随着水体富营养化现象的日益严重, 越来越多的水生态系统受到严重威胁, 蓝藻水华已成为全球性环境问题之一[1~3].铜绿微囊藻(Microcystis aeruginosa)是蓝藻水华中常见的优势藻种, 可产生有毒代谢产物, 威胁水生态系统的生物多样性和人类身体健康[4, 5].蓝藻水华暴发时, 通常会采取物理和化学方法来降低内源污染[6, 7].但实际上, 蓝藻水华并没有得到长期有效的控制, 而是陷入了“暴发-治理-暴发”的反复循环.
微藻在生长过程中会分泌大量胞外产物, 包括肽类、脂类、碳水化合物和维生素等, 藻际环境(phycosphere)是以藻类分泌物为骨架构成的微型生态结构, 生活着大量细菌群落[8].一般情况下, 这些细菌可以利用藻类的胞外代谢产物作为营养物质来生长繁殖, 并附着在藻细胞表面, 形成稳定的微生物群落[9, 10].此外, 藻际细菌在生长繁殖中也释放了多种代谢产物, 对微藻的生长、孢囊形成和死亡都产生重要影响[11, 12].不同微生物类群可在不同程度上促进或抑制藻华暴发[13], 因此, 菌藻间的相互作用在水生态平衡中起着至关重要的作用.已有不少学者对溶藻细菌进行了分离鉴定[14~16], 并研究了其抑藻性能和抑藻机制[17, 18].然而, 由于水环境的复杂性, 微藻与整个藻际细菌群落的互作关系还有待深入研究.
本研究以铜绿微囊藻为供试藻种, 以LB培养基、蛋白胨、葡萄糖、柠檬酸钠和醋酸钠为营养物质, 探究原位营养刺激后藻细胞的生长情况和生理生化响应, 分析藻际细菌群落变化, 并尝试在藻际细菌中筛选溶藻菌, 以期推进蓝藻水华防治和菌藻互作关系的研究.
1 材料与方法
1.1 藻种来源与培养方法
铜绿微囊藻(PCC7820)购自中国科学院水生生物研究所, 采用灭菌后的BG11培养基, 于光照培养箱中培养至稳定期.培养条件:温度25℃、光照强度2 000 lx、光暗比12 h∶12 h[19], 每天固定时间摇动1次藻液, 避免藻细胞贴壁生长.
1.2 实验方法
将LB培养基, 10 g·L-1的蛋白胨, 4 g·L-1的葡萄糖、柠檬酸钠和醋酸钠均置于高压锅灭菌后备用.分别取25 mL上述营养液添加至475 mL铜绿微囊藻培养液, 设为A、B、C、D和E组.A组LB培养基终浓度为1.25 g·L-1, B组蛋白胨终浓度为0.5 g·L-1, C组葡萄糖终浓度为0.2 g·L-1, D组柠檬酸钠终浓度为0.2 g·L-1, E组醋酸钠终浓度为0.2 g·L-1[20].对照组(CK组)添加等量无菌水, 每组设置3个平行, 于光照培养箱中培养, 培养条件同1.1节.每2 d取一定量藻液测定各指标.
1.3 测定方法
叶绿素a(Chl-a)采用热乙醇法测定[21], 抑藻率计算见式(1):
式中, ρ1指样品当天Chl-a浓度(mg·L-1); ρ2指对照组当天Chl-a浓度(mg·L-1).
采用16S rRNA高通量测序检测铜绿微囊藻藻际环境中细菌群落组成及物种丰度, 由上海美吉生物医药科技有限公司完成测定.
活性氧采用活性氧试剂盒测定(北京索莱宝科技有限公司, 中国).粗酶液的制备:取各实验组样品10 mL, 8 000 r·min-1离心10 min, 弃上清.用50 mmol·L-1PBS缓冲液重悬藻细胞, 细胞破碎仪(JY96-IIN, 上海沪析实业有限公司, 中国)超声15 min, 收集上清液即为粗酶液, 4℃保存, 用于蛋白质含量、丙二醛(MDA)含量、超氧化物歧化酶(SOD)和过氧化氢酶(CAT)活性的测定[22].蛋白质含量采用考马斯亮蓝法, MDA含量采用硫代巴比妥酸-分光光度法, SOD活性采用NBT(氮蓝四唑)光还原法, CAT活性采用紫外吸收法.
