 2022, Vol. 43
2022, Vol. 43



植被是陆地表面生态系统重要的组成部分, 是联结大气圈、水圈、生物圈和岩石圈的桥梁, 其变化具有时间上的复杂性和空间上的多样性[1~4].归一化植被指数(normalized difference vegetation index, NDVI)与植被叶绿素[5]、叶面积指数[6]、生物量[7]和植被覆盖度[8]等紧密相关, 故NDVI常用于植被覆盖变化[8]、水土保持和荒漠化进程[9, 10]、生态环境质量监测[11]和气候变化[3, 12, 13]等方面的研究.长江流域作为亚洲第一大流域, 其植被覆盖状况及动态变化对我国和周边国家乃至全球生态平衡有着十分重要的作用.因此, 研究长江流域植被NDVI动态变化并探测驱动其变化的主要影响因素, 对区域生态环境保护和植被资源管理有着重要的现实意义.
近年来针对长江流域植被NDVI动态变化及影响因素探测方面的研究取得了长足的进展.有研究指出1982~2015年长江流域植被NDVI整体呈上升趋势, 上升区域主要分布在长江流域中部, 而下降区域主要分布在长江流域东部[14~16].Qu等[14]进一步指出1994年后长江流域植被NDVI上升趋势相比之前更加明显.Qu等[14]和崔利芳等[17]的研究指出气温是影响长江流域植被生长的主要气候因子, 而降水对植被生长的影响较弱.Zhang等[15]进一步指出长江流域植被生长对降水和气温变化均具有滞后效应, 且超过50%的区域植被生长受气候变化的影响强于人类活动.崔浩楠等[18]的研究指出, 1982~2019年长江经济带植被NDVI呈上升趋势, 上升趋势呈明显的阶段性特征, 且非气候因素对长江经济带植被NDVI变化的影响强于气候因素.以上研究大部分是基于长江流域整体或流域部分地区, 考虑的因子多为气温和降水等气候因子和海拔等地形因子, 忽略了植被生长在不同流域单元存在的差异以及其他气候因子、人文因子和地形因子对植被生长的影响.
地理探测器[19]是一个空间分析模型, 可定量探测气候因子、人文因子和地形因子对植被NDVI空间分异的影响, 并且可以探测双因子交互作用对植被NDVI变化的影响, 因此, 被广泛应用于植被NDVI空间分异影响因素的探测.基于地理探测器, Han等[20]的研究指出气候因子是影响柴达木盆地植被退化最主要的因素, 环境因子和人文因子对柴达木盆地植被退化的影响依次减弱.Sun等[21]的研究指出气候因子对祁连山草地空间分异的影响程度高于其他植被类型.林地植被生长季起点和终点主要受气温和日照时数的影响, 草地和草甸植被生长季的起点主要受降水的影响, 而终点主要受最低气温的影响.Peng等[22]和Liu等[23]的研究指出四川和青藏高原东北部植被生长主要受土壤类型、海拔和平均气温的影响.周明星等[24]的研究指出邛崃山大熊猫栖息地植被NDVI变化除受土壤类型、海拔和平均气温影响以外, 还受降水的影响, 且自然因子对植被NDVI影响存在明显的协同效应.潘洪义等[25]的研究指出岷江中下游地区植被NPP空间分异主要受气温、海拔和土地利用类型的影响, 且各因素对植被NPP空间分异的解释力存在差异.以上研究表明, 地理探测器不仅可以有效探测区域植被NDVI空间分异的影响因素, 还可以揭示影响因素解释力的大小及影响因素间的协同作用.
综上, 已有研究大多侧重分析长江流域植被NDVI时空演变特征及其驱动因素, 但分析长江流域及其二级流域单元植被NDVI时空演变特征, 并利用地理探测器探测驱动植被NDVI空间分异的影响因素, 分析影响因素之间的交互作用的研究并不常见.鉴于此, 本文以长江流域为研究区, 以长江流域整体和二级流域为研究单元, 研究长江流域植被NDVI时空演变特征, 并在多级尺度上探测人文因子(夜间灯光和人口密度)、地形因子(坡度、坡向、高程和曲率)和气候因子(最高气温、最低气温、地表温度、平均气温、寒冷指数、温暖指数、气压、风速、降水、相对湿度、日照时数和湿度指数)对长江流域植被NDVI空间分异的影响, 以期为长江流域植被恢复工程的实施和生态环境保护政策的制定等提供参考.
1 研究区概况长江是我国第一长河, 发源于青藏高原的唐古拉山脉, 全长6 397 km, 位于90°33′~122°25′E, 24°30′~35°45′N之间, 横跨11个省级行政区, 流域面积达180万km2, 其流域范围包含金沙江石鼓以上(A1)、金沙江石鼓以下(A2)、岷沱江(A3)、嘉陵江(A4)、宜宾至宜昌(A5)、乌江(A6)、汉江(A7)、宜昌至湖口(A8)、洞庭湖水系(A9)、鄱阳湖水系(A10)、湖口以下干流(A11)和太湖水系(A12), 共计12个流域单元(图 1).长江流域高程在-70~6 444 m之间, 地势由西向东呈阶梯式下降, 横跨“第一级阶梯”、“第二级阶梯”和“第三级阶梯”, 地貌类型复杂, 区域内有高原、山地、丘陵、盆地和平原.气候类型以亚热带季风气候为主, 植被类型主要以针叶林、阔叶林、栽培植被、灌丛、草甸和草丛为主.长江流域是中国具有全球影响力的内河经济带、东中西互动合作的协调发展带和生态文明建设的先行示范带.但经济发展的同时, 受气候变化和人类活动的共同影响, 长江流域部分地区生态环境脆弱, 植被生长对气候变化和人类活动响应敏感.

