 2023, Vol. 44
2023, Vol. 44



生物气溶胶是大气气溶胶的一部分, 是含生物性粒子的多相混合的气态分散系统, 包括细菌、真菌、病毒、花粉和虫卵等, 其生物性颗粒约占总颗粒物浓度的25%[1, 2].大气微生物对生态系统和生物健康具有重要影响, 可作为冰、云和雾的凝结核促进降水, 也附着在各种非生物性粒子上进行远距离传播[3]. 大气微生物体积小质量轻, 能通过皮肤、黏膜和呼吸道等进入生物体内[4, 5].有研究表明, 大气中真菌孢子浓度和哮喘患者数量存在显著相关性[6].吸入枝孢菌属(Cladosporium spp.)、链格孢菌属(Alternaria spp.)和镰刀菌属(Fusarium spp.)类真菌会造成呼吸道过敏, 曲霉属(Aspergillus spp.)和青霉属(Penicillium spp.)的繁殖体可能造成肺部感染[7].春夏季城市行道树产生的花粉气溶胶携带大量微生物[8], 在城市形成“飞雪”一样的景象, 是致人感染的主要过敏原[9], 空气中花粉飞絮含量与急诊入院病人呈正相关[10].大气中的细菌和真菌大多来源于土壤和植被[11, 12], 受各种自然和人为因素的影响[13, 14].城市中大气污染物类型多样, 与花粉飞絮相互作用会造成更严重的健康风险[15].
西安市碑林区行道和景观树以悬铃木(Platanus acerifolia)和梾木(Cornus macrophylla Wall.)为主, 在春季释放大量花粉形成飞絮.通过采集生物气溶胶、PM2.5和飞絮, 研究了西安市交通样点和校园样点生物气溶胶的浓度、微生物的来源和群落结构, 探讨其潜在的健康风险, 以期为飞絮和生物气溶胶进一步的健康影响研究提供基础数据支持.
1 材料与方法 1.1 样品采集2021年5月的花粉季节, 在西安市碑林区兴庆路(108°59′18″E, 34°14′32″N)和西安交通大学教学二区内设置采样点(108°59′22″E, 34°14′32″N), 分别表示交通样点和校园样点, 两点相距130 m.兴庆路样点南侧为友谊东路、雁翔路和南二环路, 车流量较大, 行道树多为悬铃木、并有少量木犀(Osmanthus sp.); 校园样点位于西安交通大学教学二区校园一树林内, 植被郁闭度较高, 乔灌草分别为梾木、八角金盘(Fatsia japonica)和沿阶草(Ophiopogon bodinieri Levl.).悬铃木飞絮呈绒毛状、比表面大、易随风传播, 梾木飞絮紧实, 不易飘浮.
在2021年5月9~11日每日的08:00、13:00和18:00分别采集微生物气溶胶和PM2.5样品, 于每日的14:00采集飞絮样品.使用Anderson六级微生物采样器(青岛聚创, 中国, 28.3 L·min-1)采集生物气溶胶至琼脂培养基和沙氏培养基上, 采样时间10 min, 采样高度1.5 m.样品通过恒温培养、计数, 分析可培养微生物气溶胶的浓度和粒径特征; 使用PQ2000大气颗粒物采样器(BGI, USA, 50 L·min-1)采集PM2.5至47 mm石英膜, 采样时间2 h, 采样高度1.5 m, 结束后将膜向内对折并用无菌铝箔包裹; 使用无菌镊子夹取掉落的飞絮约1 g至离心管中, 加入30 mL无菌水制得飞絮悬浊液.PM2.5和飞絮样品均保存在-20℃冰箱中, 经高通量测序分析大气颗粒物和飞絮的微生物特征.本研究共采集微生物气溶胶样品108个, 其中可培养细菌和可培养真菌样品各54个; PM2.5样品18个; 飞絮样品6个.
Anderson六级微生物采样器分6个级别, 分别表示人体呼吸系统的不同结构.1~6级的有效截留粒径及对应的人体结构分别为7.0 μm(鼻腔)、4.7 μm(咽喉)、3.3 μm(气管和主支气管)、2.1 μm(次支气管)、1.1 μm(末端支气管)和0.65 μm(肺泡)[16].配制琼脂培养基(牛肉膏3 g, 蛋白胨10 g, NaCl 5 g, 琼脂15 g, 超纯水1 000 mL, pH=7.4)和沙氏培养基(蛋白胨10 g, 琼脂20 g, 葡萄糖40 g, 超纯水1 000 mL, pH=6.2), 倒于直径9.0 cm的培养皿中备用.
每次采样前用75%酒精对采样器和采样工具进行消毒并充分风干, 使用灭菌锅(121℃, 25 min)将超纯水灭菌制得无菌水, 同时对离心管灭菌消毒, 使用马弗炉灼烧石英膜(780℃, 3 h)和铝箔(450℃, 3 h), 灭菌同时去除杂质.