将待测样品过0.45 μm滤膜后得到溶解性有机物(DOM)溶液, 使用荧光分光光度计(FL970 Plus, 天美科技有限公司, 中国)进行组成测定.
1.4 扫描电镜观察
取LB培养基组样品10 mL, 8 000 r·min-1离心10 min, 弃上清.用2.5%戊二醛溶液固定细胞12 h后, 用磷酸盐缓冲液(PBS)洗涤细胞3次.随后, 将样品分别用30%、50%、70%、85%、95%和100%的乙醇进行梯度脱水处理, 各浓度均处理15 min, 乙酸异戊酯浸泡4 h[23].最后, CO2临界点干燥处理, 扫描电镜(Quattro C, ThermoFisher, 美国)观察细胞形态.
1.5 铜绿微囊藻藻际溶藻菌的分离鉴定
分别将A组和B组样品进行梯度稀释, 并涂布至平板.培养48 h后挑取形态各异的单菌落进行分离划线, 将纯化后的菌株与铜绿微囊藻共培养, 筛选溶藻菌[24].采用16S rRNA测序(由北京睿博兴科生物技术有限公司进行检测), 并与GenBank数据库(http://www.ncbi.nlm.nih.gov/blast)中的序列进行比对, 确定菌株种属.将所筛溶藻菌的原菌液离心弃上清, 采用无菌水重悬, 将各种菌悬液分别添加至藻液, 对照组添加等量无菌水.每组均设置3个平行, 10 d后测算各实验组的抑藻率.
1.6 数据分析
所有细菌群落组成及物种丰度的数据使用美吉生信云平台进行分析和制图; 其余数据使用Excel 365和Origin 2018软件进行分析、处理和制图, 其中误差棒代表 3次实验之间的标准偏差.
2 结果与讨论
2.1 营养物质对铜绿微囊藻生长的影响
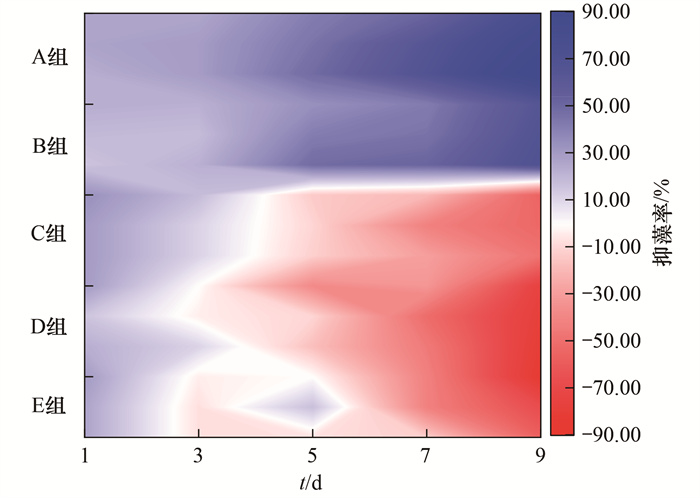
Chl-a是植物光合作用的重要色素, 测定Chl-a浓度即可了解样本中浮游植物的现存量[25, 26].因此, 本研究测定了藻液中Chl-a含量, 以分析铜绿微囊藻的生长情况.由图 1可知, 添加营养物质至铜绿微囊藻培养液后, 藻细胞生长受到严重影响.蛋白胨(有机氮源)和LB培养基(细菌培养基)分别添加至藻液后, 9 d时的抑藻率分别可高达60%和85%以上; 乙酸钠、葡萄糖和柠檬酸钠(均为碳源)分别添加至藻液后, 9 d时的增长率分别可达50%、70%和80%以上.上述结果表明LB培养基和蛋白胨(A组和B组)可明显抑制铜绿微囊藻生长, 而葡萄糖、柠檬酸钠和乙酸钠(C、D和E组)可显著促进铜绿微囊藻生长.且随着处理时间的增加, 上述5种营养物质抑制或促进作用愈发显著.然而, 丁宁等[20]的研究发现, 2216E培养基、甘油和柠檬酸钠均可抑制米氏凯伦藻(Karenia mikimotoi)生长, 抑藻率90%以上.结合本研究可初步推测:大多数细菌培养基可以抑制藻类生长, 而碳源可能促进也可能抑制藻类生长.
