 2022, Vol. 43
2022, Vol. 43

2. 青岛海洋科学与技术试点国家实验室, 青岛 266237;
3. 中国海洋大学海洋化学理论与工程技术教育部重点实验室, 青岛 266100
 , LIU Jun1,2
, LIU Jun1,2 , WANG Yi-bin1 , LIU Jia-ming1 , WU Wen-tao1,3 , FENG Yao1 , ZANG Jia-ye1 , RAN Xiang-bin1,2
, WANG Yi-bin1 , LIU Jia-ming1 , WU Wen-tao1,3 , FENG Yao1 , ZANG Jia-ye1 , RAN Xiang-bin1,2
2. Pilot National Laboratory for Marine Science and Technology (Qingdao), Qingdao 266237, China;
3. Key Laboratory of Marine Chemistry Theory and Technology, Ministry of Education, Ocean University of China, Qingdao 266100, China
磷作为浮游植物生长所必须的生源物质之一, 是海洋初级生产力的限制性营养元素[1, 2].由于人类活动的加剧, 大量的磷进入水体, 造成了富营养化等环境问题; 在近岸海域, 沉积物-水界面对水体磷循环的影响极为重要, 往往作为水体磷的主要来源[3~5], 特别是受人类活动影响较大的水产养殖区和近岸低氧区等热点区域.近岸沉积物中的磷来源主要分为外部源(河流、大气沉降和外海的交换等)[6, 7]和自生源(初级生产形成的有机颗粒物的沉降)[8].水体中的活性磷酸盐也可通过一些物理化学过程沉积在沉积物中, 包括吸附在颗粒物表面和形成自生矿物等, 从而在水体中表现为被去除或移除[9].这些被埋藏在沉积物中的磷随着有机物的再矿化作用发生形态转变, 使得无机和有机磷被释放到沉积物间隙水中, 并随着分子扩散或生物作用回到水体.这部分通过沉积物-水界面释放到水体中的磷对水环境变化和物质循环影响显著, 成为当前环境科学和海洋科学关注的热点.
磷是氧化-还原敏感参数, 底层水体溶解氧水平往往是控制沉积物-水体系磷循环的关键因子; 在富氧、低氧和无氧等不同溶解氧水平下, 磷在沉积物-水体系中呈现出不同的行为特征, 并在一定程度上改变了沉积物作为水体磷源或磷汇的角色; 溶解氧水平除影响参与沉积物-水体系磷循环的微生物群落的活性外, 还控制着其他氧化还原敏感参数的生物地球化学循环, 使得沉积物中磷的释放和保存呈现出和溶解氧变化较为规律化的响应[10~12].微生物, 特别是具有导电属性的一类微生物, 如脱硫杆菌和具有解磷作用的丝状芽孢杆菌可以通过“桥联”作用实现电子在厘米级的转移, 从而驱动沉积物中的铁和硫循环, 在一定程度上提高上层沉积物中磷的转化速率和空间[13, 14].除富营养化外, 人类活动加剧还导致水体脱氧过程的加剧, 并在全球变暖背景下(如水体层化现象加剧)变得极为典型, 潜在影响磷循环过程[15~18].有研究表明, 磷的释放通量随着底层水体溶解氧的降低而增加[19]; 这部分磷为浮游植物和其他生物的生长提供了必要的物质基础, 甚至引起有害藻类暴发等生态问题[20].可见, 近岸海域沉积物中磷的地球化学过程的变化和上覆水体环境密切相关, 是影响生态系统稳定的重要界面[15, 21, 22], 这在当前近岸海域富营养化和脱氧加剧的情况下变得尤为重要.
在机制上, 大家普遍认为铁锰氧化物在低氧条件下会发生还原溶解, 从而将大量的磷释放到水体中; 硫酸盐还原作用也会导致沉积物中有机质分解, 从而促进磷向水体的释放[23], 如波罗的海[11]和长江口[22]等.然而, 由于过程和机制的复杂性, 大多的研究集中在界面释放通量等易量化的过程上, 这导致海洋中溶解氧的变化对磷的迁移转化过程和释放机制等行为的研究还不系统, 缺少不同溶解氧水平下沉积物-水体系的对比分析和机制研究.乳山湾及其邻近海域位于山东半岛, 是重要的海洋经济贝类养殖区[24].已有研究发现, 乳山湾及其邻近海域出现了季节性低氧现象, 是进行不同溶解氧水平下沉积物-水体系磷循环研究较为理想的区域[25, 26].本研究通过沉积物柱状样的培养实验, 分析了不同溶解氧条件下柱状沉积物中磷的赋存形态、转化过程和释放通量的变化, 并利用活性物质转化模式, 从地球化学视角探讨近岸底层水体不同溶解氧状态下磷的地球化学行为和调控机制, 以期为近海环境问题研究提供科学基础.
1 材料与方法 1.1 样品采集和预处理于2016年8月在乳山湾近岸海域低氧区利用多管重力柱状取样器(KC-Denmark Ltd., Denmark)采集沉积物样品, 使用酸洗处理后的有机玻璃管(内径50 mm, 长300 mm; 预先在1% HCl溶液中浸泡48 h以上, 用Milli-Q水清洗至中性)嵌入沉积物柱状样采样器中采集柱状样, 平行采集6管; 用于培养的沉积物柱状样长150 mm, 为培养管上端留出150 mm的空间, 用于注入不同溶解氧(DO)水平的上覆水进行培养.样品采集后, 将培养管上下两端密封, 上端空间充氮气(N2)防止上层沉积物被氧化, 并抑制生物和化学转化等.采集后的柱状样品在和底层海水温度相近的条件下避光保存; 现场另采集底层海水, 经聚醚砜滤膜过滤后低温下避光保存, 用于后续培养实验.另采集1根沉积物柱状样, 在实验室充N2的手套箱内1~2 cm间隔分割, 获取沉积物湿样和间隙水样品用于各参数分析, 相关分析和观测作为背景值与培养中所获得的数据进行对比分析.
1.2 培养实验设计 1.2.1 实验装置沉积物柱状样的培养实验采用多管培养装置平行进行, 每组DO水平进行2个平行实验, 以提高实验结果的准确性.图 1是多管培养装置示意图, 上覆水体积为300 mL左右.培养时, 在进样和取样过程中沉积物-水界面应避免明显可见的物理扰动, 上覆水能够均匀地混合, 取样进样等过程尽量避免空气的污染, 上覆水溶解氧、pH、氧化还原电位(Eh)等参数在取样和进样前后能够实时监测.本实验所用玻璃和聚四氟乙烯器皿在使用前于1% HCl溶液中浸泡48 h以上, 并用Milli-Q水清洗至中性.

|
改自文献[27] 图 1 沉积物柱状样多管培养装置示意 Fig. 1 Schematic diagram of sediment columnar multi-tube culture device |
根据富氧、低氧和无氧的定义[27, 28]设置培养实验中上覆水DO条件, 将实验分为3组, 分别为富氧(213 μmol·L-1)、低氧(60 μmol·L-1)和无氧(准无氧, 2.45 μmol·L-1)培养.将上述处理好的底层海水缓慢注入到培养管上端, 进样期间水样通过进样口注入, 并顺管壁缓缓流下, 以避免沉积物表面受到扰动而出现明显的沉积物再悬浮现象.水样进样量大于上端所留空间(注满培养管后约有50 mL的水样从取样口溢出), 以赶走水柱顶端的氮气.利用蠕动泵控制培养装置中上覆水的周转时间接近于实际现场水流条件(ADCP监测显示现场往复流<0.2 m·s-1); 用于培养的上覆水通过控制氮气充入时间调节初始水样和蓄水箱的溶解氧水平, 并通过溶解氧电极实时监控, 以保证溶解氧处于稳定水平.最后将进样口和取样口密封, 培养装置置于生化培养箱(SHP 250, 上海精宏)内恒温避光进行时间序列的培养实验.培养中, 每1~2 d取样一次, 连续培养12 d; 取样结束后, 按上述进样方法加入过滤的底层海水, 使培养管上端重新充满上覆水, 并进行下一阶段实验.