1.2 样品分析已采样的琼脂培养基在37℃下培养48 h, 沙氏培养基在28℃下培养72 h, 定义总微生物是可培养细菌和真菌之和.培养结束后统计培养皿上的菌落数(colony forming units, CFU), 采用Positive-hole法进行校正, 公式如下:
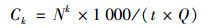
|
(1) |
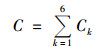
|
(2) |
式中, Ck和C分别为k级和微生物气溶胶总浓度, CFU·m-3; Nk为k级菌落数, CFU; t为采样时间, min; Q为采样器空气流量, L·min-1.
将培养结束的细菌和真菌样品分别进行革兰氏染色和棉兰染色, 在显微镜下观察其形态.将采集得到的PM2.5样品和飞絮样品委托上海派森诺生物科技有限公司进行DNA提取、扩增和高通量测序.
1.3 数据处理使用SPSS软件(R26.0.0.0, 32位)进行独立样本t检验、Spearman相关性分析, 以P<0.05作为显著性阈值.使用Origin(2018, 64bit)和Genescloud平台(https://www.genescloud.cn)进行图表制作.
2 结果与讨论 2.1 可培养生物气溶胶浓度交通样点可培养细菌浓度为校园样点的4.1倍(P=0.027), 均值分别为(1 472.3±1 351.8)CFU·m-3和(363.3±175.1)CFU·m-3; 可培养真菌浓度在两样点没有显著性差异(P=0.868), 分别为(535.9±167.5)CFU·m-3和(519.4±239.9)CFU·m-3.交通样点可培养细菌浓度均值约为真菌的2.7倍, 而校园样点真菌浓度均值约为细菌1.4倍.交通样点总微生物浓度(6 204.7 CFU·m-3)是校园样点(2 647.8 CFU·m-3)的2.3倍左右, 可培养细菌浓度占比分别为73.3%和41.1%, 细菌在交通样点的占比更高.可培养微生物浓度随地点和环境的不同而具有差异.
两采样点可培养生物气溶胶浓度日内变化呈谷型分布特征(图 1), 08:00和18:00较高而13:00低.交通样点和校园样点可培养细菌浓度峰谷值之比分别是4.1和1.8; 可培养真菌浓度峰谷值之比为1.8和1.6, 可培养细菌气溶胶变化幅度较大.校园样点可培养细菌浓度在13:00急剧减小, 可能由于午间较强的太阳辐射造成部分细菌死亡, Singer发现太阳辐射中的紫外线对细菌的生长起到抑制作用[17].Ho等[18]在中国台湾花莲的研究发现真菌气溶胶浓度在清晨(04:00~06:00)最高, 下午(14:00~16:00)最低, 随后逐渐升高.部分原因是空气的垂直混合深度和夜间的辐射逆温, 有学者称为“夜间模式”[19].清晨湿度较高, 一些孢子(Ascospores和Basidiospores)会加剧释放导致真菌浓度增加[20].在交通样点可培养细菌和真菌浓度均大于校园样点, 同样发现北京、南京、北海和西安等地大气中可培养细菌浓度高于真菌, 细菌与真菌浓度比值为1.1~4.1倍[21~23].校园样点真菌占总微生物浓度的58.9%, 可能是植物为真菌微生物的主要来源[14, 24, 25].

|
图 1 可培养微生物气溶胶浓度日内变化 Fig. 1 Diurnal variation in culturable bioaerosol concentration |
可培养细菌和真菌浓度在交通样点的变化幅度较校园样点大.交通样点人车流动频繁, 颗粒物含量显著增加.有研究表明在霾天微生物浓度会显著增加[26, 27], 空气中的细菌可与颗粒物结合形成团聚体以抵抗不利环境[28], 且颗粒物携带的水分易促进微生物的生长繁殖, 对生物气溶胶浓度的增加具有积极的作用[29, 30].但交通样点真菌浓度变化幅度较细菌小, 可能是真菌的孢子结构更能抵抗不利环境的影响.两个样点比较发现人类活动、颗粒物浓度等因素会对生物气溶胶浓度变化造成较大影响[31].
图 2(a)和2(b)为交通样点和校园样点的周边环境, 大气微生物在沙氏培养基上生长得到表面覆盖孢子、颜色较深的真菌菌落[图 2(c)], 在琼脂培养基上生长得到表面湿润光滑的细菌菌落[图 2(d)].挑取部分真菌经棉兰染色后在显微镜下可见菌丝及孢子形态[图 2(e)], 细菌经革兰氏染色后呈紫色球状[图 2(f)].