2.2 藻际环境中细菌群落组成及物种丰度
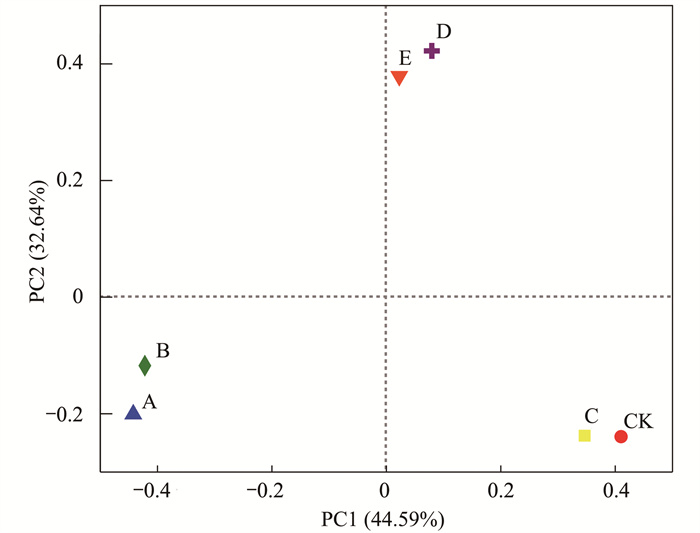
藻际环境中存在着丰富多样的细菌类群, 不同营养物质刺激后的藻际细菌群落组成差异较大.采用主坐标分析(PCoA分析)表征各实验组藻际细菌群落的相似性和差异性, 如图 2所示, A组与B组、C组与CK组、D组与E组相似性较高.此外, 蛋白胨和LB培养基的添加可使藻际环境中细菌数量显著增加(图 3), 其Shannon多样性指数分别为2.58和1.80, Chao丰富度指数分别为139.00和94.20(表 1); 而葡萄糖刺激下的藻际细菌群落组成和对照组一致性较高, 表明葡萄糖的添加几乎对藻际细菌群落无影响; 柠檬酸钠和乙酸钠均为有机酸式盐, 对藻际细菌群落影响几乎一致.表 1显示, A组和B组的Chao丰富度指数均高于CK组, A、B、C、D和E组的Shannon多样性指数均高于CK组, 表明在铜绿微囊藻培养液中添加蛋白胨和LB培养基可大幅提高藻际环境中细菌群落的多样性和丰富度.这可能是因为蛋白胨和LB培养基都含有细菌生长所需要的维生素和其他生长因子, 为细菌生长提供了营养.由于细菌生长周期比较短, 在外加营养刺激下, 细菌会在短时间内大量繁殖, 藻际细菌群落的多样性和丰富度也显著提高[20, 27].
