 2022, Vol. 43
2022, Vol. 43

 , ZHAO Lu , JI Lu-lu , YANG Song-qi , ZHENG Bao-hai , DU Yu-xin , LI Jia-xin , LI Yu-xin , SHI Jun-qiong , WU Zhong-xing
, ZHAO Lu , JI Lu-lu , YANG Song-qi , ZHENG Bao-hai , DU Yu-xin , LI Jia-xin , LI Yu-xin , SHI Jun-qiong , WU Zhong-xing
近年来, 随着气候变暖、水体富营养化程度加剧和二氧化碳浓度升高, 全球范围内蓝藻水华发生的频率、强度和持续时间均呈现增加态势[1~3].蓝藻水华的发生不仅降低了水生态系统生物多样性, 而且对生态服务功能包括饮用水、灌溉、渔业、航运和娱乐用途等产生负面效应, 造成巨大经济损失[1, 4, 5].因此, 蓝藻水华的形成特征和机制[6]、蓝藻毒素的产生[7]和蓝藻与其他物种的相互作用机制[8]等方面的研究已引起学者们广泛的关注.
有研究表明, 蓝藻水华的暴发是复杂的事件, 不同种类的水华蓝藻往往在水华频发的水体中一同发现, 在不同区域的水体中占主导地位, 并呈现出时间序列上的优势种类演替[9, 10].Soares等[11]在热带水库中观察到在持久的蓝藻水华过程中优势种从固氮蓝藻(卷曲长孢藻和拉氏拟柱孢藻)向非固氮蓝藻(铜绿微囊藻)的转变.Wu等[12]发现滇池蓝藻水华过程中出现了束丝藻与微囊藻的演替.有研究发现固氮蓝藻可以将大气中的氮(N2)转化为可利用的氨, 使其在缺乏氮的水域中比非固氮蓝藻和其他真核浮游植物具有竞争优势, 当水体中其他必需营养素充足时就容易形成水华[1], 因此氮的变化可能导致固氮蓝藻与非固氮藻类间的演替并决定优势种类的形成[13, 14].然而, Rigosi等[15]指出营养物质供应增加和温度升高对促进蓝藻的相对重要性取决于具体的种类, 例如, 长孢藻对营养物质更敏感, 而微囊藻对温度更加敏感, 表明蓝藻演替是由多种环境因素共同驱动的, 而非单一变量主导.
目前, 对水华蓝藻的研究往往只集中在一个优势种的变化及影响因素的分析.然而, 水华蓝藻作为一种多样化和异质性的组合, 不同种类在生理上对环境条件的反应各不相同, 这导致水华期间浮游植物优势种演替过程复杂多变, 且不同种群占据优势地位的因素难以确定[16].因此, 为了解形成水华的蓝藻之间的演替特征及其对浮游植物群落的影响, 并揭示其潜在的驱动因子, 本文选取南彭水库作为研究对象, 通过对其浮游植物群落结构和理化环境因子进行调查, 分析蓝藻水华过程中浮游植物群落结构变化及水华蓝藻优势种演替, 并探讨驱动水华蓝藻优势演替的生态因子, 旨在为水库蓝藻水华形成机制和富营养化治理提供科学依据.
1 材料与方法 1.1 研究区域概况与样品采集南彭水库(29°19′N, 106°41′E)是在长江一级支流花溪河上游筑坝形成的河道型水库, 位于重庆市巴南区, 属亚热带湿润季风气候, 四季分明, 春早秋迟, 年平均气温18.7℃, 年降水量约1 000 mm.南彭水库为重庆市内重要的国家一级保护水源地, 库区水域面积约为1.3 km2, 库容量1.33×107 m3.本研究根据南彭水库形态特征, 从入库口到坝前共设置S1~S5这5个监测点(图 1), 于2018年4月至2018年9月的每月中旬进行水质调查并采集水样.鉴于采样区域为河道型水库, 采样时依照河流样点设置方法, 各样点取主河道左中右表层0.5 m深的3处水样进行等量混合, 其中1 L水样现场滴加15 mL鲁格试剂固定保存用于浮游植物分析, 其余放入冰盒中于当天带回实验室做进一步分析.

|
图 1 南彭水库采样点分布示意 Fig. 1 Distributions of sampling sites in the Nanpeng Reservoir |
浮游植物样品带回实验室后于暗处静置2d左右, 虹吸浓缩至30 mL后, 补加1 mL 40%甲醛溶液长期保存.浮游植物的种类鉴定和细胞计数采用面积为20 mm×20 mm, 容量为0.1 mL的浮游计数框, 于10×40倍光学显微镜(Nikon E-Ci)下进行, 具体的种类鉴定与计数方法参照文献[17~19].
1.3 水体理化指标的测定表层水体的水温(WT)、pH、电导率(EC)和溶氧(DO)使用多参数水质分析仪DS5/DS5X(Hydrolab, USA)现场测定, 水体透明度(SD)用黑白盘测定.总磷(TP)和正磷酸盐(PO43--P)用钼酸铵分光光度法测定, 总氮(TN)和硝态氮(NO3--N)用紫外分光光度法测定, 高锰酸盐指数使用酸性滴定法测定, 叶绿素浓度(Chl-a)使用丙酮浸提分光光度法测定, 室内分析具体参照文献[20]的方法.