|
A1表示金沙江石鼓以上, A2表示金沙江石鼓以下, A3表示岷沱江, A4表示嘉陵江, A5表示宜宾至宜昌, A6表示乌江, A7表示汉江, A8表示宜昌至湖口, A9表示洞庭湖水系, A10表示鄱阳湖水系, A11表示湖口以下干流, A12表示太湖水系 图 1 研究区气象站及高程空间分布示意 Fig. 1 Spatial distribution of the meteorological station and elevation of the study area |
MODIS NDVI数据主要来源于美国国家航空航天局发布的MOD13A3 C6数据(https://ladsweb.modaps.eosdis.nasa.gov/), 时间分辨率为每月, 空间分辨率为1 km, 时间跨度为2000年2月~2020年12月.MODIS NDVI数据需要进行滤波、镶嵌、重采样、投影转换、格式转换、图像裁剪和最大值合成等预处理, 得到覆盖研究区2000~2020年植被NDVI年最大值时间序列.
气象数据主要来源于国家气象科学数据中心提供的2000~2020年中国2 416个气象站点逐日数据(https://data.cma.cn/), 包括降水、平均气温、最高气温、最低气温、平均相对湿度、日照时数、平均风速、平均气压和地表温度.研究区包含气象站点716个(图 1).对于个别缺失数据, 本研究利用IBM SPSS进行数据缺失插补.通过ANUSPLIN插值模型, 在考虑经度、纬度和高程的基础上生成各气象因子时间序列.再根据气温和降水时间序列, 计算得到寒冷指数, 温暖指数和湿度指数[26].
人口密度数据和夜间灯光数据分别来源于WorldPop发布的Population Density数据(https://www.worldpop.org/)和Harvard Dataverse发布的全球NPP-VIIRS-like扩展时间系列的夜间灯光数据(https://dataverse.harvard.edu/), 时间分辨率均为每年, 空间分辨率均为1 km, 时间跨度均为2000~2020年.DEM数据来源于中国科学院资源环境数据云平台(https://www.resdc.cn/), 数据空间分辨率为1 km.长江流域及其二级流域单元矢量数据来源于国家科技基础条件平台——国家地球系统科学数据中心(http://www.geodata.cn/).
2.2 研究方法 2.2.1 Theil-Sen Median斜率估计和Mann-Kendall显著性检验Theil-Sen Median斜率估计是一种非参数估计方法, 受异常值影响小, 常用于揭示长时间序列植被NDVI变化特征, 其分析结果能够科学且直观地反映植被NDVI在特定时间段内的变化趋势[27, 28].计算公式为:
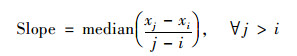
|
(1) |
式中, xi和xj分别为第i和第j年的最大合成植被NDVI值; median为中值函数.若Slope>0, 表示在2000~2020年内植被NDVI呈上升趋势; 若Slope<0, 表示在2000~2020年内植被NDVI呈下降趋势.Slope的绝对值越大, 表示植被NDVI变化趋势越明显.
Mann-Kendall显著性检验法是一种应用在监测时间序列变化趋势的检验方法, 可用来检验时间序列变化趋势的显著性[13, 29].该方法不需要样本服从正态分布, 因此常用于分析长时间序列气候、水文和植被的变化特征.本文采用Mann-Kendall显著性检验法来检验2000~2020年长江流域植被NDVI的变化趋势.对于显著性水平α, 当|Z|>Z1-α/2, 表明时间序列在α水平上存在显著性变化, 反之, 则为不显著变化.本文定义变化趋势在α=0.05下显著时, 为显著变化; 在α=0.01下显著时, 为极显著变化.为探究2000~2020年长江流域植被NDVI变化趋势的显著性, 本文根据植被NDVI变化趋势及Mann-Kendall显著性检验结果[13], 将植被NDVI变化趋势分为6类:极显著上升、显著上升、不显著上升、不显著下降、显著下降和极显著下降.
2.2.2 稳定性分析变异系数为数据标准差和数据平均值的比值, 可用来反映数据在时间序列上的稳定性和波动状况[30].本文在像元尺度上计算了2000~2020年长江流域植被NDVI的变异系数, 用于评估植被NDVI在时间序列上的稳定性.计算公式如下:
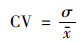
|
(2) |
式中, CV为植被NDVI变异系数; σ为植被NDVI标准差; x为植被NDVI平均值, CV值越大, 表示数据分布越离散, 植被NDVI年际变化越大; 反之, 则表示数据分布紧凑, 植被NDVI年际变化较为稳定.根据自然间断点法[31]将植被NDVI的稳定性分成5类:低波动变化(0.006~0.049)、较低波动变化(0.049~0.100)、中度波动变化(0.100~0.185)、较高波动变化(0.185~0.392)和高波动变化(0.392~1).
2.2.3 地理探测器地理探测器是探测地理现象空间分异并揭示其驱动力的一组统计学方法[19].本文主要使用因子探测、交互作用探测和生态探测来探究长江流域植被NDVI空间分异的影响因素.因子探测主要用于研究属性Y的空间分异性及影响因素X对属性Y的空间分异性的解释力度, 以q值度量.
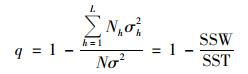
|
(3) |
式中, h为变量Y或因子X的分层, Nh和N分别为层h和全区的单元数, Nh2和σ2分别是层h和全区的Y值的方差, SSW和SST分别为层内方差之和和全区总方差.q的值域为[0, 1], q值越大, 表示X对Y的解释力越大.
交互作用主要用于探究因子与因子之间是否存在相互影响及在相互作用下对属性Y的影响程度变化.生态探测是比较因子与因子交互作用下对Y的空间分异的作用是否有显著性差异.
