 2022, Vol. 43
2022, Vol. 43



2. 厦门大学近海海洋环境科学国家重点实验室, 厦门 361102;
3. 福建省环境科学研究院, 福州 350013
2. State Key Laboratory of Marine Environmental Science, Xiamen University, Xiamen 361102, China;
3. Fujian Academy of Environmental Sciences, Fuzhou 350013, China
在全球气候变化和人类活动的叠加影响下, 水体富营养化与低氧已成为全球普遍现象, 广泛存在于水库[1]、河口[2, 3]和近海底层水体[4, 5].海岸带地区人口密集, 经济开发强度大, 低氧问题更为突出[4].“十四五”规划中明确提出建立陆海统筹的生态环境治理制度, 构建流域-河口-近海污染防治联动机制, 以促进海洋与海岸带可持续发展[6].溶解氧(dissolved oxygen, DO)是衡量水环境质量与生态系统健康的重要指标[7], 是水环境基本监测指标[8].当DO降低, 会引起水生生物生长代谢速率降低[9]、器官损伤和水生生物死亡[10, 11].此外, DO影响水体生物地球化学过程, 调控着水体氮、磷、碳和重金属等元素的转化方向与速率[12].低氧、蓝藻水华和全球暖化之间存在耦合反馈关系[13].因此, 探明流域-近海DO时空格局与低氧机制, 是实施陆海统筹战略和推进流域-海域综合治理的科学基础.
水中DO的变化趋势取决于其补充和消耗两大过程的强弱关系.DO的补充过程主要包括大气复氧和浮游植物光合作用产氧[14], DO的消耗过程主要包括有机物降解、水生生物的呼吸作用和还原性无机物的氧化等[15, 16].流域-近海系统本身具有水文连通性, 但受到筑坝拦截作用和湖库化等影响, DO变化的调控机制更为复杂.已有研究表明, 河流污染输入是影响河口和近海水环境的重要因素[17].人为营养盐氮和磷大量输入引发藻类异常增殖, 形成大量有机质的累积, 即富营养化过程[18], 而有机质的微生物分解耗氧是造成水体低氧的重要机制[19].筑坝拦截作用形成的水库和受潮汐影响较大的河口区域有着更复杂的水动力条件.夏季层化和水力停留时间长的水体(如湖泊、水库和河口)更容易出现低氧[20].除了水柱耗氧, 沉积物-水界面的有机质分解也是造成水体低氧的重要原因[21], 沉积物中还原性有机物和金属通过扰动作用进入水体也会发生耗氧过程[22].目前, 各种类型水体的低氧过程和机制已有不少研究, 但从流域-近海连续体的地学环境视角研究DO分布和低氧的调控机制未见报道.
福建省地处我国东南沿海, 河流众多, 水系发达, 梯级电站开发造成河流湖库化与富营养化, 全省约有3千多座水库[23].“十三五”以来, 各级政府加强污染治理, 全省水环境质量总体良好, 但环境形势不容乐观, 河流的主要超标因子是氨氮和总磷, 河口与近岸海域的主要超标因子是无机氮和活性磷酸盐, 部分河段和河口区的DO超标问题较为突出.长期以来, 闽江和九龙江的营养盐浓度较高[24, 25], 福建省多数水库呈现富营养化状态[26], 但有关水体DO的研究很少, 曾观测到闽江水口电站水库的低氧持续4个月之久[1].目前缺乏对全省流域-近海DO的系统性研究.本文基于2011~2020年长期监测数据, 结合气象水文和社会经济资料, 系统分析DO的时空演变规律, 探究不同水体低氧的主要影响因素和宏观调控机制, 以期为推进“十四五”流域-海域综合治理、美丽河湖和美丽海湾建设提供科学基础.
1 材料与方法 1.1 研究区域与站点分布福建省地处中国东南沿海, 属于亚热带海洋性季风气候, 光照充足, 雨量充沛, 年平均气温17~21℃, 平均降雨量1 400~2 000 mm.河流众多, 水系发达, 共有29个水系和600多条河流, 总长度约13 000 km, 水资源丰富, 河网密度达112 m ·km-2.全省较大的河流有闽江、九龙江、汀江、晋江、龙江、敖江、木兰溪和交溪等.本研究选取2011~2020年连续监测的全省135个地表水监测点位(包含64个河流点位、60个水库点位和11个河口点位)和66个近岸海域监测点位的数据进行综合分析, 以上点位覆盖全省主要河流、水库、河口和近岸海域, 具有代表性.
1.2 数据来源与监测方法水质数据来源于福建省地表水环境监测数据(包括DO、氨氮、总氮、总磷、高锰酸盐指数、化学需氧量、生化需氧量、pH和水温)和福建省近岸海域监测数据(包括DO、氨氮、化学需氧量、pH、水温和盐度).地表水监测一年6或12次(隔月或逐月), 近岸海域监测一年3次(丰、平和枯).样品的采集、固定、分析和数据处理按照《地表水和污水监测技术规范》(HJ/T 91-2002)和《近岸海域环境监测技术规范》(HJ 442-2008)执行.降水量数据来自中国气象数据网(http://data.cma.cn/).河流径流量数据来自福建省水文监测资料, 并根据汇水面积比值推算到入海断面.河口潮差数据来自海事服务网(https://www.cnss.com.cn/).国内生产总值(GDP)数据来自统计年鉴(https://tjj.fujian.gov.cn/).
