 2022, Vol. 43
2022, Vol. 43



2. 浙江省农业科学院环境资源与土壤肥料研究所, 杭州 310021
2. Institute of Environment, Resource, Soil & Fertilizer, Zhejiang Academy of Agricultural Sciences, Hangzhou 310021, China
抗生素作为一种环境新兴污染物, 在水体和土壤等各种环境介质中被频繁检出[1]. 养殖业是抗生素应用的主要领域. 据统计, 兽用抗生素的销售量占抗生素总销售量的52%[2], 进入机体后约58%的抗生素无法完成代谢, 以原药形式随粪便排出体外[3]. 伴随畜禽粪便还田利用, 这些抗生素会进入农业环境, 成为农田抗生素污染的主要来源[4]. 一些难降解的抗生素进入水体和土壤环境后长期残留并富集, 不仅影响植物的出芽和生长[5], 其产生的选择压力还进一步造成周边水体和土壤中耐药细菌种群增加, 诱导抗性基因扩散传播[6]. 采用有效手段降低粪肥中抗生素残留, 从源头上解决环境抗生素污染问题是实现绿色种养循环的重要保障.
堆肥是畜禽粪便无害化和资源再利用的重要方法[7], 既实现了物质分解和养分累积, 堆肥过程中产生的高温, 还杀灭了粪肥中的病菌和寄生虫卵[8], 降低了生态威胁. 堆肥也被证实可降低畜禽有机肥中的抗生素残留[9], 减少因畜禽粪便还田引发的抗生素环境释放. 但是由于粪便类型、抗生素种类以及堆肥工艺的差异, 不同报道中堆肥对抗生素的去除效率变异较大, 可在0% ~99%之间[10]. 有研究发现, 采用一些改良措施, 如超高温堆肥[11]和添加堆肥辅料[9]均可提高堆肥效率, 促进堆体中抗生素残留的去除. 微生物降解是堆肥和土壤等复杂环境介质中抗生素去除的主要机制. 在污染生物修复领域, 当污染环境土著微生物的污染物降解能力不足或缺乏时, 可以通过外源接种具有特定功能的微生物强化污染物去除, 即微生物强化技术. 微生物强化技术在畜禽粪便抗生素去除中也有很好的应用效果. 沈东升等[12]证实在猪粪堆肥中接种土霉素降解菌, 21 d堆肥结束时土霉素降解率从62.70%提高到82.00%. 接种部分芽孢杆菌复合制剂也被证实能有效促进堆肥中抗生素降解, 例如Zhang等[13]在堆肥过程中加入一种芽孢杆菌生物复合制剂后, 土霉素和金霉素降解率明显好于不加菌剂的试验组. 除了接种细菌, 接种白腐真菌也可有效促进粪便及其堆肥中抗生素去除. Liu等[14]在利用黄孢原毛平革菌(白腐真菌)和黑曲霉为主的微生物菌剂进行猪粪堆肥时, 多西环素和恩诺沙星降解率分别提高了40.00%和28.50%.
抗生素降解菌的运用有效加速了有机肥中抗生素的去除. 但是在实际生产中, 企业添加微生物菌剂的主要目的是促进堆肥升温腐熟. 由于缺乏抗生素残留相关的标准, 企业对单纯以抗生素降解为目的的微生物菌剂需求不强. 因此, 研究抗生素降解菌剂添加对堆肥升温和腐熟等进程的影响, 研发兼具污染物削减和堆肥升温腐熟的多功能微生物菌剂具有重要实际价值, 但是目前关于抗生素降解菌对堆肥发酵中理化和生物过程的作用研究较为有限. 此外, 外源添加的微生物菌剂在畜禽粪便堆肥中面临与土著微生物菌种的竞争, 可改变堆肥原有微生物群落结构和演替. Liu等[14]在猪粪堆肥中添加黄孢原毛平革菌、黑曲霉和芽孢杆菌复合菌剂, 引起了堆体中群落结构的变化, 促进了大量抗生素降解细菌及多重耐药菌(不动杆菌和链球菌)的繁殖, 降低抗生素抗性基因(ARGs)的总相对丰度. Du等[15]指出益生菌菌剂的添加改变了土著微生物区系结构, 在微生物区系间引入更多的竞争机制, 诱导更多的相互作用.
前期研究陆续分离获得四环素类抗生素降解菌Arthrobacter sp. OTC-16[16]、磺胺类抗生素降解菌Alcaligenes sp. SMD-FA和伴侣菌Leucobacter sp. SMD-HB[17], 试验证实这些菌株对抗生素有较好的降解效果. 本研究以报道中四环素和磺胺类抗生素检出率较高的猪粪[18]为材料, 将上述菌株混合制成复合菌剂引入其中, 明确复合菌剂发挥降解效果的可行性; 并将抗生素降解菌群与蛋白质、淀粉和纤维素分解相关细菌混合, 制成多功能菌剂, 研究该菌剂对猪粪堆肥理化进程和堆肥产物的影响, 明确了多功能菌剂的实际应用效果; 同时, 利用扩增子测序技术进一步解析了外源抗生素菌剂与土著细菌群落之间的交互效应. 本研究结果丰富了多功能菌剂应用于堆肥的理论体系, 以期为抗生素降解菌剂的实际开发利用提供初步理论依据.