长江流域植被NDVI空间分异是多种因子共同作用的结果, 综合考虑人文因子、地形因子和气候因子, 本文选取18种影响因素(表 1), 以格网为基本单元, 对长江流域植被NDVI进行影响因素影响力探测, 因子探测的q值越大, 表明该因子在对植被NDVI空间分布的解释力越强.
|
|
表 1 影响因素类别 Table 1 Category of influencing factors |
3 结果与分析 3.1 长江流域植被NDVI时空变化特征 3.1.1 长江流域植被NDVI时间变化特征
由图 2可知, 整体上, 2000~2020年长江流域植被NDVI呈波动上升趋势, 上升斜率为0.002 1 a-1.流域尺度上, 除太湖水系外, 各流域单元植被NDVI均呈上升趋势, 其中乌江植被NDVI上升趋势最为显著, 为0.003 6 a-1, 嘉陵江植被NDVI上升速率次之, 为0.003 4 a-1, 湖口以下干流植被NDVI上升趋势最为缓慢, 仅为0.000 6 a-1.太湖水系植被NDVI呈下降趋势, 下降速率为-0.002 3 a-1.汉江多年平均植被NDVI最高, 为0.805, 其次为宜宾至宜昌, 为0.790, 最低为金沙江石鼓以上, 为0.497.综上可知, 2000~2020年长江流域植被NDVI整体呈改善态势.除太湖水系外, 各流域单元植被NDVI整体上均呈上升趋势, 其中, 汉江和宜宾至宜昌植被NDVI增长趋势最为显著.汉江和宜宾至宜昌整体植被覆盖程度最好, 太湖水系和金沙江石鼓以上整体植被覆盖程度较低.

|
图 2 2000~2020年长江流域植被NDVI时间变化趋势 Fig. 2 Temporal variation in NDVI in the Yangtze River basin from 2000 to 2020 |
如图 3所示, 2000~2020年长江流域植被NDVI变化斜率在-0.034 9~0.039 7 a-1之间.由表 2可知, 植被NDVI呈上升趋势的面积占84.09%, 其中, 呈极显著上升和显著上升的面积占53.67%, 主要分布在嘉陵江、乌江、宜宾至宜昌、汉江和洞庭湖水系.植被NDVI呈下降趋势的面积占15.91%, 其中, 呈极显著下降和显著下降的面积占3.50%, 主要分布在岷沱江西部的川渝地区、汉江、宜昌至湖口和洞庭湖水系交界处、湖口以下干流和太湖水系.

|
图 3 2000~2020年长江流域植被NDVI变化趋势及显著性检验 Fig. 3 Spatial variation in NDVI and its significance test in the Yangtze River basin from 2000 to 2020 |
|
|
表 2 长江流域植被NDVI显著性变化面积占比统计/% Table 2 Statistical results of significance test of NDVI in the Yangtze River basin/% |
由表 2可知, 除太湖水系外, 其他流域单元植被NDVI呈上升趋势的面积均大于呈下降趋势的面积.乌江、宜宾至宜昌、汉江、嘉陵江和鄱阳湖水系植被NDVI呈极显著上升的面积均超过其总面积的50%.而太湖水系植被NDVI呈下降趋势的面积占其总面积的59.11%, 其中, 呈极显著下降和显著下降的区域占其总面积的30.95%.在研究时段内, 除太湖水系植被NDVI呈下降趋势的面积大于呈上升趋势的面积外, 其余流域单元植被NDVI呈上升趋势的面积均大于呈下降趋势的面积, 其中, 位于长江流域中游的乌江、宜宾至宜昌、汉江和嘉陵江的植被改善区域面积和强度均明显高于其他流域单元.
3.1.3 长江流域植被NDVI稳定性分析如图 4所示, 2000~2020年长江流域植被NDVI整体稳定性较好, 平均变异系数为0.058.低波动变化和较低波动变化的区域占研究区总面积的90.19%, 主要分布在金沙江石鼓以下、岷沱江、嘉陵江、乌江、宜宾至宜昌、洞庭湖水系、鄱阳湖水系、汉江和湖口以下干流; 中等波动变化、较高波动变化和高波动变化的区域占研究区总面积的9.81%, 主要分布在金沙江石鼓以上和太湖水系.

|
图 4 长江流域植被NDVI变异系数空间分布 Fig. 4 Spatial distribution of the coefficient of variation of NDVI in the Yangtze River basin |
流域尺度上, 金沙江石鼓以上和太湖水系植被NDVI稳定性较差, 变异系数分别为0.092和0.097, 远高于其他流域单元, 其中, 金沙江石鼓以上和太湖水系植被NDVI呈中等波动变化、较高波动变化和高波动变化的区域分别占其总面积的33.83%和36.72%, 说明以上两个流域植被NDVI变化明显, 波动性较大, 植被覆盖稳定性较差.其余流域单元植被NDVI稳定性较好, 平均变异系数在0.046~0.067之间, 其中, 金沙江石鼓以下、岷沱江、嘉陵江、宜宾至宜昌、乌江、汉江、宜昌至湖口和洞庭湖水系植被NDVI呈低波动变化和较低波动变化的区域分别占其总面积的90%以上.由上可知, 2000~2020年长江流域整体植被覆盖较为稳定, 但位于长江流域上游的金沙江石鼓以上和长江流域下游的太湖水系植被覆盖变化较大, 稳定性较差.