表 1
(Table 1)

表 1 藻际细菌群落物种多样性和丰富度指数(属水平)
Table 1 Shannon and Chao index of bacteria in the phycosphereof Microcystis aeruginosa (on genus level)
| 项目 |
CK组 |
A组 |
B组 |
C组 |
D组 |
E组 |
| Shannon指数 |
1.39 |
1.80 |
2.58 |
1.63 |
1.58 |
1.42 |
| Chao指数 |
85.10 |
94.20 |
139.00 |
61.00 |
85.00 |
70.17
|
|
表 1 藻际细菌群落物种多样性和丰富度指数(属水平)
Table 1 Shannon and Chao index of bacteria in the phycosphereof Microcystis aeruginosa (on genus level)
|
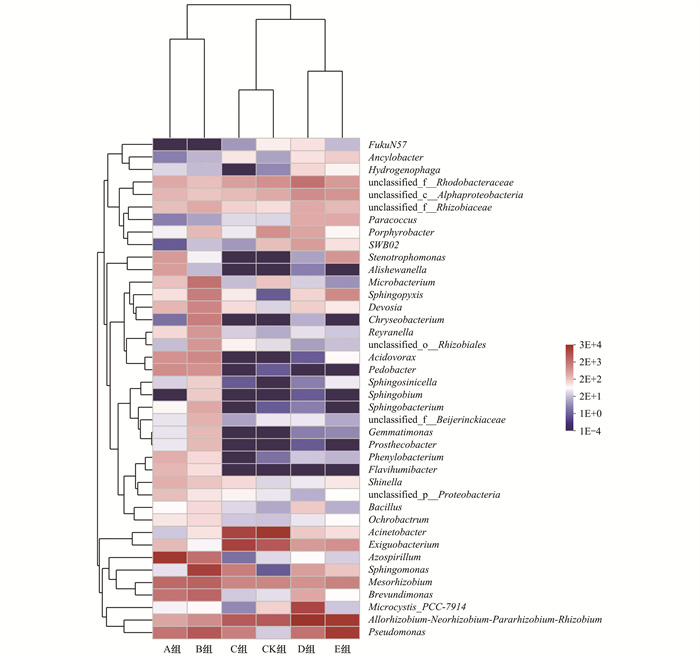
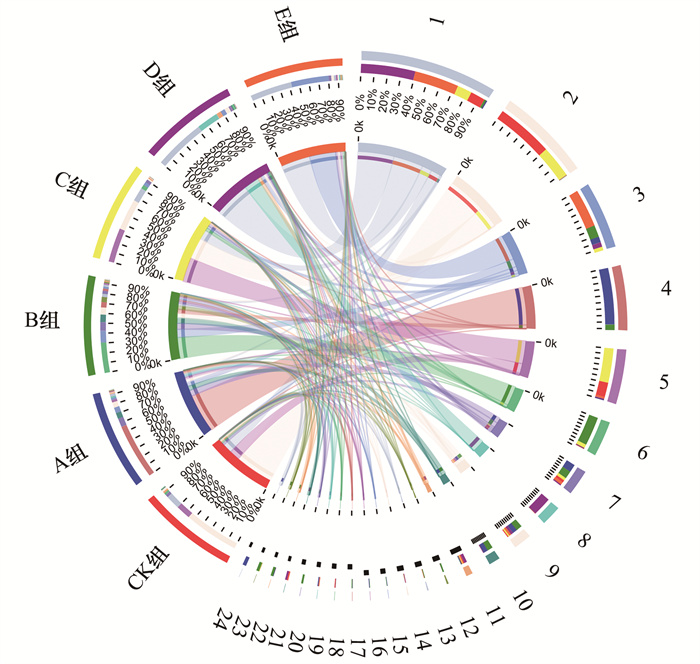
采用Circos表征各组藻际细菌的群落结构变化, 如图 4所示, 所有样本中分布最多的属是:Allorhizobium-Neorhizobium-Pararhizobium-Rhizobiu、Acinetobacter、Pseudomonas、Azospirillum和Exiguobacterium.A组和B组中分布最多的属分别是Azospirillum和Sphingomonas, 其次均是Mesorhizobium和Pseudomonas; C组和CK组中占比前3的属是Exiguobacterium、Acinetobacter和Allorhizobium-Neorhizobium-Pararhizobium-Rhizobiu; D组和E组中优势属均为Allorhizobium-Neorhizobium-Pararhizobium-Rhizobiu, 分别占比55%和45%.蛋白胨和LB培养基刺激铜绿微囊藻后, 藻际细菌群落中优势菌种变化显著, 且溶藻菌种(Pseudomonas)相对丰度显著增加[28~31], 因此, 在蓝藻暴发的水体中给予适当的营养刺激, 可一定程度上实现溶藻细菌原位抑藻.