|
图 2 采样点环境、培养的真菌和细菌及染色效果 Fig. 2 Sampling site, cultured, and stained fungi and bacteria |
可培养细菌呈双峰分布, 峰值对应粒径为>7.0 μm(第一峰)和2.1~3.3 μm(第二峰), 可培养真菌为单峰分布, 峰值对应粒径为2.1~3.3 μm, 可培养细菌第二峰与真菌峰值粒径相同(图 3).细菌气溶胶粒径分布与一项在西安秋冬季可培养细菌微生物的研究结果相似[32].有研究表明青岛室外的可培养真菌微生物在所有季节均为单峰分布且峰值出现在2.1~3.3 μm[33], 与本实验的结果一致.细菌和真菌在大气中的存在形式不同, 细菌主要附着在花粉和粉尘等颗粒物表面以团聚体存在, 真菌以细小的孢子形式存在[34].可培养微生物气溶胶浓度在各粒径分布存在日内差异(图 4).可培养细菌气溶胶第二峰对应粒径在3.3~4.7 μm和2.1~3.3 μm之间变动, 在08:00和13:00浓度峰值对应粒径为3.3~4.7 μm, 而18:00则对应2.1~3.3 μm, 从早至晚粒径向细小颗粒转移.可能是由于中午强烈的阳光导致大粒子细菌的活性降低、死亡[35].对于可培养真菌微生物, 在交通样点08:00和13:00的峰值浓度集中在2.1~3.3 μm, 18:00时峰值位于1.1~2.1 μm, 而校园样点处峰值浓度对应的粒径均倾向于集中在2.1~3.3 μm, 这可能是由于交通样点人为活动频繁、植被稀少和环境更复杂导致的.

|
图 3 微生物气溶胶粒径分布 Fig. 3 Size distribution of culturable bacterial and fungi |

|
图 4 不同样点微生物气溶胶的粒径分布 Fig. 4 Diurnal size distribution of culturable bacterial and fungi at different sampling sites |
当微生物粒径<4.7 μm时易通过人体的咽喉到达气管和肺泡等组织[19, 36].可培养细菌和真菌粒径在<4.7 μm的尺度上占比为60.00%和81.25%, 与细菌相比, 真菌气溶胶有更大的吸入风险.细颗粒物对人体有重要的健康影响, 在发展中国家急性下呼吸道感染是5岁以下幼儿死亡的主要原因之一, 在2017年因此有180万人死亡[37].有研究发现因哮喘而急诊的儿童与空气中的PM2.5之间存在显著相关的非线性的关系, 当ρ(PM2.5)达到20 μg·m-3时因哮喘而就诊的儿童数量急剧增加[38].抗生素的使用很大程度上避免了细菌对人体的进一步侵害, 但对真菌的感染引发的疾病关注较少[39].由于大气中真菌主要集中在细颗粒物上更易侵入人体, 因此后续研究需要关注真菌感染引发的健康风险.
2.3 细菌微生物群落结构组成和来源样品经高通量测序分析, 稀疏曲线趋于平稳(图 5), 测序深度可反映不同样点的物种丰富度.飞絮样品α多样性见表 1, 两样点样品丰度、覆盖度和多样性指数均有显著差异(P=0.05).交通样点飞絮样品OTUs、丰度和多样性指数均高于校园样点.交通样点飞絮主要来自悬铃木, 而校园样点飞絮主要来源为梾木, 悬铃木贡献较低, 飞絮样品覆盖度随采集类型的增加而增加.同样抽平深度下交通样点飞絮样品多样性指数更高, 说明绒毛状构造更容易负载微生物且负载的种类更丰富.PM2.5样品在每日的08:00、13:00和18:00多样性指数依次降低, 空气中微生物大多来源于周边环境且在一定时期内能够保持稳定[40], 故造成其变化的原因可能是春季清晨有利于多种微生物生存的潮湿环境和不利于其生存的下午强烈光照及较高浓度的臭氧.在本研究中可培养微生物浓度在清晨最高, 已有相关研究证明湿度有利于微生物的生长繁殖[20, 29, 30].

|
图 5 稀疏曲线 Fig. 5 Sparse curve |
|
|
表 1 飞絮样品细菌α多样性 Table 1 The α diversity of bacteria in flying catkins |
门水平上(图 6), 变形菌门(Proteobacteria)在西安春季分布最广, 不同样点和不同类型样品优势菌门不同.PM2.5样品在交通和校园样点共有的优势门类(丰度>1%)计6种, 分别为变形菌门、放线菌门(Actinobacteria)、厚壁菌门(Firmicutes)、拟杆菌门(Bacteroidetes)、蓝藻门(Cyanobacteria)和异常球菌-栖热菌门(Deinococcus-Thermus).在交通样点上述优势菌门占比分别为39.82%、23.96%、16.98%、6.25%、4.76%和2.43%, 在校园样点为45.36%、15.72%、15.59%、6.49%、9.03%和2.76%.飞絮样品在交通和校园样点共有的优势门类仅为变形菌门, 占比分别为91.27%和99.06%.变形菌门是PM2.5和飞絮样品中最丰富的门类菌, 北京、浙江和合肥的研究中有同样发现[8, 41, 42].