1 材料与方法 1.1 试验材料与菌种新鲜的猪粪采自浙江省农业科学院杨渡基地养殖场, 木屑、麸皮和生物炭、黄瓜、萝卜种子采自杭州市当地市场. 猪粪的理化性质如表 1所示. 供试微生物菌种如表 2所示, 均为本实验室保存. 本试验用抗生素购自Sigma公司, 纯度>99.00%.
|
|
表 1 原料理化性质 Table 1 Physical and chemical properties of raw materials |
|
|
表 2 复合微生物菌剂中包含的微生物菌株 Table 2 Microbial strains contained in the compound microbial inoculum |
1.2 抗生素降解菌剂的制备
将LB培养基新鲜培养的烟草节杆菌OTC-16、产碱杆菌SMD-FA和亮杆菌SMD-HB菌悬液(D600=1.80)按体积比1∶1∶1混合后制成悬液Ⅰ. 在LB培养基接入等体积的枯草芽孢杆菌、巨大芽孢杆菌、解淀粉芽孢杆菌和纤维单胞菌种子液(D600=1.80), 180 r·min-1, 30℃培养获得D600为1.80的悬液Ⅱ. 等体积混合悬液Ⅰ和Ⅱ后, 以生物炭为载体, 制备成抗生素降解菌剂, 28℃培养48 h后冷冻干燥后室温保存.
1.3 猪粪发酵/堆肥试验固态发酵培养试验:在9 cm培养皿中加入50 g猪粪, 按照0.50%接种量添加抗生素降解菌剂. 在30℃下培养10 d, 测定抗生素残留浓度. 目标猪粪中检出磺胺二甲嘧啶、磺胺甲基异唑、氧氟沙星、培氟沙星、诺氟沙星、环丙沙星、恩诺沙星、替米考星、土霉素、四环素和多西环素.
堆肥试验:在猪粪中加入磺胺二甲氧嘧啶和磺胺嘧啶混合物(质量比为1∶1)达至终浓度为4mg·kg-1; 加入木屑调节堆肥物料水分至50.00% ~60.00%, 初始C/N控制在25~30; 随后按照0.10%的重量比加入抗生素降解菌剂并混匀; 将混合均匀的物料(20 kg)移入泡沫箱(长50 cm, 宽20 cm, 高40 cm)中, 堆肥发酵. 本试验共设置CK(原始猪粪, 不接菌剂)和T(接种菌剂)两个处理. 每个处理3次重复. 自堆肥试验开始, 每天测定和记录堆体温度, 并于第0、2、6、10和14 d取堆肥样品, 一部分密封于4℃冰箱保存, 用于理化性质的测定, 另一部分放置于-80℃冰箱用于堆肥微生物的群落演替分析.
1.4 测试分析方法 1.4.1 气体采集和测定于试验的2、5、7、9和14 d进行氨气和硫化氢气体的采集和测定, 采集装置和方法如图 1所示. 氨气的采集和测定:以磷酸甘油为吸收液, 进行气体采集, 采用靛酚蓝比色法[20]进行氨气挥发量的检测. 硫化氢的采集和测定:以氨性硫酸锌为吸收液进行气体采集, 采用碘量法测定硫化氢的挥发量[21].

|
图 1 臭气采集装置示意 Fig. 1 Odor collecting system |
堆肥氨气排放通量:
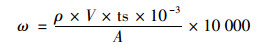
|
(1) |
式中, ω为堆体中氨气排放通量, mg·(m2·h)-1; ρ为从工作曲线中查得显色液中氮的浓度, mg·L-1; V为显色液体积, mL; ts为分取倍数; 10-3为mL换算成L的系数; A为海绵的底表面积, cm2; 10 000为cm2换算成m2的系数.
堆肥氨气累积排放量:
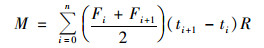
|
(2) |
式中, M为氨气累积排放量, mg·m-2; F为氨气排放通量, mg·(m2·h)-1; t为采样时间, d; i为采样次数; n为采气总次数; ti+1-ti为两次采气间隔天数; R为时间换算系数.
堆肥硫化氢排放通量:
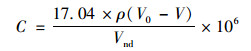
|
(3) |
式中, C为硫化氢排放通量, mg·(m3·h)-1; 17.04为摩尔质量(1/2H2S), g·mol-1; ρ为Na2S2O3标准溶液浓度, mol·L-1; Vnd为样品体积; V0和V分别为空白和样品消耗Na2S2O3体积, mL; 106为换算系数.
堆肥硫化氢累积排放量:
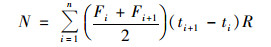
|
(4) |
式中, N为硫化氢累积排放量, mg·m-3; F为硫化氢排放通量, mg·(m3·h)-1; t为采样时间, d; i为采样次数; n为采气总次数; ti+1-ti为两次采气间隔天数; R为时间换算系数.
1.4.2 抗生素提取和测定抗生素提取和测定方法参照Qian等[22]的研究, 具体步骤如下:准确称取1 g样品到50 mL离心管中, 加入内标溶液后放置10 min; 加入15 mL提取液, 涡旋30 s, 超声30 min, 7 000 r·min-1离心10 min, 将上清液转移至圆底烧瓶, 再用15 mL提取液提取残渣一次; 将上述两次上清液混合于圆底烧瓶中, 将圆底烧瓶置于水浴温度为40℃的旋转蒸发仪上去除有机相, 将剩余溶液转移至50 mL离心管中, 并用去离子水定容至40 mL, 离心后取20 mL过SPE固相萃取小柱; 收集洗脱液于玻璃刻度管中, 氮气吹干, 用体积比1∶1的甲醇∶水溶液定容至1 mL, 过0.22 μm有机滤膜, 高效液相色谱-串联质谱(HPLC-MS/MS, AB Sciex Qtrap 5500)测定, 内标法定量. 磺胺类(SAs)、四环素类(TCs)、氟喹诺酮类(FQs)和大环内酯类(MLSs)抗生素使用的提取液分别为乙腈和EDTA-Mcllvain缓冲液(pH=4.0; 3∶7, 体积比, 下同)、Na2EDTA-磷酸盐缓冲液(pH=3.0; 1∶1)、Na2EDTA-磷酸盐缓冲液(pH=7.0; 1∶1)和Na2EDTA-磷酸盐缓冲液(pH=3.0; 1∶1).