2.3 营养物质对藻细胞活性氧、丙二醛和抗氧化酶水平的影响
2.3.1 活性氧(ROS)水平和丙二醛(MDA)含量
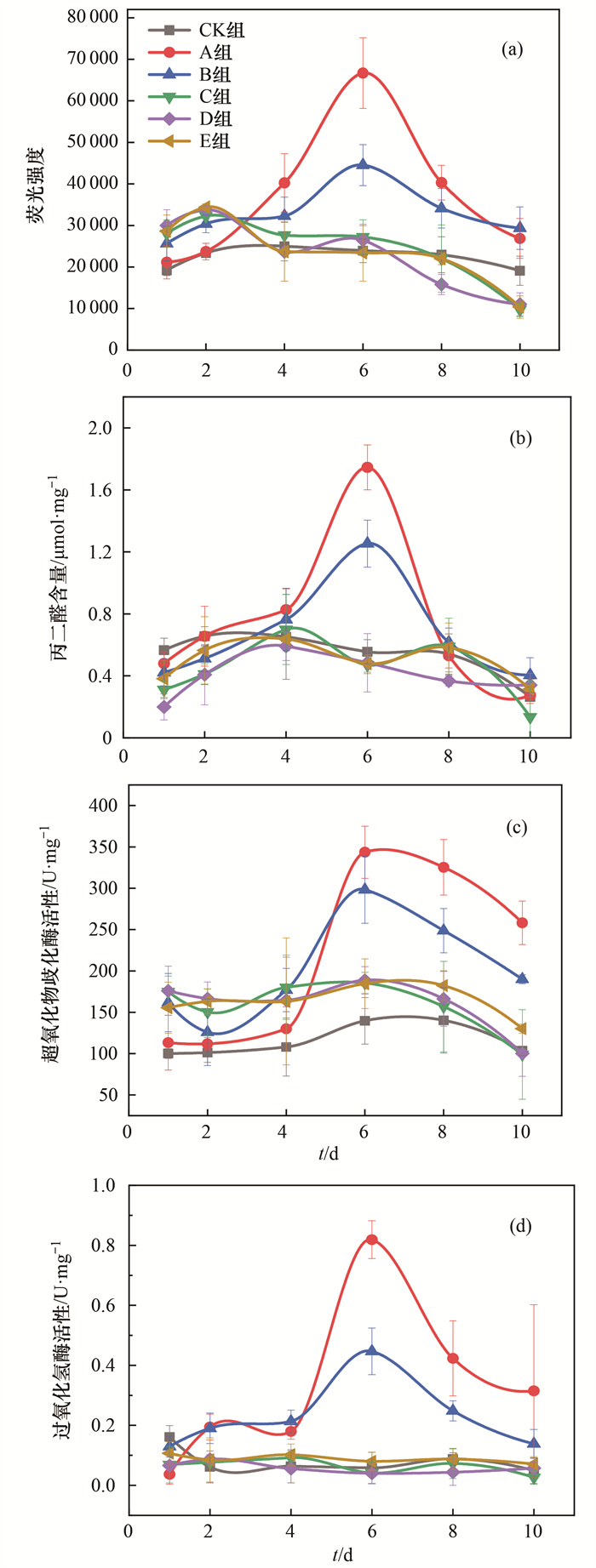
ROS是一种含有氧自由基的高活性化学物质, 对活细胞具有强毒性, 可导致细胞损伤或死亡; MDA是膜脂过氧化的产物, 可评价细胞的氧化损伤程度[32, 33].当铜绿微囊藻受到外界胁迫后, 藻细胞会发生氧化应激, ROS水平和MDA含量会急剧增加[34].将LB培养基、蛋白胨、葡萄糖、柠檬酸钠和乙酸钠分别添加到铜绿微囊藻培养液后, 整个实验过程中仅LB培养基和蛋白胨组的ROS水平有明显上升, 见图 5(a), 第6 d的荧光强度分别高达66 690和44 521; 而葡萄糖、柠檬酸钠和乙酸钠组的ROS水平一直较低且相对稳定, 与对照组相比荧光强度几乎一致, 均在10 366~28 420范围内波动.如图 5(b)所示, 不同营养物质添加到铜绿微囊藻后, 前4 d时, 随着时间的增加, MDA含量逐渐增多.第6 d时, LB培养基和蛋白胨组的MDA含量均达到最高值, 为对照组的3.18倍和2.27倍, 随后MDA含量骤降.而葡萄糖、柠檬酸钠和乙酸钠组的MDA含量始终保持在较低水平, 与对照组相近, 整体在0.13~0.59 μmol·mg-1蛋白质范围内波动.综上所述, LB培养基和蛋白胨可刺激铜绿微囊藻发生氧化应激, 诱导藻细胞膜脂过氧化, 遭受氧化损伤; 而葡萄糖、柠檬酸钠和乙酸钠均未对铜绿微囊藻造成明显的氧化损伤.
