 2022, Vol. 43
2022, Vol. 43

2. 陕西省循环农业工程技术研究中心, 杨凌 712100
 , ZHANG Zhen-jiao1,2 , ZHAO Ya-ping1,2 , XU Miao-ping1,2 , REN Cheng-jie1,2 , YANG Gai-he1,2 , FENG Yong-zhong1,2 , REN Guang-xin1,2 , WANG Xiao-jiao1,2 , HAN Xin-hui1,2
, ZHANG Zhen-jiao1,2 , ZHAO Ya-ping1,2 , XU Miao-ping1,2 , REN Cheng-jie1,2 , YANG Gai-he1,2 , FENG Yong-zhong1,2 , REN Guang-xin1,2 , WANG Xiao-jiao1,2 , HAN Xin-hui1,2
2. Shaanxi Engineering Research Center of Circular Agriculture, Yangling 712100, China
陆地生态系统的碳库主要存在于大气、植被和土壤中, 其中土壤碳库是植被和大气碳库总和的两倍[1, 2], 直接影响大气中CO2的浓度[3].近年来土壤有机碳库的研究受到了高度关注.根据有机碳分解的难易程度及周转速率, 可以将土壤有机碳库分为活性和惰性有机碳库[4].土壤微生物对活性和惰性有机碳的降解能力会直接影响到土壤有机碳的累积与稳定性[5, 6], 同时微生物分泌的酶在这过程中扮演着重要角色.因此, 土壤微生物-土壤酶-碳库之间复杂的关系直接影响整个陆地生态系统.
土壤有机碳主要来源于植物残体和根系分泌物, 其主要的化学组分有糖类、蛋白质、半纤维素、纤维素、木质素和脂类等[7, 8].土壤中的活性有机碳易被微生物利用并分解[9], 而它的分解也会释放出一定量的CO2[10].有研究表明, 土壤酶和微生物作用参与的生物化学过程使得土壤有机碳形态不断转化是导致土壤碳库及功能改变的关键因素[11].微生物分泌的胞外酶促进土壤有机碳矿化分解, 其中水解酶主要分解如糖类、淀粉和纤维素等物质, 包括β-1, 4-葡萄糖苷酶(BG)和纤维二糖水解酶(CBH)等; 氧化酶主要参与惰性有机碳氮组分(如木质素)氧化分解和土壤腐殖化等过程, 包括酚氧化酶(PO)和过氧化物酶(PER)等[12].因此, 酶活性的高低可表征土壤微生物养分需求以及与养分供给水平之间的关系.有研究表明, 土壤酶活性的动态变化与土壤有机碳存在显著正相关关系[13], 同时发现土壤转化酶与微生物量碳和可溶性有机碳显著相关, 与碳矿化速率趋势具有一致性[10].此外, 土壤酶活性与植被特征、土壤温湿度和pH等因素有关[14, 15], 以上研究结果说明酶活性与生物和非生物因素的复杂性有关.虽然土壤胞外酶活性与碳库组分含量间的关系已被广泛关注, 但是土壤微生物碳降解酶的变化及与碳库组分的响应关系仍不明确.
黄土高原自实施“退耕还林”工程以来生态恢复和水土保持的效应较好, 刺槐因其树种特性成为该地区的主栽树种.人工刺槐林演替过程中, 其类型和生境条件发生动态变化, 势必会影响森林生态系统碳库组分和酶活性的变化.目前, 研究者们对人工刺槐林恢复阶段土壤碳库储量变化和酶活性变化特征进行了大量的研究, 但对于刺槐林恢复阶段土壤中碳库组分及其相关碳降解酶的关系仍然了解不足, 限制了黄土高原退耕还林生态恢复与保护对土壤肥力演变及酶活性的影响机制的认识.为此, 本研究采用空间代替时间的方法, 选取不同演替阶段的人工刺槐林作为对象, 从土壤微生物碳降解酶及相关碳库组分的角度分析恢复阶段微生物碳降解相关酶活性及与土壤碳库转化状况的关系, 结果将有助于深入了解脆弱生境区域土壤碳库转化规律及调控机制, 以期为黄土高原植被恢复和未来经营管理提供科学依据.
