 2022, Vol. 43
2022, Vol. 43

2. 福建师范大学湿润亚热带生态地理过程教育部重点实验室, 福州 350007;
3. 福州大学环境与安全工程学院, 福州 350116
 , TONG Chuan1,2 , LUO Min3
, TONG Chuan1,2 , LUO Min3 , YANG Yang1,2 , TAN Feng-feng1,2 , PAN Zhe-yan1,2 , LIU Na1,2 , CHEN Xin3 , HUANG Jia-fang1,2
, YANG Yang1,2 , TAN Feng-feng1,2 , PAN Zhe-yan1,2 , LIU Na1,2 , CHEN Xin3 , HUANG Jia-fang1,2
2. Key Laboratory of Humid Subtropical Eco-geographical Process, Ministry of Education, Fujian Normal University, Fuzhou 350007, China;
3. College of Environment and Safety Engineering, Fuzhou University, Fuzhou 350116, China
河口潮滩湿地, 如红树林湿地、盐沼湿地和海草湿地, 所占面积不到海洋面积的0.5%, 但所固存的碳含量达到海洋沉积物碳固存总量的50%[1], 在全球碳循环中起着重要作用[2~4].土壤有机碳(soil organic carbon, SOC)库是土壤质量的重要指标[5].土壤有机碳通过影响土壤的物理、化学和生物特性, 对河口潮滩湿地生态系统服务功能产生深远的影响[6, 7].在全球土壤碳库及其相关的碳循环研究中, 陆地生态系统土壤碳库和碳循环研究比较深入[8~10], 但对于海陆交界处, 受潮汐水文影响的河口潮滩湿地的研究较单薄[11].已有研究也多集中在河口潮滩湿地SOC储量[12], 鲜见河口潮滩湿地有机碳活性组分和稳定性机制等方面的相关报道.
土壤有机碳根据土壤团聚体颗粒大小分级的方法将有机碳划分为颗粒性有机碳(particulate organic carbon, POC, >53 μm)和矿物结合态有机碳(mineral-associated organic carbon, MAOC, < 53 μm)[13, 14].土壤MAOC主要是由微生物分解产物与矿质土壤基质的强化学结合而形成, 能防止有机碳快速分解[15].因此, 土壤MAOC属于惰性有机碳库[16].相较之下, 土壤POC易受到环境变化的影响, 属于活性有机碳库[17].已有研究中常用碳库稳定性指数(carbon stability index, CSI), 即POC和MAOC的比值来表征土壤有机碳稳定性强弱[13].此外, 虽然活性有机碳组分一般占SOC不到50%, 但对土壤碳循环过程具有重要的影响[18].微生物生物量碳(microbial biomass carbon, MBC)、易分解有机碳(liable organic carbon, LOC)和溶解性有机碳(dissolved organic carbon, DOC)是土壤活性有机碳库的3个重要组分[19, 20], 其对变化的环境因子的响应比SOC更加敏感[21].以上活性组分受理化环境影响, 对于滨海潮滩湿地的碳循环过程有着十分重要的作用[22].
淹水梯度是河口潮滩湿地生态系统中的一个重要环境变量因子, 它控制着河口潮滩湿地与海洋系统之间水文、能量和营养物质的交换[23, 24].目前已经开展淹水梯度的研究包括:净初级生产力、微生物分解的速率、生态系统CO2交换通量和生态系统服务功能的影响等[12, 25~30].淹水梯度增加将显著降低河口潮滩湿地土壤的通气状况, 并导致微生物活性受到抑制, 使得枯落物分解与转化过程变缓, 从而对土壤有机碳的积累和分解产生影响[31].此外, 潮水中的阳离子如:Na+、K+、Ca2+和Mg2+等, 可通过离子交换加速团聚体解聚, 从而破坏土壤有机碳的稳定性[31, 32].目前, 尚未有淹水梯度上土壤有机碳的活性组分和稳定性机制变化的报道.了解淹水梯度对河口潮滩湿地土壤有机碳组分的影响, 对于阐明河口潮滩生态系统碳循环、迁移和转化规律具有重要意义.
鉴于此, 本文选取福建九龙江河口潮滩湿地的潮滩剖面为研究对象, 目的是探究河口潮滩湿地淹水梯度上:①有机碳碳库(SOC)储量的变化; ②活性有机碳组分MBC、DOC和LOC的含量变化; ③矿物结合态有机碳(MAOC)含量、颗粒性有机碳(POC)含量和碳库稳定性指数(CSI)的变化.通过明晰淹水梯度下河口潮滩湿地土壤有机碳及其组分的空间异质性, 以期为厘清淹水频率对河口潮滩湿地土壤碳动态的影响机制提供科学依据.
1 材料与方法 1.1 研究区概况九龙江河口潮滩湿地位于(24°22′10″N~24°28′43″N, 117°53′21″E~117°55′35″E)我国东南福建沿海[33], 面积达6 000 hm2, 属南亚热带海洋性季风气候, 该地区年平均降水量为1 400~1 800 mm, 年平均气温为19~21℃.该潮滩的潮汐为典型半日潮, 平均潮差从河口上部到河口下部为2.7~4.0 m[34].研究区(甘文尾)位于九龙江河口潮滩湿地内的龙海市红树林自然保护区.沿着河口潮滩滩面由陆向海方向依次分布着秋茄(Kandelia obovata)红树林群落、秋茄和互花米草交错带、互花米草(Spartina alterniflora)群落和光滩等4个生境(图 1).潮滩剖面的平均淹水频率由4%增加至25%(图 1), 淹水频率由Model 3001水文计记录(Solinst有限公司, 加拿大).红树林群落位于高潮滩, 交错带和互花米草群落位于中潮滩, 光滩位于低潮滩.