1.2.3 样品采集和预处理取样时, 先从培养管上端缓缓充入氮气, 用40 mL碘量瓶通过取样口收集水样, 后用聚醚砜滤膜过滤、分样; 其中10 mL样品加入0.1 mL优级纯硝酸(HNO3)后4℃冷藏保存, 用于分析铁离子(Fe2+)和锰离子(Mn2+); 另外30 mL样品在-20℃条件下冷冻保存, 用于溶解态活性磷酸盐(DRP)、氨氮(NH4+)、硝酸盐(NO3-)、亚硝酸盐(NO2-)和溶解态有机碳(DOC)的测定.连续12 d室内培养结束后, 立即在手套箱内对沉积物柱状样进行分割, 间隔为1~2 cm.分割后得样品密封于50 mL离心管, 离心后的沉积物样品用于分析各赋存形态磷的含量, 离心获得的间隙水用于分析营养盐、Fe2+和Mn2+; 上述操作均在N2的保护下进行.实验中所用试剂纯度均为优级纯.
1.3 样品分析 1.3.1 水样分析上覆水和间隙水中溶解无机氮(DIN)和DRP利用营养盐自动分析仪(SEAL Analytical Gmbh, QuAAtro, Germany)进行测定, 其检出限分别为:DRP为0.024 μmol·L-1, NO3-(以N计, 下同)为0.015 μmol·L-1, NO2-为0.003 μmol·L-1, NH4+为0.040 μmol·L-1, 相对标准偏差<0.3%, DIN为NO3-、NO2-和NH4+之和.水样中的溶解态Fe和Mn采用ICP-MS(Agilent, 7500a, America)进行测定, 其测量误差均小于5%.间隙水中所有溶解态Fe和Mn计为Fe2+和Mn2+[29].利用微电极(Field Microsensor Multimeter, Denmark)测量上覆水DO和硫化氢(H2S).
1.3.2 沉积物分析(1) 密度、孔隙度、Eh、DO和H2S沉积物干密度和含水率的测定, 先用已知质量和体积的不锈钢环刀装满沉积物样品并压实, 然后将其置于烘箱中45℃条件下烘干至恒重, 根据前后的重量差和体积得到沉积物的干密度和含水率.取一定体积的间隙水称重, 得到间隙水的密度, 再根据沉积物的含水率, 计算得到一定体积的沉积物中间隙水的体积, 间隙水体积除以对应沉积物的体积即为孔隙度.使用校准后的氧化还原电位计(AZ-8651, pH/OPR METER)分析沉积物的Eh, 利用微电极测量沉积物DO和H2S剖面.
(2) 磷赋存形态培养实验结束后, 将沉积物进行冷冻干燥并研磨, 采用改进后的连续提取法(SEDEX方法)对磷的赋存形态进行分析, 依次提取可交换态磷(Exch-P)、铁结合态磷(Fe-P)、自生磷(Auth-P)、碎屑磷(Detr-P)和有机磷(Org-P), 各形态分析的相对标准偏差<10%.颗粒态总磷(Tot-P)为Exch-P、Fe-P、Org-P、Auth-P和Detr-P的含量之和, 活性磷(Reac-P)为Exch-P、Fe-P和Org-P的含量之和[27, 30].
另外, Fe-P提取后的溶液分2份, 一份测磷酸盐, 另一份经纯硝酸酸化后采用火焰原子吸收分光光度计(Varian, AA140/240, 美国)测铁和锰, 多次测定的相对标准偏差<10%; 此过程中所分析的铁和锰记为FeCDB和MnCDB, 为铁和锰元素可还原的非晶形氧化物或氢氧化物形态[30~33].
1.3.3 底界面过程(1) 沉积物-水界面的物质交换通量沉积物-水界面的物质交换通量可以通过培养实验上覆水中相关参数的时间变化计算得到[34]:
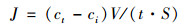
|
(1) |
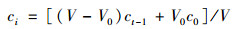
|
(2) |
式中, J为交换通量[μmol·(m2·d)-1], 正值表示溶质从沉积物中释放, 负值则表示溶质从水体清除, 即向沉积物富集; ci为进样时上覆水中物质的实际浓度(μmol·L-1); ct和ct-1为t时刻和t-1时刻上覆水样品直接测定的浓度, t为培养时间(d); V为上覆水的体积(L); S为沉积物-水界面的面积(m2); c0为每次进样时原始底层海水中物质的浓度; V0为每次取样的体积(L).本文分别计算了DRP、NH4+、NO3-、Fe2+、Mn2+、H2S和DOC等交换通量.
(2) 培养中沉积物中磷的损失量
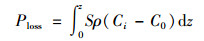
|
(3) |
式中, Ploss为培养后沉积物损失的总磷(μmol); S为培养管横截面积(cm2); ρ为沉积物的干密度(g·cm-3); C0和Ci分别为沉积物培养前后磷的含量(μmol·g-1); z为沉积物深度(cm).
1.3.4 间隙水模型基于一维稳态模型, 对间隙水剖面化学参数变率进行模拟, 得到其浓度拟合曲线和周转速率(生产和消耗)进行模拟[35]:
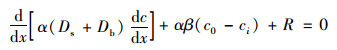
|
(4) |
式中, x为沉积物深度(cm), c为间隙水中溶质的浓度(μmol·L-1), α为孔隙度(cm3·cm-3), Ds为曲率校正后的分子扩散系数(cm2·d-1), Db为生物扩散系数(cm2·d-1), β为生物灌溉系数(cm2·d-1), c0和ci分别为上覆水和间隙水中溶质的浓度(μmol·L-1), R为净产生或消耗速率[周转速率, nmol·(cm3·d)-1], 其结果由Profile V 1.0最优模拟得到[29, 36].
1.3.5 数据处理本研究结果运用Origin 2018进行数据拟合和制图, 采用SPSS 26.0对不同培养阶段的结果进行Pearson相关分析(Pearson correlation coefficient), 在P≤0.01和P≤0.05这2个显著水平上进行统计分析.
2 结果与分析 2.1 上覆水DO、Eh、pH和H2S随时间变化在培养过程中, 富氧、低氧和无氧条件下上覆水溶解氧(μmol·L-1)范围(平均值±标准偏差)分别为: 209.9~219.6 (213.4±3.22)、55.3~65.7 (60.2±3.52)和0.1~4.4 (2.5±1.25). Eh变化依次为:富氧状态>低氧状态>无氧状态, 均表现为随时间先减小后增大的趋势(图 2). pH在富氧条件下随着培养时间的增长出现波动, 总体保持稳定; 在低氧和无氧条件下逐渐降低, 且在无氧时变化幅度更大. H2S在富氧状态下浓度较低, 无明显变化趋势, 低氧和无氧状态下呈现上升趋势, 且在培养实验后期增长幅度尤为明显.