3.2 长江流域植被NDVI影响因素地理探测 3.2.1 影响因素影响力探测由表 3可知, 海拔和气候因子(湿度指数除外)的q值均大于0.40, 说明以上因子是影响长江流域植被NDVI空间分异的主要因素.坡度、坡向、曲率、湿度指数、夜间灯光和人口密度的q值均在0.1以下, 说明以上因子对植被NDVI空间分异的影响微弱.综上可知, 海拔和气候因子对长江流域植被生长的影响程度较强, 而坡度、坡向、曲率、湿度指数、夜间灯光和人口密度对长江流域植被生长的影响程度较弱.
|
|
表 3 长江流域及各流域单元影响因素探测q值 Table 3 The q values of influencing factors in the Yangtze River basin and its sub-basin |
由地理探测器结果可知, 人文因子、地形因子和气候因子对不同流域单元植被生长的影响表现出较大的差异.地形因子对各流域单元影响力呈现出明显的地域差异, 除乌江(q=0.02)以外, 海拔对各流域单元的q值均较高, 而坡向对所有流域单元的q值均接近0, 说明海拔对各流域单元植被NDVI空间分异的影响较大, 而坡向对植被NDVI空间分异的影响微弱.气候因子对金沙江石鼓以上植被NDVI空间分异的影响程度较高, 而对其余流域单元影响程度较低.夜间灯光和人口密度对乌江、湖口以下干流和太湖水系植被NDVI影响较大, 特别是太湖水系, 夜间灯光和人口密度的q值分别为0.62和0.45, 远高于其他影响因素.综上可知, 海拔是影响长江流域各流域单元植被生长的重要因素, 而气候因子对金沙江石鼓以上植被NDVI的影响程度最高, 人文因子对乌江、湖口以下干流和太湖水系植被NDVI影响最大.
3.2.2 影响因素交互作用探测如表 4所示, 由交互作用探测结果可知, 长江流域影响因素双因子作用q值均大于单因子q值, 各影响因素交互作用均表现为双因子增强或非线性增强, 其中, 海拔∩风速的q值最高, 为0.68, 由此可知, 海拔和风速的交互作用是影响长江流域植被NDVI空间分异的主要因素.由表 5可知, 在流域尺度上, 金沙江石鼓以上、金沙江石鼓以下、岷沱江和嘉陵江的海拔与气候因子交互作用的q值最大.除乌江夜间灯光∩气压和夜间灯光∩降水和湖口以下干流以海拔∩夜间灯光和海拔∩人口密度交互作用的q值最大以外, 宜宾至宜昌、汉江、宜昌至湖口、洞庭湖水系、鄱阳湖水系和太湖水系均是夜间灯光∩海拔交互作用的q值最大.由上可知, 除乌江以外, 各流域单元海拔与其余因子交互作用的q值最大, 这也进一步说明了海拔对长江流域植被生长的影响力.
|
|
表 4 长江流域影响因素交互作用探测 Table 4 Interaction detector of influencing factors in the Yangtze River basin |
|
|
表 5 流域单元影响因素交互作用探测最大q值统计 Table 5 Statistical results of the maximum q value of interaction detector of influencing factors in sub-basin of the Yangtze River basin |
综上可知, 长江各流域单元植被生长影响因素存在明显的地域差异, 乌江和宜宾至宜昌植被NDVI空间分异主要受人文、地形和气候因子的共同作用, 以其为过渡带, 由长江流域上游至过渡带的流域单元植被NDVI空间分异主要受地形和气候因子的共同作用, 由过渡带至长江流域下游的流域单元植被NDVI空间分异主要受地形与人文因子的共同作用.
3.2.3 影响因素生态探测长江流域生态探测结果如表 6所示, 58%的因子组合对长江流域植被NDVI变化的解释力存在显著性差异, 42%的因子组合对长江流域植被NDVI变化的解释力不存在显著性差异.其中, 人文因子(夜间灯光和人口密度)与除坡度以外的地形因子和除湿度指数以外的气候因子对长江流域植被NDVI空间分异的解释力存在显著性差异.海拔与除坡向、坡度和曲率以及温暖指数、气压、降水、相对湿度和湿度指数以外的影响因子对长江流域植被NDVI空间分异的解释力存在显著性差异.坡度、坡向和曲率与除湿度指数以外的气候因子对长江流域植被NDVI空间分析的解释力均存在显著性差异.由此可知, 人文因子和地形因子是影响长江流域植被NDVI空间分异的主要因子, 这与影响因素探测和双因子交互探测的结果高度吻合.
|
|
表 6 长江流域影响因素生态探测显著性统计1) Table 6 Statistical results of the ecological detector of influencing factors in the Yangtze River basin |
基于流域单元生态探测结果表明, 因子与因子间生态探测的显著性呈现明显空间异质性.金沙江石鼓以上、嘉陵江、乌江和太湖水系海拔与其他因子对其植被NDVI空间分异的解释力存在明显差异, 而其余流域单元海拔与其余因子对其植被NDVI空间分异的解释力均不呈显著性差异.宜宾至宜昌、乌江、洞庭湖水系、鄱阳湖水系和太湖水系夜间灯光和人口密度与气候因子间均对植被NDVI空间分布的解释力不存在显著性差异, 而大部分流域夜间灯光与人口密度与坡度和坡向对其植被NDVI空间分布的解释力存在显著性差异.以上结果与影响因素探测和双因子交互探测得到的海拔、夜间灯光和人口密度等因子与长江流域各流域单元植被NDVI具有较强相关性的结果一致.