2.3.2 超氧化物歧化酶(SOD)和过氧化氢酶(CAT)活性
SOD和CAT均为抗氧化酶, SOD能够催化超氧化物阴离子(O2-)歧化生成H2O2, CAT可将H2O2分解为H2O, 从而使细胞氧化损伤有所缓解[35~37].如图 5(c)所示, LB培养基和蛋白胨添加后的第6 d, SOD活性(以蛋白质计, 下同)均大幅增加, 分别高达343.57 U·mg-1和298.22 U·mg-1, 为对照组的2.46倍和2.14倍.随后, SOD活性迅速下降, 第10 d分别降至291.09 U·mg-1和189.95 U·mg-1.整个实验期间对照组与葡萄糖、柠檬酸钠和乙酸钠组的SOD活性波动趋势相同, 均保持较低且稳定的水平.图 5(d)显示, 所有实验组CAT与SOD活性变化趋势相似, 仅LB培养基和蛋白胨组的CAT活性有显著提高.第6 d时, CAT活性达到最高值, 分别为0.82 U·mg-1和0.45 U·mg-1, 而第10 d的CAT活性骤降.综上表明, 在藻液中添加葡萄糖、柠檬酸钠和乙酸钠均未对铜绿微囊藻细胞的SOD和CAT活性产生明显影响; 而LB培养基和蛋白胨的添加可使藻细胞遭受氧化胁迫, 并激发自我防御系统以缓解氧化损伤, 使得SOD和CAT活性显著增加.随着持续胁迫, 抗氧化酶不足以清除过量ROS, 导致防御系统崩溃, 细胞损伤严重, 从而促使藻细胞死亡.
2.4 铜绿微囊藻细胞形态变化
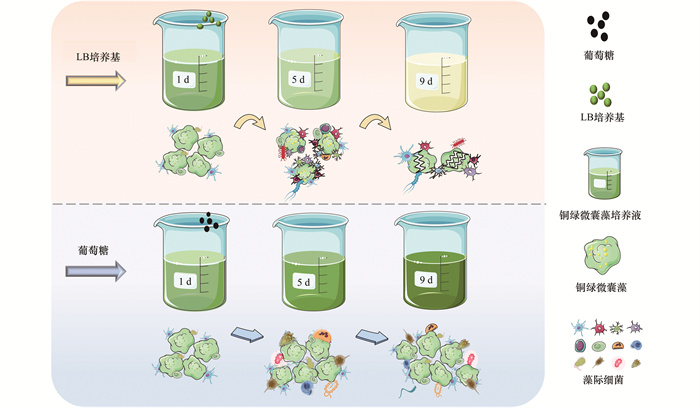
在LB培养基的刺激下, 铜绿微囊藻生长受到明显抑制.因此, 本研究观察了LB培养基胁迫下的铜绿微囊藻细胞的形态变化.图 6显示, LB培养基的添加促使藻细胞逐渐皱缩, 最终藻细胞破碎; 此外, 藻际环境中细菌群落结构变化明显, 细菌数量大幅增加.第1 d时, 藻细胞呈球形或近球形, 表面饱满, 结构完整, 藻际环境中细菌数量几乎为零; 第3 d时, 藻细胞略有皱缩, 附着在藻细胞上的细菌逐渐增多; 第5 d时, 藻细胞表面出现明显的内陷和萎缩等现象, 藻际细菌群落中优势菌种逐渐凸显, 细菌数量骤增; 第9 d时, 大部分藻细胞破碎, 细胞内含物流出, 藻细胞凋亡, 而藻际环境仍存在大量细菌.以上结果表明, 在LB培养基的胁迫下, 铜绿微囊藻细胞受到严重破坏, 藻际细菌显著增加, 初步推测这些细菌中可能存在溶藻菌.