|
图 2 上覆水体中DO、Eh、pH和H2S随时间的变化 Fig. 2 Variations in DO, Eh, pH, and H2S in the overlying seawater over time |
不同溶解氧水平下, 上覆水相关参数浓度及其在沉积物-上覆水界面交换通量均具有显著差异(图 3).富氧状态培养实验中, DRP在沉积物-水界面的交换通量以上覆水向沉积物扩散的方向呈现线性增加的趋势, 即向沉积物埋藏, 并在第3 d达到最大值81.3 μmol·(m2·d)-1, 在3~5 d交换通量快速降低并在培养后期基本保持稳定(图 3中a-1); 低氧状态下, DRP从沉积物释放到上覆水中, 其交换通量在前3 d快速增长, 达到最大值822 μmol·(m2·d)-1后降低, 在第5 d降至348 μmol·(m2·d)-1后继续增长, 但速度逐渐变慢(图 3中a-2); 无氧状态下DRP为从沉积物向上覆水释放, 在前3 d快速增长, 达到最大值1 320 μmol·(m2·d)-1后缓慢降低(图 3中a-3).培养前后, 上覆水中DRP含量在富氧条件下相对于培养前减少, 即向沉积物埋藏; 在低氧和无氧条件下相对于培养前增加, 即呈现向水体释放的趋势.

|
a表示DRP, b表示NH4+, c表示NO3-, d表示Fe2+, e表示Mn2+, f表示H2S, g表示DOC; 1表示富氧培养, 2表示低氧培养, 3表示无氧培养; DRP、NH4+、NO3-、Fe2+、Mn2+和H2S的浓度单位为μmol·L-1, 交换通量单位为μmol·(m2·d)-1; DOC的浓度单位为mg·L-1, 交换通量单位为mg·(m2·d)-1; 交换通量正值表示该要素由沉积物向上覆水扩散 图 3 沉积物培养过程中上覆水DRP、NH4+、NO3-、Fe2+、Mn2+、H2S和DOC的浓度及其在沉积物-上覆水界面交换通量的变化 Fig. 3 Concentrations and fluxes of DRP, NH4+, NO3-, Fe2+, Mn2+, H2S, and DOC in the incubation water at sediment-overlying water interface |
富氧状态下, NH4+浓度呈现先增加后减小的趋势, Fe2+和Mn2+浓度变化存在较大波动, NO3-浓度随培养时间变长逐渐增加, H2S与之相反.低氧状态下, 上覆水DRP、NH4+和Fe2+相对于培养前显著增加, 远远高于培养前的初始浓度, 交换通量均为沉积物向水体的净扩散.无氧状态下, NH4+、Mn2+和H2S浓度相对培养前增加, NO3-浓度随培养时间变长逐渐减小, Fe2+浓度在培养初期迅速增加之后逐渐减小, 其交换通量也由沉积物向水体扩散逐渐转变为由水体进入沉积物.低氧状态下上覆水DOC浓度缓慢升高, 无氧状态下浓度存在一定的波动(图 3).
2.3 培养前后间隙水各要素浓度变化3种培养条件下沉积物柱状样间隙水中的Fe2+和Mn2+存在显著差异, 总体上Mn2+的含量在培养后相对于培养前均有所减少; 在富氧状态下Fe2+的含量相比培养前减少且含量较低, 其他2种状态下Fe2+含量的变化相似, 在0~2 cm层相比培养前减少, 在5~10 cm层增加且变化程度随深度增大, 低氧和无氧状态下Fe2+含量显著高于富氧状态.间隙水中的DRP含量在富氧状态下相比培养前几乎不变, 在低氧和无氧状态下含量相比培养前增加, 且增加程度随深度呈增加趋势.间隙水中NH4+的含量在富氧状态下相比培养前减少, 低氧和无氧状态下在3~10 cm层相比培养前增加, 并呈现随深度增加的趋势.间隙水中NO3-的含量相比培养前减少, 且变化程度在3~4 cm层达到峰值, 在一定深度以下变化幅度减小, 且浓度较低.尽管不同培养条件下, 间隙水各要素发生了较大变化, 但总体上各要素在培养前后随深度分布呈现较为相似的变化规律(图 4).

|
a表示DRP, b表示NH4+, c表示NO3-, d表示Fe2+, e表示Mn2+; 1表示培养前, 2表示富氧培养, 3表示低氧培养, 4表示无氧培养; DRP、NH4+、NO3-、Fe2+和Mn2+的浓度单位为μmol·L-1, 周转速率单位为10-2μmol·(L·d)-1 图 4 培养前后沉积物间隙水中DRP、NH4+、NO3-、Fe2+和Mn2+的浓度变化和周转速率 Fig. 4 Concentration changes and turnover rates of DRP, NH4+, NO3-, Fe2+, and Mn2+ in the pore water of sediment pre-incubation and post-incubation experiments |
培养前后Detr-P均为主要的颗粒磷的赋存形态, 且变化较小.培养前沉积物柱状样中Exch-P含量随深度波动较小, 在8~10 cm层有显著增加; Auth-P含量总体上随深度缓慢增大; Fe-P、Detr-P、Org-P和Tot-P的含量在0~4 cm层随深度变化剧烈; FeCDB的含量在0~4 cm层和6~10 cm层随深度先升高后减小; MnCDB的变化和FeCDB相反, 在4~6 cm层含量先减小后增大, 而后含量逐渐减小.富氧培养前后, 沉积物中磷的形态变化主要发生在0~3 cm层, 其中Exch-P、Fe-P和Org-P含量随深度变化剧烈, Auth-P含量在整个深度范围下培养前后变化较小.低氧培养前后, 各组分含量在0~4 cm层变化剧烈.无氧条件下培养前后, Auth-P和Org-P含量在0~4 cm层变化剧烈, 在4~6 cm层含量基本保持不变.3种培养方式FeCDB的含量变化主要发生在沉积物上层(0~4 cm), 而MnCDB在整个沉积物中变化均较大(图 5).

|
a表示Exch-P, b表示Fe-P, c表示Auth-P, d表示Detr-P, e表示Org-P, f表示Reac-P, g表示Tot-P, h表示FeCDB, i表示MnCDB; 1表示富氧培养, 2表示低氧培养, 3表示无氧培养; 含量单位均为μmol·g-1 图 5 不同培养条件下沉积物柱状样不同赋存形态磷、FeCDB和MnCDB在培养前后的剖面变化 Fig. 5 Vertical changes in phosphorus forms, FeCDB, and MnCDB between pre-incubation and post-incubation under different oxygen levels |
间隙水中磷的形态和含量变化取决于溶解氧、pH、氧化-还原敏感元素浓度的高低或组成和沉积物的理化性质的差异[37].间隙水中的NO3-的剖面变化可作为氧渗透深度的定性指标, 同时NO3-和NH4+的剖面变化也可作为沉积物中有机碳降解和硝酸盐异化还原为铵作用强度的指标[38], 二者在沉积物中的变化和溶解氧的变化相关联.富氧状态下溶解氧剖面的渗透深度在1.5~2.5 cm, 低氧状态下渗透深度在1.5~2.0 cm, 即低氧状态下这个深度会变浅.由于沉积物中存在氧和多种氧化物, 因此沉积物中有机质在早期成岩过程中可能存在多种氧化还原序列(反硝化、铁锰氧化物还原和硫酸盐还原等)共存的机制, 影响间隙水中各要素的剖面分布(图 4和图 5).