1.3 数据分析方法 1.3.1 数据质控与溶解氧饱和度计算对原始的监测数据进行异常值筛选和质控, 低于检测限的数据以检测限的一半赋值统计.因水温有季节变化, 地表水与海水盐度有显著差异, 根据温、盐和大气压将现场测定DO(mg ·L-1)转化为溶解氧饱和度(%), 以提高数据可比性.溶解氧饱和度的计算过程见式(1)~(4).在标准大气压和100% 相对湿度下, 氧在水中的溶解度C0(mmol ·kg-1)由式(1)计算[27].实际大气压下的氧溶解度C(mmol ·kg-1)由式(2)计算.根据玻尔兹曼气体分子在重力场中按高度的分布规律, 不同站位的大气压通过和海拔高度H(基准为海平面)的对应关系进行计算, 具体见式(3).
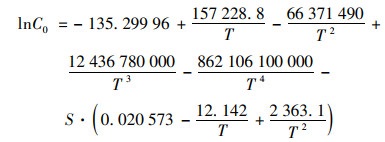
|
(1) |
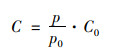
|
(2) |
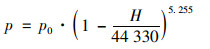
|
(3) |
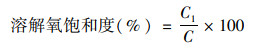
|
(4) |
式中, T为水温(℃), S为盐度(‰), H为海拔高度(基准为海平面), p为实际大气压(kPa), p0为海平面大气压(101 kPa), C1为实际大气压下的氧溶解度(mmol ·kg-1), C为水气平衡氧溶解度(mmol ·kg-1).
1.3.2 低氧检出频率定义目前对于低氧的定义没有通用标准, 国际上将低氧定义为DO < 2 mg ·L-1[4].有研究表明, 不同生物维持其正常生命活动所需的DO不同, 而鱼类生长代谢的DO阈值一般较高(通常为6.0 mg ·L-1)[7].当溶解氧饱和度低于70%时鱼类代谢降低[9], 或DO低于7.0 mg ·L-1时鱼类生长受到影响[28].我国地表水和海水水质标准对DO的限值不同, 地表水I~Ⅱ类水质标准(GB 3838-2002)规定DO≥6.0 mg ·L-1, 而海水一、二类水质标准(GB 3097-1997)规定DO>5.0 mg ·L-1.
为全面分析全省流域-近海DO的分布特征和低氧机制, 将2011~2020年地表水和海水溶解氧饱和度数据排序, 统一将10%分位数作为低氧限值, 对应饱和度67.0%, 换算为标准条件(25℃, 101 kPa)的地表水DO约为5.5 mg ·L-1, 海水(25℃, 101 kPa, 盐度30‰)约为4.6 mg ·L-1.采用本文定义的低氧限值进行分析, 基本可以反映河流、水库、河口和近海这4种类型水体的水质状况.将各种类型水体溶解氧饱和度低于67%的数据样本量占其总样本量的比例定义为低氧检出频率.利用该指标可以直观展示福建省流域-近海DO的时空格局.
1.3.3 随机森林模型分析为探明流域-近海低氧的主要影响因素, 采用随机森林(random forest, RF)模型量化分析理化参数和水文条件等因素对水体DO变化的贡献率.相较于常规的统计方法, RF模型适用于分析非线性和非正态分布数据, 在处理多重共线性方面具有稳健性[29].使用python 3.7软件“sklearn”库, 针对不同类型水体构建年和季节尺度的RF模型.RF模型输入变量包括化学参数(pH、氨氮、总氮、总磷、生化需氧量、化学需氧量和高锰酸盐指数)、物理参数(水温、降水量、潮差和径流量), 响应变量为溶解氧饱和度.所有输入变量经归一化处理, 设定决策树个数参数(n_estimators)为100.
1.3.4 数据处理与统计数据分析流程见图 1.①利用SPSS 19.0和Excel 2019计算各监测点位溶解氧饱和度和低氧检出频率, 并利用ArcGIS作图得到2011~2015年和2016~2020年福建省溶解氧饱和度均值和低氧检出频率空间分布.②基于高清遥感影像工作底图目视解译, 将所有监测点位划分为河流、水库、河口和近海这4种类型水体, 利用OriginPro 2021b分析溶解氧饱和度、低氧检出频率和其他水质参数的时空差异和演变规律.③利用SPSS 19.0进行Spearman相关分析, 再针对相关性强的参数, 基于RF模型重点分析河流、水库和河口这3种类型水体DO变化的主要贡献因子, 结合已有的环境生物地球化学过程知识, 研究福建省流域-近海低氧的主要调控机制.

|
图 1 数据分析流程示意 Fig. 1 Flowchart of data analysis |
将2011~2020年划分为“十二五”期间(2011~2015年)和“十三五”期间(2016~2020年), 对比两个时期的溶解氧饱和度均值、低氧检出频率及其变化方向和幅度(图 2).