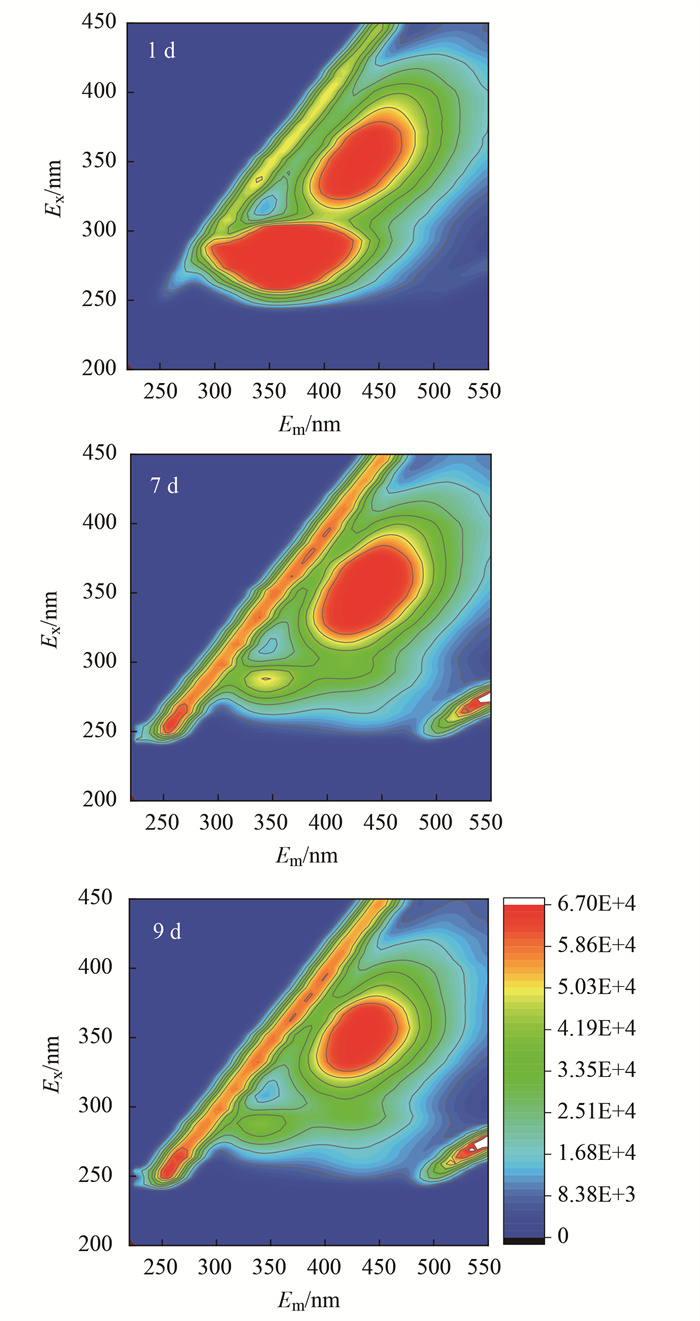
2.5 藻液中溶解性有机物(DOM)的变化
LB培养基营养丰富, 能够为细菌生长提供所需的碳源、氮源和无机盐等.因此, 对LB培养基刺激下的铜绿微囊藻胞外DOM组成变化进行分析.如图 7所示, 第1 d的光谱中出现2个较强的荧光峰, 激发和发射(Ex/Em)中心位置分别为350 nm/450 nm(FL-a)和280 nm/350 nm(FL-b).第7 d时, FL-a的荧光强度略有增强, 但FL-b的荧光强度大幅降低, 第9 d时几乎消失.FL-a为类腐殖质荧光峰, 一种难降解的DOM, 含有大量长链烷烃和少量芳香基和碳水化合物[38]; FL-b为类蛋白质色氨酸荧光峰, 可作为细菌生长所需的养分[39, 40].综上所述, LB培养基刺激藻液中类蛋白质迅速被微生物降解, 导致异养型藻附生菌大大增殖.