4 讨论 4.1 长江流域植被NDVI时空演变特征从时间尺度上看, 2000~2020年长江流域植被NDVI整体呈上升趋势.流域尺度上, 除太湖水系外, 其余流域单元植被NDVI均呈上升趋势, 且乌江和嘉陵江植被NDVI上升速率高于其他流域单元, 这与已有的研究结果一致[14, 16, 18, 32~34].从空间尺度上看, 植被NDVI呈上升趋势的区域主要分布在乌江、宜宾至宜昌、汉江、嘉陵江、洞庭湖水系和鄱阳湖水系.以上流域植被类型以阔叶林、灌丛和栽培植被为主, 植被群落较为稳定, 抵御风险的能力较强, 植被覆盖程度较高, 植被稳定性较好.自2000年以来, 得益于长江流域防护林工程、岩溶地区石漠化综合治理工程、天然林保护工程和退耕还林还草工程等林业生态工程的相继实施, 以及洞庭湖湿地生态环境保护政策和鄱阳湖流域山江湖生态治理措施的实行和三峡水电站的开发和水库群的建设, 使得以上流域生态环境质量得到改善, 促进了植被群落的繁衍和演替进程, 从而提高了植被覆盖程度, 这与已有的研究结果一致[14, 16, 35~37].
植被NDVI呈下降趋势的区域主要分布在岷沱江西部的川渝地区, 汉江、宜昌至湖口和洞庭湖水系三大流域交界处, 湖口以下干流和太湖水系.以上地区主要位于成渝经济圈、武汉城市圈、长株潭城市群和长江三角洲经济圈, 随着经济水平的不断提高和城市化进程的日益加快, 导致以上地区建筑用地急剧扩展, 持续占用林地、草地和农用地, 使得区域植被覆盖程度下降, 植被NDVI呈下降趋势.除此以外, 流域单元内高强度的人类活动和农业活动导致部分地区土壤质地变差、土壤肥力流失、土壤侵蚀和水土流失加剧, 从而导致植被NDVI下降, 植被稳定性较差, 这与已有的研究结果较一致[13, 16, 36, 38, 39].金沙江石鼓以上植被稳定性较差, 主要由于该流域为国家最大的水电基地, 加之部分地区受流域地质状况、地貌类型、气候条件和人类活动的共同影响, 水土流失严重, 生态环境敏感且脆弱, 因此, 流域内部分地区植被NDVI呈下降趋势, 植被稳定性较差, 这与已有的研究结果一致[40, 41].
4.2 长江流域植被NDVI影响因素分析海拔、温度、日照时数和风速是影响长江流域整体植被生长的关键因素.海拔变化是引起植被NDVI空间分异的主要原因, 长江流域海拔高度由西向东逐级递减, 逐渐由“第一级阶梯”过渡到“第三级阶梯”.植被生长主要受区域水分、热量和光照等条件的影响, 而风速、温度和日照时数等气候因子与海拔高度息息相关.且已有研究表明, 北半球增温能够引起植被生长季的起点的提前以及生长季的延长, 从而促进植被生长[42], 故海拔、温度、日照时数和风速对长江流域整体植被NDVI空间分异影响较大.从流域单元上看, 影响长江流域植被NDVI变化的影响因素存在明显地域差异.长江流域中上游的金沙江石鼓以下、岷沱江、宜宾至宜昌、汉江、宜昌至湖口、洞庭湖水系和鄱阳湖水系植被生长主要受海拔影响, 而乌江、湖口以下干流和太湖水系植被生长主要受人类活动(夜间灯光)的影响, 这与已有的研究结果一致[18, 30, 43~45].乌江为长江流域和珠江流域防护林工程等林业生态工程的覆盖区, 但位于乌江的贵阳及其周边城市经济发展迅速, 城区面积急剧扩张, 使得部分地区植被NDVI呈下降趋势.湖口以下干流和太湖水系地处长江三角洲核心地带, 随着经济水平和城市化水平的不断提高, 城市建设面积的不断扩张, 持续占用植被资源, 地表植被覆盖锐减, 部分植被NDVI呈下降趋势.夜间灯光可用于表征区域经济发展状况和人口空间分布, 由此可知, 人类活动对以上3个流域单元植被NDVI变化影响显著, 这与已有的研究结果较一致[18, 32, 34].
交互作用探测结果表明, 长江流域各流域单元植被生长影响因素存在明显的地域差异, 以乌江和宜宾至宜昌为过渡带, 以上两个流域植被NDVI空间分异主要受人文、地形和气候条件的共同作用, 由长江流域上游至过渡带的流域单元的植被NDVI空间分异主要受地形和气候条件的共同作用, 由过渡带至长江流域下游的流域单元的植被NDVI空间分异主要受地形与人文因素的共同作用.受长江流域防护林等林业生态工程的影响, 长江流域整体植被覆盖较高[46], 但长江流域上游流域海拔较高, 人口稀疏, 这使得地形因素和气候因素对长江流域上游植被生长影响较大[16].而中游各流域单元地处“第二级阶梯”到“第三级阶梯”的过渡带, 地形起伏较大, 气候条件多变, 人类活动强度逐渐加强, 因此, 植被生长受人文、地形和气候条件的共同作用.而下游各流域单元植被类型主要为栽培植被, 且温暖湿润的气候条件以及纵横交错的水系和星罗漫布的湖泊和水库降低了植被生长对气候变化的敏感性, 加之以上各流域地势平坦, 经济发达, 人类活动频繁[43~45, 47], 因此, 长江流域下游各流域单元植被分异主要受地形和人文因子的共同影响.
5 结论(1) 2000~2020年长江流域植被NDVI呈波动上升趋势, 上升斜率为0.002 1 a-1.流域尺度上, 除太湖水系植被NDIV呈下降趋势外, 其余流域单元植被NDVI均呈上升趋势, 其中乌江和嘉陵江植被NDVI上升趋势最为显著.长江流域植被NDVI变化斜率呈现出明显的空间异质性, 整体呈“中部高, 两翼低”的空间分布格局.
(2) 2000~2020年长江流域整体植被覆盖较为稳定.位于长江流域上游的金沙江石鼓以上和长江流域下游的太湖水系植被覆盖变化较大, 稳定性较差, 而金沙江石鼓以下、岷沱江、嘉陵江、宜宾至宜昌、乌江、汉江、宜昌至湖口和洞庭湖水系植被覆盖较为稳定.