富氧状态下, 上覆水的DRP浓度随培养时间增加不断地减小(图 3中a-1), 同时沉积物上层间隙水中的DRP浓度处于较低水平, 这种变化可能是由于DRP被富氧环境下沉积物中的铁氧化物所吸附, 并以Fe-P的形式向沉积物沉淀[39].上覆水体中的H2S浓度随培养时间增加而减少(图 3中f-1), 并呈现向间隙水和沉积物双向转移的特征, 沉积物中存在以H2S作为还原剂的铁氧化物还原, 这一过程向间隙水释放DRP.在富氧环境下, 上覆水Eh较高, 间隙水释放的活性铁或水体存在的活性铁会被氧化并吸附水体中的磷[40], 所以间隙水中的DRP由于存在吸附和转化等过程其含量和培养前一样均维持在较低水平(图 4中a-1和a-2), 并导致Fe2+含量相较培养前显著减少(图 4中d-1和d-2), 即通过形态转化、向上覆水释放和形成Fe-P沉淀等系列过程实现磷在沉积物-上覆水-沉积物的旋回式跨界面转移.
在低氧状态下, 上覆水DRP、NH4+和Fe2+相对于培养前显著增加, 交换通量均为沉积物向水体的净释放, 上覆水体中H2S浓度随培养时长增加而增加(图 3); H2S从沉积物中释放出来, 表明沉积物中有微生物硫酸盐还原作用和铁氧化物还原溶解作用两个过程共同存在的现象.间隙水中的DRP在培养前后含量显著提高, 这一DRP变化量和Fe2+变化量呈显著正相关[(P≤0.01, 图 6(b)], 且培养后的DRP和Fe2+随深度呈现相似的变化趋势(图 4中a-3和d-3).由模拟可知, 间隙水中DRP有显著生成趋势(图 4中a-3).在低氧状态下, 上覆水的Eh较低(图 2), 铁氧化物在沉积物-水界面多被还原, DRP从而由铁氧化物中释放出来[41, 42], 这一反应过程在更深的缺氧或无氧的沉积层中表现尤为明显, 进而促进了有机物的矿化降解和铁氧化物还原溶解, 使得DRP和Fe2+进入间隙水; 释放出的DRP一部分会吸附在沉积物的细颗粒物上, 另一部分会扩散到上覆水中.上述转化过程还引起上层沉积物中释放出去的Fe2+被表层沉积物吸附后发生再氧化, 导致上层间隙水中Fe2+浓度相较培养前变小.低氧状态下上覆水中DOC交换通量大于0, 且大小随着培养时间上下波动(图 3中g-2), 这表明沉积物中存在一定的有机质降解过程, 从而向水体中释放DOC.在多数情况下, 沉积物-水体系的活性磷大多来自有机质的降解; 在有机质埋藏时, 部分沉积的有机质发生分解, 这一过程也会释放出一定量的DRP[43].低氧状态下上覆水的DRP和NH4+含量显著高于富氧状态, 表明在此条件下有更多的DRP和NH4+通过沉积物释放到水体中, 从而较富氧环境具有更强的调节和补充水体中DRP浓度的能力[29].总体而言, 低氧状态下沉积物-水界面的地球化学过程和理想的早期成岩模式最为接近, 且有利于更多的DRP由沉积物进入水环境.

|
图 6 不同培养条件培养前后沉积物剖面磷形态和间隙水中各参数变化量相关性分析 Fig. 6 Correlation of phosphorus species in sediment vertical profile and variation in pore water parameters between the pre- and post-incubations under different incubation conditions |
在无氧状态下, 上覆水体中Eh值小于零(图 2), 水体呈现较富氧和低氧更强的还原性.培养中, 上覆水Fe2+与NO3-和H2S与NH4+两组参数的浓度变化趋势大致相反, 其中Fe2+和NO3-浓度随培养时间变长逐渐地减小, 且Fe2+浓度在培养后期趋于零(图 3中d-3), 表明沉积物存在较弱的Fe2+释放过程.DRP的交换通量和NH4+交换通量显著正相关[P≤0.01, 图 7(c)], 显示沉积物处于无氧状态时发生了硝酸盐异化还原成铵和硫酸盐还原作用两个主导过程.硫酸盐还原产生的H2S可以作为铁氧化物的还原剂发生铁异化还原产生铁硫化物沉淀, 降低了DRP在铁氧化物上吸附, 从而促使其释放到水体中[44].上覆水中DOC浓度在培养后期交换通量接近于零(图 3中g-3), 表明有机质矿化降解在无氧条件下不像低氧状态下那样活跃.

|
图 7 不同培养条件沉积物-水界面DRP交换通量和其他参数交换通量相关性分析 Fig. 7 Correlation of DRP exchange fluxes and other parameters at sediment-water interface from different incubations |
影响沉积物-水界面磷释放的过程包括氧化还原反应、吸附解吸、矿物相溶解和有机物的矿化等[45~47], 其中具有活性的磷形态是沉积物中较易发生转化的部分(图 5), 同时控制了界面DRP的交换通量(图 3).在不同的海洋生态系统中, 沉积物中总磷含量和总储量的变化受颗粒物沉积速率和浮游植物初级生产力水平的影响[48, 49].在季节性缺氧的河口和近海海洋沉积物中, 磷的永久埋藏主要为有机磷、自生钙磷或生物钙磷的形式[50], 即转化沉积或埋藏过程.在永久性缺氧近海, 铁结合磷也可作为磷汇的赋存形态长期埋藏[51, 52], 而Reac-P是能从沉积物释放到上覆水体中并被藻类和浮游植物消耗的磷形态的上限[48, 53, 54].沉积物氧气的浓度和穿透深度在一定程度上控制着上述早期成岩过程, 从而在沉积物-间隙水体系中产生了多样化的氧化还原序列(图 4和图 5).
富氧条件下, Tot-P含量相对于培养前有所增加, 且培养前后Exch-P的变化量和Fe-P变化量呈显著正相关[P≤0.01, 图 6(a)].Fe-P受环境氧化还原条件变化的影响较大[40], 导电细菌是沉积物中Fe-P形成的驱动因素之一[13], 底层水体富氧状态使得铁氧化物和Fe-P在表层沉积物中积累, Fe-P含量因此显著增加(表 1), 磷被铁氧化物结合后从间隙水转移到沉积物中, 且和间隙水中Fe2+的变化过程相一致, 这也使得Fe-P对Tot-P的含量的贡献最大.由于富氧状态下上覆水环境呈弱碱性, pH存在较小的波动(图 2), 限制了部分磷的转换, 这使得培养前后沉积物剖面Auth-P变化较小(图 5中c-1).通常在偏碱性水环境下, Auth-P几乎不会发生溶解, 只有在介质中pH突然下降时, Auth-P才有部分发生溶解[30, 42].上层沉积物中的Org-P在富氧状态下相较于培养前减少(图 5中e-1), 有机质发生矿化向间隙水中释放DRP, 但在深层沉积物中的Org-P在培养前后几乎保持不变.富氧条件下Reac-P含量显著增加(表 1), 沉积物表现为水体磷汇的作用.Detr-P是海洋环境中最不活跃的磷组分[30, 55], 培养前后几乎保持不变(图 5中d-1).
