 2022, Vol. 43
2022, Vol. 43



土壤是陆地生态系统中最大的碳库, 土壤呼吸的微小变化, 都会对全球大气碳平衡产生很大的影响[1].不同尺度上的土壤呼吸都具有强烈的空间异质性[2], 给评估陆地生态系统的碳预算带来困难.有研究表明, 温度和水分作为非常重要的环境变量, 可以很好地解释土壤呼吸大部分的时间变异, 但不能独立解释其空间分异性, 土壤呼吸还会受到地形、生物及土壤理化因子的影响而呈现空间异质性[3, 4]. Song等[5]的研究发现复杂的地形是影响土壤呼吸空间分异性的主导因子, 地形改变了植被根系分布, 使得高碳源区域表现出较强的空间异质性, Tian等[6]的研究也支持了这一观点.土壤有机碳和根系生物量能直接影响土壤呼吸的空间变化, 尤其在生长季, 根活性强, 可以更好地解释土壤呼吸的异质性[7].植物分布对土壤呼吸空间异质性的响应是动态的, 复杂的, 也有研究认为植被活动越强, 土壤呼吸空间变异越大[8].有研究表明土壤物理和化学因素对土壤呼吸的协同交互作用可通过线性关系表示[9].在不同生态环境下, 土壤呼吸空间变化的主导因子不同, 因此需要进行更深入地探究.近年来, 学者在群落尺度上对土壤呼吸的空间变化及机制进行了研究[10~13], 但在区域尺度上对于影响土壤呼吸的因素及其驱动机制耦合作用方面还没有一致的认知.为了精确量化区域土壤呼吸碳通量, 提高估计大气和陆地生态系统之间碳通量的能力和维持全球生态系统碳收支平衡, 需深入研究土壤呼吸的空间分异性.
地理探测器模型是一种探测事物及地理要素空间分异性及量化其驱动力的统计方法[14].与相关和回归分析法相比, 该方法的一个独特优势是能够探测两因子对因变量的交互作用.一般而言, 在回归模型中往往通过两因子的乘积项来表达两因子对因变量的交互作用.然而, 研究表明, 两因子的交互作用不一定就是相乘关系[15].地理探测器通过单因子的影响力和双因子叠加后的影响力, 判断两因子对因变量是否存在交互作用, 以及交互作用的强弱及其表现形式. 因此, 在分析地理要素空间分异和格局演变方面具有非常广泛的应用[16~19], 但对土壤呼吸空间分异性影响因素的分析的应用还未见报道.土壤呼吸的空间格局是多因子交互作用的结果, 传统的统计分析方法很难将其交互作用区分开.基于此, 本文以庞泉沟自然保护区为试验区, 运用地理探测器模型定量揭示气候、地形、土壤和生物因子对庞泉沟自然保护区土壤呼吸空间分异的单因子和不同因子之间的交互作用影响.以揭示环境因子对土壤呼吸空间分异性的作用机制, 以期为区域碳循环的可持续发展提供理论支持和决策参考.
1 材料与方法 1.1 研究区与试验地概况研究在庞泉沟自然保护区进行, 该区地处山西省吕梁市中西部(111°22′~111°33′E, 37°45′~37°55′N), 面积约10 443.5 hm2, 海拔1 600~2 831 m.该区属暖温带大陆性季风气候, 受到地形和海拔的影响, 温度较同纬度低, 空气湿度较高, 为典型的山地气候.年均温4.3℃, 夏季最高温达32.0℃, 冬季最低温为-26.0℃.降水主要集中在7~8月, 年平均降水量822.6 mm, 最大为2 023.8 mm, 最小为310.9 mm.土壤呈现垂直分布规律, 类型主要有:黄绵土、山地褐土、山地淋溶褐土、山地棕壤和草甸土.植被类型主要有华北落叶松林(Larix principis-rupprechtii Mayr)、云杉林(Picea asperata Mast)、辽东栎林(Quercus liaotungensis)、山杨林(Populus davidiana šuma)、青杄(Picea wilsonii)、白杄(Picea meyeri)、桦树(Betula)、油松林(Pinus tabulaeformis)、侧柏林(Platycladus orientalis)、鬼箭锦鸡儿灌丛(Caraganajubata)、银露梅灌丛(Potentilla.glabra)、沙棘灌丛(Hippophae rhamnoides shrub)、薹草草甸(Carex meadow)和嵩草草甸(Kobresia spp)等[20, 21].