2.6 藻际细菌群落中溶藻菌的分离与鉴定
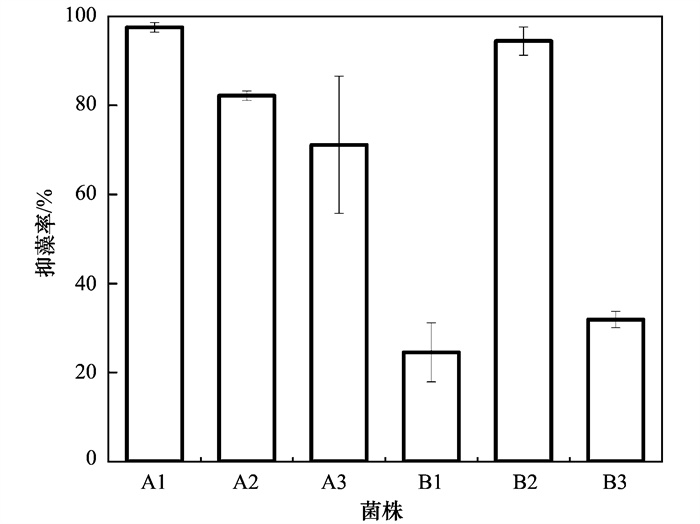
在铜绿微囊藻培养液中添加LB培养基和蛋白胨, 藻际环境中细菌激增, 并且藻细胞的生长受到强烈抑制作用.因此, 对其中的溶藻菌进行筛选, 并分离出6株具有明显抑藻效果的菌株.通过16S rRNA基因测序, 鉴定出3株溶藻菌为芽孢杆菌属(Bacillus sp.)、2株假单胞菌属(Pseudomonas sp.)和1株寡氧单胞菌属(Stenotrophomonas sp.).各菌株与铜绿微囊藻共培养后的抑藻情况如图 8所示, A1和B2抑藻效果显著且稳定, 抑藻率分别高达97.55%和94.48%, 分别为芽孢杆菌属和假单胞菌属.Wu等[41]也从富营养化严重的太湖分离出能够强效抑制铜绿微囊藻9110的溶藻菌Bacillus sp. strain S51107.本研究中, A1、B1和B3同为芽孢杆菌属, 但B1和B3抑藻率分别仅为24.54%和31.90%, 抑藻效果远不如A1, 表明同一种属细菌在不同环境中, 功能特性差异显著.有报道称, 原位环境可能会调节细菌自身的功能表达[42, 43].通常溶藻菌可裂解藻细胞, 但当有机营养发生变化时, 溶藻菌和藻细胞可能演变为共生关系[44].此外, 许多溶藻菌需要达到特定浓度才可激发其溶藻活性[45, 46].因此, 环境中现存的溶藻菌不一定意味着目前可以溶藻.综上所述, 适当的营养刺激会激发藻际环境中细菌的溶藻活性, 大量增殖的溶藻菌可有效抑制藻细胞生长(图 9), 这为微生物防控蓝藻水华提供了一定的理论参考.
3 结论
(1) 营养物质是影响铜绿微囊藻生长的关键因子.LB培养基和蛋白胨对铜绿微囊藻的生长具有强烈的抑制作用, 而葡萄糖、醋酸钠和乙酸钠则对铜绿微囊藻的生长具有显著的促进作用.
(2) 原位营养刺激下铜绿微囊藻藻际细菌群落结构变化明显, 物种丰富度和群落多样性显著升高.藻液中添加蛋白胨后Chao丰富度指数从85.10增加到139.00, Shannon多样性指数从1.39增加到2.58.
(3) 铜绿微囊藻细胞受LB培养基和蛋白胨胁迫后, 氧化损伤严重, 随后自身抗氧化酶系统被激活.藻细胞ROS水平和MDA含量显著升高, 抗氧化酶SOD和CAT活性先升高后降低, 最终藻细胞防御系统受损, 藻细胞破碎.
(4) 营养刺激下的藻际环境会激发溶藻菌的溶藻活性.添加LB培养基和蛋白胨后, 大量溶藻菌快速生长繁殖, 溶藻活性增强, 所筛溶藻菌A1和B2的抑藻率分别高达97.55%和94.48%.
 2022, Vol. 43
2022, Vol. 43