|
|
表 1 3种培养条件下各赋存形态磷相比于初始状态的损失量1) Table 1 Loss of phosphorus forms under three incubation conditions |
低氧条件下, Exch-P在上层沉积物中相对于培养前有所减少(图 5中a-2), Exch-P由于较高的活性, 是沉积物中生物利用度最高的磷形态; 当上覆水体中磷浓度较低时, Exch-P很容易通过沉积物-水界面释放扩散到上覆水中并被浮游生物利用[56, 57]. 铁氧化物是沉积物中磷酸盐的储库, 在铁氧化物溶解时, DRP可被释放到间隙水中[58].上层沉积物培养后Fe-P和FeCDB含量高于培养前(图 5中b-2和h-2), 且沉积物柱状样中的FeCDB前后变化量和间隙水中的DRP和Fe2+呈显著负相关[P≤0.01, 图 6(b)], 表明该部分Fe-P主要为可还原性铁氧化物结合磷, 下层含量随深度降低, 且变化趋势和培养前大致相当, 这也说明Fe-P在沉积物-间隙水体系磷转化中地位重要.除向DRP转化外, 部分铁氧化物结合磷在无氧条件下还可能进一步转化为还原性铁结合磷的次生矿物(蓝铁矿), 这一产物对磷酸盐的保存能力较强[59].低氧状态下培养前后Auth-P变化量和Fe-P变化量呈显著负相关[P≤0.01, 图 6(b)], 且上层沉积物Auth-P含量较培养前减少(图 5中c-2), 上覆水的pH随培养时间逐渐减小, 表明溶解氧降低使得上层沉积物中部分Auth-P溶解.和Detr-P一样, Auth-P在弱碱性水环境中的活性较低, 化学性质较为稳定[53, 60], 对沉积物间隙水或上覆水中磷的积累的贡献因此很小.在沉积物深层, Org-P含量在培养前后几乎保持不变, 有机质矿化降解作用较弱.可见, 在沉积物中Org-P在低氧条件下比富氧状况下的积累程度更高, 即有利于其保存.
无氧状态下Exch-P的变化量和间隙水中Mn2+的变化量呈显著负相关(P≤0.01), 和NH4+正相关[P≤0.05, 图 6(c)], 表明沉积物中存在硝酸盐异化还原成铵作用过程[38]. Fe-P含量在0~5 cm层相较于培养前减少, 在5~10 cm层相对培养前随深度增加(图 5中b-3), 且Fe-P前后变化量和间隙水中的DRP和Fe2+呈显著正相关(P≤0.01), 同时Org-P前后变化量和间隙水中的DRP呈显著负相关[P≤0.01, 图 6(c)], 表明沉积物在无氧状态下发生了甲烷厌氧氧化反应, 这一过程也向水体中释放出DRP, 并在沉积物更深处生成还原态铁结合磷, 导致Fe-P含量的增加.同时在溶解氧偏低的状态下, 导电细菌的活性较低, 也会抑制磷的释放[13].无氧状态下, Auth-P和Fe-P变化量及间隙水DRP、NH4+和Fe2+变化量均呈显著正相关[P≤0.05, 图 6(c)], 说明培养过程中Auth-P和Fe-P的生成可能有共同的来源.当底层水体中氧化还原条件发生变化时, 特别是溶解氧趋于耗尽时, 上层沉积物中的Auth-P和Fe-P的转化作用向水体中释放出DRP, 而在深层沉积物中Auth-P对磷的埋藏起重要作用.无氧状态下培养前后Detr-P变化量和Exch-P变化量呈显著正相关[P≤0.05, 图 6(c)], 也表明Detr-P所依存的颗粒物对Exch-P的吸附作用较强.
对比培养前后, Reac-P在表层沉积物中的含量为: 富氧>低氧>培养前>无氧, 表明低氧和无氧状态对表层沉积物活性磷含量和组成的影响较大, 沉积物呈现出更强的磷源的特征(图 5), 且3种溶解氧条件下Reac-P培养前后变化量和Fe-P变化量均为正相关关系(图 6), 表明Fe-P的变化对于Reac-P变化起主导作用.培养前后磷形态和交换通量的显著变化表明无氧和低氧水体中沉积物底界面的磷释放可作为磷源影响上层水体磷循环, 并在调节初级生产力方面发挥重要作用.
3.3 溶解氧变化下沉积物-水界面磷的转化浮游植物所产生的有机质降解释放的NH4+通量和DRP通量表现为接近16∶1的量比[61].当水体中的DRP均来源于沉积物中铁氧化物的还原溶解, 则Fe2+和DRP的比值应在2~20范围之间[60, 62, 63].在富氧状态下, NH4+的交换通量和DRP的交换通量的平均比值(N/P)为4.46, 低氧状态下N/P平均值为5.77, 无氧状态下N/P平均值为7.69(表 2), 远低于Redfield比值16, 这表明近海沉积物中磷来源较为复杂.这一比值和长江口等低氧水域较为接近[29], 即反映了磷组成的多源性.
|
|
表 2 3种溶解氧条件下磷的转化速率、N/P和Fe/P对比1) Table 2 Phosphorus transformation, N/P and Fe/P under three incubation conditions |
富氧培养条件下, 沉积物-水界面Fe2+的交换通量和DRP的交换通量的比值(Fe/P)均接近或低于2.低氧条件下, Fe/P绝大部分在2~20范围内, 且间隙水中N/P均小于16, N/P和NH4+交换通量正相关[P≤0.05, 图 7(b)].当水体溶解氧较低时, 由于沉积物对磷的保存能力降低, DRP向间隙水和上覆水中增加释放.Fe/P和NO3-交换通量显著负相关[P≤0.05, 图 7(b)], Fe2+的释放受到沉积物硝酸盐异化还原作用的影响, 且铁氧化物和NO3-的还原存在竞争关系.可见, 在富氧和低氧条件下, 除Fe-P还原溶解释放DRP之外, 还存在Org-P等其他磷源的转化.无氧条件下, Fe/P比值均在2~20范围之外, 随着培养时间的增加Fe2+从沉积物向水体释放逐渐转变为水体向沉积物扩散, 说明Fe-P还原溶出的Fe2+进入水体后又被颗粒物吸附从而离开水体[64].无氧状态下, Fe/P和Fe2+交换通量显著正相关(P≤0.01, 图 6), 且硫酸盐还原产生了大量的硫化物, 硫化物和Fe2+反应形成FeS并埋藏, Fe2+浓度随之急剧下降, 从而导致Fe/P比例较低[65].
本研究所采用的培养装置所营造的实验条件较为接近现场状况, 培养中使用了多根柱状沉积物进行平行实验, 以降低实验的误差, 且采用了数值模拟的手段验证培养结果.如图 8所示, 本文中培养实验所采用溶解氧条件参考世界其他海域, 并将结果和世界其他海域进行对比, 在低氧和富氧条件下DRP交换通量(表 2)和溶解氧的关系与其他海域的研究结果较为一致; 且在之前涉及培养实验相关的研究中发现, 培养时长超过12 d后各要素的转化速率大致趋于稳定[27], 另外, 由于自生磷生成速率较慢, 培养过程中可能转化不完全[29], 以上均说明本实验研究的合理性和可行性.不过, 本研究所得交换通量未考虑实际海洋环境中溶解氧条件变化下其他物理过程和生物活动等的协同作用, 因此跟实际状况相比会有所出入.此外, 尽管这些沉积物均来自同一采泥器, 然而这些沉积物在同一水平面上也会存在细微的理化性质的差异, 如Detr-P(一种较为稳定的磷形态)培养前后的变化可能由不同沉积物柱状样品细微的区域性差异所致, 这一差异也会对实验结果的对比分析产生一定的影响.将培养结果和理想状态下早期成岩过程进行对比, 发现低氧条件下的变化比较符合理想状态下氧化还原特征(图 9)[66, 67].相关实验明确了不同溶解氧水平下磷的地球化学转化模式和相关机制, 可为近岸不同溶解氧水平下沉积物-间隙水体系磷转化埋藏和释放规律以及水环境演变研究提供科学支撑.