1.4 数据处理与分析以浮游植物群落资源利用效率(resource use efficiency, RUE)作为评价单位养分影响生物量产量的生态过程指标[15], 可以通过每单位的资源量所产生的浮游植物生物量来衡量[21].因此本研究通过以下公式计算浮游植物的资源利用效率:
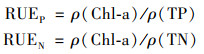
|
式中, RUEP和RUEN分别为浮游植物磷的利用效率和氮的利用效率; ρ(Chl-a)为浮游植物生物量测量值(mg ·L-1); ρ(TP)和ρ(TN)分别为总磷和总氮的浓度(mg ·L-1).
浮游植物的Manaughton优势度指数(Y)不小于0.02时, 可认定该物种为优势种[22], 优势度指数计算公式为:
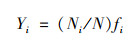
|
式中, Ni为第i种浮游植物的丰度, N为浮游植物总丰度, fi为第i种在各采样点出现的频率.
物种丰富度(S)、Shannon-Wiener多样性指数(H)和均匀度(J)等指标可以衡量浮游植物群落物种多样性, 其中Shannon多样性指数(H) 的计算公式为:
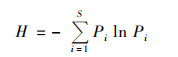
|
Pielou均匀度指数(J) 的计算公式为:
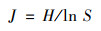
|
式中, Pi为物种i的相对丰度.
南彭水库各月份间浮游植物群落多样性指数及水体理化因子的差异采用单因素方差分析(one-way analysis of variance, ANOVA)进行并作LSD事后检验, 对于不满足ANOVA方法条件的因子采用Wilcoxon非参数检验.不同月份间周边浮游植物群落的差异采用置换多元方差分析(permutational multivariate analysis of variance, PERMANOVA)进行显著性检验.
为分析蓝藻水华优势种是否以及在多大程度上影响周边浮游植物群落的结构组成, 使用非度量多维标度法(non-metric multidimensional scaling, NMDS)减少群落数据的冗余性, 并以此被动拟合水华优势种丰度, 这一过程中外部变量与降维排序空间中所承担方差载体的关系是后计算的, 不影响群落排序的计算过程, 二者关联强度以R2来衡量[21].此外, 采用方差分解方法(variance partitioning analysis, VPA) 来评估水华优势蓝藻和水体环境因子对周边浮游植物群落组成和结构变化的相对贡献.
为确定水体环境因子与水华优势种丰度之间的联系, 本研究利用广义加性模型(generalized additive model, GAM)来探索响应变量与自变量之间线性或者非线性的关系.GAM先前已被证明可用于阐明人类发病率和气候条件之间的非线性统计关系[23].在具体分析时, 由于GAM中具有较低广义交叉验证值的模型具有更高的预测能力[23], 因此在对不同参数比较后选用最大自由度为3的样条函数作为自变量的局部平滑方法, 并对水华蓝藻丰度数据进行lg(x+1)转换.
所有数据整理采用Excel 2019, 具体分析与统计使用R 4.0.3中stats、vegan和mgcv等程序包进行, 并用ggplot2程序包作图.
2 结果与分析 2.1 水体理化特征在调查的6个月中, 所有环境因子在时间序列上均呈现显著差异(P < 0.05, 表 1).水体温度从4月逐步上升直到8月达到峰值, 随后下降; pH值的变化与水温类似, 峰值出现在7月; EC在夏季显著低于其他时间; DO在5月达到最低值、在7月达到最高值; SD随时间的变化与pH的相反, 7月平均值最低.此外, 水体氮磷营养元素指标普遍偏高, 尤其是TN, 除7月外样点浓度平均值都在2 mg ·L-1以上; 而样点ρ(TP)平均值在6月最低, 为0.05 mg ·L-1, 处于较高水平.水体中5月浮游植物磷的平均资源利用效率(RUEP)较其他月份显著更低, 仅为0.26±0.11; 而浮游植物氮的平均资源利用效率(RUEN)在7月最高, 为0.023.
|
|
表 1 南彭水库环境因子月份变化1) Table 1 Environmental factors of Nanpeng Reservoir changes in different months |
2.2 浮游植物群落结构与优势种
调查期间, 南彭水库共检出浮游植物8门59属108种.其中绿藻种类最多, 为26属50种, 占比46.3%, 其次为蓝藻(16.7%)和硅藻(13.0%)分别为12属18种和9属14种, 随后裸藻5属11种和甲藻3属10种占比分别为10.2%和9.3%, 而其余3门种类较少, 包括隐藻2属3种, 金藻和黄藻各1种[图 2(a)].尽管蓝藻种类相对绿藻较少, 但藻密度上占据明显优势, 平均值达7.84×107个·L-1, 占浮游植物总密度的94.1%[图 2(b)].从浮游植物密度组成时间变化来看, 蓝藻在各个月份中均占据优势地位, 并且在5月和7月中出现两个峰值, 分别为(2.01±0.71)×108个·L-1和(1.13±0.18)×108个·L-1, 与之相比4月最低, 仅为(6.17±1.07)×106个·L-1.