基于物种丰度矩阵进行主成分分析(principal components analysis, PCA, 图 7), PC1和PC2的解释度分别为51%和34.9%, 总计85.9%.PC1对交通样点和校园样点(09)飞絮的解释度较大, PC2对PM2.5样品(除校园样点13:00样品)和校园样点飞絮(10和11)的解释度高于PC1.因此PM2.5和校园样点飞絮样品受PC2的贡献更大, 而交通样点飞絮受PC1的贡献更高.微生物来自土壤、植被、海洋、动物粪便和废物设施等, 其中最主要的来源为陆地环境, 如土壤和植被可通过雾化将细菌扩散到大气中, 源的类型直接影响微生物的群落结构组成[31, 43].变形菌门、放线菌门、厚壁菌门和拟杆菌门是典型的土壤源菌, 也是多个城市的优势菌门[40, 42, 44~46].同时变形菌门是多种飞絮微生物的优势菌[47].可以确定, PM2.5和飞絮样品微生物的主要来源为土壤和植被, 受PC1主导时, 飞絮中变形菌门丰度>90%.综上, PC1是植被源, PC2为土壤源, 即飞絮样品的微生物主要来自植被源, PM2.5中的微生物主要来自土壤源.在主要受土壤源影响时微生物的优势门类较多, 但对于受到土壤源更多贡献的校园样点飞絮(10和11)多样性较交通样点低, 这可能是飞絮类型导致的.本研究采集由植物掉落地面的飞絮, 其不仅有植物源微生物, 也含地表微生物.呈绒毛状、比表面更大的悬铃木飞絮更易负载物质, 蓝藻门非典型城市土壤源菌, 在一些城市的研究中并非优势菌属[1, 40, 46], 但在两样点PM2.5样品中占比较高.蓝藻门是初级生产者的关键组成部分, 在旱地土壤结皮和沙漠广泛存在, 也是水生生态系统的基本组成部分, 与矿物有很深的亲和力, 与赤潮等现象密切相关[48~50].但在杭州西湖的研究发现蓝藻门并非优势菌门[46], 水体不是大气微生物的主要来源.Xie等[51]在西安的研究发现细菌可跨地域传播.在北京的研究发现, 蓝藻在微生物种群中占比最高, 它来源于俄罗斯和蒙古沙漠地区的矿物盐, 通过风进行迁移[50].本研究PM2.5样品微生物群落中的蓝藻门可能是大陆性季风带来的北部沙漠矿物盐产生.

|
F1-1、F1-2、F1-3、F09-1、F10-1和F11-1表示交通样点; F2-1、F2-2、F2-3、F09-2、F10-2和F11-2表示校园样点 图 6 PM2.5和飞絮样品的微生物群落结构 Fig. 6 Bacterial community structure of PM2.5 and fluffy samples |

|
JT表示交通样点的PM2.5, XY表示校园样点的PM2.5, 09、10和11分别表示2021-05-09、2021-05-10和2021-05-11的样品 图 7 主成分分析 Fig. 7 Principal component analysis |
属水平上, PM2.5比飞絮样品菌种丰度更高.PM2.5样品中两个样点丰度均>1%的菌种有:Chloroplast、乳杆菌属(Lactobacillus)、盐单胞菌属(Halomonas)、副球菌属(Paracoccus)、假单胞菌属(Pseudomonas)、Nesterenkonia、鞘氨醇单胞菌属(Sphingomonas)和Rubellimicrobium.另有痤疮丙酸杆菌属(Cutibacterium)和不动杆菌属(Acinetobacter)在交通样点丰度>1%;泛菌属(Pantoea)、明串珠菌(Leuconostoc)、WPS-2、异常球菌(Deinococcus)和链球菌属(Streptococcus)在校园样点丰度>1%.泛菌属广泛存在于植物中, 痤疮丙酸杆菌属、不动杆菌属、明串珠菌、异常球菌和链球菌属均为典型的土壤源菌.在北京, 平球菌属(Planococcus)、库克菌属(Kocuria)和芽孢杆菌属(Bacillus)是春季大气的优势菌[52]; 在兰州, 以乳球菌属和芽孢杆菌属为代表[45]; 在西安春季最丰富的菌属是Prevotella_9[53].导致生物气溶胶来源变化的地理变化强烈影响微生物的丰度. WPS-2从未被培养过, 也没基因组数据, 近来才逐渐被认识.它存在于泥炭地、沼泽地以及南极裸土、北极冰锥和受金属污染等有机质贫瘠的土壤和干燥的裸露土壤中[54, 55]. WPS-2可以适应极端环境, 具有提供初级生产力的功能[55], 校园样点植被类型单一、群落结构简单和空间异质性低, 这可能是WPS-2细菌存在的原因.在交通样点, WPS-2丰度不足1%, 可能采样点周边并无WPS-2菌生存的条件.
交通样点飞絮样品丰度>1%的有: 假单胞菌属(55.96%)、Stenotrophomonas(4.66%)和泛菌属(1.67%); 校园样点中丰度>1%的有: 泛菌属(38.71%)、假单胞菌属(10.85%)和肠杆菌属(Enterobacter, 4.88%).交通样点的优势属为假单胞菌属, 丰度为校园样点的5倍, 校园样点丰度最高的泛菌属是交通样点的23倍, 可知不同飞絮类型对负载的微生物的选择倾向不同.Xu等[8]在北京不同地点采集杨树(Populus)飞絮, 发现副球菌属、Rubellimicrobium、库克菌属、鞘氨醇单胞菌属和Serratia是主要的优势菌, 但不同样点优势菌属不同.综上飞絮类型和微生物来源环境均会导致飞絮的优势菌属发生变化.