本研究综合考虑植被类型、土壤类型、海拔高度和坡度等环境因子的代表性以及可达性, 选择了31个样地进行土壤呼吸及其环境因子观测(图 1).海拔介于1 600~1 800、1 800~2 000、2 000~2 200、2 200~2 400和2 400~2 700 m的样地数目分别为6、14、7、2和2(表 1).用GPS测定样地的经纬度和海拔(ELE).植被类型涵盖了针叶林(主要以华北落叶松为主)、针阔混交林(云杉、华北落叶松、辽东栎林、山杨林、白桦混交)、阔叶林(山杨林、辽东栎林)、阔叶混交林(杨桦混交、山杨辽东栎混交)、亚高山草甸和灌丛(沙棘)这6种不同的植被型组.

|
图 1 研究区土壤呼吸测定样地示意 Fig. 1 Sample sites for soil respiration measurement in the study area |
|
|
表 1 不同海拔的植被类型 Table 1 Vegetation types at different altitudes |
1.2 数据来源及处理 1.2.1 土壤呼吸测定
土壤呼吸速率(Rs)用LI-COR-6400便携式气体分析系统连接LI-6400-09土壤呼吸叶室测定.测定前1 d, 每个样地随机放置12个PVC环, 并将环内表面的植物去除.测定时, 每个PVC环测定3个数据, 取12个样点的平均值作为该样地当天的呼吸速率.测定于2018年9月12~15日进行, 天气状况均为晴朗, 测定时间为每天10:00~16:00.
1.2.2 环境因子测定用LI-COR-6400的土壤温度探针测定10 cm深度的土壤温度(T). 0~10 cm深度的土壤含水量(SWC)采用土钻法测定. 0~10 cm深度的土壤容重(BD)采用环刀法测定. Rs测定完成后, 用土钻取PVC环中0~10 cm深度的土壤样品, 带回实验室用于测定土壤水分、土壤全氮(N)和总碳(C).N和C使用vario max常量元素分析仪(Elementar, Germany)测定.测得的土壤C、N和BD的含量见图 2.

|
图 2 不同样地的土壤全碳(C)、全氮(N)和容重(BD) Fig. 2 Soil total carbon (C), total nitrogen (N), and bulk density (BD) of different sampling plots |
从美国国家航天航空局(NASA)网站(https://ladsweb.modaps.eosdis.nasa.gov/search/)订购了中分辨率成像光谱仪(MODIS)的Rs测定时期的MOD13A1产品, 提取了研究区的归一化植被指数(NDVI)数据, 用于作为生物因子的指标分析对Rs空间分异性的影响.DEM高程数据来源于地理空间数据云(http://www.gscloud.cn/search), 空间分辨率为30 m.利用ArcGIS 10.2根据采样点的经纬度提取了各样地的坡度(slope), 用于分析地形因子对土壤呼吸空间变异性的影响.
1.3 研究方法 1.3.1 传统方法采用变异系数(CV)分析土壤呼吸和环境因子的空间变异程度.采用皮尔逊相关法分析气象因子、生物因子、土壤因子和地形因子与Rs的相关关系.采用Origin 7.0(Microcal)绘制所有图.
1.3.2 标准差分级法由于地理探测器模型运行的数据是离散变量, 而Rs影响因子数据属于连续变量, 因此需将连续变量离散化.标准差分级法是以均值为中心, 以标准差倍数为级差, 分别向大于和小于均值的两个方向进行分级[22].该方法避免人为因素对数据干扰, 根据原始数据的数值特征和分布规律进行分级, 使计算结果更具客观性.选择不同分级方法得到的q值最大的分类数, 分级效果最佳.本文通过比较不同倍标准差下q值, 采用1倍标准差分级效果最好.
1.3.3 地理探测器模型地理探测器能够判断影响因子决定力的大小, 分析要素的空间分布是否与地理事物趋势密切相关, 及影响因素间的耦合关系.
(1) 因子探测主要通过因子解释力的大小探究不同变量对Rs的贡献率, 引入决定力指标q.假设Rs为Y, A={Ah}为可能影响Rs的一个因子, h=1, 2, …, L, L为按照标准分级法得到的因子分类数, h为因子A的不同类型.为了探究因子A与Y的空间关联性, 将Rs图层与因子A图层叠合[23].计算公式如下:
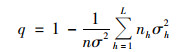
|
(1) |
式中, n和nh分别为总样本数和h层样本量; σ2和σh2分别为总样本方差和h层样本方差; q取值范围为[0, 1], q值越大, 因子对Rs的解释力越强, q=0时, 影响因子对Rs没有影响, q=1时, 影响因子完全解释Rs.