|
图 1 九龙江河口潮滩湿地采样点示意 Fig. 1 Study area and sampling sites in the Jiulong River estuary |
于2019年12月进行野外采样.在红树林群落、交错带、互花米草群落和光滩等4个不同生境各选取1个典型样地(图 1), 每个样地S形采集4个土样(重复).采样时, 先去除土壤地表的凋落物, 然后用内直径为10 cm的土钻采集40 cm土柱, 并分为0~20 cm和20~40 cm两层.将土样分别装入样品袋, 挤出袋子内部空气以密封保存, 将样品装入放有冰袋的保温箱中带回实验室备用.原位测定土壤电导、pH和氧化还原电位(ORP).在实验室先混匀各样品, 然后取部分土样测定土壤含水率和容重.将剩余土样过60目筛, 去除土样中的残余根系和砾石.将过筛后的鲜土样分成2份, 一份用于测定土壤微生物生物量碳(MBC)和可溶性有机碳(DOC)含量, 需4℃冷藏保存.另一份用于测定易分解有机碳(LOC)、颗粒性有机碳(POC)、矿物结合态有机碳(MAOC)和土壤有机碳(SOC)含量, 需自然风干后研磨过筛.
1.3 测定方法土壤pH值采用pH450便携式pH计(Spectrum Technologies, 美国)原位测定; 土壤氧化还原电位ORP运用Ag/AgCl参比电极的FJA-6型ORP仪(南京传滴仪器设备有限公司, 检测限=0.1 mV)原位测定; 电导率EC运用EC450便携式电导仪(Spectrum Technologies, 美国)原位测定; 容重和含水率采用烘干称重法测定; 土壤粒径采用粒径分析仪(马尔文帕纳科, MasterSizer 2000, 英国)测定.
土壤SOC在土壤酸化后采用土壤碳氮元素分析仪(艾力蒙塔贸易有限公司; Elementar Vario MAX, 德国)测定; 即先用浓度为0.5 mol·L-1的HCl去除土壤中的无机碳, 用蒸馏水洗至中性后风干研磨上机测定.
土壤DOC采用无二氧化碳水浸提法测定[35], 具体操作为:称取5 g过筛后的鲜土样, 依次加入25 mL的无二氧化碳水(沸腾后自然冷却的去离子水)浸提, 浸提液使用0.45 μm玻璃纤维滤膜过滤, 样品于24 h内上机测定.测定仪器采用TOC-L CPH总有机碳分析仪(岛津, 日本, 检测限为2 μmol·L-1).
土壤MBC采用氯仿熏蒸法与0.5 mol·L-1 K2SO4浸提法测定[36].具体操作为:用25 mL玻璃烧杯称取5 g鲜土进行熏蒸, 同时设置对照(不熏蒸).熏蒸样品需在干燥皿中放入氯仿(烧杯底部铺有一层玻璃珠)及0.2 mol·L-1的NaOH溶液, 并保持真空密封.二者均在室温下避光培养24 h.培养完成后分别用20 mL浓度为0.5 mol·L-1的K2SO4溶液浸提, 上清液过0.45 μm的玻璃纤维滤膜(滤膜在使用前经马弗炉400℃烘2 h).液体样品于48 h内采用TOC-L CPH总有机碳分析仪(岛津, 日本, 检测限为2 μmol·L-1)测定有机碳含量.则土壤MBC的计算如公式(1):
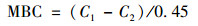
|
(1) |
式中, C1为熏蒸样有机碳含量(g·kg-1); C2为未熏蒸样有机碳含量(g·kg-1); MBC为所测土壤MBC含量(mg·kg-1).
土壤LOC采用KMnO4氧化法测定[37].具体步骤为:2 g过10目筛风干土样, 混合25 mL浓度为333 mmol·L-1的KMnO4溶液, 振荡使其充分反应, 而后离心提取上清液.加入去离子水稀释上清液(样水比为1 ∶250), 稀释液需于2 h内上机测定.测定仪器为UV-1800紫外可见分光光度计(岛津, 日本), 对应比色波段为565 nm.
土壤POC与MAOC采用湿筛法测定[38].具体步骤为:10 g风干土样(已过10目筛, W0), 加入50 mL浓度为5 g·L-1的六偏磷酸钠溶液, 室温25℃速度90 r·min-1振荡18 h使其充分混合反应, 振荡后的混合液搅匀后过300目筛, 并用蒸馏水反复冲洗筛上物质至土壤颗粒无法下渗.收集筛上物质于100℃烘箱烘干, 确保土样完全烘干至恒重并称重(W1), 研磨过筛备用.筛下的混合液静置24 h后离心, 用蒸馏水冲洗数次至上清液澄清, 将沉淀物烘干称重(W2), 研磨过筛备用.使用土壤碳氮元素分析仪(艾力蒙塔贸易有限公司; Elementer Vario MAX CN, 德国)分别测定筛上、筛下物质有机碳含量.则土壤POC和MAOC含量计算依次如公式(2)和公式(3):
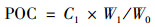
|
(2) |
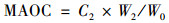
|
(3) |
式中, C1为筛上物质有机碳含量(g·kg-1); C2为筛下物质有机碳含量(g·kg-1); W1为筛上颗粒物干重(g); W2为筛下沉淀物干重(g); W0即为10(g); POC即为所测土壤POC含量(g·kg-1); MAOC即为所测土壤MAOC含量(g·kg-1).
1.4 稳定性指数土壤碳库稳定性指数(carbon stability index, CSI)通常由矿物结合态有机碳含量和颗粒性有机碳含量的比值来表征[13, 14], 如公式(4):
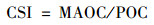
|
(4) |
式中, MAOC为矿物结合态有机碳含量(g·kg-1); POC为颗粒性有机碳含量(g·kg-1).其中, 土壤矿物结合态有机碳MAOC的相对含量越高, 土壤稳定性越强.
1.5 数据处理文中所有数据分析均采用SPSS 25.0软件完成.首先检验数据的正态分布和方差齐次性, 不符合正态分布的数据需进行置换而后再分析.检验不同生境及土层深度对土壤有机碳组分及稳定性系数的影响采用双因素方差分析(Two-way ANOVA).双因素分析有显著差异结果的数据进行Tukey事后检验分析, 以明晰相同处理下的组间差异.不同有机碳组分及占比与理化因子之间的相关性采用Pearson相关分析检验.数据分析显著性均保证P=0.05, 文中所有数据均以平均值(mean)±标准差(standard deviation)的形式呈现.