2.4 飞絮传播对人类健康的潜在影响悬铃木作为优秀的行道树, 在中国从南到北均有栽培, 梾木在城市园林中多有使用.这两种树在中国分布广泛, 每年花季产生大量飞絮, 尤其是悬铃木, 飞絮蓬松细小且容易附着在人体黏膜和鼻腔等部位, 部分粒子甚至可达人体肺部.人们普遍认为呼吸道微生物种群的组成是由气道生理环境决定的, 当暴露在巨变的大气环境中时将导致人体呼吸道的微生物种群改变, 从而引起炎症、激活人体特异性免疫调节和导致气道生理特征改变进一步引发哮喘、鼻炎、肺炎和结膜炎等疾病[56].飞絮可携带大量的生物性颗粒, 部分细菌、真菌、植物属和病毒具有致病致敏的危害.如细菌:假单胞菌属; 真菌:念珠菌; 植物:松树(Pinus); 病毒:疱疹病毒(Herpesviridae)[8].本研究发现的肠杆菌属和假单胞菌属等是潜在的人体致病菌.肠杆菌属是革兰氏阳性菌, 病原体可引起腹泻、软组织、尿路和呼吸道感染等[57].假单胞菌属在交通样点飞絮样品中丰度达56%, 其病原体可导致休克, 中耳炎、脑膜炎、败血症、呼吸道和尿路感染[58]等.在开花季节人体接触大量飞絮, 除其携带的微生物会对人体产生健康影响外, 飞絮本身也是引起人体过敏的重要因素.Cebrino等[59]在西班牙的研究中发现草花粉、橄榄花粉、梧桐花粉和荨麻科花粉会引起人群感染, 所有花粉与鼻炎和结膜炎的发生呈正比.Ribeiro等[10]的研究发现每年季节性住院高峰人数与当地槭树(Acer)、悬铃木、杨树和栎树(Quercus)的每月花粉浓度呈显著正相关.北京杨柳造成的“四月雪”经常引起人群感染, 在蓬松的柳絮上经常发现疱疹病毒、疣状病毒(Potyviridae)和黄病毒(Flaviviridae)等, 它们可感染动物和人类, 并在受感染的宿主体内持续存在[59].以上结果表明, 西安市蓬松的悬铃木飞絮和梾木飞絮可诱发腹泻、呼吸道感染、气管炎、肺炎和鼻炎等疾病, 影响人体健康.
3 结论(1) 交通样点及校园样点可培养微生物浓度变化差异明显, 交通样点总微生物浓度是校园样点的2.3倍, 它们的日内峰值浓度呈现谷型变化特征, 可培养细菌在日内变化幅度较真菌更大.
(2) 可培养细菌和真菌微生物粒径分布存在明显差异, 可培养细菌粒径变化呈双峰分布, 第二峰粒径在日内从早至晚存在向细粒子转移的现象.可培养真菌粒径变化为单峰分布, 且具有更大的吸入风险.
(3) 变形菌门是飞絮和PM2.5样品中共有和丰度最高的门水平菌, 植被和土壤是大气微生物的主要来源, 植被源对飞絮的贡献更大, 土壤源细菌多样性更高, 表现为PM2.5比飞絮含有更多丰度>1%的细菌, 导致生物气溶胶来源变化的地理变化强烈影响微生物的丰度, 且不同飞絮携带的微生物类型有差异.春季飘扬的飞絮及生物性颗粒在人口密集的城市更易造成人群感染, 因此更需加强防护.
| [1] |
李天佑, 祁建华, 马曼曼, 等. 不同空气质量下生物气溶胶中细菌群落特征研究[J]. 环境科学与技术, 2021, 44(1): 58-68. Li T Y, Qi J H, Ma M M, et al. Characteristics of bacterial community in atmospheric bioaerosols under different air quality[J]. Environmental Science & Technology, 2021, 44(1): 58-68. DOI:10.19672/j.cnki.1003-6504.2021.01.008 |
| [2] |
李彦鹏, 马天峰, 杜胜利, 等. 大气生物气溶胶的源排放与源解析研究进展[J]. 地球科学与环境学报, 2021, 43(2): 315-331. Li Y P, Ma T F, Du S L, et al. Review on source emission and source identification of bioaerosols in the atmosphere[J]. Journal of Earth Sciences and Environment, 2021, 43(2): 315-331. DOI:10.19814/j.jese.2020.11018 |
| [3] | Ariya P A, Sun J, Eltouny N A, et al. Physical and chemical characterization of bioaerosols-Implications for nucleation processes[J]. International Reviews in Physical Chemistry, 2009, 28(1): 1-32. DOI:10.1080/01442350802597438 |
| [4] |
宫静, 祁建华, 李鸿涛. 青岛近海生物气溶胶中总微生物的分布特征[J]. 环境科学, 2019, 40(8): 3477-3488. Gong J, Qi J H, Li H T. Distribution of total microbes in atmospheric bioaerosols in the coastal region of Qingdao[J]. Environmental Science, 2019, 40(8): 3477-3488. DOI:10.13227/j.hjkx.201901262 |
| [5] |
谢雯文, 路瑞, 慕飞飞, 等. 西安市秋冬季市区与山区微生物气溶胶组成特征及来源[J]. 环境科学, 2020, 41(5): 2044-2049. Xie W W, Lu R, Mu F F, et al. Characteristics and sources of microbial aerosols in urban and mountainous areas in autumn and winter in Xi'an, China[J]. Environmental Science, 2020, 41(5): 2044-2049. DOI:10.13227/j.hjkx.201911010 |
| [6] | Nasser N I, Al-Hadrawi M K, Oleiwi S A, et al. The diversity in dust fungal spores concentration at four districts of Al-Najaf environment and their potential correlation with Asthma[J]. Journal of Pure and Applied Microbiology, 2019, 13(4): 2169-2176. DOI:10.22207/JPAM.13.4.29 |
| [7] | Vacher G, Niculita-Hirzel H, Roger T. Immune responses to airborne fungi and non-invasive airway diseases[J]. Seminars in Immunopathology, 2015, 37(2): 83-96. DOI:10.1007/s00281-014-0471-3 |
| [8] | Xu S Y, Yao M S. Plant flowers transmit various bio-agents through air[J]. Science China Earth Sciences, 2020, 63(10): 1613-1621. DOI:10.1007/s11430-020-9629-2 |
| [9] | Cariñanos P, Casares-Porcel M. Urban green zones and related pollen allergy: a review. Some guidelines for designing spaces with low allergy impac[J]. Landscape Urban Plan, 2011, 10(3): 205-214. |