1.4.3 基本理化性质测定采用恒温烘箱(DHG-9920, 上海一恒)干燥法, 105℃烘干样品至恒重, 计算样品的含水率. 取新鲜样品按照1∶10固液比浸提后, 取上清液采用pH计进行检测. 全磷含量采用钒钼黄比色法检测, 全钾含量采用火焰光度法检测, 全氮含量采用元素分析仪(Elementar vario MAX, 德国)测定[23].
采用Bruker Avance Ⅱ 400光谱仪对样品进行磁共振碳谱分析, 运用MestReNova 9.0.1软件整理分析堆肥样品中官能团比例.
1.4.4 种子发芽试验为检测堆肥的腐熟程度, 以黄瓜和萝卜种子为对象, 参照栾润宇等[24]的方法在发酵结束后进行种子发芽率试验. 具体步骤如下:称取5 g堆肥样品加蒸馏水25 mL, 200 r·min-1振荡浸提2 h后, 滤纸过滤获得堆肥浸提液. 在9 cm培养皿中放置无菌滤纸后加入5 mL浸提液. 在滤纸上均匀撒入20粒各供试种子, 22℃、相对湿度80.00%和培养48 h后测定种子发芽率.
发芽率(SG)和发芽指数(GI)计算公式如下:

|
(5) |

|
(6) |
称取500 mg堆肥样品, 采用DNA快速提取试剂盒E.Z.N.A.® Stool DNA Kit进行基因组DNA的提取. 经1%琼脂糖凝胶电泳检测质量达标后, 委托联川生物科技有限公司对原核16S V3-V4高变区进行PCR扩增和测序. PCR扩增使用引物对序列为341F(5′-CCTACGGGNGGCWGCAG-3′)和805R(5′- GACTACHVGGGTATCTAATCC-3′). PCR产物经2%琼脂糖凝胶电泳确认无误后由AMPure XT beads(Beckman Coulter Genomics, Danvers, MA, USA)纯化和Qubit(Invitrogen, USA)定量. 上述16S rDNA PCR产物在Illumina NovaSeq平台测序, 利用overlap拼接双端数据并进行质控和嵌合体过滤. 利用DADA2(divisive amplicon denoising algorithm)[25]通过“去重复”(dereplication, 相当于以100%相似度聚类)等步骤获得单碱基精度的代表序列. 采用ASVs(amplicon sequence variants)概念构建类OTUs(operational taxonomic units)[26], 获得最终feature特征表以及特征序列. 测序所得数据进行Blast序列比对, 用SILVA(Release 132, https://www.arb-silva.de/documentation/release-132/)数据库和70%的置信阈值, 对特征序列进行注释, 样本中所有α多样性指标均采用QIIME2计算. 序列提交NCBI数据库, 登录号为PRJNA 797708.
1.4.6 数据分析利用Microsoft Excel 2007和SPSS 19.0软件对所得数据进行整理分析, 用Microsoft Excel 2007进行作图. 为明确微生物ASVs之间的潜在联系, 运用R语言里的corr.test函数分组计算了单个处理组中所有Species之间的Spearman相关性系数. 选择r>0.70, P<0.01的数据, 用Gephi 0.9.2进行可视化分析. 为明确微生物属水平之间的潜在联系, 利用联川生物云平台(https://www.omicstudio.cn/tool)计算了细菌属水平之间及理化性质间的Spearman相关性系数, 选择r>0.70, P<0.01的数据, 利用R version 3.6.1软件构建细菌群落互作网络.
2 结果与讨论 2.1 抗生素降解菌剂在猪粪抗生素去除中的作用效果接种复合微生物菌剂可促进猪粪中抗生素降解. 如表 3所示, 接种抗生素降解菌剂去除猪粪中81.95%的抗生素残留. 与对照相比, 接种菌剂处理的猪粪中抗生素残留总量下降了42.18%, 其中磺胺类抗生素残留浓度下降41.85%, 四环素类残留浓度下降42.39%. 本结果表明, 添加的四环素类和磺胺类抗生素降解菌在猪粪抗生素去除中发挥了作用. 此外, 添加菌剂处理中氟喹诺酮类和大环内酯类抗生素含量也分别较对照降低29.44%和20.20%. 一方面, 复合菌剂中的其他菌种可能也具有一定抗生素分解作用, 另一方面菌剂的添加有助于形成适合抗生素降解的环境. 例如, Wu[27]的研究指出, 微生物添加剂促进了土著抗生素降解菌群的代谢活动. 目前通过微生物菌剂添加促进粪便中抗生素降解的研究不在少数, 例如肖礼等[28]通过添加白腐真菌、氨化和硝化菌剂使得粪便中抗生素去除率增加了10.00%. Liang等[29]的研究表明, 在赭曲霉、念珠菌和伯克霍尔德菌复合菌剂的添加下, 粪便中四环素和多西环素残留也有所下降. 相比上述报道的菌剂, 本文研究的复合微生物菌剂在畜禽粪便中的抗生素去除效果较好, 可降解的抗生素类型更丰富.
|
|
表 3 接种菌剂对猪粪中抗生素降解的影响1) Table 3 Effect of inoculating microbial agent on antibiotic degradation in pig manure |
2.2 接种抗生素降解菌剂对堆肥理化进程的影响 2.2.1 堆肥温度、水分、pH和总养分变化
温度是影响堆肥质量的重要环境参数[30]. 高温可促进堆体基质中有机和有毒物质分解, 还能抑制且杀死粪肥中残留的病菌和虫卵[31]. 添加菌剂处理在堆肥第5 d达到高温峰值55℃, 在高温时段(50~55℃)持续2 d; 未添加菌剂的对照升温缓慢且高温峰值低, 仅在堆肥第7 d时达到48℃[图 2(a)]. 因此, 抗生素降解菌接种加快堆体升温.