将ELE、T、SWC、NDVI、slope、N、C和BD这8个影响因子分别与研究区Rs进行因子探测分析, 计算得到各因子对Rs的解释力q, 对影响研究区Rs空间分异性的主导因素进行诊断.然后利用SPSS 22软件, 根据q值大小, 用聚类分析法探究Rs空间分异性的主要驱动力.
(2) 生态探测主要比较因子x1和x2在对Rs空间分布的影响是否存在显著差异[24], 通过F检验来衡量.
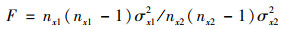
|
(2) |
式中, nx1和nx2分别为因子x1和x2的样本量; σx12和σx22分别为因子x1和x2的方差.
(3) 交互探测用于探究不同因子对Rs空间变化的削弱或增强的交互作用或相互独立关系.用“∩”表示两因子间交互作用. ①非线性拮抗:q(x1∩x2)<Min[q(x1), q(x2)], 表示两个因子的共同作用小于其中任何一个因子的作用; ②单拮抗:Min[q(x1), q(x2)]<q(x1∩x2)<Max[q(x1), q(x2)], 表示两个因子的共同作用大于其中一个因子的作用且小于另一个因子的作用; ③双协同:Max[q(x1), q(x2)]<q(x1∩x2)<q(x1)+q(x2), 表示两个因子的共同作用大于其中任何一个因子的作用且小于两个因子作用之和; ④独立:q(x1∩x2)=q(x1)+q(x2), 表示两个因子的共同作用等于两个因子作用之和; ⑤非线性协同:q(x1∩x2)>q(x1)+q(x2), 表示两个因子的共同作用大于两个因子作用之和[25].
2 结果与分析 2.1 土壤呼吸与环境因子的相关性分析研究区Rs在0.86~12.29 μmol ·(m2 ·s)-1, 平均值为(6.13±2.83) μmol ·(m2 ·s)-1. Rs和各环境因子的变异系数(CV)的变化范围为12% ~57%, 均为中等变异.其中, SWC的CV最大为57%, ELE、T和BD较低分别为12%、14%和15%, 其余各因子的CV与Rs的差异不大, 在30% ~48%之间(表 2).皮尔逊相关性分析表明, Rs与NDVI、T和N呈极显著正相关(P<0.01), 与ELE、slope和SWC呈极显著负相关(P<0.01), 与BD显著负相关(P<0.05), 与C相关性不显著(P>0.05)(表 3).
|
|
表 2 土壤呼吸及其驱动因子的描述统计分析1) Table 2 Descriptive statistical analysis of soil respiration and its driving factors |
|
|
表 3 土壤呼吸与环境因子的相关性分析1) Table 3 Correlation between soil respiration and environmental factors |
2.2 土壤呼吸空间分异性的多元线性逐步回归分析
多元线性逐步回归的结果表明, Tolerance值大于0.1, VIF值小于10, 说明模型不存在共线性问题.模型保留参数的回归系数的Sig.值均小于0.01, 表明各系数都达到显著性水平.NDVI和T构成的多元线性模型可以解释Rs空间分异的64.3%(表 4).
|
|
表 4 多元线性逐步回归分析 Table 4 Multiple linear stepwise regression analysis |
2.3 基于地理探测器的土壤呼吸空间分异性分析 2.3.1 环境因子对土壤呼吸空间分异性的影响
因子探测器结果表明, 各影响因子q值均大于0, 且均通过显著性检验(P<0.000 1), 表明Rs在各驱动力分区的空间分布存在空间异质性.影响Rs的各因子驱动力q值从大到小依次为:ELE(0.64)>T(0.59)>NDVI(0.48)>SWC(0.32)>slope(0.27)>N(0.14)>C(0.13)>BD(0.10).以上分析表明, ELE和T是影响研究区Rs空间分异性的主要驱动因子, 可以解释Rs空间变化的64%和59%, 其次为NDVI和SWC, 其解释力为0.48和0.32.土壤因子的解释力较低, N、C和BD的解释力均低于0.15.聚类分析表明, 8个环境因子可以分为2类, 一类以ELE、T和NDVI为代表, 其解释力q值高于0.48, 称为主导驱动力; 另一类以SWC、slope、N、C和BD为代表, 其解释力q值低于0.32, 为重要驱动力.