| [10] | Ribeiro H, Oliveira M, Ribeiro N, et al. Pollen allergenic potential nature of some trees species: a multidisciplinary approach using aerobiological, immunochemical and hospital admissions data[J]. Environmental Research, 2009, 109(3): 328-333. DOI:10.1016/j.envres.2008.11.008 |
| [11] | Barberán A, Ladau J, Leff J W, et al. Continental-scale distributions of dust-associated bacteria and fungi[J]. Proceedings of the National Academy of Sciences of the United States of America, 2015, 112(18): 5756-5761. DOI:10.1073/pnas.1420815112 |
| [12] | Gandolfi I, Bertolini V, Bestetti G, et al. Spatio-temporal variability of airborne bacterial communities and their correlation with particulate matter chemical composition across two urban areas[J]. Applied Microbiology and Biotechnology, 2015, 99(11): 4867-4877. DOI:10.1007/s00253-014-6348-5 |
| [13] |
闫威卓, 王步英, Montana O F, 等. 北偏西大风对北京冬季生物气溶胶的影响[J]. 环境科学, 2017, 38(9): 3561-3568. Yan W Z, Wang B Y, Montana O F, et al. Impact of gusty northwesterly winds on biological particles in winter in Beijing[J]. Environmental Science, 2017, 38(9): 3561-3568. DOI:10.13227/j.hjkx.201701110 |
| [14] |
方治国, 欧阳志云. 城市室内外空气真菌群落及影响因素研究进展[J]. 生态环境学报, 2009, 18(1): 386-393. Fang Z G, Ouyang Z Y. Advance of airborne fungal community and the influencing factors in indoor and outdoor environments in urban ecosystem[J]. Ecology and Environmental Sciences, 2009, 18(1): 386-393. DOI:10.3969/j.issn.1674-5906.2009.01.071 |
| [15] | Bosch-Cano F, Bernard N, Sudre B, et al. Human exposure to allergenic pollens: a comparison between urban and rural areas[J]. Environmental Research, 2011, 111(5): 619-625. DOI:10.1016/j.envres.2011.04.001 |
| [16] | Andersen A A. New sampler for the collection, sizing, and enumeration of viable airborne particles[J]. Journal of Bacteriology, 1958, 76(5): 471-484. DOI:10.1128/jb.76.5.471-484.1958 |
| [17] | Singer C E, Ames B N. Sunlight ultraviolet and bacterial DNA base ratios: natural exposure to ultraviolet may be an evolutionary pressure toward high guanine plus cytosine in DNA[J]. Science, 1970, 170(3960): 822-826. DOI:10.1126/science.170.3960.822 |
| [18] | Ho H M, Rao C Y, Hsu H H, et al. Characteristics and determinants of ambient fungal spores in Hualien, Taiwan[J]. Atmospheric Environment, 2005, 39(32): 5839-5850. DOI:10.1016/j.atmosenv.2005.06.034 |
| [19] | Eisenberg W V. The microbiology of the atmosphere[J]. Journal of Association of Official Agricultural Chemists, 1963, 46(2): 347-348. |
| [20] | Burch M, Levetin E. Effects of meteorological conditions on spore plumes[J]. International Journal of Biometeorology, 2002, 46(3): 107-117. DOI:10.1007/s00484-002-0127-1 |
| [21] | 陈梅玲, 胡庆轩, 徐秀芝, 等. 南京市大气微生物污染情况调查[J]. 中国公共卫生, 2000, 16(6): 504-505. DOI:10.3321/j.issn:1001-0580.2000.06.015 |
| [22] |
陈皓文. 北海市空气微生物含量时空分布[J]. 广西科学, 1998, 5(2): 83-86. Chen H W. Spatial and temporal distributions of air borne microbes above Beihai city[J]. Guangxi Sciences, 1998, 5(2): 83-86. |
| [23] |
胡庆轩, 车凤翔, 张松乐, 等. 京、津地区大气微生物的浓度[J]. 环境科学, 1989, 10(5): 30-35. Hu Q X, Che F X, Zhang S L, et al. A study on concentrations of microorgan-isms in the atmosphere of Beijing and Tianjin areas[J]. Environmental Science, 1989, 10(5): 30-35. |
| [24] | Awad A H A. Vegetation: a source of air fungal bio-contaminant[J]. Aerobiologia, 2005, 21(1): 53-61. DOI:10.1007/s10453-004-5878-1 |
| [25] | Fang Z G, Ouyang Z Y, Hu L F, et al. Culturable airborne fungi in outdoor environments in Beijing, China[J]. Science of the Total Environment, 2005, 350(1-3): 47-58. DOI:10.1016/j.scitotenv.2005.01.032 |
| [26] | Dong L J, Qi J H, Shao C C, et al. Concentration and size distribution of total airborne microbes in hazy and foggy weather[J]. Science of the Total Environment, 2016, 541: 1011-1018. DOI:10.1016/j.scitotenv.2015.10.001 |
