, 2022, 10. DOI:10.3389/FEART.2022.799493 |
| [29] |
田智慧, 任祖光, 魏海涛. 2000~2020年黄河流域植被时空演化驱动机制[J]. 环境科学, 2022, 43(2): 743-751. Tian Z H, Ren Z G, Wei H T. Driving mechanism of the spatiotemporal evolution of vegetation in the Yellow River Basin from 2000 to 2020[J]. Environmental Science, 2022, 43(2): 743-751. |
| [30] |
洪辛茜, 黄勇, 孙涛. 我国西南喀斯特地区2001—2018年植被净初级生产力时空演变[J]. 生态学报, 2021, 41(24): 9836-9846. Hong X Q, Huang Y, Sun T. Spatiotemporal evolution of vegetation net primary productivity in the karst region of Southwest China from 2001 to 2018[J]. Acta Ecologica Sinica, 2021, 41(24): 9836-9846. |
| [31] | Wang Y H, Dai E F, Wu C S. Spatiotemporal heterogeneity of net primary productivity and response to climate change in the mountain regions of Southwest China[J]. Ecological Indicators, 2021, 132. DOI:10.1016/j.ecolind.2021.108273 |
| [32] |
马炳鑫, 靖娟利, 徐勇, 等. 2000—2019年滇黔桂岩溶区植被NPP时空变化及与气候变化的关系研究[J]. 生态环境学报, 2021, 30(12): 2285-2293. Ma B X, Jing J L, Xu Y, et al. Spatial-temporal changes of NPP and its relationship with climate change in karst areas of Yunnan, Guizhou and Guangxi from 2000 to 2019[J]. Ecology and Environmental Sciences, 2021, 30(12): 2285-2293. |
| [33] |
杨金虎, 张强, 王劲松, 等. 近60年来西南地区旱涝变化及极端和持续性特征认识[J]. 地理科学, 2015, 35(10): 1333-1340. Yang J H, Zhang Q, Wang J S, et al. Extreme and persistent feature of drought and flood of Southwest China in Past 60 years[J]. Scientia Geographica Sinica, 2015, 35(10): 1333-1340. |
| [34] |
赵志平, 吴晓莆, 李果, 等. 2009—2011年我国西南地区旱灾程度及其对植被净初级生产力的影响[J]. 生态学报, 2015, 35(2): 350-360. Zhao Z P, Wu X P, Li G, et al. Drought in southwestern China and its impact on the net primary productivity of vegetation from 2009—2011[J]. Acta Ecologica Sinica, 2015, 35(2): 350-360. |
| [35] |
孙治娟, 谢世友. 基于地理探测器的云南省净初级生产力时空演变及因子探测[J]. 生态学杂志, 2021, 40(12): 3836-3848. Sun Z J, Xie S Y. Spatiotemporal variation in net primary productivity and factor detection in Yunnan Province based on geodetector[J]. ChineseJournal of Ecology, 2021, 40(12): 3836-3848. |
| [36] |
王淑静, 赖佩玉, 郝斌飞, 等. 西南地区2001~2019年森林损失特征遥感监测与时空分析[J]. 遥感技术与应用, 2021, 36(3): 552-563. Wang S J, Lai P Y, Hao B F, et al. Remote sensing monitoring and spatio-temporal pattern of deforestation in Southwest China from 2001 to 2019[J]. Remote Sensing Technology and Application, 2021, 36(3): 552-563. |
| [37] |
耿庆玲, 陈晓青, 赫晓慧, 等. 中国不同植被类型归一化植被指数对气候变化和人类活动的响应[J]. 生态学报, 2022, 42(9): 3557-3568. Geng Q L, Chen X Q, He X H, et al. Vegetation dynamics and its response to climate change and human activities based on different vegetation types in China[J]. Acta Ecologica Sinica, 2022, 42(9): 3557-3568. |
| [38] | Khalifa M, Elagib N A, Ribbe L, et al. Spatio-temporal variations in climate, primary productivity and efficiency of water and carbon use of the land cover types in Sudan and Ethiopia[J]. Science of the Total Environment, 2018, 624: 790-806. |
| [39] |
王震, 闫文德, 刘曙光, 等. 基于MODIS数据的中国三种主要土地类型变化的空间特征分析[J]. 生态学报, 2017, 37(10): 3295-3301. Wang Z, Yan W D, Liu S G, et al. Spatial-temporal characteristics of three main land-use types in China based on MODIS data[J]. Acta Ecologica Sinica, 2017, 37(10): 3295-3301. |
| [40] | Wang J, Meng J J, Cai Y L. Assessing vegetation dynamics impacted by climate change in the southwestern karst region of China with AVHRR NDVI and AVHRR NPP time-series[J]. Environmental Geology, 2008, 54(6): 1185-1195. |
| [41] | Liu H Y, Zhang M Y, Lin Z S. Relative importance of climate changes at different time scales on net primary productivity-a case study of the Karst area of Northwest Guangxi, China[J]. Environmental Monitoring and Assessment, 2017, 189(11). DOI:10.1007/s10661-017-6251-5 |
| [42] | De Jong R, Verbesselt J, Zeileis A, et al. Shifts in global vegetation activity trends[J]. Remote Sensing, 2013, 5(3): 1117-1133. |
| [43] |
徐勇, 周清华, 窦世卿, 等. 基于ZGS和TW模型的长江流域植被NPP时空演变特征[J]. 水土保持通报, 2022, 42(1): 225-232. Xu Y, Zhou Q H, Dou S Q, et al. Spatiotemporal characteristics of vegetation net primary productivity in Yangtze River Basinbased on ZGS and Thornthwaite Memorial models[J]. Bulletin of Soil and Water Conservation, 2022, 42(1): 225-232. |
| [44] | Ding Z, Zheng H, Li H, et al. Afforestation-driven increases in terrestrial gross primary productivity are partly offset by urban expansion in Southwest China[J]. Ecological Indicators, 2021, 127. DOI:10.1016/j.ecolind.2021.107641 |
| [45] | Li W J, Wang Y, Xie S Y, et al. Spatiotemporal evolution scenarios and the coupling analysis of ecosystem health with land use change in Southwest China[J]. Ecological Engineering, 2022, 179. DOI:10.1016/j.ecoleng.2022.106607 |