2 结果与分析 2.1 淹水梯度上不同生境的理化指标分析九龙江河口潮滩湿地土壤理化性质如表 1所示.土壤电导率EC的变化范围为(6.85±0.29)~(14.00±1.94)mS·cm-1, 不同生境间土壤电导率EC有显著差异(P<0.001), 光滩和互花米草生境的电导率EC高于红树林和交错带.不同深度土层之间土壤电导率EC无显著差异.土壤pH的变化范围为(5.00±0.03)~(7.00±0.12), 土壤pH值随着淹水频率增加而增大(P<0.001), 光滩0~20 cm土层的pH高于20~40 cm土层, 其他生境不同深度土层之间pH无明显差异.土壤氧化还原电位ORP处于(-255.91±8.81)~(-151.62±3.46)mV, 土壤氧化还原电位ORP随着淹水频率增加而减小(P<0.001). 0~20 cm土层的氧化还原电位ORP含量均高于20~40 cm土层(P<0.001).土壤容重处于(0.59±0.07)~(0.87±0.07)g·cm-3, 光滩和互花米草的容重要显著高于红树林和交错带(P<0.001).含水率处于(43.36±0.67)% ~(53.88±2.27)%, 随着淹水频率的增加, 土壤含水率也逐渐增加(P<0.001).不同生境下土壤粒径均以粉粒为主[(81.50±0.93)% ~(82.55± 2.28)%], 黏粒[(9.97±0.27)% ~(16.10±0.48%)]次之, 砂粒[(2.40±0.72)% ~(8.20±0.44)%]最少.由陆向海, 土壤黏粒占比随着淹水频率增加而减少, 而砂粒占比显著随着淹水频率的增加而增加(P<0.001), 粉粒含量无显著变化.不同深度土层之间土壤容重、含水率、黏粒、粉粒和砂粒的含量均无明显区别.
|
|
表 1 九龙江河口潮滩湿地土壤理化性质1) Table 1 Soil physicochemical properties in the tidal wetland of the Jiulong River estuary |
2.2 淹水梯度上不同生境的土壤有机碳总量的差异
九龙江河口潮滩湿地土壤ω(SOC)的变化范围为(8.63±0.07)~(21.08±1.13)g·kg-1(图 2).不同生境之间土壤SOC储量差异显著, 整体表现为:红树林>交错带>互花米草>光滩(P<0.001).红树林、交错带和互花米草0~20 cm土层的SOC储量均高于20~40 cm土层(P<0.001); 而光滩不同深度土层之间土壤SOC储量无显著差异(图 2).

|
不同大写字母表示不同生境之间差异显著(P<0.05); 不同星号表示同一生境下不同深度土层之间差异显著(*表示P<0.05, **表示P<0.01, ***表示P<0.001), 下同 图 2 九龙江河口潮滩湿地土壤SOC储量的变化 Fig. 2 Change in the soil organic carbon storage in the tidal wetland of the Jiulong River estuary |
土壤ω(MBC)变化范围为(24.49±3.63)~(123.63±7.47)mg·kg-1[图 3(a)], 土壤MBC/SOC变化范围为0.3% ~0.6%[图 3(b)].土壤MBC含量和MBC/SOC均随着淹水频率增加而减少[P<0.001, 图 3(a)和3(b)].不同深度土层之间MBC含量和MBC/SOC差异显著, 0~20 cm土层的MBC含量和MBC/SOC均高于20~40 cm土层[P<0.001, 图 3(a)和3(b)].

|
图 3 九龙江河口潮滩湿地土壤有机碳活性组分及其在SOC的占比的变化 Fig. 3 Change in active soil organic carbon component contents and their proportions to SOC storage in the tidal wetland of the Jiulong River estuary |
土壤ω(DOC)变化范围为(44.20±0.66)~(58.32±2.72)mg·kg-1[图 3(c)], 土壤DOC/SOC变化范围为0.3% ~0.5%[图 3(d)].土壤DOC含量随着淹水频率的增加逐渐减少, 但DOC/SOC随着淹水频率增加而增大[P<0.001, 图 3(c)和3(d)].从深度上看, 0~20 cm土层DOC含量显著高于20~40 cm土层(P<0.05), 不同深度土层之间DOC/SOC无显著差异[图 3(c)和3(d)].
土壤ω(LOC)变化范围为(2.74±0.15)~(6.87±0.32)g·kg-1[图 3(e)], 土壤LOC/SOC变化范围为24% ~40%[图 3(f)].土壤LOC含量随着淹水频率增加而减少, 而LOC/SOC随着淹水频率增加而增大[P<0.001, 图 3(e)和3(f)].从深度上看, 0~20 cm土层的LOC含量和LOC/SOC均显著高于20~40 cm土层[P<0.01, 图 3(e)和3(f)].
2.4 淹水梯度上不同生境的土壤颗粒性有机碳、矿物结合态有机碳和碳库稳定性指数的差异土壤ω(POC)变化范围为(1.28±0.26)~(8.80±1.16)g·kg-1[图 4(a)], 土壤ω(MAOC)变化范围为(7.35±0.30)~(12.28±0.49)g·kg-1[图 4(b)].土壤POC含量和MAOC含量均随着淹水频率增加而减少(P<0.001, 图 4).红树林生境下0~20 cm土层的POC含量和MAOC含量均大于20~40 cm土层(P<0.05, 图 4).而其他生境下不同深度土层之间POC含量和MAOC含量均无显著差异(图 4).

|
图 4 九龙江河口潮滩剖面土壤POC和MAOC含量的变化 Fig. 4 Changes in the soil POC and MAOC in the tidal wetland of the Jiulong River estuary |
土壤有机碳稳定性指数CSI变化范围为(1.42±0.24)~(6.10±2.02)(图 5)土壤CSI随着淹水频率增加而增加(P<0.001, 图 5).红树林生境下0~20 cm土层的CSI小于20~40 cm土层(P<0.01, 图 5), 而其他生境下不同深度土层之间土壤CSI均无显著差异(图 5).