(3) 影响因素影响力探测结果表明, 日照时数、寒冷指数、风速、温度和海拔是影响长江流域植被生长的主导因素.影响因素交互作用探测结果表明, 长江流域双因子作用q值均大于单因子q值, 各影响因素交互作用结果表现为双因子增强或非线性增强, 其中, 海拔∩风速的q值最高, 为0.68.生态探测结果表明, 58%的因子组合对长江流域植被NDVI变化的解释力存在显著性差异, 42%的因子组合对长江流域植被NDVI变化的解释力不存在显著性差异.
| [1] | Jiao W Z, Wang L X, Smith W K, et al. Observed increasing water constraint on vegetation growth over the last three decades[J]. Nature Communications, 2021, 12(1), doi: 0.1038/s41467-021-24016-9. |
| [2] | Cheng J, Wu H B, Liu Z Y, et al. Vegetation feedback causes delayed ecosystem response to East Asian Summer Monsoon Rainfall during the Holocene[J]. Nature Communications, 2021, 12(1). DOI:10.1038/s41467-021-22087-2 |
| [3] | Slater S M, Twitchett R J, Danise S, et al. Substantial vegetation response to Early Jurassic global warming with impacts on oceanic anoxia[J]. Nature Geoscience, 2019, 12(6): 462-467. DOI:10.1038/s41561-019-0349-z |
| [4] | Xu C G, McDowell N G, Fisher R A, et al. Increasing impacts of extreme droughts on vegetation productivity under climate change[J]. Nature Climate Change, 2019, 9(12): 948-953. DOI:10.1038/s41558-019-0630-6 |
| [5] |
帅爽, 张志, 吕新彪, 等. 矿山恢复治理区植被物候与健康状况遥感监测[J]. 农业工程学报, 2021, 37(4): 224-234. Shuai S, Zhang Z, Lyu X B, et al. Remote sensing monitoring of vegetation phenological characteristics and vegetation health status in mine restoration areas[J]. Transactions of the Chinese Society of Agricultural Engineering, 2021, 37(4): 224-234. |
| [6] | Forzieri G, Miralles D G, Ciais P, et al. Increased control of vegetation on global terrestrial energy fluxes[J]. Nature Climate Change, 2020, 10(4): 356-362. DOI:10.1038/s41558-020-0717-0 |
| [7] | Aalto J, Niittynen P, Riihimäki H, et al. Cryogenic land surface processes shape vegetation biomass patterns in northern European tundra[J]. Communications Earth & Environment, 2021, 2(1). DOI:10.1038/s43247-021-00292-7 |
| [8] |
何宝忠, 丁建丽, 张喆, 等. 新疆植被覆盖度趋势演变实验性分析[J]. 地理学报, 2016, 71(11): 1948-1966. He B Z, Ding J L, Zhang Z, et al. Experimental analysis of spatial and temporal dynamics of fractional vegetation cover in Xinjiang[J]. Acta Geographica Sinica, 2016, 71(11): 1948-1966. |
| [9] | Macedo P M S, Oliveira P T S, Antunes M A H, et al. New approach for obtaining the C-factor of RUSLE considering the seasonal effect of rainfalls on vegetation cover[J]. International Soil and Water Conservation Research, 2021, 9(2): 207-216. DOI:10.1016/j.iswcr.2020.12.001 |
| [10] | Zhumanova M, Mönnig C, Hergarten C, et al. Assessment of vegetation degradation in mountainous pastures of the Western Tien-Shan, Kyrgyzstan, using eMODIS NDVI[J]. Ecological Indicators, 2018, 95: 527-543. DOI:10.1016/j.ecolind.2018.07.060 |
| [11] | Yan Y B, Chai Z Y, Yang X S, et al. The temporal and spatial changes of the ecological environment quality of the urban agglomeration on the northern slope of Tianshan Mountain and the influencing factors[J]. Ecological Indicators, 2021, 133. DOI:10.1016/j.ecolind.2021.108380 |
| [12] | Zhe M, Zhang X Q. Time-lag effects of NDVI responses to climate change in the Yamzhog Yumco Basin, South Tibet[J]. Ecological Indicators, 2021, 124. DOI:10.1016/J.ECOLIND.2021.107431 |
| [13] |
徐勇, 黄雯婷, 窦世卿, 等. 2000~2020年西南地区植被NDVI对气候变化和人类活动响应特征[J]. 环境科学, 2022, 43(6): 3230-3240. Xu Y, Huang W T, Dou S Q, et al. Responding mechanism of vegetation cover to climate change and human activities in southwest China from 2000 to 2020[J]. Environmental Science, 2022, 43(6): 3230-3240. |
| [14] | Qu S, Wang L C, Lin A W, et al. What drives the vegetation restoration in Yangtze River basin, China: climate change or anthropogenic factors?[J]. Ecological Indicators, 2018, 90: 438-450. DOI:10.1016/j.ecolind.2018.03.029 |
| [15] | Zhang W, Wang L C, Xiang F F, et al. Vegetation dynamics and the relations with climate change at multiple time scales in the Yangtze River and Yellow River Basin, China[J]. Ecological Indicators, 2020, 110. DOI:10.1016/j.ecolind.2019.105892 |
| [16] | Chen S S, Wen Z F, Zhang S L, et al. Effects of long-term and large-scale ecology projects on forest dynamics in Yangtze River Basin, China[J]. Forest Ecology and Management, 2021, 496. DOI:10.1016/J.FORECO.2021.119463 |
| [17] |