2.3.2 各环境因子对土壤呼吸空间分异性的影响显著性分析通过探测两个影响因子对Rs的空间变化的驱动作用是否存在显著差异, 即相对重要性是否显著, 进行显著性分析(表 5).可以看出, ELE除了与slope对Rs的空间分异性存在显著差异外, 与其他因子均无显著差异; T除了与ELE对Rs的空间分布变化无显著差异外, 与其他因子均存在显著差异; slope与ELE、SWC、T和NDVI对Rs的空间分布变化具有显著差异; SWC在对Rs的空间分异性上与T和NDVI有显著差异; NDVI与slope、SWC和T对Rs的空间分布变化具有显著差异; BD、C和N均与其他所有因子对Rs的空间分布变化不存在显著差异.结合因子探测器的结果, 可以判断出T、ELE和NDVI是土壤呼吸空间分异性的主要驱动因子.
|
|
表 5 生态探测器结果1) Table 5 Results of ecoprobe |
2.3.3 环境因子对土壤呼吸空间分异性的交互作用
交互探测器主要用来探测2个影响因子共同作用时是否会增加或者减弱对Rs空间分异性的决定力, 或这些因子对Rs的影响是相互独立的, 从而确定2因子对Rs的交互作用是非线性减弱、单因子非线性减弱、双因子增强、独立或者非线性增强.交互探测结果表明, 8个影响因子两两间的交互作用的q值都大于任何因子单独作用的q值(表 6), 表明两因子的交互作用均会增强对Rs空间分异性的解释能力.ELE和T与其他7个因子两两之间的交互作用均为双协同作用.NDVI除与BD为非线性协同作用外, 与其他6个因子两两之间的交互作用均为双协同作用.SWC、slope、BD、C和N这5个因子两两之间的交互作用均为非线性协同作用, 双因子的解释能力高于任何一个单因子的q值.总体来看, 解释力低的单因子两两间的交互作用都高于两个因子q值的和, 而解释力高的单因子与其他因子的交互作用则表现为双协同作用, 高于两个变量的最大q值, 低于两个变量的q值和. 8个因子中, ELE、T与其他驱动因子的交互作用解释Rs空间分异性的能力最强, 均为双协同作用, 解释力在0.70左右, 其中ELE∩BD(q=0.73)和T∩slope(q=0.74)的解释力最强.与两个因子的最大q值相比, SWC、slope、BD、C和N这5个因子两两之间的交互作用的q值提高幅度较大, 解释力都增加了0.15~0.48.表明自然条件下, 气象、地形、生物和土壤因子对Rs的空间变化不是单独起作用, 也不是简单的双因子叠加, 而是通过非线性协同和双协同的双向促进作用来影响Rs的空间分布.
|
|
表 6 各驱动因子对土壤呼吸空间分异性的交互作用探测结果 Table 6 Interactive effects of driving factors on spatial variability of soil respiration |
3 讨论
土壤呼吸的空间变异程度可通过变异系数量化, 不同生态系统的土壤微环境不同, 所以变异程度不同.多数研究表明Rs的CV介于20% ~70%[3, 9, 26], 属于中等变异.张义辉等[27]的研究发现太原盆地土壤呼吸的空间CV为32% ~59%; Tian等[6]的研究指出湖南省八大公山自然保护区Rs的平均CV为(40.7±2.1)%; 郭全恩等[28]对绿洲荒漠过渡带土壤呼吸进行了研究, 结果发现土壤呼吸受到表层水分的限制, CV为64.1% ~78.1%.本研究中, 庞泉沟Rs的CV为46%, 属于中等变异范围, 位于报道的范围内.
地理探测器模型通过探测因变量与自变量之间空间分布格局的一致性, 评价自变量对因变量的解释能力, 比一般统计量能更明确地揭示因果关系[15].因子探测器分析表明, 海拔、温度和植被指数是影响研究区Rs空间变化的主导驱动因子, 分别可以解释研究区土壤呼吸空间分布的64%、59%和48%.不同海拔样点数存在差异, 通过方差齐性检验结果为方差齐性, 因此对土壤呼吸的空间响应的影响不大.多元线性回归分析表明Rs在空间位置产生的变异主要是由温度和植被指数共同影响的, 可以解释Rs空间分异的64.3%.温度与海拔相关性很强, 存在一定的共线性.相关分析表明(表 3), ELE与T和NDVI呈显著负相关(r为-0.80和-0.59, P<0.01), 与SWC呈显著正相关(r=0.54, P<0.01), 表明海拔是多因素的综合反应, 随着海拔的升高, 温度降低、土壤水分增加, 相应的植被类型也出现了有规律的变化(阔叶林-针叶林-亚高山草甸), 从而对Rs空间格局产生重要的影响.土壤中的生物化学和非生物过程都会受到温度的影响.