|
图 2 2011~2015年和2016~2020年福建省流域-近海溶解氧饱和度和低氧检出频率空间分布 Fig. 2 Spatial distribution of dissolved oxygen saturation and detection frequency of hypoxia in 2011-2015 and 2016-2020 of the watershed-coastal system in Fujian province |
福建省流域-近海2011~2015年溶解氧饱和度均值为52.1% ~107.0%[图 2(a)], 2016~2020年溶解氧饱和度均值为55.3% ~111.7%[图 2(b)]. 和2011~2015年相比, 2016~2020年溶解氧饱和度整体有所增加, 但流域和河口部分点位有所下降[图 2(c)]. 2011~2015年低氧检出频率大于0的流域点位90个(共124个点位), 河口点位7个(共11个点位), 近海监测点位3个(共66个点位)[图 2(d)]. 2016~2020年低氧检出频率大于0的流域点位66个, 河口点位10个, 近海监测点位0个[图 2(e)]. 2011~2015年低氧检出频率较高(>10%)的监测点位主要分布在汀江水库和沙溪梯级电站2个库区, 闽江干流、九龙江北溪上游(漳平)和西溪下游城市(漳州)河段, 以及龙江、木兰溪、晋江和九龙江4个河口. 2016~2020年河流和水库的低氧检出频率整体有所下降, 但闽江、九龙江和晋江流域有个别站位上升较多; 河口区域变化不大, 其中晋江口、漳江口和诏安湾的低氧检出频率有所增加[图 2(f)].
2.2 不同类型水体DO和其他水质因子差异按河流、水库、河口和近海这4种类型水体进行统计, 溶解氧饱和度和其他水质指标(高锰酸盐指数、化学需氧量、氨氮、总氮和总磷)存在组间差异(图 3). 2011~2020年福建省河流、水库、河口和近海溶解氧饱和度分别为(86.1±14.3)%、(83.5±14.3)%、(79.2±17.9)%和(98.2±10.2)%.其中, 河口溶解氧饱和度最低, 同时化学需氧量、高锰酸盐指数、氨氮、总氮和总磷浓度最高, 与河流、水库和近海存在显著性差异(P < 0.05).福建省河口各项指标的平均浓度最高, 不同河口间的水质差异较大.河口化学需氧量为(14.7±8.8) mg ·L-1, 氨氮浓度为(0.58±0.53) mg ·L-1, 总氮浓度为(2.41±1.70) mg ·L-1, 总磷浓度为(0.154±0.101) mg ·L-1, 高锰酸盐指数为(3.5±1.8) mg ·L-1.从河流、水库到河口这一连续体呈现DO降低的趋势, 水库的水质优于河流, 但平均溶解氧饱和度低于河流[图 3(a)].近海的溶解氧饱和度最高, 化学需氧量和氨氮浓度最低.

|
误差棒为标准偏差, 不同小写字母表示组间差异具有统计学意义(P < 0.05), 近海没有监测总氮、总磷和高锰酸盐指数 图 3 2011~2020年4种类型水体水质参数比较 Fig. 3 Comparison of water quality parameters of four types of water bodies from 2011 to 2020 |
对比两个时期河流、水库和河口这3种类型水体(近海监测指标和频率不同, 且基本未检出低氧, 在此未作比较)水质指标(高锰酸盐指数、化学需氧量、氨氮、总氮和总磷)的变化, 发现2016~2020年河流和水库各水质指标浓度均明显降低, 但是河口氨氮和总磷浓度变化不大, 总氮浓度略有增加(表 1).
|
|
表 1 3种类型水体水质参数浓度两个时期比较(2011~2020年)1)/mg ·L-1 Table 1 Comparison of water quality concentration of three types of water bodies in two periods from 2011 to 2020/mg ·L-1 |
2.3 不同类型水体DO年际变化和季节差异
为探究福建省流域-近海系统DO时间变化规律, 从年际和季节两个时间尺度对河流、水库、河口和近海这4种类型水体的DO进行分析.图 4显示, 与2011~2015年相比, 2016~2020年这4种类型水体的溶解氧饱和度均有所增加, 河流、水库和近海溶解氧饱和度在两个时期间存在显著差异[P < 0.05, 图 4(a)]; 低氧检出频率均有所下降, 其中近海低氧检出频率降低至0%, 且只有河流和水库存在显著差异[P < 0.05, 图 4(b)].对于河口来说, 与2011~2015年相比, 2016~2020年的溶解氧饱和度均值增加, 低氧检出频率均值降低, 但两个时期无显著差异.

|
对4种类型水体进行两个时期的组间平均值比较, 不同小写字母表示组间差异具有统计学意义(P < 0.05); 箱体上下限为上下四分位数, 外界为1.5IQR内的范围, 箱体中线为中位线, 点为均值, 均值连线表示两个时期变化趋势 图 4 溶解氧饱和度和低氧检出频率两个时期比较 Fig. 4 Comparison of dissolved oxygen saturation and detection frequency of hypoxia between two periods |
近海没有完整的4个季节监测(一年三期), 也基本未观测到低氧问题, 不进行季节变化分析.河流、水库和河口的溶解氧饱和度和低氧检出频率存在一定的季节差异(图 5).3种类型水体的溶解氧饱和度均是夏季高, 河流和水库冬季低, 河口春季低[图 5(a)].低氧检出频率均是夏季低, 河流春冬季高, 水库秋季高, 河口春季高[图 5(c)].如果筛选出低氧检出频率大于0%的监测点位数据进行统计, DO的季节变化规律有所不同, 此时河口溶解氧饱和度呈现夏季低, 春冬季高的趋势[图 5(b)], 河口低氧检出频率为夏季最高, 河流和水库低氧检出频率在秋季最高[P < 0.05, 图 5(d)].