|
图 5 九龙江河口潮滩剖面土壤有机碳稳定性指数CSI的变化 Fig. 5 Changes in the soil carbon stability indexes in the tidal wetland of the Jiulong River estuary |
土壤有机碳各组分之间的相关性分析如表 2所示. SOC、MBC、DOC、LOC、POC和MAOC等不同碳组分之间均呈显著正相关(r≥0.762, n=32, P<0.01).土壤有机碳各组分和占比与理化因子之间的相关性分析如图 6所示. MBC/SOC与土壤电导率EC呈显著负相关关系[P<0.01, 图 6(a)], 表明潮滩湿地海向盐度变化会影响微生物活性. LOC/SOC与土壤氧化还原电位ORP呈显著负相关关系[P<0.01, 图 6(b)], 说明土壤LOC受氧化还原环境影响.土壤POC与土壤pH呈显著负相关关系[P<0.01, 图 6(c)], 该结果表明淹水梯度上POC的减少是由于土壤pH增加而引起. 土壤MAOC与土壤黏粒呈显著正相关关系[P<0.01, 图 6(d)], 该结果表明淹水梯度上MAOC的减少是由于淹水梯度上土壤黏粒含量的下降造成的.
|
|
表 2 土壤有机碳各组分之间的相关性分析1) Table 2 Correlation between the organic carbon components |

|
图 6 有机碳组分和占比与土壤理化性质之间的相关分析 Fig. 6 Correlation between the organic carbon components, their proportion and soil physicochemical properties |
淹水频率是影响河口潮滩湿地固碳潜力的关键环境因子之一[12].本研究选取我国东南沿海九龙江河口潮滩湿地作为研究对象, 分析不同淹水频率下SOC储量的变化.本研究结果表明, 表层(0~20 cm)SOC的储量均随着淹水频率的增加而减少59%, 深层(20~40 cm)SOC的储量均随着淹水频率的增加而逐渐减少48%(图 2).有研究发现, 在美国乔治亚海岸潮滩湿地, 低潮滩相对于高潮滩土壤有机碳储量减少25%[39]; 在中国江苏省大丰自然保护区潮滩湿地, 由陆向海土壤有机碳储量减少57%[40]; 在中国崇明岛东滩湿地, 土壤SOC储量随着淹水频率的增加而减少57%[41].以上研究中土壤SOC储量在空间上均与本研究表现出相同的分布特征, 即:高潮滩>中潮滩>低潮滩.SOC储量河口潮滩湿地土壤中的有机碳主要来自本地植物如:地上植物凋落物、地下根系的残体和根系分泌物等[42~46].在九龙江河口潮滩湿地由陆向海方向上, 分别为秋茄红树林群落、红树林互花米草交错带、互花米草群落和无植被的光滩(图 1).有研究表明, 红树林生境地表年凋落物产量可达618.79 g·(m2·a)-1[47], 而互花米草生境地表年凋落物产量仅为139.30 g·(m2·a)-1[48], 由于红树植物根系发达, 具有高归还率, 年凋落物产量和碳输入量远高于红树林互花米草交错带生境和互花米草生境[42, 46].因此, 高潮滩(红树林)SOC储量(19.02 g·kg-1)显著高于中潮滩交错带(12.55 g·kg-1)和互花米草(12.07 g·kg-1)(图 2), 且有植被的生境SOC储量均显著高于无植被的光滩SOC储量(8.75 g·kg-1).由此, 随着淹水频率的增加, SOC储量逐渐减少.在红树林、交错带和互花米草生境中, 表层土壤的SOC储量均高于深层土壤(图 2), 这是由于有植被覆盖的表层土壤接收来自地上凋落物的输入, 因此, 其SOC储量在深度方向上分异显著[49].光滩生境中的表层和深层土壤SOC储量差异并不显著(图 2), 这是由于光滩壤淋溶作用最强, 故SOC储量在深度方向上变化不显著.
3.2 土壤有机碳活性组分沿淹水梯度分布特征土壤微生物量碳(MBC)、溶解性有机碳(DOC)和易分解有机碳(LOC)均是重要的SOC活性组分, 它们对环境因子的响应比土壤SOC储量更加敏感[21].土壤LOC是含量最高的活性组分, 其次为DOC和MBC(图 3).高灯州等[42]在闽江河口潮汐湿地不同淹水频率下土壤有机碳组分的研究中发现, 随着淹水频率的增加, 土壤MBC、DOC和LOC含量分别减少了49%、63%和69%.Li等[40]在东南沿海盐沼湿地土壤固碳的研究中发现, 由陆向海土壤MBC含量减少94%, DOC含量减少58%.本研究发现SOC活性组分均随着淹水频率的增加而减少, MBC、DOC和LOC含量分别减少了80%、24%和60%[图 3(a)、3(c)和3(e)], 这与上述研究的结果一致.根系分泌物和植物凋落物是土壤SOC活性组分的主要碳源, 故导致土壤SOC活性组分沿淹水梯度分布特征与SOC储量一致.此外, 土壤MBC的含量与DOC(r=0.843, n=32, P<0.01, 表 2)和LOC(r=0.935, n=32, P<0.01, 表 2)的含量呈现极显著的正相关, 说明土壤LOC和DOC均为河口潮滩湿地土壤微生物潜在的碳源.