|
图 2 浮游植物种类和藻密度组成 Fig. 2 Compositions of phytoplankton species and density |
各月份中共发现4门13个优势种(Y ≥ 0.02), 包括蓝藻7种、绿藻2种、硅藻2种和隐藻2种(表 2).南彭水库优势种在不同月份各不相同, 既有交叉又有演替, 但假鱼腥藻在各月均以优势种或亚优势种出现, 并且在4月和5月占据最高优势度(0.592和0.899), 而在6~9月, 拉氏拟柱孢藻的优势度最高(优势度分别为0.518、0.912、0.424和0.441).以整个调查期数据分析, 结果显示优势种为3种丝状蓝藻, 其中水华束丝藻作为6月、8月和9月的优势种替代了之前的绿藻(狭形纤维藻和四尾栅藻)和硅藻(尖针杆藻和披针曲壳藻), 但其优势度较低.
|
|
表 2 浮游植物优势种优势度指数(Y) 1) Table 2 Dominance index (Y) of dominant phytoplankton species |
2.3 水华优势蓝藻对浮游植物群落的影响
整个研究期间, 各月份浮游植物群落组成的空间差异都不显著(P>0.05), 各样点间的水华优势蓝藻(拉氏拟柱孢藻和假鱼腥藻)密度与组成的变化趋势相一致(图 3).然而, 在此次蓝藻水华过程中发生了明显的优势种演替现象, 在5月发生了以假鱼腥藻为主的蓝藻水华, 在6月则呈现出假鱼腥藻和拟柱孢藻共同主导占优的现象, 而在7月拟柱孢藻水华暴发, 藻密度达到最大值, 此后蓝藻水华开始消退, 并在8月和9月呈现出多种蓝藻共存但仍以拟柱孢藻为主的现象(图 3和表 2).

|
图 3 水华蓝藻优势种藻密度时空分布 Fig. 3 Temporal and spatial distribution of dominant cyanobacteria in algal bloom |
蓝藻水华对浮游植物群落的影响在生物多样性层面有所体现.南彭水库浮游植物群落物种丰富度在夏季7月藻密度达到最高峰时最低, 各样点丰富度平均值仅为11.6, 相反在9月水华末期时物种丰富度最高, 平均值达到27[图 4(a)].此外, 各样点Shannon多样性指数平均值在5月和7月较低, 分别为0.79和0.57, 与其余4个月份有显著性差异(P < 0.05) [图 4(b)]. Pielou均匀度指数的变化趋势与Shannon多样性指数基本一致, 同样在5月和7月较其他月份显著更低(P < 0.05), 样点平均值都仅为0.24[图 4(c)].

|
不同小写字母表示具有显著性差异(P < 0.05) 图 4 南彭水库浮游植物的多样性指数 Fig. 4 Diversity index of phytoplankton in Nanpeng Reservoir |
在非约束群落排序分析中, NMDS二维平面上各月样本具有明显的区分(PERMANOVA, F=7.86, P < 0.001, 图 5).与假鱼腥藻(R2=0.48, P=0.002)相比, 拟柱孢藻(R2=0.58, P=0.002)与周边环境浮游植物群落的关联程度更高.而VPA结果表明, 相比于环境变量对浮游植物群落组成变化的解释(47.5%), 两种水华蓝藻假鱼腥藻和拟柱孢藻的相对贡献相差不大, 分别解释了12.74%和12.04%的方差(图 6).

|
2D stress表示二维排序空间最终应力函数值 图 5 非蓝藻水华优势种浮游植物群落NMDS图 Fig. 5 NMDS ordination plots of the non-dominant cyanobacteria bloom specialist in phytoplankton community |

|
图 6 拟柱孢藻、假鱼腥藻与环境因子对周边浮游植物群落相对贡献的方差分解分析 Fig. 6 Variation partitioning analysis of the relative contributions of C. raciborskii, Pseudoanabaena sp., and environmental factors to ambient phytoplankton community |
为探究水华优势蓝藻沿不同类型环境梯度下的变化, 以蓝藻丰度的对数值为响应变量, 水体理化因子作为自变量构建广义可加模型. 结果发现拟柱孢藻丰度对数受到WT、pH、SD、RUEP和RUEN的显著影响(P < 0.05), 模型的整体解释率R2为0.957; 而假鱼腥藻丰度的对数受到EC、DO和高锰酸盐指数的显著影响(P < 0.05), 模型的整体解释率R2为0.856(表 3).显著环境变量对蓝藻丰度影响的平滑回归曲线表明, 拟柱孢藻丰度的对数随水温增加而增加, 在28℃左右存在一个拐点; 随RUEP增加而降低, 在0.5左右趋于平稳; 在pH、SD和RUEN的环境梯度下受线性影响, 其中与pH和RUEN为正相关, 与SD为负相关(图 7).而假鱼腥藻丰度的对数随EC和DO的增加而减少, 随高锰酸盐指数的增加而增加, 其中EC在大于260时影响更为强烈(图 7).