|
(a)和(c)为2011~2020年所有数据, (b)和(d)为筛选出低氧检出频率大于0%的数据; 对每种类型水体进行季节的组间比较, 不同小写字母表示组间差异具有统计学意义(P < 0.05); 箱体上下限为上下四分位数, 外界为1.5IQR内的范围, 箱体中线为中位线, 点为均值, 均值连线表示季节变化趋势 图 5 溶解氧饱和度和低氧检出频率的季节差异 Fig. 5 Seasonal variation in dissolved oxygen saturation and detection frequency of hypoxia |
溶解氧饱和度和其他参数的相关分析结果见表 2.河流、水库和河口的溶解氧饱和度与化学需氧量、高锰酸盐指数、氨氮、总氮和总磷均呈显著负相关.河流和水库的溶解氧饱和度与降水量、水温和pH正相关, 但河口的溶解氧饱和度与降水量和水温没有相关性.3种类型水体的溶解氧饱和度与GDP负相关, 其中河口的相关系数最大.
|
|
表 2 溶解氧饱和度和环境经济参数的Spearman相关性 Table 2 Spearman correlation between dissolved oxygen saturation and environmental and economic parameters |
选取直接影响DO且相关性较强的9个参数进行RF模型分析.图 6显示, 2016~2020年, 氨氮和总氮对河流DO的贡献率较高, 降水量和pH对水库DO的贡献率较高, 总磷和总氮对河口DO的贡献率较高.和2011~2015年相比, 2016~2020年氨氮对河流DO的贡献率从10.3%增加为27.1%[图 6(a)-1和6(a)-2], pH对水库DO的贡献率由11.9%增加为20.8%[图 6(b)-1和6(b)-2], 总氮对河口DO的贡献率由10.9%增加为16.1%[图 6(c)-1和6(c)-2].对比所有监测点位和低氧频率大于10%的点位的分析结果, 贡献率差异不大.

|
(a)表示河流, (b)表示水库, (c)表示河口; 1和2表示低氧检出频率大于10%的点位(N=56), 3和4表示所有点位(N=135); 1和3表示2011~2015年, 2和4表示2016~2020年; 数值对应贡献率(%)中最高的两个参数 图 6 河流、水库和河口2011~2015年和2016~2020年随机森林模型贡献率 Fig. 6 Contribution of random forest models for rivers, reservoirs, and estuaries between 2011-2015 and 2016-2020 |
基于低氧检出频率大于10%的点位数据进行RF模型分析, DO的贡献率呈现季节差异(图 7).对河流DO贡献率较大的因子, 春夏季是氨氮, 秋季是氨氮和总氮, 冬季是氨氮和高锰酸盐指数[图 7(a)].降水量对水库DO的贡献率春冬季低, 夏秋高, pH的贡献率在春夏季较高[图 7(b)].对河口DO贡献率较大的因子, 春季是总氮和总磷, 夏秋季是总磷[图 7(c)].

|
(a)表示河流, (b)表示水库, (c)表示河口; 1、2、3和4分别表示春季、夏季、秋季和冬季; 数值对应贡献率(%)中最高的两个参数 图 7 河流、水库和河口不同季节随机森林模型贡献率(2011~2020年) Fig. 7 Contribution of random forest models for rivers, reservoirs and estuaries in different seasons from 2011 to 2020 |
选择低氧检出频率大于10%的4个河口(晋江口、九龙江口、木兰溪口和龙江口)进行RF模型分析.生化需氧量、总磷和总氮对河口DO贡献率最大, 贡献率分别为16.3%、13.8%和13.2%.降水量、潮差和径流量对河口DO也有一定的贡献, 贡献率分别为9.5%、8.0%和7.7%.河口的溶解氧饱和度与总氮、总磷和生化需量氧之间存在显著的负相关关系(P < 0.05, 图 8).总体上, 这些河口低氧检出频率大于0%时, 对应的氮磷浓度较高.

|
图 8 典型河口溶解氧饱和度与总氮、总磷和生化需氧量的关系 Fig. 8 Relationship between TN, TP, BOD, and dissolved oxygen saturation in typical estuaries |
流域-近海溶解氧饱和度和低氧检出频率的空间差异主要与污染排放有关.从河流、水库、河口到近海, 溶解氧饱和度先降低后增加, 而化学需氧量和营养盐浓度先增加后降低(图 3).相关分析表明溶解氧饱和度和氨氮、总氮和总磷等水质参数均呈负相关关系(表 2), 这和珠江口等研究结果类似[15, 30].表明不同类型水体污染输入差异是影响流域-近海DO的关键因素.大量生活污水、农业废水和工业废水排入河流, 特别是城市河段的低氧检出频率会升高.例如, 九龙江北溪上游的漳平市河段和西溪下游的漳州市河段的低氧检出频率均较高(图 2), 这些河段的营养盐浓度也较高.本课题组研究发现, 九龙江高氨氮污水排入河流以后, 氨氧化细菌(AOB)增加, 促进硝化作用和耗氧[31].汤云等[24]研究报道, 闽江流域沙溪下游段水质较差, 主要与三明市工业污染与生活污染较重有关.受水文联通作用, 流域污染物通过河流汇集到河口, 容易产生低氧现象[32].位于东南与南海之间的福建近海(台湾海峡), 冬春季受浙闽沿岸流主导, 夏季受南海暖流主导, 因狭管效应水动力扩散条件好, 大气复氧快, 污染物稀释作用强, 耗氧量少[33], 因此溶解氧饱和度最高, 低氧检出频率最低(图 4).在近海点位中, 三沙湾是一个例外, “十二五”期间溶解氧饱和度均值为85%, 小于三沙湾两条入海河流(88%)与河口(90%).三沙湾属于封闭型海湾, 水动力条件较差, 加上城市生活和海水养殖等污染较重, DO较低[34].