有机碳活性组分的占比如MBC/SOC、DOC/SOC和LOC/SOC, 均可反映土壤碳库的循环转化过程.研究结果表明, 随着淹水频率的增加, 土壤MBC/SOC下降29%[图 3(b)], 而DOC/SOC和LOC/SOC分别增加80%和26%[图 3(d)和3(f)], 说明当淹水频率增加后, 活性组分中DOC和LOC的相对累积速率较快, 而MBC的相对累积速率相对变缓.土壤DOC/SOC主要受潮滩水动力的调节, 随着淹水频率的增加, 水动力增强, 加速土壤有机质的淋溶, 导致土壤DOC的累积加快.土壤LOC/SOC与土壤氧化电位ORP呈现显著的负相关关系[r=-0.355, n=32, P<0.01, 图 6(b)], 这说明LOC的周转受到氧化还原电位ORP的调节.随着淹水频率的增加, 土壤氧化还原电位ORP逐渐减少(表 1).因此, 与代谢相关的微生物活性迅速减少, 造成LOC/SOC随着淹水频率的增加而累积[图 3(f)].随着淹水频率的增加, 土壤电导率EC由6.85 mS·cm-1增加到14.00 mS·cm-1(表 1), 同时, 土壤MBC/SOC与土壤电导率EC呈现显著负相关关系[r=-0.497, n=32, P<0.01, 图 6(a)].说明土壤的盐度的增加会增强微生物的离子胁迫, 促进有毒的低价HS-或Fe2+离子的累积, 抑制微生物活性[31].因此, 淹水频率增加造成的土壤盐胁迫引起土壤MBC的产生速率降低.
3.3 土壤颗粒有机碳、矿物结合态有机碳和有机碳稳定性指数沿淹水梯度分布特征本研究中, 土壤颗粒有机碳(POC)的含量随着淹水梯度的增加而减少81%, 矿物结合态有机碳(MAOC)的含量随着淹水梯度的增加而减少35%(图 4).淹水梯度上土壤POC的含量下降, 可能是由于多种因素共同作用的结果.首先, 随着淹水频率的增加, 强烈的水动力作用将严重破坏土壤团聚体结构[50], 降低团聚体胶结物质与能力[21].其次, 土壤POC含量的减少可能是由于土壤pH增加引起的.本研究发现从高潮滩至光滩, 土壤pH由5.00逐渐增加至7.00(表 1), 且土壤POC含量与pH呈现显著负相关关系[r=-0.822, n=32, P<0.01, 图 6(c)].土壤pH值增大伴随着土壤团聚体吸附有机质的能力降低[51], 即土壤pH值增加会限制土壤大团聚体的结合能力, 尤其是导致大团聚体的吸附减少, 进而减少POC的含量[52].本研究发现黏粒含量沿着淹水频率的增加逐渐减少34%(表 1).这主要是由于近海区域水动力作用强烈, 大量黏粒随着“潮泵”作用被潮水带至附近海域中[32].土壤MAOC的含量与土壤黏粒含量呈现正相关关系[r=0.678, n=32, P<0.01, 图 6(d)], 该结果与盐沼土壤的研究结果一致[53~55].这主要是由于MAOC是封闭或者吸附在微团聚体内部矿物上的有机质, 而河口潮滩湿地土壤中的矿物主要分布在黏粒中[56].因此, 与粗粒土(如砂粒含量和黏粒含量)相比, 细粒的黏土含量与河口潮滩湿地土壤中MAOC的相关性更高[57].因此, 淹水频率增加后, 由于黏粒含量大量减少, 因此导致河口潮滩湿地MAOC的含量也逐渐减少.
对土壤稳定性指数CSI的研究结果表明, 在九龙江河口潮滩湿地, 随着淹水频率的增加, 土壤CSI增加246%, 土壤有机碳稳定性增强.土壤CSI始终大于1.00(图 5), 说明矿物结合态有机碳含量始终大于颗粒性有机碳含量, 矿物结合态有机碳始终是九龙江河口潮滩湿地土壤有机碳的主要赋存形态.土壤CSI由红树林生境的1.76逐渐增加为光滩的6.02, 说明相较于微团聚体, 大团聚体受淹水增加扰动更为敏感[56].因此, 受大团聚体保护的、不稳定的POC首当其冲被破坏释放, 损失远高于相对稳定的MAOC, 大团聚体有机碳逐渐向微团聚体转移, 受微团聚体保护的MAOC则逐渐成为土壤有机碳固存的主要形式, 土壤有机碳稳定性逐渐增强.
3.4 河口潮滩湿地生境、土壤碳库和稳定性对未来海平面上升的响应20世纪60年代以来, 人类频繁的工业活动导致大气中温室气体浓度急剧增加, 并引起全球气候变暖, 最直接的后果之一为全球海平面上升[58].仅2006~2018年间, 海平面上升速率可达3.2~4.2 mm·a-1[59].海平面上升已成为21世纪人类面临的重大环境问题, 九龙江河口潮滩湿地生境理化特征、土壤碳库、有机碳活性组分以及有机碳稳定性沿着淹水梯度的变化见图 7.本文预测未来海平面上升将对河口潮滩湿地土壤理化特征和蓝碳产生影响.首先, 河口潮滩湿地土壤理化性质将发生改变, 土壤pH增加, 氧化还原电位ORP减少, 土壤中黏粒的含量也会随着海平面上升-淹水增加而减少.其次, 随着湿地植物生物量的减少, 土壤SOC储量也随之减少.有机碳的活性组分中, DOC和LOC的占比会增加, 而MBC的占比会减少.MBC的占比减少也暗示着微生物丰度也会随着淹水频率的增加而减少.MAOC是河口潮滩湿地最重要的有机碳赋存形式, 随着海平面上升-淹水频率和程度的增加, MAOC的占比将增加, 土壤稳定性指数CSI增加, 说明随着海平面上升(淹水增加)后, 河口潮滩湿地土壤有机碳的稳定性将增加.

|
图 7 河口潮滩湿地有机碳储量、活性组分和稳定性指数沿淹水梯度分布 Fig. 7 Conceptual diagram of soil organic carbon storage, active components and carbon stability index along a flooding gradient in an estuarine tidal wetland |
(1) 淹水频率对河口潮滩湿地SOC储量有显著影响.主要是由于淹水频率增加导致的植被生物量减少影响SOC储量.
(2) 随着淹水频率的增加, 有机碳的活性组分溶解性有机碳(DOC)、微生物生物量碳(MBC)和易分解有机碳(LOC)均呈现减少的趋势, 其中DOC/SOC和LOC/SOC增加, 而MBC/SOC减少, 说明活性组分中DOC和LOC的累积速率提高, 而MBC的累积速率相对减少.