崔利芳, 王伦澈, 屈赛, 等. 气温、降水量和人类活动对长江流域植被NDVI的影响[J]. 地球科学, 2020, 45(6): 1905-1917. Cui L F, Wang L C, Qu S, et al. Impacts of temperature, precipitation and human activity on vegetation NDVI in Yangtze river basin, China[J]. Earth Science, 2020, 45(6): 1905-1917. |
| [18] |
崔浩楠, 罗海江, 张学珍. 1982—2019年长江经济带植被覆盖变化的时空特征[J]. 生态学杂志, 2021, 40(8): 2517-2529. Cui H N, Luo H J, Zhang X Z. Temporal and spatial characteristics of green vegetation cover changes in the Yangtze River Economic Belt from 1982 to 2019[J]. Chinese Journal of Ecology, 2021, 40(8): 2517-2529. |
| [19] |
王劲峰, 徐成东. 地理探测器: 原理与展望[J]. 地理学报, 2017, 72(1): 116-134. Wang J F, Xu C D. Geodetector: principle and prospective[J]. Acta Geographica Sinica, 2017, 72(1): 116-134. |
| [20] | Han J J, Wang J P, Chen L, et al. Driving factors of desertification in Qaidam Basin, China: an 18-year analysis using the geographic detector model[J]. Ecological Indicators, 2021, 124. DOI:10.1016/j.ecolind.2021.107404 |
| [21] | Sun Y F, Guan Q Y, Wang Q Z, et al. Quantitative assessment of the impact of climatic factors on phenological changes in the Qilian Mountains, China[J]. Forest Ecology and Management, 2021, 499. DOI:10.1016/j.foreco.2021.119594 |
| [22] | Peng W F, Kuang T T, Tao S. Quantifying influences of natural factors on vegetation NDVI changes based on geographical detector in Sichuan, western China[J]. Journal of Cleaner Production, 2019, 233: 353-367. DOI:10.1016/j.jclepro.2019.05.355 |
| [23] | Liu C L, Li W L, Wang W Y, et al. Quantitative spatial analysis of vegetation dynamics and potential driving factors in a typical alpine region on the northeastern Tibetan Plateau using the Google Earth Engine[J]. CATENA, 2021, 206. DOI:10.1016/J.CATENA.2021.105500 |
| [24] |
周明星, 李登秋, 邹建军. 基于稠密Landsat数据的邛崃山大熊猫栖息地植被变化研究[J]. 植物生态学报, 2021, 45(4): 355-369. Zhou M X, Li D Q, Zou J J. Vegetation change of giant panda habitats in Qionglai Mountains through dense Landsat Data[J]. Chinese Journal of Plant Ecology, 2021, 45(4): 355-369. |
| [25] |
潘洪义, 黄佩, 徐婕. 基于地理探测器的岷江中下游地区植被NPP时空格局演变及其驱动力研究[J]. 生态学报, 2019, 39(20): 7621-7631. Pan H Y, Huang P, Xu J. The spatial and temporal pattern evolution of vegetation NPP and its driving forces in middle-lower areas of the Min river based on geographical detector analyses[J]. Acta Ecologica Sinica, 2019, 39(20): 7621-7631. |
| [26] |
谢春平, 吴显坤, 薛晓明, 等. 浙江楠适生区与气候环境关系的分析[J]. 四川农业大学学报, 2020, 38(3): 264-271. Xie C P, Wu X K, Xue X M., et al. Analysis on relationship between suitable habitat of Phoebe chekiangensis and climatic environmental factors[J]. Journal of Sichuan Agricultural University, 2020, 38(3): 264-271. |
| [27] | Sen P K. Estimates of the regression coefficient based on Kendall's tau[J]. Journal of the American Statistical Association, 1968, 63(324): 1379-1389. DOI:10.1080/01621459.1968.10480934 |
| [28] |
赵伟, 高博, 卢清, 等. 2006~2019年珠三角地区臭氧污染趋势[J]. 环境科学, 2021, 42(1): 97-105. Zhao W, Gao B, Lu Q, et al. Ozone pollution trend in the Pearl River Delta Region during 2006-2019[J]. Environmental Science, 2021, 42(1): 97-105. |
| [29] |
杨玉莲, 杨昆, 罗毅, 等. 1998~2016中国八大经济区植被覆盖对PM2.5浓度时空分布的影响[J]. 环境科学, 2021, 42(11): 5100-5108. Yang Y L, Yang K, Luo Y, et al. Effect of vegetation coverage on the temporal and spatial distribution of PM2.5 concentration in China's Eight Major Economic Regions from 1998 to 2016[J]. Environmental Science, 2021, 42(11): 5100-5108. |
| [30] |
李华林, 张建军, 张耀方, 等. 基于不同赋权方法的北运河上游潜在非点源污染风险时空变化特征分析[J]. 环境科学, 2021, 42(6): 2796-2809. Li H L, Zhang J J, Zhang Y F, et al. Analysis of spatial-temporal variation characteristics of potential non-point source pollution risks in the Upper Beiyun River basin using different weighting methods[J]. Environmental Science, 2021, 42(6): 2796-2809. |
| [31] |
刘彦随, 李进涛. 中国县域农村贫困化分异机制的地理探测与优化决策[J]. 地理学报, 2017, 72(1): 161-173. Liu Y S, Li J T. Geographic detection and optimizing decision of the differentiation mechanism of rural poverty in China[J]. Acta Geographica Sinica, 2017, 72(1): 161-173. |
| [32] |
石悦樾, 银正彤, 郑文锋. 基于MODIS数据乌江流域植被覆盖变化与气候变化关系研究[J]. 林业资源管理, 2017(1): 127-134. Shi Y Y, Yin Z T, Zheng W F. Study on the response of vegetation cover change and climate change in Wujiang River basin based on MODIS data[J]. Forest Resources Management, 2017(1): 127-134. |