流域-近海溶解氧饱和度和低氧检出频率的季节变化受气象水文条件和生化过程的综合影响.河流和水库DO的季节变化规律相似, 对于有低氧检出的点位进行统计分析, 河流和水库均为秋季低氧检出频率最高[图 5(d)], 夏秋季氨氮和总氮对河流DO的贡献率较高, 其中秋季最高[图 7(a)].福建省位于亚热带季风气候区, 春末夏初进入汛期, 在降水冲刷作用下, 流域地表污染物大量入河, 流域-近海系统水体的耗氧物质增加, 同时夏季水温升高促进有机质分解, 秋季汛期结束, 流量开始下降, 水力停留时间延长, 水中营养盐和有机质呈现富集效应, 持续耗氧, 造成秋季低氧检出频率在全年中最高.河口低氧的季节性特征与流域不同, 主要与水动力条件的季节差异有关.河口的低氧检出频率夏季最高, 从春季到夏秋季, 总磷对DO的贡献率增加(图 7).汛期流域污染物通过河流输入河口, 加上水温升高可促进微生物耗氧过程, 导致河口夏秋的低氧检出频率明显高于其他季节[图 5(d)].
3.2 水库低氧机制水库的溶解氧饱和度和低氧检出频率与上游污染输入、库区富营养化和层化引起的底层生化耗氧过程加剧有关.降水量和pH对水库DO的贡献率较大[图 6(b)], 春夏季pH对DO的贡献率增加[图 7(b)], 水库溶解氧饱和度和pH正相关(表 2), 库区pH升高主要是由于CO2降低[35, 36], 这是藻类光合作用增强的结果[1].水库春夏季表层水光合作用较强, 低氧检出频率较低[图 5(d)].夏季高温水体分层明显, 底层水无法进行水气交换, 表层水强光合作用产生生源有机质, 随后沉降至底层水, 加上河流携带的陆源有机质的大量输入, 在水中开始耗氧分解; 秋季开始降温, 垂向混合加强, 低氧的底层水和表层水混合, 同时通过坝下泄流[1], 造成秋季梯级电站水库坝下河段表层水低氧检出频率整体增加[图 5(d)].
针对水库低氧点位的数据专门分析, 发现低氧检出频率较高的点位主要分布在汀江水库和沙溪梯级电站河段.水库低氧受到水库库容、水力停留时间和入库河流水质等因素影响[37].筑坝以后, 水库水力停留时间延长, 流速变缓, 径流携带的陆源颗粒物进入库区以后容易沉降, 从河流区、过渡区到湖泊区透明度增加, 促进富营养化过程(通过初级生产向水体供应有机质)[38].福建省的水系属于山区性河流, 大多数水库水深大于20 m, 高温季节易出现热力学分层现象, 垂向混合弱, 底层水的大气供氧不足, 耗氧过程强烈, 最终造成水库下层水体的DO持续下降.与其他水库不同, 汀江水库的监测点位处于水库出口, 监测的是下泄的底层水, 溶解氧饱和度较低.闽江沙溪段分布有多座大坝, 属于梯级电站水库河段, 水库底层低氧水的下泄造成整个河段低氧检出频率较高的点位较多(图 2).沙溪流经三明市和沙县, 沿河城镇有污水输入, 水质较差[24], 加上水库的拦截与层化效应, 是低氧的主要成因.本课题组基于长期观测和培养实验研究, 发现闽江水口水库(水深60 m)的季节性低氧与层化和有机质耗氧的水文-生地化耦合作用有关[1].
3.3 河口低氧机制河口的低氧与污染输入、潮水顶托和还原性有机物耗氧过程有关.河口位于海陆界面, 受径流和潮汐的叠加影响, 大量陆源颗粒物(含有机物与矿物颗粒)通过河流或沿岸排污口进入河口区域.受潮水顶托作用, 污染物滞留在河口(含感潮段)和海湾内, 难以向外海排出.表观上, 河口溶解氧饱和度与总磷、总氮和生化需氧量负相关, 降水量、潮差和径流量等水动力参数对河口溶解氧饱和度也有一定的贡献.与河流不同, “十三五”期间河口的溶解氧饱和度与“十二五”相比变化不大, 部分河口的低氧检出频率不降反升[图 2(f)]; 河口低氧检出频率最高的是夏季, 而河流是秋季, 这种差异暗示河口地区的低氧问题和本地污染输入关系可能更为密切.有研究表明, 全球海水养殖规模近20年大幅扩张[39], 福建是海水养殖大省, 河口湾养殖密度逐年增大[40].通过养殖尾水排放, 大量的氮、磷和有机物进入河口区[41].此外, 福清市(龙江口)、莆田市荔城区(木兰溪口)和泉州市丰泽区(晋江口)2020年常住人口数相较于2015年分别增加4.8%、14.0%和12.2%, 高于全省平均增加幅度(4.4%).因此, 海岸带地区人口增加, 生活污水输入也可能是河口低氧检出频率居高不下的原因之一.