|
|
表 3 GAM中环境变量对拟柱孢藻或假鱼腥藻丰度对数的响应结果1) Table 3 Response of environmental variables to the abundance of C. raciborskii or Pseudoanabaena sp. in the GAM |

|
s()为经样条函数变化后的自变量, 阴影部分为95%的置信区间 图 7 显著环境变量与拟柱孢藻或假鱼腥藻丰度的广义可加模型分析图 Fig. 7 Relationship between significant environmental variables and the abundance of C. raciborskii or Pseudoanabaena sp. based on the generalized additive models |
南彭水库在4~9月调查期间的浮游植物群落组成与西南地区其他水库相似, 以绿藻、蓝藻和硅藻为主(图 2)[24~26].河流筑坝形成水库之后, 由激流环境转变为静水环境, 水体流速减缓、滞留时间变长并截留营养物质, 有利于浮游植物的生存与繁殖[27], 这解释了南彭水库浮游植物群落以蓝、绿藻等富营养型静水种为主的现象.尽管蓝藻种类较绿藻少, 但就丰度来看, 蓝藻占据了绝对优势.Paerl等[28, 29]指出蓝藻作为原核生物, 在较高的温度下具有更高的生长速率, 这使得在营养丰富的条件下蓝藻与其他真核初级生产者(包括硅藻、绿藻和甲藻等)具有明显的竞争优势.在调查期间, 南彭水库的水温在22~32℃之间, 水体TN和TP浓度远超水体富营养化的临界浓度[ρ(TN)为0.2 mg ·L-1, ρ(TP)为0.02 mg ·L-1], 这些条件为蓝藻的生长并占据优势地位提供了良好的外部条件.
优势种组成可以在一定程度上反映研究水库的污染现状[25], 尽管南彭水库浮游植物优势种种类数较多, 但其优势度在不同月份存在差异, 大部分种类仅在某月份成为优势种属(表 2).而蓝藻作为最典型的富营养水体优势类群, 在南彭水库中优势种类最多, 表明水体污染现状不容乐观.朱广伟等[30]调查发现硅藻在气温16℃以上时, 生物量开始快速增加, 当气温达到26℃之后便开始下降, 并且相比于尖针杆藻, 曲壳藻的最佳生长温度更高.这符合本研究中尖针杆藻在4月成为优势种, 披针曲壳藻在5月成为优势种的现象.而当9月水体温度和光照减弱时, 隐藻由于独特的辅助色素能用于光合作用, 并能凭借鞭毛调节自身在水体中位置的特性所具备竞争优势[31, 32], 成为优势种类.
蓝藻优势种类的大量生长将通过种间相互作用(资源竞争和化感作用等)直接抑制其他浮游植物种类的生长, 当增殖暴发形成水华时将引起水质的改变从而间接影响浮游植物群落结构[6, 33, 34].在本研究中, 蓝藻密度在5月和7月形成了两个峰值, 此时浮游植物群落的Shannon多样性指数和Pielou均匀度指数与其他月份相比都显著更低[图 4(b)和图 4(c)], 表明蓝藻水华暴发时将降低浮游植物群落结构稳定性, 使得浮游植物群落结构趋于简单, 这与陈莎等[24]的研究结果相一致, 在夏季较高温度和光照条件下, 蓝藻优势种类的大量生长对其他藻类形成抑制, 导致生物多样性降低.
3.2 不同水华蓝藻优势种对浮游植物群落的影响调查期间所发现的3种优势种均为丝状蓝藻, 并远超过蓝藻水华阈值(107个·L-1)[35], 与同气候地区调查结果一致, 表明丝状蓝藻的优势普遍存在[25, 36].在5月和7月水华暴发时假鱼腥藻和拟柱孢藻分别在浮游植物中占据绝对优势地位(图 3), 两种蓝藻在大量生长繁殖时都对周边浮游植物群落产生了显著的影响, 相较于假鱼腥藻, 拟柱孢藻与周边群落的关联程度更高(图 5). Figueredo等[37]的研究证实拟柱孢藻藻株的渗出物对不同藻类的光合活性具有较强的抑制作用.化感作用被认为在湖泊浮游植物结构和演替中起重要作用, 并与水生生境的生态相互作用紧密联系[38], 主要原因在于由于本地物种缺乏与入侵物种长期共存, 并且没有进化到能够忍受入侵物种产生的化感物质[39].有研究表明拟柱孢藻作为一种全球性的入侵蓝藻与其潜在的产毒能力[40], 可能改变物种优势结构、养分动态和初级生产力水平, 从而严重影响被入侵的群落和物种多样性, 相较于假鱼腥藻这种广布性物种, 拟柱孢藻在大量增殖形成水华时对被入侵群落的结构组成将产生更大的影响[41].因此, 尽管在两种藻华暴发期间浮游植物群落的Shannon指数和Pielou指数无显著性差异, 但丰富度存在显著差异, 且拟柱孢藻水华期间更低(图 4).