随着温度升高, 土壤微生物酶活性增强, 有机质加速分解, 加快了碳周转速率, 进而影响Rs[29].植被指数和叶面积指数是表征植物光合作用的关键指标, 影响Rs的底物供应[30].本研究中, 归一化植被指数是第3个影响Rs的空间分异的因子, 与Rs呈显著正相关(P<0.01).其原因是地上植被通过光合作用固碳, 合成的光合产物分配给根系, 根系生物依赖地下的同化产物量, 直接影响Rs的空间变化.研究区土壤呼吸与土壤水分呈现负相关, 能够解释Rs空间格局的32%.这可能是因为, 本区属于典型的山地气候, 海拔较高(1 600~2 831 m), 全年土壤温度较低, 降水充足, 土壤蒸发和植物蒸腾较弱, 土壤水分维持在一个较高的水平, 不存在水分限制Rs的情况[31], Rs对温度的变化更为敏感, 随着温度的升高而增加, 而土壤温度与土壤水分呈极显著负相关(r=-0.58), 因此Rs表现出与土壤水分呈现负相关的结果.坡度主要通过影响土壤水分、土壤养分含量等进而影响土壤呼吸, 随着坡度的增加, 土壤总碳和总氮含量显著减低(r为-0.58和-0.42), 因此Rs与坡度呈极显著负相关(r=-0.45), 其解释力q值为0.27.此外, 本研究中, 土壤全氮、总碳和容重分别能解释Rs的14%、13%和10%的空间变化.氮是植物生长的重要元素, 不仅会影响光合作用, 也是根活力指示因子, 对Rs至关重要[29].土壤中氮的变化会影响微生物活性进而影响呼吸, 本研究中Rs与土壤全氮呈极显著正相关(r=0.33), 与刘燕飞等[32]的研究结果相一致.土壤有机质含量对土壤的通风性、阳离子保持能力等都有重要的影响, 是微生物呼吸的物质基础[33].本研究中, 相关分析和因子探测器都表明, 土壤全碳含量对Rs影响较小, 可能与本区海拔较高, 土壤有机质含量较高, 样地间土壤有机碳含量空间变异较小有关, 与董利军等[34]的研究结果相一致.土壤容重反映了Rs的畅通程度, 研究结果显示Rs与容重为显著负相关(r=-0.26), 可能与容重较高, 孔隙度较低有关.
自然条件下, Rs同时受多因素的交互作用, 通常对最限制它的因子做出响应[29].在温度低于5℃时, Rs对水分的变化不敏感, 但温度在10~20℃时反应敏感.同样, 水分含量低时对温度的变化不敏感, 水分含量高时随着温度的变化而变化[35].当温度和水分含量都不存在限制的时候, 二者对Rs交互作用明显[35].本研究中温度和水分的交互作用高于两个因子的单独作用.其他的因子也可能与温度和水分交互作用影响Rs.当温度维持在14℃时, Rs受水分和pH的共同影响.当水分维持在田间持水量的60%时, Rs主要受pH和土壤有机质含量的共同影响[36].研究区土壤有机质也与其他因子一起影响Rs, 交互作用解释力增大.光合作用大部分产物都被分配到土壤易分解的碳库中[37], 因此, 生长季Rs随温度变化而产生的变化更大.本研究中, 两因子共同作用时均会增加对研究区土壤呼吸空间分布的解释能力, 表现为双协同作用或者非线性协同作用, 其中ELE∩BD(q=0.73)和T ∩ slope(q=0.74)的解释力最强, 证明了研究区Rs同时受到气象、生物、地形和土壤等多个因子相互作用和相互影响, 各要素通过其自身性质或与其他要素间相互耦合胁迫来影响Rs的空间变化.与传统的多元线性回归分析法相比, 地理探测器无线性假设, 能够探测两变量真正的交互效应, 判断两因子是否存在交互作用, 以及交互作用的强弱、方向、线性还是非线性, 不仅仅局限于用两个变量的乘积来表示交互, 而且对自变量间共线性不做要求, 避免出现过度拟合的现象[14, 15].
4 结论庞泉沟自然保护区Rs及其影响因子的空间变异均为中等变异. Rs与NDVI、T和N呈极显著正相关(P<0.01), 与ELE、slope和SWC呈极显著负相关(P<0.01), Rs与BD显著负相关(P<0.05), 与C相关性不显著(P>0.05). Rs影响因子驱动力从大到小依次为ELE>T>NDVI>SWC>slope>N>C>BD. ELE、T和NDVI为主导驱动力, 其解释力q值分别为0.64、0.59和0.48.各因子间的交互作用表现为双协同作用和非线性协同作用, 表明两因子的交互作用均会增强对Rs空间分异性的解释能力, 其中ELE∩BD(q=0.73)和T ∩ slope(q=0.74)的解释力最强.表明田间条件下, 气象、地形、生物和土壤因子对Rs的空间变化不是单独起作用, 也不是简单的双因子叠加, 而是通过非线性协同和双协同的双向促进作用来影响Rs的空间分布.