(3) 随着淹水频率的增加, 颗粒性有机碳(POC)和矿物结合态有机碳(MAOC)也呈现减少的趋势, 土壤POC的减少与pH的增加有关, 土壤MAOC的减少与黏粒含量的减少有关.
(4) 随着淹水频率的增加, 土壤有机碳稳定性指数CSI增加, 矿物结合态有机碳(MAOC)是土壤有机碳的主要存在形态.随着淹水频率的增加, 土壤有机碳稳定性指数会持续增大, 土壤有机碳稳定性增强.
| [1] | Duarte C M. Reviews and syntheses: hidden forests, the role of vegetated coastal habitats in the ocean carbon budget[J]. Biogeosciences, 2017, 14(2): 301-310. DOI:10.5194/bg-14-301-2017 |
| [2] | Doetterl S, Stevens A, Six J, et al. Soil carbon storage controlled by interactions between geochemistry and climate[J]. Nature Geoscience, 2015, 8(10): 780-783. DOI:10.1038/ngeo2516 |
| [3] | Braun K N, Theuerkauf E J, Masterson A L, et al. Modeling organic carbon loss from a rapidly eroding freshwater coastal wetland[J]. Scientific Reports, 2019, 9(1). DOI:10.1038/s41598-019-40855-5 |
| [4] | Spivak A C, Sanderman J, Bowen J L, et al. Global-change controls on soil-carbon accumulation and loss in coastal vegetated ecosystems[J]. Nature Geoscience, 2019, 12(9): 685-692. DOI:10.1038/s41561-019-0435-2 |
| [5] | Benbi D K, Brar K, Toor A S, et al. Sensitivity of labile soil organic carbon pools to long-term fertilizer, straw and manure management in rice-wheat system[J]. Pedosphere, 2015, 25(4): 534-545. DOI:10.1016/S1002-0160(15)30034-5 |
| [6] | Bongiorno G, Bünemann E K, Oguejiofor C U, et al. Sensitivity of labile carbon fractions to tillage and organic matter management and their potential as comprehensive soil quality indicators across pedoclimatic conditions in Europe[J]. Ecological Indicators, 2019, 99: 38-50. DOI:10.1016/j.ecolind.2018.12.008 |
| [7] | Culman S W, Snapp S S, Freeman M A, et al. Permanganate oxidizable carbon reflects a processed soil fraction that is sensitive to management[J]. Soil Science Society of America Journal, 2012, 76(2): 494-504. DOI:10.2136/sssaj2011.0286 |
| [8] |
徐广平, 李艳琼, 沈育伊, 等. 桂林会仙喀斯特湿地水位梯度下不同植物群落土壤有机碳及其组分特征[J]. 环境科学, 2019, 40(3): 1491-1503. Xu G P, Li Y Q, Shen Y Y, et al. Soil organic carbon distribution and components in different plant communities along a water table gradient in the Huixian Karst wetland in Guilin[J]. Environmental Science, 2019, 40(3): 1491-1503. |
| [9] |
罗晓虹, 王子芳, 陆畅, 等. 土地利用方式对土壤团聚体稳定性和有机碳含量的影响[J]. 环境科学, 2019, 40(8): 3816-3824. Luo X H, Wang Z F, Lu C, et al. Effects of land use type on the content and stability of organic carbon in soil aggregates[J]. Environmental Science, 2019, 40(8): 3816-3824. |
| [10] |
刘琪, 李宇虹, 李哲, 等. 不同水分条件和微生物生物量水平下水稻土有机碳矿化及其影响因子特征[J]. 环境科学, 2021, 42(5): 2440-2448. Liu Q, Li Y H, Li Z, et al. Characteristics of paddy soil organic carbon mineralization and influencing factors under different water conditions and microbial biomass levels[J]. Environmental Science, 2021, 42(5): 2440-2448. |
| [11] | Gattuso J P, Magnan A K, Bopp L, et al. Ocean solutions to address climate change and its effects on marine ecosystems[J]. Frontiers in Marine Science, 2018, 5. DOI:10.3389/fmars.2018.00337 |
| [12] | Mueller P, Granse D, Nolte S, et al. Unrecognized controls on microbial functioning in blue carbon ecosystems: the role of mineral enzyme stabilization and allochthonous substrate supply[J]. Ecology and Evolution, 2020, 10(2): 998-1011. DOI:10.1002/ece3.5962 |
| [13] | Chen L Y, Liu L, Qin S Q, et al. Regulation of priming effect by soil organic matter stability over a broad geographic scale[J]. Nature Communications, 2019, 10(1). DOI:10.1038/s41467-019-13119-z |
| [14] | Lugato E, Lavallee J M, Haddix M L, et al. Different climate sensitivity of particulate and mineral-associated soil organic matter[J]. Nature Geoscience, 2021, 14(5): 295-300. DOI:10.1038/s41561-021-00744-x |
| [15] | Cotrufo M F, Ranalli M G, Haddix M L, et al. Soil carbon storage informed by particulate and mineral-associated organic matter[J]. Nature Geoscience, 2019, 12(12): 989-994. DOI:10.1038/s41561-019-0484-6 |
| [16] | Witzgall K, Vidal A, Schubert D I, et al. Particulate organic matter as a functional soil component for persistent soil organic carbon[J]. Nature Communications, 2021, 12(1). DOI:10.1038/s41467-021-24192-8 |
| [17] | Jagadamma S, Essington M E, Xu S T, et al. Total and active soil organic carbon from long-term agricultural management practices in West Tennessee[J]. Agricultural & Environmental Letters, 2019, 4(1). DOI:10.2134/ael2018.11.0062 |
| [18] | Paul E A. The nature and dynamics of soil organic matter: plant inputs, microbial transformations, and organic matter stabilization[J]. Soil Biology and Biochemistry, 2016, 98: 109-126. DOI:10.1016/j.soilbio.2016.04.001 |
| [19] | Yuan G Y, Huan W W, Song H, et al. Effects of straw incorporation and potassium fertilizer on crop yields, soil organic carbon, and active carbon in the rice-wheat system[J]. Soil and Tillage Research, 2021, 209. DOI:10.1016/j.still.2021.104958 |