|
图 8 世界不同海域磷酸盐交换通量和底层水溶解氧的关系 Fig. 8 Relationships between benthic DRP fluxes and bottom water oxygen in different coastal seas around the world |

|
(a)富氧培养下沉积物间隙水各要素的剖面周转变化; (b)低氧培养下沉积物间隙水各要素的剖面周转变化; (c)无氧培养下沉积物间隙水各要素的剖面周转变化; 框中实线表示该要素在培养中发生富集, 虚线表示该要素在培养中被消耗; 灰色方框区域表示间隙水各要素的在该深度上的剖面周转变化和磷的剖面富集对应于左侧理想的早期成岩模式 图 9 不同溶解氧控制的早期成岩作用下磷循环过程对比 Fig. 9 Phosphorus cycling and early diagenesis processes under different oxygen conditions |
(1) 沉积物-水界面磷酸盐交换通量受到溶解氧水平的影响存在较大差异, 低氧和无氧条件下沉积物具有较富氧状态高的磷释放通量, 且沉积物-水界面磷循环和N、Fe、S等多种元素相耦合, 低氧状态下间隙水化学参数变化和理想状态下的早期成岩模式较为接近.
(2) 溶解氧变化控制着沉积物中磷赋存形态的转化, 从而影响了磷的生物有效性, 特别是Exch-P和Fe-P; 沉积物中Reac-P含量在溶解氧水平高的条件下增加, 增大了其生物可利用的潜力.
(3) 富氧条件下沉积物具有较高的磷保存能力, 低氧水体的沉积物对磷的保留能力减弱, 低氧或无氧条件下沉积物中还存在硫酸盐还原和硝酸盐异化还原成铵等早期成岩路径影响磷循环过程.同时, 低氧条件下, Fe-P的还原溶解是水体中DRP的最主要来源, 有机质的矿化也显著促进了磷的迁移和释放.
| [1] | Lomas M W, Burke A L, Lomas D A, et al. Sargasso Sea phosphorus biogeochemistry: an important role for dissolved organic phosphorus (DOP)[J]. Biogeosciences, 2010, 7(2): 695-710. DOI:10.5194/bg-7-695-2010 |
| [2] | Lürling M, van Oosterhout F. Controlling eutrophication by combined bloom precipitation and sediment phosphorus inactivation[J]. Water Research, 2013, 47(17): 6527-6537. DOI:10.1016/j.watres.2013.08.019 |
| [3] | Pan F, Guo Z R, Cai Y, et al. Cyclical patterns and (im)mobilization mechanisms of phosphorus in sediments from a small creek estuary: evidence from in situ monthly sampling and indoor experiments[J]. Water Research, 2020, 171. DOI:10.1016/j.watres.2020.115479 |
| [4] | Smith L, Watzin M C, Druschel G. Relating sediment phosphorus mobility to seasonal and diel redox fluctuations at the sediment-water interface in a eutrophic freshwater lake[J]. Limnology and Oceanography, 2011, 56(6): 2251-2264. DOI:10.4319/lo.2011.56.6.2251 |
| [5] |
刘永九, 黄素珍, 张璐, 等. 洪湖国际重要湿地沉积物磷空间分布特征及释放风险[J]. 环境科学, 2021, 42(7): 3198-3205. Liu Y J, Huang S Z, Zhang L, et al. Spatial distribution characteristics of phosphorus fractions and release risk in sediments of Honghu international importance wetland[J]. Environmental Science, 2021, 42(7): 3198-3205. DOI:10.13227/j.hjkx.202009090 |
| [6] | Mahowald N, Jickells T D, Baker A R, et al. Global distribution of atmospheric phosphorus sources, concentrations and deposition rates, and anthropogenic impacts[J]. Global Biogeochemical Cycles, 2008, 22(4). DOI:10.1029/2008GB003240 |
| [7] | Paytan A, Cade-Menun B J, McLaughlin K, et al. Selective phosphorus regeneration of sinking marine particles: evidence from 31P-NMR[J]. Marine Chemistry, 2003, 82(1-2): 55-70. DOI:10.1016/S0304-4203(03)00052-5 |
| [8] | Paytan A, McLaughlin K. The oceanic phosphorus cycle[J]. Chemical Reviews, 2007, 107(2): 563-576. DOI:10.1021/cr0503613 |
| [9] | Defforey D, Paytan A. Phosphorus cycling in marine sediments: advances and challenges[J]. Chemical Geology, 2018, 477: 1-11. DOI:10.1016/j.chemgeo.2017.12.002 |
| [10] | Kang M X, Peng S, Tian Y M, et al. Effects of dissolved oxygen and nutrient loading on phosphorus fluxes at the sediment-water interface in the Hai River Estuary, China[J]. Marine Pollution Bulletin, 2018, 130: 132-139. DOI:10.1016/j.marpolbul.2018.03.029 |
| [11] | Hylén A, van de Velde S J, Kononets M, et al. Deep-water inflow event increases sedimentary phosphorus release on a multi-year scale[J]. Biogeosciences, 2021, 18(9): 2981-3004. DOI:10.5194/bg-18-2981-2021 |
| [12] | Pan F, Guo Z R, Cai Y, et al. Remobilization and hypoxia-dependent migration of phosphorus at the coastal sediment-water interface[J]. Journal of Hazardous Materials, 2021, 411. DOI:10.1016/j.jhazmat.2021.125078 |
| [13] | Sulu-Gambari F, Seitaj D, Meysman F J R, et al. Cable bacteria control iron-phosphorus dynamics in sediments of a coastal hypoxic basin[J]. Environmental Science & Technology, 2016, 50(3): 1227-1233. |
| [14] | Seitaj D, Schauer R, Sulu-Gambari F, et al. Cable bacteria generate a firewall against euxinia in seasonally hypoxic basins[J]. Proceedings of the National Academy of Sciences of the United States of America, 2015, 112(43): 13278-13283. DOI:10.1073/pnas.1510152112 |
| [15] | Rabalais N, Cai W J, Carstensen J, et al. Eutrophication-driven deoxygenation in the coastal ocean[J]. Oceanography, 2014, 27(1): 172-183. DOI:10.5670/oceanog.2014.21 |
| [16] | Breitburg D, Levin L A, Oschlies A, et al. Declining oxygen in the global ocean and coastal waters[J]. Science, 2018, 359(6371). DOI:10.1126/science.aam7240 |
| [17] | Carstensen J, Andersen J H, Gustafsson B G, et al. Deoxygenation of the Baltic Sea during the last century[J]. Proceedings of the National Academy of Sciences of the United States of America, 2014, 111(15): 5628-5633. DOI:10.1073/pnas.1323156111 |
| [18] | Altieri A H, Gedan K B. Climate change and dead zones[J]. Global Change Biology, 2015, 21(4): 1395-1406. DOI:10.1111/gcb.12754 |
| [19] | Conley D J, Carstensen J, Vaquer-Sunyer R, et al. Ecosystem thresholds with hypoxia[J]. Hydrobiologia, 2009, 629(1): 21-29. DOI:10.1007/s10750-009-9764-2 |
| [20] | Smith V H, Schindler D W. Eutrophication science: where do we go from here?[J]. Trends in Ecology & Evolution, 2009, 24(4): 201-207. |
| [21] | Lenz C, Jilbert T, Conley D J, et al. Are recent changes in sediment manganese sequestration in the euxinic basins of the Baltic Sea linked to the expansion of hypoxia?[J]. Biogeosciences, 2015, 12(16): 4875-4894. DOI:10.5194/bg-12-4875-2015 |
| [22] |
刘军, 臧家业, 冉祥滨, 等. 长江口低氧区沉积物中磷的形态及其环境意义[J]. 环境科学, 2017, 38(8): 3243-3253. Liu J, Zang J Y, Ran X B, et al. Sedimentary phosphorus speciation in the coastal hypoxic area of Changjiang Estuary and its environmental significance[J]. Environmental Science, 2017, 38(8): 3243-3253. DOI:10.13227/j.hjkx.201701140 |
| [23] | Picard A, Gartman A, Cosmidis J, et al. Authigenic metastable iron sulfide minerals preserve microbial organic carbon in anoxic environments[J]. Chemical Geology, 2019, 530. DOI:10.1016/j.chemgeo.2019.119343 |