|
CK:对照, T:接种菌剂, 下同 图 2 接种菌剂对堆肥温度、水分、pH和总养分的影响 Fig. 2 Effect of inoculating microbial agent on temperature, moisture, pH, and total nutrients of compost |
降低畜禽粪便中水分含量是堆肥的重要目标之一[31]. 如图 2(b)所示, 相比对照处理, 接种抗生素降解菌剂处理组的水分含量下降速度更快, 整体数值更低, 表明该菌剂接种促进水分去除, 有助于堆肥干燥. 升温引发的水汽蒸发是堆肥干燥的重要原因. 因此, 抗生素降解菌剂接种可能通过促进升温, 加快水分去除.
pH值影响堆体中微生物的生长和繁殖, 故而是影响堆肥腐熟效果的另一重要环境因素. 图 2(c)显示抗生素降解菌剂接种对堆肥过程pH变化影响不大. 在堆肥过程中, 所有处理组的pH值2 d后均呈快速上升趋势, 在6 d后逐渐下降并趋于平稳. 堆肥过程初期pH的迅速上升可能是因为高温导致的有机酸挥发以及微生物对含氮有机物的降解导致的NH3释放和NH4+积累所致, 而后期的pH逐渐降低则是缘于有机酸的产生以及微生物的脱氮作用[32].
微生物菌剂对畜禽粪便堆肥氮磷钾总含量的影响如图 2(d)所示. 与王亚飞等[33]的研究结果相似, 堆肥过程中总养分含量整体呈上升趋势. 相比对照处理, 接种抗生素降解菌剂处理促进堆肥总养分积累. 在堆肥结束时, 接种抗生素降解菌剂处理堆肥中的氮磷钾(NPK)含量相比对照处理增加了6.80%(P<0.05). 添加菌剂可能通过促进堆肥养分的转化, 减少养分流失, 进而提高猪粪有机肥养分含量.
2.2.2 臭气释放量变化氨气和硫化氢是堆肥过程中臭气的主要成分, 其与堆肥中微生物的代谢活动和有机质分解密切相关. 已有研究指出, 在堆肥中接种微生物菌剂改变微生物代谢活动, 降低氨气和硫化氢的产生, 促进二者的氧化分解, 从而降低排放, 减少污染[34, 35]. 因此, 关注抗生素降解菌剂对堆肥在臭气减排中的作用效果具有重要意义. 如图 3(a)所示, 堆肥过程中硫化氢排放速率随着堆肥进程呈直线下降, 其中接种抗生素降解菌剂处理堆肥中的硫化氢排放整体低于对照处理, 因此本抗生素降解菌剂在硫化氢减排中有一定的贡献. 从氨气减排上看, 与对照相比, 添加菌剂的处理在高峰期氨气释放更少[图 3(b)]. 图 3(c)和3(d)计算了整个堆肥过程中臭气的总排放量, 结果表明添加微生物复合菌剂能显著(P<0.05)降低氨气和硫化氢的总排放.

|
(a)和(b)表示硫化氢和氨气的排放通量, (c)和(d)表示硫化氢和氨气的累计排放量; 不同小写字母表示不同处理间差异显著(P<0.01) 图 3 接种菌剂对硫化氢和氨气排放通量和累积排放量的影响 Fig. 3 Effect of inoculating microbial agent on emission flux and cumulative emission of H2S and NH3 |
种子发芽指数(GI)能有效反映基质对植物毒性的大小, 可反映堆肥的腐熟程度. 一般认为当GI>50.00%时堆肥基本腐熟, 毒性在植物可承受范围内, GI ≥80.00%时, 认为堆肥已完全腐熟, 没有毒性[36].
如表 4所示, 接种抗生素降解菌剂处理堆肥的黄瓜和萝卜种子发芽指数(GI)和发芽率(SG)均大于对照, 且发芽指数均大于70.00%, 其中萝卜种子的发芽指数高达293.09%, 这表明堆肥浸提液不仅对萝卜种子无毒, 且对萝卜种子的萌发具有促进作用. 综上, 接种抗生素降解菌剂促进堆肥腐熟, 甚至促进萝卜种子萌发.
|
|
表 4 接种菌剂对种子发芽率和发芽指数的影响/% Table 4 Effect of inoculating microbial agent on seed germination rate and germination index/% |
2.3.2 堆肥有机碳组分变化
采用磁共振碳谱分析了猪粪堆肥产物, 对比接种抗生素降解菌剂对猪粪堆肥后有机碳组分的影响, 根据文献[37~39]中对碳谱的归类方法得出堆肥发酵产物的主要官能团及其所占比例, 结果见表 5.
|
|
表 5 不同处理堆肥产物官能团的相对比例/% Table 5 Relative proportion of functional groups in composts from different treatments/% |
相比对照, 接种菌剂处理堆肥的芳香类碳含量更高, 表明接种抗生素降解菌剂增加堆肥中稳定性有机质形成. 相比对照, 菌剂接种处理堆肥中木质素、纤维素和半纤维素等难降解有机质的含量降低, 烷基碳含量增加, 暗示抗生素降解菌剂促进堆体中纤维素、半纤维素和木质素的分解, 并向可利用的烷基类碳转化. 综上, 抗生素降解菌剂的应用促进堆肥有机质腐殖化. 易分解有机质向稳定性有机质的转变是堆肥腐熟的重要指标[40]. 因此, 菌剂接种对碳谱的影响进一步反映了菌剂促进堆肥腐熟.