| [1] | Shi B K, Gao W F, Cai H Y, et al. Spatial variation of soil respiration is linked to the forest structure and soil parameters in an old-growth mixed broadleaved-Korean pine forest in northeastern China[J]. Plant and Soil, 2016, 400(1-2): 263-274. DOI:10.1007/s11104-015-2730-z |
| [2] | Liang G P, Cai A D, Wu H J, et al. Soil biochemical parameters in the rhizosphere contribute more to changes in soil respiration and its components than those in the bulk soil under nitrogen application in croplands[J]. Plant and Soil, 2019, 435(1-2): 111-125. DOI:10.1007/s11104-018-3886-0 |
| [3] | Han M G, Shi B K, Jin G Z. Spatial patterns of soil respiration in a spruce-fir valley forest, Northeast China[J]. Journal of Soils and Sediments, 2019, 19(1): 10-22. DOI:10.1007/s11368-018-2018-z |
| [4] |
郑甲佳, 黄松宇, 贾昕, 等. 中国森林生态系统土壤呼吸温度敏感性空间变异特征及影响因素[J]. 植物生态学报, 2020, 44(6): 687-698. Zheng J J, Huang S Y, Jia X, et al. Spatial variation and controlling factors of temperature sensitivity of soil respiration in forest ecosystems across China[J]. Chinese Journal of Plant Ecology, 2020, 44(6): 687-698. |
| [5] | Song Q H, Tan Z H, Zhang Y P, et al. Spatial heterogeneity of soil respiration in a seasonal rainforest with complex terrain[J]. IForest-Biogeosciences and Forestry, 2013, 6(2): 65-72. DOI:10.3832/ifor0681-006 |
| [6] | Tian Q X, Wang D Y, Tang Y N, et al. Topographic controls on the variability of soil respiration in a humid subtropical forest[J]. Biogeochemistry, 2019, 145(1-2): 177-192. DOI:10.1007/s10533-019-00598-x |
| [7] |
焦振. 帽儿山温带森林土壤呼吸组分时空动态及其影响因子[D]. 哈尔滨: 东北林业大学, 2019. Jiao Z. Spatiotemporal dynamics in soil respiration components of the temperate forests in the Maoershan region of Northeast China[D]. Harbin: Northeast Forestry University, 2019. |
| [8] | Gao G Q, Goebel M, Wang Y, et al. Spatial-temporal variations of absorptive fine roots in the organic and soil layers of a Larix gmelinii forest[J]. Trees, 2021, 35(3): 1013-1023. DOI:10.1007/s00468-021-02101-2 |
| [9] | Cai Y H, Nishimura T, Ida H, et al. Spatial variation in soil respiration is determined by forest canopy structure through soil water content in a mature beech forest[J]. Forest Ecology and Management, 2021, 501. DOI:10.1016/J.FORECO.2021.119673 |
| [10] |
廖艳. 我国典型低温区土壤有机碳变化及土壤呼吸特征[D]. 北京: 中国地质大学(北京), 2011. Liao Y. Soil organic carbon change and soil respiration characteristics in the typical regions with low temperature in China[D]. Beijing: China University of Geosciences (Beijing), 2011. |
| [11] |
毛亚辉. 青藏高原高寒沼泽草甸土壤呼吸动态变化及对增温的响应——以瓦颜山为例[D]. 青海: 青海师范大学, 2016. Mao Y H. Respiration dynamic change of the Qinghai-Tibet plateau alpine swamp meadow soil and its response to warming——A case study of WayanShan Mountain[D]. Qinghai: Qinghai Normal University, 2016. |
| [12] |
丁梦凯, 胡晓农, 曹建华, 等. 桂林毛村不同土地利用方式下土壤呼吸空间变异特征[J]. 环境科学, 2019, 40(2): 924-933. Ding M K, Hu X N, Cao J H, et al. Spatial variability in soil respiration under different land use patterns in Maocun Village, Guilin[J]. Environmental Science, 2019, 40(2): 924-933. |
| [13] |