| [24] |
崔毅, 马绍赛, 陈碧鹃, 等. 乳山湾生物理化环境现状研究[J]. 中国水产科学, 1999, 6(4): 76-80. Cui Y, Ma S S, Chen B J, et al. Study on the status of bio-physic-chemical environment in Rushan Bay[J]. Journal of Fishery Sciences of China, 1999, 6(4): 76-80. DOI:10.3321/j.issn:1005-8737.1999.04.018 |
| [25] |
刘军, 臧家业, 冉祥滨, 等. 乳山湾外低氧海域沉积物中有机碳、氮、磷及其形态特征分析[J]. 海洋科学, 2012, 36(7): 70-78. Liu J, Zang J Y, Ran X B, et al. Characteristics of different forms of nitrogen and phosphorus and organic carbon in the sediments of low-oxygen zones in adjacent Rushan Bay[J]. Marine Science, 2012, 36(7): 70-78. |
| [26] |
冉祥滨, 臧家业, 韦钦胜, 等. 乳山湾邻近海域低氧现象及成因浅析[J]. 海洋科学进展, 2012, 30(3): 347-356. Ran X B, Zang J Y, Wei Q S, et al. Hyposia and its cause of formation in the adjacent waters of Rushan Bay[J]. Advances in Marine Science, 2012, 30(3): 347-356. DOI:10.3969/j.issn.1671-6647.2012.03.005 |
| [27] | Liu J, Krom M D, Ran X B, et al. Sedimentary phosphorus cycling and budget in the seasonally hypoxic coastal area of Changjiang Estuary[J]. Science of the Total Environment, 2020, 713. DOI:10.1016/j.scitotenv.2019.136389 |
| [28] | Middelburg J J, Levin L A. Coastal hypoxia and sediment biogeochemistry[J]. Biogeosciences, 2009, 6(7): 1273-1293. DOI:10.5194/bg-6-1273-2009 |
| [29] | Liu J, Zang J Y, Zhao C Y, et al. Phosphorus speciation, transformation, and preservation in the coastal area of Rushan Bay[J]. Science of the Total Environment, 2016, 565: 258-270. DOI:10.1016/j.scitotenv.2016.04.177 |
| [30] | Ruttenberg K C. Development of a sequential extraction method for different forms of phosphorus in marine sediments[J]. Limnology and Oceanography, 1992, 37(7): 1460-1482. DOI:10.4319/lo.1992.37.7.1460 |
| [31] | Huerta-Diaz M A, Tovar-Sánchez A, Filippelli G, et al. A combined CDB-MAGIC method for the determination of phosphorus associated with sedimentary iron oxyhydroxides[J]. Applied Geochemistry, 2005, 20(11): 2108-2115. DOI:10.1016/j.apgeochem.2005.07.009 |
| [32] | Slomp C P, Epping E H G, Helder W, et al. A key role for iron-bound phosphorus in authigenic apatite formation in north Atlantic continental platform sediments[J]. Journal of Marine Research, 1996, 54(6): 1179-1205. DOI:10.1357/0022240963213745 |
| [33] | Slomp C P, Mort H P, Jilbert T, et al. Coupled dynamics of iron and phosphorus in sediments of an oligotrophic coastal basin and the impact of anaerobic oxidation of methane[J]. PLoS One, 2013, 8(4). DOI:10.1371/journal.pone.0062386 |
| [34] | Fisher M M, Reddy K R. Phosphorus flux from wetland soils affected by long-term nutrient loading[J]. Journal of Environmental Quality, 2001, 30(1): 261-271. DOI:10.2134/jeq2001.301261x |
| [35] | Boudreau B P. On the equivalence of nonlocal and radial-diffusion models for porewater irrigation[J]. Journal of Marine Research, 1984, 42(3): 731-735. DOI:10.1357/002224084788505924 |
| [36] | Berg P, Risgaard-Petersen N, Rysgaard S. Interpretation of measured concentration profiles in sediment pore water[J]. Limnology and Oceanography, 1998, 43(7): 1500-1510. DOI:10.4319/lo.1998.43.7.1500 |
| [37] | Lukkari K, Hartikainen H, Leivuori M. Fractionation of sediment phosphorus revisited. Ⅰ: fractionation steps and their biogeochemical basis[J]. Limnology and Oceanography: Methods, 2007, 5(12): 433-444. DOI:10.4319/lom.2007.5.433 |
| [38] | Homoky W B, Severmann S, McManus J, et al. Dissolved oxygen and suspended particles regulate the benthic flux of iron from continental margins[J]. Marine Chemistry, 2012, 134-135: 59-70. DOI:10.1016/j.marchem.2012.03.003 |
| [39] | Pan F, Liu H T, Guo Z R, et al. Effects of tide and season changes on the iron-sulfur-phosphorus biogeochemistry in sediment porewater of a mangrove coast[J]. Journal of Hydrology, 2019, 568: 686-702. DOI:10.1016/j.jhydrol.2018.11.002 |
| [40] | Zhou A M, Wang D S, Tang H X. Phosphorus fractionation and bio-availability in Taihu Lake (China) sediments[J]. Journal of Environmental Sciences, 2005, 17(3): 384-388. DOI:10.3321/j.issn:1001-0742.2005.03.007 |
| [41] | Schenau S J, De Lange G J. Phosphorus regeneration vs. burial in sediments of the Arabian Sea[J]. Marine Chemistry, 2001, 75(3): 201-217. DOI:10.1016/S0304-4203(01)00037-8 |
| [42] | Mao C P, Li T N, Rao W B, et al. Chemical speciation of phosphorus in surface sediments from the Jiangsu coast, East China: influences, provenances and bioavailabilities[J]. Marine Pollution Bulletin, 2021, 163. DOI:10.1016/j.marpolbul.2020.111961 |
| [43] | Yang B, Gao X L, Zhao J M, et al. The influence of summer hypoxia on sedimentary phosphorus biogeochemistry in a coastal scallop farming area, north Yellow Sea[J]. Science of the Total Environment, 2021, 759. DOI:10.1016/j.scitotenv.2020.143486 |
| [44] | An S U, Mok J S, Kim S H, et al. A large artificial dyke greatly alters partitioning of sulfate and iron reduction and resultant phosphorus dynamics in sediments of the Yeongsan River estuary, Yellow Sea[J]. Science of the Total Environment, 2019, 665: 752-761. DOI:10.1016/j.scitotenv.2019.02.058 |
| [45] | House W A, Denison F H. Factors influencing the measurement of equilibrium phosphate concentrations in river sediments[J]. Water Research, 2000, 34(4): 1187-1200. DOI:10.1016/S0043-1354(99)00249-3 |
| [46] | Kaiserli A, Voutsa D, Samara C. Phosphorus fractionation in lake sediments-Lakes Volvi and Koronia, N. Greece[J]. Chemosphere, 2002, 46(8): 1147-1155. DOI:10.1016/S0045-6535(01)00242-9 |
| [47] | Kleeberg A, Dudel G E. Changes in extent of phosphorus release in a shallow lake (Lake Groβer Müggelsee; Germany, Berlin) due to climatic factors and load[J]. Marine Geology, 1997, 139(1-4): 61-75. DOI:10.1016/S0025-3227(96)00099-0 |
| [48] | Bastami K D, Neyestani M R, Raeisi H, et al. Bioavailability and geochemical speciation of phosphorus in surface sediments of the southern Caspian Sea[J]. Marine Pollution Bulletin, 2018, 126: 51-57. DOI:10.1016/j.marpolbul.2017.10.095 |