| [27] |
路瑞, 李婉欣, 宋颖, 等. 西安市不同天气下可培养微生物气溶胶浓度变化特征[J]. 环境科学研究, 2017, 30(7): 1012-1019. Lu R, Li W X, Song Y, et al. Characteristics of culturable bioaerosols in various weather in Xi'an City, China[J]. Research of Environmental Sciences, 2017, 30(7): 1012-1019. |
| [28] | Kobayashi F, Maki T, Kakikawa M, et al. Bioprocess of Kosa bioaerosols: effect of ultraviolet radiation on airborne bacteria within Kosa (Asian dust)[J]. Journal of Bioscience and Bioengineering, 2015, 119(5): 570-579. DOI:10.1016/j.jbiosc.2014.10.015 |
| [29] | Li W L, Yang J S, Zhang D Z, et al. Concentration and community of airborne bacteria in response to cyclical haze events during the fall and midwinter in Beijing, China[J]. Frontiers in Microbiology, 2018, 9. DOI:10.3389/fmicb.2018.01741 |
| [30] | Yan D, Zhang T, Su J, et al. Structural variation in the bacterial community associated with airborne particulate matter in Beijing, China, during hazy and nonhazy days[J]. Applied and Environmental Microbiology, 2018, 84(9). DOI:10.1128/AEM.00004-18 |
| [31] | Ruiz-Gil T, Acuña J J, Fujiyoshi S, et al. Airborne bacterial communities of outdoor environments and their associated influencing factors[J]. Environment International, 2020, 145. DOI:10.1016/j.envint.2020.106156 |
| [32] | Yang L, Shen Z X, Wang D W, et al. Diurnal variations of size-resolved bioaerosols during autumn and winter over a semi-arid megacity in northwest China[J]. GeoHealth, 2021, 5(5). DOI:10.1029/2021GH000411 |
| [33] | Li M F, Qi J H, Zhang H D, et al. Concentration and size distribution of bioaerosols in an outdoor environment in the Qingdao coastal region[J]. Science of the Total Environment, 2011, 409(19): 3812-3819. DOI:10.1016/j.scitotenv.2011.06.001 |
| [34] |
方治国, 欧阳志云, 胡利锋, 等. 北京市三个功能区空气微生物中值直径及粒径分布特征[J]. 生态学报, 2005, 25(12): 3220-3224. Fang Z G, Ouyang Z Y, Hu L F, et al. Study on median diameters and size distributions of airborne microbes in three functional regions in Beijing[J]. Acta Ecologica Sinica, 2005, 25(12): 3220-3224. |
| [35] | Lighthart B, Shaffer B T, Marthi B, et al. Trajectory of aerosol droplets from a sprayed bacterial suspension[J]. Applied and Environmental Microbiology, 1991, 57(4): 1006-1012. |
| [36] | Pastuszka J S, Paw U K T, Lis D O, et al. Bacterial and fungal aerosol in indoor environment in Upper Silesia, Poland[J]. Atmospheric Environment, 2000, 34(22): 3833-3842. |
| [37] | Shi T, McLean K, Campbell H, et al. Aetiological role of common respiratory viruses in acute lower respiratory infections in children under five years: a systematic review and meta-analysis[J]. Journal of Global Health, 2015, 5(1). DOI:10.7189/jogh.05.010408 |
| [38] | Bouazza N, Foissac F, Urien S, et al. Fine particulate pollution and asthma exacerbations[J]. Archives of Disease in Childhood, 2018, 103(9): 828-831. |
| [39] | Roe K. How major fungal infections can initiate severe autoimmune diseases[J]. Microbial Pathogenesis, 2021, 161. DOI:10.1016/j.micpath.2021.105200 |
| [40] | Chen H L, Du R, Zhang Y T, et al. Evolution of PM2.5 bacterial community structure in Beijing's suburban atmosphere[J]. Science of the Total Environment, 2021, 799. DOI:10.1016/j.scitotenv.2021.149387 |
| [41] | Liu H, Zhang X, Zhang H, et al. Effect of air pollution on the total bacteria and pathogenic bacteria in different sizes of particulate matter[J]. Environmental Pollution, 2018, 233: 483-493. |
| [42] |
姜少毅, 孙博文, 代海涛, 等. 合肥市夏季大气颗粒物中微生物群落的高通量测序分析[J]. 环境科学, 2021, 42(5): 2169-2178. Jiang S Y, Sun B W, Dai H T, et al. High-throughput sequencing analysis of microbial communities in summertime atmospheric particulate matter in Hefei city[J]. Environmental Science, 2021, 42(5): 2169-2178. |