2.4 接种抗生素降解菌剂对堆肥细菌群落演替的影响 2.4.1 添加菌剂对堆肥细菌群落结构的影响粪肥样品的16S V3-V4区测序结果经质控筛选后, 共获得3 661 248条有效序列, 从60个样品中鉴定出14 561 ASVs. 经Blast比对后选取丰度最高的前30个物种构建柱状堆叠, 结果如图 4所示. 从中可知, 与Mao等[34]的研究发现一致, 猪粪堆肥中在门水平上占优势的菌群主要有Firmicutes(厚壁菌门)、Proteobacteria(变形菌门)、Actinobacteria(放线菌门)和Bacteridetes(拟杆菌门), 但其相对丰度在堆肥的不同时期各异. 随着时间的推移, 堆肥中的微生物群落结构发生了明显的变化. 尽管在各个阶段Firmicutes门细菌在处理和对照中均占据绝对优势, 但是其他细菌的优势种群出现了更迭, 如发酵伊始除Firmicutes外, Actinobacteria门细菌占比最高, 2 d后Bacteridetes(拟杆菌门)细菌成为优势种群, 6 d后Proteobacteria细菌数量显著增加并于发酵末期占据优势, 而在发酵初期作为微生物重要组成的Tenericutes(软壁菌门)细菌在堆肥结束后几乎检测不到[图 4(a)]. 堆肥发酵2 d后随着升温阶段的到来微生物群落结构在属水平上也发生了显著变化, 原先占绝对优势的Sporosarcina(芽孢八叠球菌)被Cerasibacillus(樱桃样芽胞杆菌)、Bacillus(芽孢杆菌)和Halobacillus(嗜盐芽孢杆菌)替代[图 4(b)].

|
(a)和(b)分别表示门水平和属水平; 横坐标中的数字0、2、6、10和14表示采样时间0、2、6、10和14 d 图 4 不同堆肥处理中细菌群落组成在门水平和属水平的相对丰度 Fig. 4 Relative abundance of taxa of bacterial community composition at the phylum level and genus level in different composting treatments |
外源添加菌剂改变了原有微生物的群落结构, 提高了早期堆肥中Actinobacteria的丰度. 此外, 在堆肥早期和中期(0~10 d), 添加菌剂处理组的厚壁菌门(Firmicutes)的相对丰度也高于对照. 由于菌剂中Actinobacteria和Firmicute细菌占较大优势, 因此上述细菌门相对丰度增加可能与外源菌剂中微生物的定殖有一定关系. 不过, 除了Bacillus属细菌丰度显著增加外(P<0.05), 未能在属或种水平上鉴定到另外几株外源接种菌, 这也可能与高通量测序技术在菌种鉴定方面的局限性有关.除Bacillus以外, 添加菌剂还诱导Cerasibacillus、Pseudogracilibacillus(假纤细芽胞杆菌属)、Halobacillus、Paenalcaligenes(产碱杆菌属)、Virgibacillus(枝芽孢杆菌属)、Lactobacillus(乳杆菌属)和Sphingobacteriaceae(鞘脂杆菌)等属细菌数量增加.
Bacillus属细菌是常见的从环境中分离得到的木质纤维素降解微生物[41], 因此本试验中添加菌剂处理的堆肥产物中纤维素和半纤维含量的下降可能与Bacillus菌群数量的增加有关. 有报道证实, 一些Lactobacillus属细菌能通过酶类物质氧化分解堆体中产生的硫化氢, 减少臭气的挥发量[42], 而其产生的乳酸能够抑制堆体中有害微生物的活性[43]. 在本研究中, 随着堆肥的进行, 添加菌剂处理组的Lactobacillus的相对丰度逐渐增加, 直至堆肥末期相对丰度最高, 这可能是菌剂处理后堆肥除臭效果更佳和种子发芽率更高的一个原因. 肠杆菌科细菌含有多种人体致病菌, 因此常被认为与疾病相关. 在本研究中堆肥发酵10 d后, 添加菌剂处理的肠杆菌(Enterobacter)相对丰度已无法检出, 而对照中肠杆菌的相对丰度在发酵14 d后仍残留0.60%. 因此, 接种抗生素降解菌剂有助于杀灭有害微生物.
2.4.2 微生物菌群网络共存分析基于Spearman相关性分析, 在ASV水平上构建了堆肥细菌互作网络拓扑[图 5(a)和5(b)]. 拓扑特性通常用于描述细菌相互关系的复杂模式. 结果表明, 对照堆肥的细菌群落网络由93个节点组成, 连接1 583条边, 而添加菌剂的处理组互作网络节点和连接大幅下降[图 5(a)和5(b)]. 添加菌剂的处理组的直径(即最远处两个节点之间的距离)小于对照, 平均网络距离增大, 表明添加菌剂的处理组细菌网络连通性低于对照. 网络中的正相关(positive)和负相关(negative)可以用来表征微生物之间的相关关系, 如微生物间存在互生或共生的积极关系呈正相关, 若存在竞争或捕食关系则呈负相关[44]. 图 5表明, 对照堆肥细菌群落网络中正相关作用占32.20%, 而接种菌剂处理中正相关作用下降至20.80%. 综上, 添加菌剂大幅降低了细菌群落互作网络复杂度和连通性, 细菌之间的共生和协同作用减弱.