陈全胜. 内蒙古锡林河流域草原群落土壤呼吸的时空变异及其影响因子研究[D]. 北京: 中国科学院大学(中国科学院植物研究所), 2002. Chen Q X. Study on the spatio-temporal variation of soil respiration and the controlling factors in the Xilin River Basin of inner Mongolia[D]. Beijing: University of Chinese Academy of Sciences (Institute of Botany, the Chinese Academy of Sciences), 2002. |
| [14] |
王劲峰, 徐成东. 地理探测器: 原理与展望[J]. 地理学报, 2017, 72(1): 116-134. Wang J F, Xu C D. Geographic detectors: Principles and prospects[J]. Acta Geographica Sinica, 2017, 72(1): 116-134. |
| [15] | Wang J F, Hu Y. Environmental health risk detection with GeogDetector[J]. Environmental Modelling & Software, 2012, 33: 114-115. |
| [16] |
付明花. 武汉市不透水层时空格局分析及其在城市内涝成因分析中的应用[D]. 武汉: 武汉大学, 2017. Fu M H. Exploring the spatio-temporal patterns of impervious layer in Wuhan and its application in the analysis of urban waterlogging[D]. Wuhan: Wuhan University, 2017. |
| [17] |
张莹, 张学玲, 蔡海生. 基于地理探测器的江西省万安县生态脆弱性时空演变及驱动力分析[J]. 水土保持通报, 2018, 38(4): 207-214. Zhang Y, Zhang X L, Cai H S. Temporal and spatial evolutions and its driving factors of ecological vulnerability in Wan'an county of Jiangxi province based on geogdetector[J]. Bulletin of Soil and Water Conservation, 2018, 38(4): 207-214. |
| [18] |
杨忍, 刘彦随, 龙花楼, 等. 基于格网的农村居民点用地时空特征及空间指向性的地理要素识别——以环渤海地区为例[J]. 地理研究, 2015, 34(6): 1077-1087. Yang R, Liu Y S, Long H L, et al. Spatial-temporal characteristics of rural residential land use change and spatial directivity identification based on grid in the Bohai Rim in China[J]. Geographical Research, 2015, 34(6): 1077-1087. |
| [19] | Wu Q M, Hu W Y, Wang H F, et al. Spatial distribution, ecological risk and sources of heavy metals in soils from a typical economic development area, Southeastern China[J]. Science of the Total Environment, 2021, 780. DOI:10.1016/J.SCITOTENV.2021.146557 |
| [20] |
李世广, 张峰. 山西庞泉沟国家级自然保护区生物多样性与保护管理[M]. 北京: 中国林业出版社, 2014. Li S G, Zhang F. Biodiversity, conservation and management of Pangquangou National Nature Reserve, Shanxi[M]. Beijing: China Forestry Publishing House, 2014. |
| [21] | 马子清. 山西植被[M]. 北京: 中国科学技术出版社, 2001. |
| [22] |
孙道亮. 都江堰市农村居民点时空演变与布局优化研究[D]. 成都: 四川师范大学, 2020. Sun D L. Study on the spatial-temporal evolution and layout optimization of rural settlement areas in Dujiangyan county[D]. Chengdu: Sichuan Normal University, 2020. |
| [23] |
刘彦随, 李进涛. 中国县域农村贫困化分异机制的地理探测与优化决策[J]. 地理学报, 2017, 72(1): 161-173. Liu Y S, Li J T. Geographic detection and optimizing decision of the differentiation mechanism of rural poverty in China[J]. Acta Geographica Sinica, 2017, 72(1): 161-173. |
| [24] |
潘洪义, 黄佩, 徐婕. 基于地理探测器的岷江中下游地区植被NPP时空格局演变及其驱动力研究[J]. 生态学报, 2019, 39(20): 7621-7631. Pan H Y, Huang P, Xu J. The spatial and temporal pattern evolution of vegetation NPP and its driving forces in middle-lower areas of the Min river based on geographical detector analyses[J]. Acta Ecologica Sinica, 2019, 39(20): 7621-7631. |
| [25] |
周伟, 李丽丽, 周旭, 等. 基于地理探测器的土壤重金属影响因子分析及其污染风险评价[J]. 生态环境学报, 2021, 30(1): 173-180. Zhou W, Li L L, Zhou X, et al. Influence factor analysis of soil heavy metal based on geographic detector and its pollution risk assessment[J]. Ecology and Environmental Sciences, 2021, 30(1): 173-180. |
| [26] |
姚雄, 余坤勇, 曾琪, 等. 福建西部山地水土流失区土壤呼吸的空间异质性[J]. 环境科学, 2016, 37(12): 4789-4799. Yao X, Yu K Y, Zeng Q, et al. Spatial heterogeneity of soil respiration in the soil erosion area of west mountains in Fujian province, China[J]. Environmental Science, 2016, 37(12): 4789-4799. |