此外, 调查期间各月份的浮游植物群落结构组成具有显著性差异, VPA结果表明造成这种差异绝大部分是由于环境条件差异所导致的, 而水华蓝藻种类仅能解释较少的一部分方差(图 6).但这并不能说明水华优势种对周边群落的影响不大, 相反优势蓝藻形成水华后会对水环境产生显著改变.藻体的大量聚集降低了水体透明度, 同时死亡的藻体被微生物分解诱发的缺氧条件等, 都间接影响其它浮游植物的生长繁殖[6].观察到7月拟柱孢藻水华时透明度降至最低, 而pH和DO处于较高水平(表 2), 这与其他地区蓝藻水华发生时的环境特征相一致[42, 43].相反, 5月假鱼腥藻水华时DO处于较低水平, 并且GAM结果发现假鱼腥藻丰度的对数随DO增加而减少(图 7).有研究表明DO在蓝藻水华暴发期间往往明显升高并达到峰值, 但后期常常降至同期最低点[43, 44].因此, 调查发现7月的藻密度相较于5月更高[图 2(b)], 推测在5月假鱼腥藻水华已开始衰退, 而在7月拟柱孢藻水华正处于暴发峰值, 假鱼腥藻的优势地位正逐渐被拟柱孢藻所取代.
3.3 影响两种水华蓝藻优势种演替的环境因素在蓝藻水华频繁发生的湖泊中不同优势种类竞争并经历生态演替, 蓝藻水华形成种的相对成功是复杂和协同环境因素的结果, 而不是单一的支配变量, 重要的是要探讨形成水华类群的外部驱动因素[45].在淡水生态系统中, 最普遍的水华蓝藻模式就是固氮蓝藻和非固氮蓝藻之间主导地位的确定与转换, 而温度与营养物质被证明是关键的影响因素[36, 45, 46].传统上认为当N缺乏时, 固氮蓝藻将占据优势地位, 但在本研究中无论是固氮的拟柱孢藻还是非固氮的假鱼腥藻都不受TN和NO3--N梯度的影响(表 3).考虑到N和P等营养物质对浮游植物群落具有系统性的影响, 因此浮游植物群落的资源利用效率(RUE)相比水体中的N和P浓度更能体现营养的限制情况[21].
陶敏等[25]对四川丘陵区水库的浮游植物群落优势种研究中发现, 假鱼腥藻全年都占据优势地位, 生态响应速率高, 相比其他优势蓝藻包括拟柱孢藻是相对泛化的种群, 处于不断掠夺其他物种资源的扩张状态.本研究发现在5月较低的氮磷比和RUEP(表 1), 分别表明此时水体中P相对丰富, 浮游植物群落对P的竞争利用不激烈.而南彭水库作为富营养的水体, 尤其是长期处于高氮磷比的情况下, 在P得到满足时, 长期适应本地环境的假鱼腥藻相较于拟柱孢藻更具有竞争优势.
从RUEP响应拟柱孢藻丰度对数的平滑回归曲线可以看出, 当RUEP大于0.5之后, 拟柱孢藻不再受到RUEP的负面影响(图 7), 表明在浮游植物群落对P的竞争利用较为激烈时, 水体中的P成为其他浮游植物的限制因素, 相反由于拟柱孢藻在P限制水体中具有出色的磷吸收和利用能力[47, 48], 使其更具竞争优势, 这也解释了拟柱孢藻为何能够在6~9月RUEP较高时成为主要的优势种(表 1和表 2).而7月拟柱孢藻水华暴发时观察到的RUEN峰值(表 1)以及拟柱孢藻丰度的对数随RUEN增加而增加的现象(图 7), 都表明具有固氮能力的拟柱孢藻在N相对限制的情况下具有竞争优势.此外, 固氮作用作为高昂成本的生物反应, 与其他酶催化过程类似, 在有可利用氮源(如硝酸盐和铵盐)时会得到抑制, 而较高的温度下固氮作用将会增强[49].因此, 7月较高的温度将有利于起源于热带地区的拟柱孢藻形成水华[40].
4 结论(1) 调查期间共检出浮游植物8门59属108种, 蓝藻在丰度和优势种组成上占据绝对优势地位, 5~7月水华优势种由假鱼腥藻向拟柱孢藻演替, 生物多样性较低.
(2) 水华蓝藻主要通过改变水环境特征来间接影响周边浮游植物群落, 相比本地的假鱼腥藻, 入侵蓝藻-拟柱孢藻对水生生态系统的影响更强烈.