|
(a)和(b)为ASV水平上构建的对照和菌剂处理堆肥的细菌群落互作网络拓扑, 每一个节点表示ASV, 节点的颜色表示ASV聚类模块, 连接线表示它们之间的相互作用; 红色线表示显著正相关, 绿色线表示显著负相关; (c)和(d)为基于对照和菌剂处理堆肥中种群丰度前30的细菌属和堆肥温度间相关性构建的网络, 每个圆形节点表示一个细菌属, 三角形表示温度; 节点的大小表示与该指标关联的其他指标的数目多少, 连接线表示它们之间的相互作用; 灰色线表示显著负相关, 黄色线表示显著正相关 图 5 细菌共存模式的网络分析 Fig. 5 Network analysis of the bacterial coexistence pattern |
进一步选取种群丰度前30的细菌属和堆肥温度构建了相关性网络[图 5(c)和5(d)]. 可见, 对照堆肥中连接性最高的5个节点对应的细菌属为Proteiniphilum、Fastidiosipila、Terrisporobacter、Bacteroides和Turicibacter; 而添加菌剂的处理组连接性最高的属为Pseudogracilibacillus、Bacteroides、Bacillus、Fermentimonas和Proteiniphilum. 与ASV水平的细菌共现网络结果相同, 属水平的细菌相关性网络在对照处理中也更为复杂. 接种菌剂降低了属水平的细菌相关性网络的复杂性, 使得Bacillus和Pseudogracilibacillus在相关性网络中的重要性增加. 相比对照, 接种菌剂处理堆肥中Bacillus和Pseudogracilibacillus与其他细菌种群之间的竞争关系减弱, 多建立互生或共生的积极关系.堆体温度(temperature)与Bacillus和Pseudogracilibacillus呈极显著正相关(P<0.01, R2=0.999). 在接种的菌剂中, Bacillus是重要的菌种之一. Pseudogracilibacillus被报道在大分子有机物分解中发挥重要作用[45]. 因此, Bacillus和Pseudogracilibacillus在细菌互作网络中重要性的增加以及与其他细菌竞争性的下降解释了菌剂处理组堆体升温更快、温度更高和粪肥腐熟效果更好的原因. 此外, Bacillus和Pseudogracilibacillus在堆肥中产生积极的正相关促进关系, 与致病菌Enterobacter等产生竞争或抑制的负相关关系, 这与上述外源菌剂添加后堆温升高和致病菌下降等现象具有一致性. 综上, 外源抗生素降解菌剂的进入改善了原有土著细菌群落之间的相互关系, 优化了有益细菌与其他菌群的生态关系, 为建立新的和更加健康的堆肥细菌群落奠定了基础.
3 结论抗生素降解菌剂的应用不仅可促进猪粪中抗生素去除, 也可改善堆肥发酵效果, 包括有效促进了堆肥升温, 加速堆体水分去除, 提高了堆肥产物中氮磷钾养分总含量, 减少了臭气的排放; 促进堆肥中纤维素和半纤维素等难利用物质向稳定性芳香碳和可利用烷基碳类物质转变, 提高堆肥产物种子发芽率; 改变了堆体中微生物群落结构, 增加有助于堆体升温、有机物异化和臭气分解的细菌, 减少疾病相关的肠杆菌. 堆肥中土著细菌群落之间存在紧密的相互作用, 而外源抗生素降解菌剂的进入打破了原有土著细菌群落之间的平衡, 为新的且更加健康的堆肥细菌群落的建立奠定了基础. 本试验结果为菌剂未来的实际应用提供了初步理论依据.
| [1] | Wang K, Zhuang T, Su Z X, et al. Antibiotic residues in wastewaters from sewage treatment plants and pharmaceutical industries: occurrence, removal and environmental impacts[J]. Science of the Total Environment, 2021, 788. DOI:10.1016/j.scitotenv.2021.147811 |
| [2] |
张晓娇, 柏杨巍, 张远, 等. 辽河流域地表水中典型抗生素污染特征及生态风险评估[J]. 环境科学, 2017, 38(11): 4553-4561. Zhang X J, Bai Y W, Zhang Y, et al. Occurrence, distribution, and ecological risk of antibiotics in surface water in the Liaohe River Basin, China[J]. Environmental Science, 2017, 38(11): 4553-4561. |
| [3] | Wang L J, Wang J, Wang J H, et al. Distribution characteristics of antibiotic resistant bacteria and genes in fresh and composted manures of livestock farms[J]. Science of the Total Environment, 2019, 695. DOI:10.1016/j.scitotenv.2019.133781 |
| [4] | Hill D, Morra M J, Stalder T, et al. Dairy manure as a potential source of crop nutrients and environmental contaminants[J]. Journal of Environmental Sciences, 2021, 100: 117-130. DOI:10.1016/j.jes.2020.07.016 |
| [5] |
王小彬, 闫湘, 李秀英. 畜禽粪污厌氧发酵沼液农用之环境安全风险[J]. 中国农业科学, 2021, 54(1): 110-139. Wang X B, Yan X, Li X Y. Environmental safety risk for application of anaerobic fermentation biogas slurry from livestock manure in agricultural land in China[J]. Scientia Agricultura Sinica, 2021, 54(1): 110-139. |
| [6] |
张宁, 李淼, 刘翔. 土壤中抗生素抗性基因的分布及迁移转化[J]. 中国环境科学, 2018, 38(7): 2609-2617. Zhang N, Li M, Liu X. Distribution and transformation of antibiotic resistance genes in Soil[J]. China Environmental Science, 2018, 38(7): 2609-2617. DOI:10.3969/j.issn.1000-6923.2018.07.029 |
| [7] | Zhu L J, Zhao Y, Zhang W S, et al. Roles of bacterial community in the transformation of organic nitrogen toward enhanced bioavailability during composting with different wastes[J]. Bioresource Technology, 2019, 285. DOI:10.1016/j.biortech.2019.121326 |
| [8] | Mahboob M, Jahan R. A new and a known species of the genus Mesorhabditis (Osche, 1952) Dougherty, 1953 associated with the larva of longhorn beetle (Cerambycidae) and ground beetle (Scarabaeidae)[J]. Archives of Phytopathology and Plant Protection, 2021, 54(15-16): 1087-1101. DOI:10.1080/03235408.2021.1887662 |
| [9] | Zhang J, Lin H, Ma J W, et al. Compost-bulking agents reduce the reservoir of antibiotics and antibiotic resistance genes in manures by modifying bacterial microbiota[J]. Science of the Total Environment, 2019, 649: 396-404. DOI:10.1016/j.scitotenv.2018.08.212 |
| [10] | Liu H D, Ye X H, Chen S L, et al. Chitosan as additive affects the bacterial community, accelerates the removals of antibiotics and related resistance genes during chicken manure composting[J]. Science of the Total Environment, 2021, 792. DOI:10.1016/j.scitotenv.2021.148381 |
| [11] | Liao H P, Zhao Q, Cui P, et al. Efficient reduction of antibiotic residues and associated resistance genes in tylosin antibiotic fermentation waste using hyperthermophilic composting[J]. Environment International, 2019, 133. DOI:10.1016/j.envint.2019.105203 |