除了流域污染输入, 在涨退潮、洪水急流或风力扰动作用下, 河口厌氧沉积物的再悬浮和颗粒物相关的耗氧过程, 也是河口低氧的成因.有研究认为, 河口颗粒有机物的耗氧可能起到主要作用[21].总磷对河口溶解氧饱和度的贡献率最大[图 6(c)], 河口夏季的低氧检出频率最高[图 5(d)], 对应总磷浓度最高(总氮、氨氮和生化需氧量是春冬季最高).磷的转化与DO没有直接关系, 但磷属于颗粒活性物质, 易与金属颗粒结合, 暗示水中可能含有未监测的还原性物质(如还原态的金属颗粒)参与了耗氧过程.研究发现沉积物中羟基氧化铁可以吸附磷酸盐, 在沉积物处于缺氧时逐渐转化成硫化亚铁并使磷酸盐解吸, 在水体扰动作用下颗粒物中的磷溶解在水中, 水体磷酸盐与二价铁Fe(Ⅱ)浓度均增加, Fe(Ⅱ)会消耗氧导致水中DO降低[42].受长江冲淡水控制的表层海水具有较高浓度的可溶态二价铁Fe(Ⅱ), 长江口(盐度 < 30‰)Fe(Ⅱ)在Fe(Ⅱ+Ⅲ)中所占的比例高达50%以上[43].此外, 福建省的铁矿石丰富[44], 河口区的低氧可能和矿物金属耗氧过程有关, 未来需要进一步开展深入研究.
4 结论(1) 福建省流域-近海多年平均的溶解氧饱和度近海>河流>水库>河口, 近海最高[(98.2±10.2)%], 河口最低[(79.2±17.9)%].与“十二五”(2011~2015年)相比, “十三五”(2016~2020年)河流和水库的低氧检出频率有明显降低, 但河口变化不大.统计有低氧检出的点位, 河流和水库的多年平均低氧检出频率在秋季最高, 河口在夏季最高.
(2) 流域-近海DO的空间差异与水质和水动力条件有关, 不同水体的污染输入差异是影响流域-近海DO的关键因素.流域污染物通过河流汇集到水库和河口区域, 导致水库和河口的低氧检出频率高于河流与近海.流域-近海DO的季节变化主要受气象水文条件和生化过程的综合影响, 温度的季节差异主要影响藻类光合作用、微生物降解过程和水体分层, 降水的季节差异主要影响外源污染的冲刷输入.
(3) 水库和河口低氧问题最为突出但调控机制不同, 水库低氧检出频率在夏秋季节较高, 与上游污染输入、库区富营养化和层化引起的底层生化耗氧过程加剧有关, 河口的低氧与污染输入(特别是本地源)、潮水顶托和还原性物质耗氧过程有关.亟需建立系统治理与分区管控制度, 加大流域和河口海域污染的治理力度, 以减缓水体富营养化和低氧问题.
| [1] | Yan J, Chen N W, Wang F F, et al. Interaction between oxygen consumption and carbon dioxide emission in a subtropical hypoxic reservoir, Southeastern China[J]. Journal of Geophysical Research: Biogeosciences, 2021, 126(4). DOI:10.1029/2020JG006133 |
| [2] | Zhao Y Y, Uthaipan K, Lu Z M, et al. Destruction and reinstatement of coastal hypoxia in the South China Sea off the Pearl River estuary[J]. Biogeosciences, 2021, 18(8): 2755-2775. DOI:10.5194/bg-18-2755-2021 |
| [3] | Zhang Z R, Wang B, Li S Y, et al. On the intra-annual variation of dissolved oxygen dynamics and hypoxia development in the Pearl River Estuary[J]. Estuaries and Coasts, 2022, 45(5): 1305-1323. DOI:10.1007/s12237-021-01022-0 |
| [4] | Breitburg D, Levin L A, Oschlies A, et al. Declining oxygen in the global ocean and coastal waters[J]. Science, 2018, 359(6371). DOI:10.1126/science.aam7240 |
| [5] | Kemp W M, Boynton W R, Adolf J E, et al. Eutrophication of chesapeake bay: historical trends and ecological interactions[J]. Marine Ecology Progress Series, 2005, 303: 1-29. DOI:10.3354/meps303001 |
| [6] |
宋爽, 郭丽峰, 张辉, 等. 流域-河口-近岸海域污染防治机制研究[J]. 海洋环境科学, 2021, 40(6): 838-842. Song S, Guo L F, Zhang H, et al. Research on pollution control mechanism of watershed, estuary and coastal sea areas[J]. Marine Environmental Science, 2021, 40(6): 838-842. DOI:10.13634/j.cnki.mes.2021.06.005 |
| [7] | Gray J S, Wu R S S, Or Y Y. Effects of hypoxia and organic enrichment on the coastal marine environment[J]. Marine Ecology Progress Series, 2002, 238: 249-279. DOI:10.3354/meps238249 |
| [8] | Tanaka Y, Ishan I, Jumat A, et al. Multivariate analysis of the surface water in tropical river-estuary-sea continuums with changing land use[J]. Environmental Monitoring and Assessment, 2022, 194(5). DOI:10.1007/S10661-022-10006-X |
| [9] | Hammer C. Fatigue and exercise tests with fish[J]. Comparative Biochemistry and Physiology Part A: Physiology, 1995, 112(1): 1-20. DOI:10.1016/0300-9629(95)00060-K |
| [10] | LaBone E D, Rose K A, Justic D, et al. Effects of spatial variability on the exposure of fish to hypoxia: a modeling analysis for the Gulf of Mexico[J]. Biogeosciences, 2021, 18(2): 487-507. DOI:10.5194/bg-18-487-2021 |
| [11] | Ferreira A M, Marques J C, Seixas S. Integrating marine ecosystem conservation and ecosystems services economic valuation: implications for coastal zones governance[J]. Ecological Indicators, 2017, 77: 114-122. DOI:10.1016/j.ecolind.2017.01.036 |
| [12] | Middelburg J J, Levin L A. Coastal hypoxia and sediment biogeochemistry[J]. Biogeosciences, 2009, 6(7): 1273-1293. DOI:10.5194/bg-6-1273-2009 |