(3) 在N缺乏和温度升高的协同影响作用下, 具有固氮能力的拟柱孢藻取代假鱼腥藻成为蓝藻水华的优势类群.
| [1] | Huisman J, Codd G A, Paerl H W, et al. Cyanobacterial blooms[J]. Nature Reviews Microbiology, 2018, 16(8): 471-483. DOI:10.1038/s41579-018-0040-1 |
| [2] | O'Neil J M, Davis T W, Burford M A, et al. The rise of harmful cyanobacteria blooms: the potential roles of eutrophication and climate change[J]. Harmful Algae, 2012, 14: 313-334. DOI:10.1016/j.hal.2011.10.027 |
| [3] | Paerl H W, Paul V J. Climate change: links to global expansion of harmful cyanobacteria[J]. Water Research, 2012, 46(5): 1349-1363. DOI:10.1016/j.watres.2011.08.002 |
| [4] |
马健荣, 邓建明, 秦伯强, 等. 湖泊蓝藻水华发生机理研究进展[J]. 生态学报, 2013, 33(10): 3020-3030. Ma J R, Deng J M, Qin B Q, et al. Progress and prospects on cyanobacteria bloom-forming mechanism in lakes[J]. Acta Ecologica Sinica, 2013, 33(10): 3020-3030. |
| [5] |
秦伯强, 王小冬, 汤祥明, 等. 太湖富营养化与蓝藻水华引起的饮用水危机——原因与对策[J]. 地球科学进展, 2007, 22(9): 896-906. Qin B Q, Wang X D, Tang X M, et al. Drinking water crisis caused by eutrophication and cyanobacterial bloom in Lake Taihu: cause and measurement[J]. Advances in Earth Science, 2007, 22(9): 896-906. |
| [6] | Paerl H W, Otten T G. Harmful cyanobacterial blooms: causes, consequences, and controls[J]. Microbial Ecology, 2013, 65(4): 995-1010. DOI:10.1007/s00248-012-0159-y |
| [7] | Merel S, Walker D, Chicana R, et al. State of knowledge and concerns on cyanobacterial blooms and cyanotoxins[J]. Environment International, 2013, 59: 303-327. DOI:10.1016/j.envint.2013.06.013 |
| [8] |
彭宇科, 路俊玲, 陈慧萍, 等. 蓝藻水华形成过程对氮磷转化功能细菌群的影响[J]. 环境科学, 2018, 39(11): 4938-4945. Peng Y K, Lu J L, Chen H P, et al. Dynamic changes of nitrogen-transforming and phosphorus-accumulating bacteria along with the formation of cyanobacterial blooms[J]. Environmental Science, 2018, 39(11): 4938-4945. |
| [9] | Zhang M, Zhang Y C, Yang Z, et al. Spatial and seasonal shifts in bloom-forming cyanobacteria in Lake Chaohu: patterns and driving factors[J]. Phycological Research, 2016, 64(1): 44-55. DOI:10.1111/pre.12112 |
| [10] |
贾晓会, 施定基, 史绵红, 等. 巢湖蓝藻水华形成原因探索及"优势种光合假说"[J]. 生态学报, 2011, 31(11): 2968-2977. Jia X H, Shi D J, Shi M H, et al. Formation of cyanobacterial blooms in Lake Chaohu and the photosynthesis of dominant species hypothesis[J]. Acta Ecologica Sinica, 2011, 31(11): 2968-2977. |
| [11] | Soares M C S, de A Rocha M I, Marinho M M, et al. Changes in species composition during annual cyanobacterial dominance in a tropical reservoir: physical factors, nutrients and grazing effects[J]. Aquatic Microbial Ecology, 2009, 57(2): 137-149. |
| [12] | Wu Y L, Li L, Zheng L L, et al. Patterns of succession between bloom-forming cyanobacteria Aphanizomenon flos-aquae and Microcystis and related environmental factors in large, shallow Dianchi Lake, China[J]. Hydrobiologia, 2016, 765(1): 1-13. DOI:10.1007/s10750-015-2392-0 |
| [13] | Paterson M J, Schindler D W, Hecky R E, et al. Comment: lake 227 shows clearly that controlling inputs of nitrogen will not reduce or prevent eutrophication of lakes[J]. Limnology and Oceanography, 2011, 56(4): 1545-1547. DOI:10.4319/lo.2011.56.4.1545 |
| [14] | Schindler D W, Hecky R E, Findlay D L, et al. Eutrophication of lakes cannot be controlled by reducing nitrogen input: results of a 37-year whole-ecosystem experiment[J]. Proceedings of the National Academy of Sciences of the United States of America, 2008, 105(32): 11254-11258. DOI:10.1073/pnas.0805108105 |
| [15] | Rigosi A, Carey C C, Ibelings B W, et al. The interaction between climate warming and eutrophication to promote cyanobacteria is dependent on trophic state and varies among taxa[J]. Limnology and Oceanography, 2014, 59(1): 99-114. DOI:10.4319/lo.2014.59.1.0099 |
| [16] | Dokulil M T, Teubner K. Cyanobacterial dominance in lakes[J]. Hydrobiologia, 2000, 438(1-3): 1-12. |