| [12] |
沈东升, 何虹蓁, 汪美贞, 等. 土霉素降解菌TJ-1在猪粪无害化处理中的作用[J]. 环境科学学报, 2013, 33(1): 147-153. Sheng D S, He H Z, Wang M Z, et al. The role of oxytetracycline-degrading bacterium TJ-1 on the hazard-free treatment of pig manure[J]. Acta Scientiae Circumstantiae, 2013, 33(1): 147-153. |
| [13] | Zhang S B, Xia T Y, Wang J L, et al. Role of Bacillus inoculation in rice straw composting and bacterial community stability after inoculation: unite resistance or individual collapse[J]. Bioresource Technology, 2021, 337. DOI:10.1016/j.biortech.2021.125464 |
| [14] | Liu Y T, Zheng L, Cai Q J, et al. Simultaneous reduction of antibiotics and antibiotic resistance genes in pig manure using a composting process with a novel microbial agent[J]. Ecotoxicology and Environmental Safety, 2021, 208. DOI:10.1016/j.ecoenv.2020.111724 |
| [15] | Du G L, Shi J P, Zhang J X, et al. Exogenous probiotics improve fermentation quality, microflora phenotypes, and trophic modes of fermented vegetable waste for animal feed[J]. Microorganisms, 2021, 9(3): 644. DOI:10.3390/microorganisms9030644 |
| [16] | Shi Y K, Lin H, Ma J W, et al. Degradation of tetracycline antibiotics by Arthrobacter nicotianae OTC-16[J]. Journal of Hazardous Materials, 2021, 403. DOI:10.1016/j.jhazmat.2020.123996 |
| [17] | 林辉, 马军伟, 孙万春, 等. 一种磺胺类抗生素降解产碱菌及其应用[P]. 中国专利: CN 108823115A, 2018-11-16. |
| [18] | Ghirardini A, Grillini V, Verlicchi P. A review of the occurrence of selected micropollutants and microorganisms in different raw and treated manure - Environmental risk due to antibiotics after application to soil[J]. Science of the Total Environment, 2020, 707. DOI:10.1016/j.scitotenv.2019.136118 |
| [19] | Tortosa G, Fernández-González A J, Lasa A V, et al. Involvement of the metabolically active bacteria in the organic matter degradation during olive mill waste composting[J]. Science of the Total Environment, 2021, 789. DOI:10.1016/j.scitotenv.2021.147975 |
| [20] |
邹娟, 胡学玉, 张阳阳, 等. 不同地表条件下生物炭对土壤氨挥发的影响[J]. 环境科学, 2018, 39(1): 348-354. Zou J, Hu X Y, Zhang Y Y, et al. Effect of biochar on ammonia volatilization from soils of different surface conditions[J]. Environmental Science, 2018, 39(1): 348-354. DOI:10.3969/j.issn.1000-6923.2018.01.039 |
| [21] | Yuan J, Li Y, Zhang H Y, et al. Effects of adding bulking agents on the biodrying of kitchen waste and the odor emissions produced[J]. Journal of Environmental Sciences, 2018, 67: 344-355. DOI:10.1016/j.jes.2017.08.014 |
| [22] | Qian M R, Wu H Z, Wang J M, et al. Occurrence of trace elements and antibiotics in manure-based fertilizers from the Zhejiang Province of China[J]. Science of the Total Environment, 2016, 559: 174-181. DOI:10.1016/j.scitotenv.2016.03.123 |
| [23] |
王飞, 李清华, 何春梅, 等. 稻秆与紫云英联合还田提高黄泥田氮素利用率和土壤肥力[J]. 植物营养与肥料学报, 2021, 27(1): 66-74. Wang F, Li Q H, He C M, et al. Combined returning of milk vetch and rice straw improves fertilizer nitrogen recovery and fertility of yellow-mud paddy soil[J]. Journal of Plant Nutrition and Fertilizers, 2021, 27(1): 66-74. |
| [24] |
栾润宇, 高珊, 徐应明, 等. 不同钝化剂对鸡粪堆肥重金属钝化效果及其腐熟度指标的影响[J]. 环境科学, 2020, 41(1): 469-478. Luan R Y, Gao S, Xu Y M, et al. Effect of different passivating agents on the stabilization of heavy metals in chicken manure compost and its maturity evaluating indexes[J]. Environmental Science, 2020, 41(1): 469-478. |
| [25] | Callahan B J, McMurdie P J, Rosen M J, et al. DADA2: high-resolution sample inference from Illumina amplicon data[J]. Nature Methods, 2016, 13(7): 581-583. DOI:10.1038/nmeth.3869 |
| [26] | Blaxter M, Mann J, Chapman T, et al. Defining operational taxonomic units using DNA barcode data[J]. Philosophical Transactions of the Royal Society B: Biological Sciences, 2005, 360(1462): 1935-1943. DOI:10.1098/rstb.2005.1725 |
| [27] | Wu Z, Luo Y N, Bao J Z, et al. Additives affect the distribution of metabolic profile, microbial communities and antibiotic resistance genes in high-moisture sweet corn kernel silage[J]. Bioresource Technology, 2020, 315. DOI:10.1016/j.biortech.2020.123821 |
| [28] |
肖礼, 黄懿梅, 赵俊峰, 等. 外源菌剂对猪粪堆肥质量及四环素类抗生素降解的影响[J]. 农业环境科学学报, 2016, 35(1): 172-178. Xiao L, Huang Y M, Zhao J F, et al. Effects of exogenous microbial agents on pig manure compost quality and tetracycline antibiotic degradation[J]. Journal of Agro-Environment Science, 2016, 35(1): 172-178. |