| [43] | Wei M, Xu C H, Xu X M, et al. Size distribution of bioaerosols from biomass burning emissions: characteristics of bacterial and fungal communities in submicron (PM1.0) and fine (PM2.5) particles[J]. Ecotoxicology and Environmental Safety, 2019, 171: 37-46. |
| [44] | Xu C H, Wei M, Chen J M, et al. Bacterial characterization in ambient submicron particles during severe haze episodes at Ji'nan, China[J]. Science of the Total Environment, 2017, 580: 188-196. |
| [45] |
赵炜, 李杰, 谢慧娜, 等. 兰州市春季微生物气溶胶浓度、粒径及细菌群落结构分布特性[J]. 环境科学, 2021, 42(4): 1668-1678. Zhao W, Li J, Xie H N, et al. Concentration and particle size distribution characteristics of microbial aerosol and bacterial community structure during spring in Lanzhou city, China[J]. Environmental Science, 2021, 42(4): 1668-1678. |
| [46] |
楼秀芹, 郭卫俊, 姚文冲, 等. 杭州市西湖景区春季空气细菌多样性特征[J]. 生态学报, 2019, 39(8): 2946-2953. Lou X Q, Guo W J, Yao W C, et al. Diversity of airborne bacterial community in spring in the West Lake scenic areas of Hangzhou, China[J]. Acta Ecologica Sinica, 2019, 39(8): 2946-2953. |
| [47] | Manirajan B A, Maisinger C, Ratering S, et al. Diversity, specificity, co-occurrence and hub taxa of the bacterial-fungal pollen microbiome[J]. FEMS Microbiology Ecology, 2018, 94(8). DOI:10.1093/femsec/fiy112 |
| [48] | Lu T, Zhang Q, Zhang Z Y, et al. Pollutant toxicology with respect to microalgae and cyanobacteria[J]. Journal of Environmental Sciences, 2021, 99: 175-186. |
| [49] | Muñoz-Martín M A, Becerra-Absalón I, Perona E, et al. Cyanobacterial biocrust diversity in Mediterranean ecosystems along a latitudinal and climatic gradient[J]. New Phytologist, 2019, 221(1): 123-141. |
| [50] | Park E H, Heo J, Kim H, et al. The major chemical constituents of PM2.5 and airborne bacterial community phyla in Beijing, Seoul, and Nagasaki[J]. Chemosphere, 2020, 254. DOI:10.1016/j.chemosphere.2020.126870 |
| [51] | Xie Z S, Du S L, Ma T F, et al. High time-resolved characterization of airborne microbial community during a typical haze pollution process[J]. Journal of Hazardous Materials, 2021, 415. DOI:10.1016/j.jhazmat.2021.125722 |
| [52] | Du P R, Du R, Ren W S, et al. Seasonal variation characteristic of inhalable microbial communities in PM2.5 in Beijing city, China[J]. Science of the Total Environment, 2018, 610-611: 308-315. |
| [53] | Wang S, Liu W Y, Li J, et al. Seasonal variation characteristics of bacteria and fungi in PM2.5 in typical basin cities of Xi'an and Linfen, China[J]. Atmosphere, 2021, 12(7). DOI:10.3390/atmos12070809 |
| [54] | Holland-Moritz H, Stuart J, Lewis L R, et al. Novel bacterial lineages associated with boreal moss species[J]. Environmental Microbiology, 2018, 20(7): 2625-2638. |
| [55] | Sheremet A, Jones G M, Jarett J, et al. Ecological and genomic analyses of candidate phylum WPS-2 bacteria in an unvegetated soil[J]. Environmental Microbiology, 2020, 22(8): 3143-3157. |
| [56] | Xue Y F, Chu J, Li Y W, et al. The influence of air pollution on respiratory microbiome: a link to respiratory disease[J]. Toxicology Letters, 2020, 334: 14-20. |
| [57] | 国家病原微生物资源库. 国家病原微生物资源库[EB/OL]. https://www.nprc.org.cn/, 2021-11-16. |
| [58] | Cillóniz C, Gabarrús A, Ferrer M, et al. Community-acquired pneumonia due to multidrug- and non-multidrug-resistant pseudomonas aeruginosa[J]. Chest, 2016, 150(2): 415-425. |
| [59] | Cebrino J, de la Cruz S P, Barasona M J, et al. Airborne pollen in Córdoba city (Spain) and its implications for pollen allergy[J]. Aerobiologia, 2017, 33(2): 281-291. |