| [13] | Smucker N J, Beaulieu J J, Nietch C T, et al. Increasingly severe cyanobacterial blooms and deep water hypoxia coincide with warming water temperatures in reservoirs[J]. Global Change Biology, 2021, 27(11): 2507-2519. DOI:10.1111/gcb.15618 |
| [14] | D'Asaro E, McNeil C. Air-sea gas exchange at extreme wind speeds measured by autonomous oceanographic floats[J]. Journal of Marine Systems, 2007, 66(1-4): 92-109. DOI:10.1016/j.jmarsys.2006.06.007 |
| [15] | He B Y, Dai M H, Zhai W D, et al. Hypoxia in the upper reaches of the Pearl River Estuary and its maintenance mechanisms: a synthesis based on multiple year observations during 2000-2008[J]. Marine Chemistry, 2014, 167: 13-24. DOI:10.1016/j.marchem.2014.07.003 |
| [16] | Oschlies A, Brandt P, Stramma L, et al. Drivers and mechanisms of ocean deoxygenation[J]. Nature Geoscience, 2018, 11(7): 467-473. DOI:10.1038/s41561-018-0152-2 |
| [17] | Justić D, Kourafalou V, Mariotti G, et al. Transport processes in the Gulf of Mexico along the river-estuary-shelf-ocean continuum: a review of research from the Gulf of Mexico research initiative[J]. Estuaries and Coasts, 2022, 45(3): 621-657. DOI:10.1007/s12237-021-01005-1 |
| [18] | Nixon S W. Coastal marine eutrophication: a definition, social causes, and future concerns[J]. Ophelia, 1995, 41(1): 199-219. DOI:10.1080/00785236.1995.10422044 |
| [19] | Qian W, Gan J P, Liu J W, et al. Current status of emerging hypoxia in a eutrophic estuary: the lower reach of the Pearl River Estuary, China[J]. Estuarine, Coastal and Shelf Science, 2018, 205: 58-67. DOI:10.1016/j.ecss.2018.03.004 |
| [20] | Li D, Gan J P, Hui R, et al. Vortex and biogeochemical dynamics for the hypoxia formation within the coastal transition zone off the Pearl River Estuary[J]. Journal of Geophysical Research: Oceans, 2020, 125(8). DOI:10.1029/2020JC016178 |
| [21] | Liu Z F, Xue J H. The lability and source of particulate organic matter in the northern Gulf of Mexico hypoxic zone[J]. Journal of Geophysical Research: Biogeosciences, 2020, 125(9). DOI:10.1029/2020JG005653 |
| [22] | Johnson G M, Sutherland D A, Roering J J, et al. Estuarine dissolved oxygen history inferred from sedimentary trace metal and organic matter preservation[J]. Estuaries and Coasts, 2019, 42(5): 1211-1225. DOI:10.1007/s12237-019-00580-8 |
| [23] |
朱珍香, 杨军. 福建水库空间分布特征: 沿海密度高水量少、内陆密度低水量多[J]. 湖泊科学, 2018, 30(2): 567-580. Zhu Z X, Yang J. The spatial distribution of reservoirs in Fujian Province: higher density but lower storage in coastal than inland regions[J]. Journal of Lake Sciences, 2018, 30(2): 567-580. |
| [24] |
汤云, 卢毅敏, 吴升. 闽江流域水质时空分布特征及污染源解析[J]. 长江科学院院报, 2019, 36(8): 30-35, 48. Tang Y, Lu Y M, Wu S. Spatio-temporal distribution and source identification of water pollutants in Minjiang River Basin[J]. Journal of Yangtze River Scientific Research Institute, 2019, 36(8): 30-35, 48. |
| [25] |
陈能汪. 全球变化下九龙江河流-河口系统营养盐循环过程、通量与效应[J]. 海洋地质与第四纪地质, 2018, 38(1): 23-31. Chen N W. Nutrient cycling processes, fluxes and effects in the Jiulong river-estuary system under global change[J]. Marine Geology & Quaternary Geology, 2018, 38(1): 23-31. |
| [26] | Yang J, Yu X Q, Liu L M, et al. Algae community and trophic state of subtropical reservoirs in southeast Fujian, China[J]. Environmental Science and Pollution Research, 2012, 19(5): 1432-1442. DOI:10.1007/s11356-011-0683-1 |
| [27] | Benson B B, Krause Jr D. The concentration and isotopic fractionation of oxygen dissolved in freshwater and seawater in equilibrium with the atmosphere[J]. Limnology and Oceanography, 1984, 29(3): 620-632. DOI:10.4319/lo.1984.29.3.0620 |
| [28] | Pedersen C L. Energy budgets for juvenile rainbow trout at various oxygen concentrations[J]. Aquaculture, 1987, 62(3-4): 289-298. DOI:10.1016/0044-8486(87)90171-2 |