| [29] | Liang J D, Jin Y M, Wen X, et al. Adding a complex microbial agent twice to the composting of laying-hen manure promoted doxycycline degradation with a low risk on spreading tetracycline resistance genes[J]. Environmental Pollution, 2020, 265. DOI:10.1016/j.envpol.2020.114202 |
| [30] | Ajmal M, Shi A P, Awais M, et al. Ultra-high temperature aerobic fermentation pretreatment composting: parameters optimization, mechanisms and compost quality assessment[J]. Journal of Environmental Chemical Engineering, 2021, 9(4). DOI:10.1016/j.jece.2021.105453 |
| [31] | Bao Y Y, Feng Y Z, Qiu C W, et al. Organic matter- and temperature-driven deterministic assembly processes govern bacterial community composition and functionality during manure composting[J]. Waste Management, 2021, 131: 31-40. DOI:10.1016/j.wasman.2021.05.033 |
| [32] | Wang K, Ma X C, Yin X Q, et al. Difference and interplay of microbial communities, metabolic functions, trophic modes and influence factors between sludge and bulking agent in a composting matrix[J]. Bioresource Technology, 2021, 336. DOI:10.1016/j.biortech.2021.125085 |
| [33] |
王亚飞, 李梦婵, 邱慧珍, 等. 不同畜禽粪便堆肥的微生物数量和养分含量的变化[J]. 甘肃农业大学学报, 2017, 52(3): 37-45. Wang Y F, Li M C, Qiu H Z, et al. Changes of microbial quantity and nutrient content in different composting of livestock manure[J]. Journal of Gansu Agricultural University, 2017, 52(3): 37-45. |
| [34] | Mao H, Lv Z Y, Sun H D, et al. Improvement of biochar and bacterial powder addition on gaseous emission and bacterial community in pig manure compost[J]. Bioresource Technology, 2018, 258: 195-202. DOI:10.1016/j.biortech.2018.02.082 |
| [35] | Wang Y, Bi L L, Liao Y H, et al. Influence and characteristics of Bacillus stearothermophilus in ammonia reduction during layer manure composting[J]. Ecotoxicology and Environmental Safety, 2019, 180: 80-87. DOI:10.1016/j.ecoenv.2019.04.066 |
| [36] | López-González J A, del Carmen Vargas-García M, López M J, et al. Biodiversity and succession of mycobiota associated to agricultural lignocellulosic waste-based composting[J]. Bioresource Technology, 2015, 187: 305-313. DOI:10.1016/j.biortech.2015.03.124 |
| [37] | Alekseev I, Abakumov E. 13C-NMR spectroscopy of humic substances isolated from the agricultural soils of Puchuncavi (El Melón and Puchuncavi areas), central Chile[J]. Soil and Water Research, 2020, 15(3): 191-198. DOI:10.17221/81/2019-SWR |
| [38] | Suzuki R, Uesawa Y, Okada Y, et al. Use of 13C-NMR chemical shifts; application of principal component analysis for categorizing structurally similar methoxyflavones and correlation analysis between chemical shifts and cytotoxicity[J]. Chemical and Pharmaceutical Bulletin, 2021, 69(2): 199-202. DOI:10.1248/cpb.c20-00778 |
| [39] | Chung T L, Chen J S, Chiu C Y, et al. 13C-NMR spectroscopy studies of humic substances in subtropical perhumid montane forest soil[J]. Journal of Forest Research, 2012, 17(6): 458-467. DOI:10.1007/s10310-011-0319-9 |
| [40] | Luo Y J, van Veelen H P J, Chen S Y, et al. Effects of sterilization and maturity of compost on soil bacterial and fungal communities and wheat growth[J]. Geoderma, 2022, 409. DOI:10.1016/j.geoderma.2021.115598 |
| [41] |
郭亚萍, 张国庆, 陈青君, 等. 双孢蘑菇堆肥过程中细菌群落结构分析[J]. 应用与环境生物学报, 2014, 20(5): 832-839. Guo Y P, Zhang G Q, Chen Q J, et al. Bacterial community structure analysis for mushroom (Agaricus bisporus) compost using PCR-DGGE technique[J]. Chinese Journal of Applied & Environmental Biology, 2014, 20(5): 832-839. |
| [42] | Li W C, Liu Y H, Hou Q C, et al. Lactobacillus plantarum improves the efficiency of sheep manure composting and the quality of the final product[J]. Bioresource Technology, 2020, 297. DOI:10.1016/j.biortech.2019.122456 |
| [43] |
马石霞, 摆倩文, 周魏, 等. 微生物除臭剂应用于畜禽养殖场的研究现状[J]. 浙江农业学报, 2021, 33(8): 1552-1564. Ma S X, Bai Q W, Zhou W, et al. Application of microbial deodorant in livestock and poultry farms[J]. Acta Agriculturae Zhejiangensis, 2021, 33(8): 1552-1564. DOI:10.3969/j.issn.1004-1524.2021.08.22 |
| [44] | Layeghifard M, Hwang D M, Guttman D S. Disentangling interactions in the microbiome: a network perspective[J]. Trends in Microbiology, 2017, 25(3): 217-228. DOI:10.1016/j.tim.2016.11.008 |
| [45] | Li H, Li J, Wan Q, et al. Bioremediation mechanism of Monensin contaminated chicken manure by a combination of housefly larvae and Stenotrophomonas sp. DM- 2[J]. Environmental Technology & Innovation, 2021, 21. DOI:10.1016/j.eti.2020.101269 |