| [27] |
张义辉, 李洪建, 荣燕美, 等. 太原盆地土壤呼吸的空间异质性[J]. 生态学报, 2010, 30(23): 6606-6612. Zhang Y H, Li H J, Rong Y M, et al. Study on spatial heterogeneity of soil respiration in Taiyuan Basin[J]. Acta Ecologica Sinica, 2010, 30(23): 6606-6612. |
| [28] |
郭全恩, 李保国, 南丽丽, 等. 疏勒河流域绿洲荒漠过渡带土壤呼吸特征及其影响因素[J]. 生态环境学报, 2016, 25(6): 934-938. Guo Q E, Li B G, Nan L L, et al. Characteristics of soil respiration and its influence factors in oasis-desert transitional belt of Shule River Basin[J]. Ecology and Environmental Sciences, 2016, 25(6): 934-938. |
| [29] |
严俊霞, 李洪建, 李君剑, 等. 山西高原落叶松人工林土壤呼吸的空间异质性[J]. 环境科学, 2015, 36(5): 1793-1801. Yan J X, Li H J, Li J J, et al. Spatial heterogeneity of soil respiration in a planted larch forest in Shanxi Plateau[J]. Environmental Science, 2015, 36(5): 1793-1801. |
| [30] |
魏书精, 罗碧珍, 孙龙, 等. 森林生态系统土壤呼吸时空异质性及影响因子研究进展[J]. 生态环境学报, 2013, 22(4): 689-704. Wei S J, Luo B Z, Sun L, et al. Spatial and temporal heterogeneity and effect factors of soil respiration in forest ecosystems: a review[J]. Ecology and Environmental Sciences, 2013, 22(4): 689-704. |
| [31] | Yan J X, Zhang X, Liu J, et al. MODIS-Derived estimation of soil respiration within five cold temperate coniferous forest sites in the Eastern Loess Plateau, China[J]. Forests, 2020, 11(2). DOI:10.3390/f11020131 |
| [32] |
刘燕飞, 张羽, 赖金美, 等. 土壤氮水交互对马尾松和杉木COS和CO2通量的影响[J]. 生态学报, 2020, 40(16): 5729-5738. Liu Y F, Zhang Y, Lai J M, et al. Effects of soil nitrogen and water interaction on COS and CO2 fluxes of Pinus massoniana and Cunninghamia lanceolata[J]. Acta Ecologica Sinica, 2020, 40(16): 5729-5738. |
| [33] |
李键, 刘鑫铭, 姚成硕, 等. 武夷山国家公园不同林地土壤呼吸动态变化及其影响因素[J]. 生态学报, 2021, 41(9): 3588-3602. Li J, Liu X M, Yao C S, et al. Dynamic change of soil respiration and its effecting factors in different forest lands of Wuyishan National Park[J]. Acta Ecologica Sinica, 2021, 41(9): 3588-3602. |
| [34] |
董利军, 李金花, 陈珊, 等. 若尔盖湿地高寒草甸退化过程中土壤有机碳含量变化及成因分析[J]. 植物生态学报, 2021, 45(5): 507-515. Dong L J, Li J H, Chen S, et al. Changes in soil organic carbon content and their causes during the degradation of alpine meadows in Zoige Wetland[J]. Chinese Journal of Plant Ecology, 2021, 45(5): 507-515. |
| [35] |
左嫚, 陈奇伯, 黎建强, 等. 枯落物输入变化对云南松林地CO2释放的影响[J]. 生态学报, 2021, 41(11): 4552-4561. Zuo M, Chen Q B, Li J Q, et al. Effects of alteration in forest litter input on CO2 release in Pinus yunnanensis forestland in central Yunnan Plateau[J]. Acta Ecologica Sinica, 2021, 41(11): 4552-4561. |
| [36] |
李一凡, 王玉杰, 王彬, 等. 三峡库区典型林分土壤呼吸及其组分对模拟酸雨的响应[J]. 环境科学, 2019, 40(3): 1457-1467. Li Y F, Wang Y J, Wang B, et al. Response of soil respiration and its components to simulated acid rain in a typical forest stand in the Three Gorges Reservoir Area[J]. Environmental Science, 2019, 40(3): 1457-1467. |
| [37] |
宋明华, 陈锦, 蒋婧, 等. 13 CO2示踪不同化学形态氮素添加对高寒草甸植物光合碳分配的影响[J]. 生态学报, 2020, 40(11): 3688-3697. Song M H, Chen J, Jiang J, et al. Effect of nitrogen addition with different chemical forms on dynamics of photosynthate using 13 CO2 tracing in an alpine grassland[J]. Acta Ecologica Sinica, 2020, 40(11): 3688-3697. |