| [17] | 胡鸿钧, 魏印心. 中国淡水藻类——系统、分类及生态[M]. 北京: 科学出版社, 2006. |
| [18] | 章宗涉, 黄祥飞. 淡水浮游生物研究方法[M]. 北京: 科学出版社, 1991. |
| [19] | 中国科学院中国孢子植物志编辑委员会. 中国淡水藻志-第十二卷-硅藻门, 异极藻科[M]. 北京: 科学出版社, 2004. |
| [20] | 国家环境保护总局. 水和废水监测分析方法[M]. 北京: 中国环境科学出版社, 2002. |
| [21] | Olli K, Klais R, Tamminen T. Rehabilitating the cyanobacteria-niche partitioning, resource use efficiency and phytoplankton community structure during diazotrophic cyanobacterial blooms[J]. Journal of Ecology, 2015, 103(5): 1153-1164. DOI:10.1111/1365-2745.12437 |
| [22] | Lampitt R S, Wishner K F, Turley C M, et al. Marine snow studies in the northeast Atlantic Ocean: distribution, composition and role as a food source for migrating plankton[J]. Marine Biology, 1993, 116(4): 689-702. DOI:10.1007/BF00355486 |
| [23] | Xu L, Stige L C, Chan K S, et al. Climate variation drives dengue dynamics[J]. Proceedings of the National Academy of Sciences of the United States of America, 2017, 114(1): 113-118. DOI:10.1073/pnas.1618558114 |
| [24] |
陈莎, 谢青, 付梅, 等. 三峡库区典型支流水库浮游动植物群落结构特征及其与环境因子的关系[J]. 环境科学, 2021, 42(5): 2303-2312. Chen S, Xie Q, Fu M, et al. Structural characteristics of zooplankton and phytoplankton communities and its relationship with environmental factors in a typical tributary reservoir in the Three Gorges Reservoir region[J]. Environmental Science, 2021, 42(5): 2303-2312. |
| [25] |
陶敏, 岳兴建, 岳珊, 等. 四川丘陵区水库浮游植物群落结构与蓝藻水华风险——基于优势种生态位与种间联结研究[J]. 生态学报, 2021, 41(23): 9457-9469. Tao M, Yue X J, Yue S, et al. Phytoplankton community structure and cyanobacteria bloom risk of reservoirs in hilly regions of Sichuan Province based on dominant species niche and interspecific association[J]. Acta Ecologica Sinica, 2021, 41(23): 9457-9469. |
| [26] |
张耀文, 李洪, 李嘉, 等. 西南山区典型河道型水库藻类功能群时空演替特征及其影响因素: 以紫坪铺水库为例[J]. 环境科学, 2018, 39(6): 2680-2687. Zhang Y W, Li H, Li J, et al. Spatiotemporal succession characteristics of algal functional groups and its impact factors for a typical channel-type reservoir in a southwest mountainous area[J]. Environmental Science, 2018, 39(6): 2680-2687. |
| [27] |
曹承进, 郑丙辉, 张佳磊, 等. 三峡水库支流大宁河冬、春季水华调查研究[J]. 环境科学, 2009, 30(12): 3471-3480. Cao C J, Zheng B H, Zhang J L, et al. Systematic investigation into winter and spring algal blooms in Daning River of Three Gorges Reservoir[J]. Environmental Science, 2009, 30(12): 3471-3480. |
| [28] | Paerl H W, Hall N S, Calandrino E S. Controlling harmful cyanobacterial blooms in a world experiencing anthropogenic and climatic-induced change[J]. Science of the Total Environment, 2011, 409(10): 1739-1745. DOI:10.1016/j.scitotenv.2011.02.001 |
| [29] | Paerl H W, Huisman J. Blooms like it hot[J]. Science, 2008, 320(5872): 57-58. DOI:10.1126/science.1155398 |
| [30] |
朱广伟, 金颖薇, 任杰, 等. 太湖流域水库型水源地硅藻水华发生特征及对策分析[J]. 湖泊科学, 2016, 28(1): 9-21. Zhu G W, Jin Y W, Ren J, et al. Characteristics of diatom blooms in a reservoir-water supply area and the countermeasures in Taihu Basin, China[J]. Journal of Lake Sciences, 2016, 28(1): 9-21. |
| [31] | Abirhire O, North R L, Hunter K, et al. Environmental factors influencing phytoplankton communities in Lake Diefenbaker, Saskatchewan, Canada[J]. Journal of Great Lakes Research, 2015, 41(S2): 118-128. |
| [32] | Tardio M, Tolotti M, Novarino G, et al. Ecological and taxonomic observations on the flagellate algae characterising four years of enclosure experiments in Lake Tovel (Southern Alps)[J]. Hydrobiologia, 2003, 502(1-3): 285-296. DOI:10.1023/B:HYDR.0000004286.28881.75 |
| [33] | Moustaka-Gouni M, Vardaka E, Michaloudi E, et al. Plankton food web structure in a eutrophic polymictic lake with a history of toxic cyanobacterial blooms[J]. Limnology and Oceanography, 2006, 51(1 Pt 2): 715-727. |
| [34] | Paerl H W, Fulton R S, Moisander P H, et al. Harmful freshwater algal blooms, with an emphasis on cyanobacteria[J]. The Scientific World Journal, 2001, 1: 76-113. DOI:10.1100/tsw.2001.138 |