| [49] | Ruttenberg K C, Goñi M A. Phosphorus distribution, C∶N∶P ratios, and δ13 Coc in arctic, temperate, and tropical coastal sediments: tools for characterizing bulk sedimentary organic matter[J]. Marine Geology, 1997, 139(1-4): 123-145. DOI:10.1016/S0025-3227(96)00107-7 |
| [50] | Slomp C P, Van Cappellen P. The global marine phosphorus cycle: sensitivity to oceanic circulation[J]. Biogeosciences, 2007, 4(2): 155-171. DOI:10.5194/bg-4-155-2007 |
| [51] | Mort H P, Slomp C P, Gustafsson B G, et al. Phosphorus recycling and burial in Baltic Sea sediments with contrasting redox conditions[J]. Geochimica et Cosmochimica Acta, 2010, 74(4): 1350-1362. DOI:10.1016/j.gca.2009.11.016 |
| [52] | Slomp C P, Thomson J, de Lange G J. Controls on phosphorus regeneration and burial during formation of eastern Mediterranean sapropels[J]. Marine Geology, 2004, 203(1-2): 141-159. DOI:10.1016/S0025-3227(03)00335-9 |
| [53] | Andrieux F, Aminot A. A two-year survey of phosphorus speciation in the sediments of the Bay of Seine (France)[J]. Continental Shelf Research, 1997, 17(10): 1229-1245. DOI:10.1016/S0278-4343(97)00008-3 |
| [54] | Kang X M, Song J M, Yuan H M, et al. Phosphorus speciation and its bioavailability in sediments of the Jiaozhou Bay[J]. Estuarine, Coastal and Shelf Science, 2017, 188: 127-136. DOI:10.1016/j.ecss.2017.02.029 |
| [55] | Acharya S S, Panigrahi M K, Kurian J, et al. Speciation of phosphorus in the continental shelf sediments in the eastern Arabian Sea[J]. Continental Shelf Research, 2016, 115: 65-75. DOI:10.1016/j.csr.2016.01.005 |
| [56] | Chen J J, Lu S Y, Zhao Y K, et al. Effects of overlying water aeration on phosphorus fractions and alkaline phosphatase activity in surface sediment[J]. Journal of Environmental Sciences, 2011, 23(2): 206-211. DOI:10.1016/S1001-0742(10)60394-4 |
| [57] | Zhuang W, Gao X L, Zhang Y, et al. Geochemical characteristics of phosphorus in surface sediments of two major Chinese mariculture areas: the Laizhou Bay and the coastal waters of the Zhangzi Island[J]. Marine Pollution Bulletin, 2014, 83(1): 343-351. DOI:10.1016/j.marpolbul.2014.03.040 |
| [58] | Lentini C J, Wankel S D, Hansel C M. Enriched iron(Ⅲ)-reducing bacterial communities are shaped by carbon substrate and iron oxide mineralogy[J]. Frontiers in Microbiology, 2012, 3. DOI:10.3389/fmicb.2012.00404 |
| [59] | Sundby B. Transient state diagenesis in continental margin muds[J]. Marine Chemistry, 2006, 102(1-2): 2-12. DOI:10.1016/j.marchem.2005.09.016 |
| [60] | Slomp C P, Van der Gaast S J, Van Raaphorst W. Phosphorus binding by poorly crystalline iron oxides in North Sea sediments[J]. Marine Chemistry, 1996, 52(1): 55-73. DOI:10.1016/0304-4203(95)00078-X |
| [61] | Redfield A C, Ketchum B H, Richards F A. The influence of organisms on the chemical composition of seawater[A]. In: Hill M N, The Sea. Vol. 2[C]. New York: Interscience, 1963. |
| [62] | Noffke A, Hensen C, Sommer S, et al. Benthic iron and phosphorus fluxes across the Peruvian oxygen minimum zone[J]. Limnology and Oceanography, 2012, 57(3): 851-867. DOI:10.4319/lo.2012.57.3.0851 |
| [63] | Anschutz P, Zhong S J, Sundby B, et al. Burial efficiency of phosphorus and the geochemistry of iron in continental margin sediments[J]. Limnology and Oceanography, 1998, 43(1): 53-64. DOI:10.4319/lo.1998.43.1.0053 |
| [64] | Gunnars A, Blomqvist S. Phosphate exchange across the sediment-water interface when shifting from anoxic to oxic conditions an experimental comparison of freshwater and brackish-marine systems[J]. Biogeochemistry, 1997, 37(3): 203-226. DOI:10.1023/A:1005744610602 |
| [65] | Rozan T F, Taillefert M, Trouwborst R E, et al. Iron-sulfur-phosphorus cycling in the sediments of a shallow coastal bay: implications for sediment nutrient release and benthic macroalgal blooms[J]. Limnology and Oceanography, 2002, 47(5): 1346-1354. DOI:10.4319/lo.2002.47.5.1346 |
| [66] | Canfield D E, Thamdrup B. Towards a consistent classification scheme for geochemical environments, or, why we wish the term 'suboxic' would go away[J]. Geobiology, 2009, 7(4): 385-392. DOI:10.1111/j.1472-4669.2009.00214.x |
| [67] | Egger M, Rasigraf O, Sapart C J, et al. Iron-mediated anaerobic oxidation of methane in brackish coastal sediments[J]. Environmental Science & Technology, 2015, 49(1): 277-283. |
| [68] | Jahnke R A. Early diagenesis and recycling of biogenic debris at the seafloor, Santa Monica Basin, California[J]. Journal of Marine Research, 1990, 48(2): 413-436. |
| [69] | Reimers C E, Jahnke R A, McCorkle D C. Carbon fluxes and burial rates over the continental slope and rise off central California with implications for the global carbon cycle[J]. Global Biogeochemical Cycles, 1992, 6(2): 199-224. |
| [70] | Andrieux-Loyer F, Philippon X, Bally G, et al. Phosphorus dynamics and bioavailability in sediments of the Penzé Estuary (NW France): in relation to annual P-fluxes and occurrences of Alexandrium Minutum[J]. Biogeochemistry, 2008, 88(3): 213-231. |
| [71] | Bonaglia S, Deutsch B, Bartoli M, et al. Seasonal oxygen, nitrogen and phosphorus benthic cycling along an impacted Baltic Sea estuary: regulation and spatial patterns[J]. Biogeochemistry, 2014, 119(1-3): 139-160. |
| [72] | Sisma-Ventura G, Herut B, Silverman J, et al. P fluxes and prokaryotic cycling at benthic boundary layer in the deep southeastern Mediterranean Sea[J]. Journal of Geophysical Research: Biogeosciences, 2021, 126(4). DOI:10.1029/2020JG006110 |
| [73] | Hall P O J, Rosell E A, Bonaglia S, et al. Influence of natural oxygenation of baltic proper deep water on benthic recycling and removal of phosphorus, nitrogen, silicon and carbon[J]. Frontiers in Marine Science, 2017, 4. DOI:10.3389/fmars.2017.00027 |