| [29] | Strobl C, Malley J, Tutz G. An introduction to recursive partitioning: rationale, application, and characteristics of classification and regression trees, bagging, and random forests[J]. Psychological Methods, 2009, 14(4): 323-348. DOI:10.1037/a0016973 |
| [30] | Kemp W M, Testa J M, Conley D J, et al. Temporal responses of coastal hypoxia to nutrient loading and physical controls[J]. Biogeosciences, 2009, 6(12): 2985-3008. DOI:10.5194/bg-6-2985-2009 |
| [31] | Lin J J, Chen N W, Yuan X, et al. Impacts of human disturbance on the biogeochemical nitrogen cycle in a subtropical river system revealed by nitrifier and denitrifier genes[J]. Science of the Total Environment, 2020, 746. DOI:10.1016/j.scitotenv.2020.141139 |
| [32] | Castro A L C, Eschrique S A, Silveira P C A, et al. Physicochemical properties and distribution of nutrients on the inner continental shelf adjacent to the Gulf of Maranhao (Brazil) in the equatorial Atlantic[J]. Applied Ecology and Environmental Research, 2018, 16(4): 4829-4847. DOI:10.15666/aeer/1604_48294847 |
| [33] |
吴林妮, 韦钦胜, 冉祥滨, 等. 南黄海-长江口海域溶解氧分布和低氧、酸化特征及其控制因素[J]. 中国环境科学, 2021, 41(3): 1311-1324. Wu L N, Wei Q S, Ran X B, et al. Dissolved oxygen distributions, hypoxic and acidification characteristics, and their controlling factors in the southern Yellow Sea and off the Changjiang Estuary[J]. China Environmental Science, 2021, 41(3): 1311-1324. DOI:10.3969/j.issn.1000-6923.2021.03.035 |
| [34] |
王颢, 魏姗姗, 阳藻, 等. 福建三沙湾夏季海水溶解氧分布及低氧现象初探[J]. 海洋湖沼通报, 2014(3): 167-174. Wang H, Wei S S, Yang Z, et al. Preliminary studies on DO distributions and hypoxia of Sansha Bay of Fujian in summer[J]. Transactions of Oceanology and Limnology, 2014(3): 167-174. |
| [35] | Wallace R B, Baumann H, Grear J S, et al. Coastal ocean acidification: the other eutrophication problem[J]. Estuarine, Coastal and Shelf Science, 2014, 148: 1-13. DOI:10.1016/j.ecss.2014.05.027 |
| [36] | Sobek S, Tranvik L J, Cole J J. Temperature independence of carbon dioxide supersaturation in global lakes[J]. Global Biogeochemical Cycles, 2005, 19(2). DOI:10.1029/2004GB002264 |
| [37] | Dos Santos Sá A K D, Cutrim M V J, Costa D S, et al. Algal blooms and trophic state in a tropical estuary blocked by a dam (northeastern Brazil)[J]. Ocean and Coastal Research, 2021, 69. DOI:10.1590/2675-2824069.20-006akddss |
| [38] | Moss B, Kosten S, Meerhoff M, et al. Allied attack: climate change and eutrophication[J]. Inland Waters, 2011, 1(2): 101-105. DOI:10.5268/IW-1.2.359 |
| [39] | Naylor R L, Hardy R W, Buschmann A H, et al. A 20-year retrospective review of global aquaculture[J]. Nature, 2021, 591(7851): 551-563. DOI:10.1038/s41586-021-03308-6 |
| [40] | Del Valle T M, Wu J, Xu C B, et al. Spatiotemporal dynamics and resource use efficiency in mariculture production: a case study in Southeastern China[J]. Journal of Cleaner Production, 2022, 340. DOI:10.1016/j.jclepro.2022.130743 |
| [41] |
李纯厚, 黄洪辉, 林钦, 等. 海水对虾池塘养殖污染物环境负荷量的研究[J]. 农业环境科学学报, 2004, 23(3): 545-550. Li C H, Huang H H, Lin Q, et al. Environmental carrying capacity of the pollutants from the prawn seawater culture ponds[J]. Journal of Agro-Environment Science, 2004, 23(3): 545-550. DOI:10.3321/j.issn:1672-2043.2004.03.030 |
| [42] | Krom M D, Berner R A. Adsorption of phosphate in anoxic marine sediments[J]. Limnology and Oceanography, 1980, 25(5): 797-806. |
| [43] | Huang Y M, Yuan D X, Zhu Y, et al. Real-time redox speciation of iron in estuarine and coastal surface waters[J]. Environmental Science & Technology, 2015, 49(6): 3619-3627. |
| [44] |
陈心怡, 黄奇瑜, 邵磊. 福建闽江和九龙江现代沉积物重矿物特征及其物源意义[J]. 古地理学报, 2018, 20(4): 637-650. Chen X Y, Huang Q Y, Shao L. Characteristics of heavy minerals in modern sediments of Minjiang and Jiulongjiang Rivers, Fujian Province and their provenance implication[J]. Journal of Palaeogeography, 2018, 20(4): 637-650. |