| [33] |
张婷, 薛东剑, 段金亮, 等. 2000~2019嘉陵江流域植被覆盖时空变化特征及气候响应分析[J]. 长江流域资源与环境, 2021, 30(5): 1110-1120. Zhang T, Xue D J, Duan J L, et al. Spatio-temporal variation characteristics and climate response analysis of vegetation coverage in Jialing River basin from 2000 to 2019[J]. Resources and Environment in the Yangtze Basin, 2021, 30(5): 1110-1120. |
| [34] | Jiang L G, Liu Y, Wu S, et al. Analyzing ecological environment change and associated driving factors in China based on NDVI time series data[J]. Ecological Indicators, 2021, 129. DOI:10.1016/J.ECOLIND.2021.107933 |
| [35] |
刘栩位, 周启刚, 周浪, 等. 基于RSEI的三峡库区重庆段水土保持生态功能区生态环境质量动态监测[J]. 水土保持研究, 2021, 28(5): 278-286. Liu X W, Zhou Q G, Zhou L, et al. RSEI—Based dynamic monitoring of ecological quality of the soil and water conservation functional area in the Chongqing section of the three gorges reservoir area[J]. Research of Soil and Water Conservation, 2021, 28(5): 278-286. |
| [36] |
张亮, 丁明军, 张华敏, 等. 1982—2015年长江流域植被覆盖度时空变化分析[J]. 自然资源学报, 2018, 33(12): 2084-2097. Zhang L, Ding M J, Zhang H M, et al. Spatiotemporal variation of the vegetation coverage in Yangtze River basin during 1982-2015[J]. Journal of Natural Resources, 2018, 33(12): 2084-2097. |
| [37] | Zhang P P, Cai Y P, Yang W, et al. Contributions of climatic and anthropogenic drivers to vegetation dynamics indicated by NDVI in a large dam-reservoir-river system[J]. Journal of Cleaner Production, 2020, 256. DOI:10.1016/j.jclepro.2020.120477 |
| [38] | Wang J Y, Delang C O, Hou G L, et al. Net primary production increases in the Yangtze River Basin within the latest two decades[J]. Global Ecology and Conservation, 2021, 26. DOI:10.1016/J.GECCO.2021.E01497 |
| [39] |
冯磊, 杨东, 黄悦悦. 2000—2017年川渝地区植被NDVI特征及其对极端气候的响应[J]. 生态学杂志, 2020, 39(7): 2316-2326. Feng L, Yang D, Huang Y Y. Vegetation NDVI characteristics and response to extreme climate in Sichuan and Chongqing from 2000 to 2017[J]. Chinese Journal of Ecology, 2020, 39(7): 2316-2326. |
| [40] |
姜丽光, 姚治君, 王蕊, 等. 金沙江梯级水电开发区NDVI时空变化及其驱动因子研究[J]. 资源科学, 2014, 36(11): 2431-2441. Jiang L G, Yao Z J, Wang R, et al. Spatio-temporal variation of NDVI change and driving forces in the cascade hydropower development zone of the Jinsha River[J]. Resources Science, 2014, 36(11): 2431-2441. |
| [41] |
周长艳, 王顺久, 彭骏. 金沙江流域及邻近地区空中水资源的气候特征分析[J]. 资源科学, 2010, 32(12): 2433-2440. Zhou C Y, Wang S J, Peng J. Climatic characteristics of water vapor resources over the Jinsha River reaches and its surroundings[J]. Resources Science, 2010, 32(12): 2433-2440. |
| [42] | Xu X Y, Riley W J, Koven C D, et al. Earlier leaf-out warms air in the north[J]. Nature Climate Change, 2020, 10(4): 370-375. DOI:10.1038/s41558-020-0713-4 |
| [43] |
刘珞丹, 李晶, 柳彩霞, 等. 2000-2015年长江经济带植被覆盖时空变化特征及影响因素分析[J]. 水土保持研究, 2021, 28(6): 330-336, 347. Liu L D, Li J, Liu C X, et al. Analysis on the characteristics of temporal and spatial changes and influencing factors of vegetation coverage in the Yangtze River Economic Belt from 2000 to 2015[J]. Research of Soil and Water Conservation, 2021, 28(6): 330-336, 347. |
| [44] |
潘颖, 林杰, 佟光臣, 等. 2004—2014年太湖流域植被覆盖时空变化特征[J]. 生态与农村环境学报, 2018, 34(1): 37-45. Pan Y, Lin J, Tong G C, et al. Spatio-temporal variations of vegetation coverage in Taihu Lake Basin from 2004 to 2014[J]. Journal of Ecology and Rural Environment, 2018, 34(1): 37-45. |
| [45] |
陶帅, 邝婷婷, 彭文甫, 等. 2000—2015年长江上游NDVI时空变化及驱动力——以宜宾市为例[J]. 生态学报, 2020, 40(14): 5029-5043. Tao S, Kuang T T, Peng W F, et al. Analyzing the spatio-temporal variation and drivers of NDVI in upper reaches of the Yangtze River from 2000 to 2015: a case study of Yibin City[J]. Acta Ecologica Sinica, 2020, 40(14): 5029-5043. |
| [46] | Kong R, Zhang Z X, Zhang F Y, et al. Increasing carbon storage in subtropical forests over the Yangtze River basin and its relations to the major ecological projects[J]. Science of the Total Environment, 2020, 709. DOI:10.1016/j.scitotenv.2019.136163 |
| [47] | Yang H F, Zhong X N, Deng S Q, et al. Assessment of the impact of LUCC on NPP and its influencing factors in the Yangtze River basin, China[J]. CATENA, 2021, 206. DOI:10.1016/j.catena.2021.105542 |