| [20] | Wang C, Fang Y Y, An W L, et al. Acid rain mediated nitrogen and sulfur deposition alters soil nitrogen, phosphorus and carbon fractions in a subtropical paddy[J]. CATENA, 2020, 195. DOI:10.1016/j.catena.2020.104876 |
| [21] | Luo M, Zeng C S, Tong C, et al. Iron reduction along an inundation gradient in a tidal sedge (Cyperus malaccensis) marsh: the rates, pathways, and contributions to anaerobic organic matter mineralization[J]. Estuaries and Coasts, 2016, 39(6): 1679-1693. DOI:10.1007/s12237-016-0094-0 |
| [22] | Bockelmann A C, Bakker J P, Neuhaus R, et al. The relation between vegetation zonation, elevation and inundation frequency in a Wadden sea salt marsh[J]. Aquatic Botany, 2002, 73(3): 211-221. DOI:10.1016/S0304-3770(02)00022-0 |
| [23] | Kirwan M L, Langley J A, Guntenspergen G R, et al. The impact of sea-level rise on organic matter decay rates in Chesapeake Bay brackish tidal marshes[J]. Biogeosciences, 2013, 10(3): 1869-1876. DOI:10.5194/bg-10-1869-2013 |
| [24] | Langley J A, Mozdzer T J, Shepard K A, et al. Tidal marsh plant responses to elevated CO2, nitrogen fertilization, and sea level rise[J]. Global Change Biology, 2013, 19(5): 1495-1503. DOI:10.1111/gcb.12147 |
| [25] | Morris J T, Sundareshwar P V, Nietch C T, et al. Responses of coastal wetlands to rising sea level[J]. Ecology, 2002, 83(10): 2869-2877. DOI:10.1890/0012-9658(2002)083[2869:ROCWTR]2.0.CO;2 |
| [26] | Redelstein R, Dinter T, Hertel D, et al. Effects of inundation, nutrient availability and plant species diversity on fine root mass and morphology across a saltmarsh flooding gradient[J]. Frontiers in Plant Science, 2018, 9. DOI:10.3389/FPLS.2018.00098 |
| [27] | Ishtiaq K S, Abdul-Aziz O I. Ecological parameter reductions, environmental regimes, and characteristic process diagram of carbon dioxide fluxes in coastal salt marshes[J]. Scientific Reports, 2020, 10. DOI:10.1038/s41598-020-72066-8 |
| [28] | Duarte C M, Losada I J, Hendriks I E, et al. The role of coastal plant communities for climate change mitigation and adaptation[J]. Nature Climate Change, 2013, 3(11): 961-968. DOI:10.1038/nclimate1970 |
| [29] | Neubauer S C. Ecosystem Responses of a tidal freshwater marsh experiencing saltwater intrusion and altered hydrology[J]. Estuaries and Coasts, 2013, 36(3): 491-507. DOI:10.1007/s12237-011-9455-x |
| [30] | Tully K, Gedan K, Epanchin-Niell R, et al. The invisible flood: the chemistry, ecology, and social implications of coastal saltwater intrusion[J]. BioScience, 2019, 69(5): 368-378. DOI:10.1093/biosci/biz027 |
| [31] | Herbert E R, Boon P, Burgin A J, et al. A global perspective on wetland salinization: ecological consequences of a growing threat to freshwater wetlands[J]. Ecosphere, 2015, 6(10): 1-43. |
| [32] | Kim K H, Michael H A, Field E K, et al. Hydrologic shifts create complex transient distributions of particulate organic carbon and biogeochemical responses in beach aquifers[J]. Journal of Geophysical Research: Biogeosciences, 2019, 124(10): 3024-3038. DOI:10.1029/2019JG005114 |
| [33] |
蔡锋, 黄敏芬, 苏贤泽, 等. 九龙江河口湾泥沙运移特点与沉积动力机制[J]. 台湾海峡, 1999, 18(4): 418-418. Cai F, Huang M F, Su X Z, et al. Characteristics of silt movement and sedimentary dynamic mechanism in Jiulongjiang Estuary[J]. Journal of Oceanography in Taiwan Strait, 1999, 18(4): 418-424. DOI:10.3969/j.issn.1000-8160.1999.04.012 |
| [34] | 全球潮汐预报服务平台[EB/OL]. http://global-tide.nmdis.org.cn/, 2021-08-16. |
| [35] | Chantigny M H. Dissolved and water-extractable organic matter in soils: A review on the influence of land use and management practices[J]. Geoderma, 2002, 113(3-4): 357-380. |
| [36] | Jenkinson D S, Brookes P C, Powlson D S. Measuring soil microbial biomass[J]. Soil Biology and Biochemistry, 2004, 36(1): 5-7. DOI:10.1016/j.soilbio.2003.10.002 |
| [37] | Blair G J, Lefroy R D B, Lisle L. Soil carbon fractions based on their degree of oxidation, and the development of a carbon management index for agricultural systems[J]. Australian Journal of Agricultural Research, 1995, 46(7): 1459-1466. DOI:10.1071/AR9951459 |
| [38] | Zhang M K, He Z L. Long-term changes in organic carbon and nutrients of an ultisol under rice cropping in Southeast China[J]. Geoderma, 2004, 118(3-4): 167-179. DOI:10.1016/S0016-7061(03)00191-5 |
| [39] | Więski K, Guo H Y, Craft C B, et al. Ecosystem functions of tidal fresh, brackish, and salt marshes on the Georgia coast[J]. Estuaries and Coasts, 2010, 33(1): 161-169. DOI:10.1007/s12237-009-9230-4 |
| [40] | Li N, Shao T Y, Zhu T S, et al. Vegetation succession influences soil carbon sequestration in coastal alkali-saline soils in southeast China[J]. Scientific Reports, 2018, 8(1). DOI:10.1038/s41598-018-28054-0 |
| [41] |