| [35] |
况琪军, 马沛明, 胡征宇, 等. 湖泊富营养化的藻类生物学评价与治理研究进展[J]. 安全与环境学报, 2005, 5(2): 87-91. Kuang Q J, Ma P M, Hu Z Y, et al. Study on the evaluation and treatment of lake eutrophication by means of algae biology[J]. Journal of Safety and Environment, 2005, 5(2): 87-91. |
| [36] | Hu L L, Shan K, Huang L C, et al. Environmental factors associated with cyanobacterial assemblages in a mesotrophic subtropical plateau lake: a focus on bloom toxicity[J]. Science of the Total Environment, 2021, 777. DOI:10.1016/j.scitotenv.2021.146052 |
| [37] | Figueredo C C, Giani A, Bird D F. Does allelopathy contribute to Cylindrospermopsis raciborskii (cyanobacteria) bloom occurrence and geographic expansion?[J]. Journal of Phycology, 2007, 43(2): 256-265. DOI:10.1111/j.1529-8817.2007.00333.x |
| [38] | Leão P N, Vasconcelos M T S D, Vasconcelos V M. Allelopathy in freshwater cyanobacteria[J]. Critical Reviews in Microbiology, 2009, 35(4): 271-282. DOI:10.3109/10408410902823705 |
| [39] | Mack R N, Simberloff D, Lonsdale W M, et al. Biotic invasions: causes, epidemiology, global consequences, and control[J]. Ecological Applications, 2000, 10(3): 689-710. DOI:10.1890/1051-0761(2000)010[0689:BICEGC]2.0.CO;2 |
| [40] | Antunes J T, Leão P N, Vasconcelos V M. Cylindrospermopsis raciborskii: review of the distribution, phylogeography, and ecophysiology of a global invasive species[J]. Frontiers in Microbiology, 2015, 6. DOI:10.3389/fmicb.2015.00473 |
| [41] |
李红敏, 孙炯明, 张莎莎, 等. 拟柱孢藻(Cylindrospermopsis raciborskii)入侵对水生态系统群落结构和物种多样性的影响[J]. 生态学杂志, 2018, 37(12): 3757-3768. Li H M, Sun J M, Zhang S S, et al. The impacts of Cylindrospermopsis raciborskii invasion on community structure and species diversity of aquatic ecosystem[J]. Chinese Journal of Ecology, 2018, 37(12): 3757-3768. |
| [42] |
吴阿娜, 朱梦杰, 汤琳, 等. 淀山湖蓝藻水华高发期叶绿素a动态及相关环境因子分析[J]. 湖泊科学, 2011, 23(1): 67-72. Wu E N, Zhu M J, Tang L, et al. Dynamics of chlorophyll-a and analysis of environmental factors in Lake Dianshan during summer and autumn[J]. Journal of Lake Sciences, 2011, 23(1): 67-72. |
| [43] |
张宁红, 黎刚, 郁建桥, 等. 太湖蓝藻水华暴发主要特征初析[J]. 中国环境监测, 2009, 25(1): 71-74. Zhang N H, Li G, Yu J Q, et al. Character of blue-green algal blooms outbreak in Taihu Lake[J]. Environmental Monitoring in China, 2009, 25(1): 71-74. |
| [44] | Yan X C, Xu X G, Wang M Y, et al. Climate warming and cyanobacteria blooms: looks at their relationships from a new perspective[J]. Water Research, 2017, 125: 449-457. DOI:10.1016/j.watres.2017.09.008 |
| [45] | Shan K, Song L R, Chen W, et al. Analysis of environmental drivers influencing interspecific variations and associations among bloom-forming cyanobacteria in large, shallow eutrophic lakes[J]. Harmful Algae, 2019, 84: 84-94. DOI:10.1016/j.hal.2019.02.002 |
| [46] | Paerl H W, Otten T G. Duelling 'CyanoHABs': unravelling the environmental drivers controlling dominance and succession among diazotrophic and non-N2-fixing harmful cyanobacteria[J]. Environmental Microbiology, 2016, 18(2): 316-324. DOI:10.1111/1462-2920.13035 |
| [47] | Bai F, Shi J Q, Yang S Q, et al. Interspecific competition between Cylindrospermopsis raciborskii and Microcystis aeruginosa on different phosphorus substrates[J]. Environmental Science and Pollution Research, 2020, 27(34): 42264-42275. DOI:10.1007/s11356-020-08652-0 |
| [48] | Wu Z X, Shi J Q, Li R H. Comparative studies on photosynthesis and phosphate metabolism of Cylindrospermopsis raciborskii with Microcystis aeruginosa and Aphanizomenon flos-aquae[J]. Harmful Algae, 2009, 8(6): 910-915. DOI:10.1016/j.hal.2009.05.002 |
| [49] | Brauer V S, Stomp M, Rosso C, et al. Low temperature delays timing and enhances the cost of nitrogen fixation in the unicellular cyanobacterium Cyanothece[J]. The ISME Journal, 2013, 7(11): 2105-2115. DOI:10.1038/ismej.2013.103 |