王红丽, 肖春玲, 李朝君, 等. 崇明东滩湿地土壤有机碳空间分异特征及影响因素[J]. 农业环境科学学报, 2009, 28(7): 1522-1528. Wang H L, Xiao C L, Li C J, et al. Spatial variability of organic carbon in the soil of wetlands in Chongming Dongtan and its influential factors[J]. Journal of Agro-Environment Science, 2009, 28(7): 1522-1528. DOI:10.3321/j.issn:1672-2043.2009.07.033 |
| [42] |
高灯州, 曾从盛, 章文龙, 等. 闽江口湿地土壤有机碳及其活性组分沿水文梯度分布特征[J]. 水土保持学报, 2014, 28(6): 216-221, 227. Gao D Z, Zeng C S, Zhang W L, et al. Spatial distributions of soil organic carbon and active composition along a hydrologic gradient in Min River estuarine wetland[J]. Journal of Soil and Water Conservation, 2014, 28(6): 216-221, 227. |
| [43] | Zhao X C, Rivera-Monroy V H, Farfán L M, et al. Tropical cyclones cumulatively control regional carbon fluxes in Everglades mangrove wetlands (Florida, USA)[J]. Scientific Reports, 2021, 11. DOI:10.1038/s41598-021-92899-1 |
| [44] | Donnelly J P, Bertness M D. Rapid shoreward encroachment of salt marsh cordgrass in response to accelerated sea-level rise[J]. Proceedings of the National Academy of Sciences of the United States of America, 2001, 98(25): 14218-14223. DOI:10.1073/pnas.251209298 |
| [45] | Guan B, Zhang H X, Wang X H, et al. Salt is a main factor shaping community composition of arbuscular mycorrhizal fungi along a vegetation successional series in the Yellow River Delta[J]. CATENA, 2020, 185. DOI:10.1016/j.catena.2019.104318 |
| [46] |
陈桂香, 高灯州, 陈刚, 等. 互花米草入侵对我国红树林湿地土壤碳组分的影响[J]. 水土保持学报, 2017, 31(6): 249-256. Chen G X, Gao D Z, Chen G, et al. Effects of Spartina alterniflora invasion on soil carbon fractions in mangrove wetlands of China[J]. Journal of Soil and Water Conservation, 2017, 31(6): 249-256. |
| [47] |
张雨雪, 胡伟芳, 罗敏, 等. 闽江口秋茄红树林凋落物产量及分解动态[J]. 环境科学学报, 2019, 39(4): 1312-1323. Zhang Y X, Hu W F, Luo M, et al. Litter production and decomposition dynamics of Kandelia obovata from a mangrove swamp in Min River Estuary[J]. Acta Scientiae Circumstantiae, 2019, 39(4): 1312-1323. |
| [48] | Liao C Z, LuoYQ, Fang C M, et al. Litter pool sizes, decomposition, and nitrogen dynamics in Spartina alterniflora-invaded and native coastal marshlands of the Yangtze Estuary[J]. Oecologia, 2008, 156(3): 589-600. DOI:10.1007/s00442-008-1007-0 |
| [49] |
刘国华, 傅伯杰, 吴钢, 等. 环渤海地区土壤有机碳库及其空间分布格局的研究[J]. 应用生态学报, 2003, 14(9): 1489-1493. Liu G H, Fu B J, Wu G, et al. Soil organic carbon pool and its spatial distribution pattern in the Circum Bohai Region[J]. Chinese Journal of Applied Ecology, 2003, 14(9): 1489-1493. |
| [50] | Zhao Q Q, Bai J H, Wang X, et al. Soil organic carbon content and stock in wetlands with different hydrologic conditions in the Yellow River Delta, China[J]. Ecohydrology & Hydrobiology, 2020, 20(4): 537-547. |
| [51] | Rabbi S M F, Minasny B, McBratney A B, et al. Microbial processing of organic matter drives stability and pore geometry of soil aggregates[J]. Geoderma, 2020, 360. DOI:10.1016/j.geoderma.2019.114033 |
| [52] | Avena M J, Koopal L K. Kinetics of humic acid adsorption at solid-water interfaces[J]. Environmental Science & Technology, 1999, 33(16): 2739-2744. |
| [53] | Morrissey E M, Gillespie J L, Morina J C, et al. Salinity affects microbial activity and soil organic matter content in tidal wetlands[J]. Global Change Biology, 2014, 20(4): 1351-1362. |
| [54] | Bai J H, Zhang G L, Zhao Q Q, et al. Depth-distribution patterns and control of soil organic carbon in coastal salt marshes with different plant covers[J]. Scientific Reports, 2016, 6(1). DOI:10.1038/srep34835 |
| [55] | Yang W, An S Q, Zhao H, et al. Impacts of Spartina alterniflora invasion on soil organic carbon and nitrogen pools sizes, stability, and turnover in a coastal salt marsh of eastern China[J]. Ecological Engineering, 2016, 86: 174-182. DOI:10.1016/j.ecoleng.2015.11.010 |
| [56] | Kleber M, Bourg I C, Coward E K, et al. Dynamic interactions at the mineral-organic matter interface[J]. Nature Reviews Earth & Environment, 2021, 2(6): 402-421. |
| [57] | Bucka F B, Pihlap E, Kaiser J, et al. A small-scale test for rapid assessment of the soil development potential in post-mining soils[J]. Soil and Tillage Research, 2021, 211. DOI:10.1016/j.still.2021.105016 |
| [58] | Larsen L, Moseman S, Santoro A E, et al. A complex-systems approach to predicting effects of sea level rise and nitrogen loading on nitrogen cycling in coastal wetland ecosystems[A]. In: American Society of Limnology and Oceanography (Ed. ). Ecological Dissertations in the Aquatic Sciences Symposium Proceedings Ⅷ[M]. The American Society of Limnology and Oceanography, 2010. 67-92. |
| [59] | IPCC. IPCC Climate Change 2021: the physical science basis. The working group I contribution to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change[M]. Cambridge, United Kingdom: Cambridge University Press, 2021: 5-6. |