1 材料与方法 1.1 研究区概况本研究选择陕西省延安市安塞区五里湾(109°19′~109°22′E, 36°51′~36°53′N)为研究区域, 平均海拔1 371.9 m.该地区属暖温带半干旱大陆性季风气候, 年均气温8.8℃, 年均降雨量为505.3 mm, 降雨多集中于7~9月, 该流域属黄土丘陵沟壑区, 土壤类型主要为黄绵土(Calcaric Cambisols, FAO-UNESCO), 土质疏松, 抗蚀力差, 属最具代表性的黄土侵蚀区之一.在退耕前, 该地区主要种植的作物是玉米, 退耕后, 该地区种植大面积刺槐, 并采取自然经营, 未经间伐抚育.刺槐林下植被主要有异叶败酱(Patrinia heterophylla)、苔草(Carex tristachya)、碱菀(Tripolium pannonicum)和野菊(Dendranthema indicum)等.
1.2 样地选择和采样本试验于2019年8月开展, 试验选取立地条件相似和干扰程度较低的土地, 分别种植14、20、30和45 a的刺槐林(编号为RP14、RP20、RP30和RP45, 林龄通过当地林业部门造林记录和使用生长锥获取的林木生长数据确定进行研究), 各年限刺槐林在造林前均为耕地, 主要种植作物为玉米(Zea mays); 并以农田(FL)作为对照.样地基本特征信息见表 1.在每个年限刺槐林选择3块立地条件相近的重复样地, 使用GIS和坡度仪等仪器进行样地坡向、坡度和海拔的测量并记录; 每个样地内设置大小为20 m×20 m的标准样方, 随机选取样方内15~20棵树, 通过每木检尺法测量不同林龄刺槐的树高和胸径; 并且在样方内随机设置6个1 m×1 m的小样方, 通过目测法计算林下植被覆盖度; 同时将样方内按“S”形采样法选取10个采样点, 去除表面枯落物和植物残体后, 用土钻(直径为5 cm)采集距离树干1 m以外的0~10 cm土壤, 将同一样方内5个样点的土样混匀过2 mm土筛后分成两份, 一份在-20℃冷藏保存用于测定微生物生物量养分和酶活性; 一份在阴凉处风干保存用于测定土壤养分.
|
|
表 1 样地基本特征信息1) Table 1 Basics feature information of the sample site |
1.3 土壤理化性质的测定
土壤有机碳(SOC)含量采用重铬酸钾-浓硫酸外加热容量法测定[16]; 微生物量碳(MBC)氯仿熏蒸培养-K2SO4浸提, TOC仪测定; 土壤惰性有机碳采用浓度为6 mol ·L-1HCl酸水解法[17], 并用重铬酸钾-浓硫酸外加热容量法测定碳含量; 采用环刀法(100 cm3)测定土壤容重.称取5 g土壤样品加入100 mL 0.5 mol ·L-1的(NaPO3)6溶液中, 置于180 r ·min-1旋转式摇床上(25℃)持续振荡6 h, 通过Malvern MS 2000激光粒度仪(Malvern Instruments, 英国)测定土壤黏粒(clay, < 2 μm)、粉粒(silt, 2~50 μm)和砂粒(sand, 50~2 000 μm)的质量分数.
1.4 土壤微生物呼吸和酶活性的测定土壤微生物呼吸即土壤基础呼吸的测定采用碱液吸收法[18].具体操作: 将不同年限刺槐林土壤分别在(25±0.5)℃下预培养7 d[19].在进行培养试验之前, 将20 g鲜土和5 mg石英砂(防止土壤压实)放入一个50 mL的管中, 将含水量调整到土壤持水量的60%.二氧化碳的吸收装置(装有20 mL 2 mol ·L-1 NaOH的50 mL离心管)、去离子水(装有20 mL去离子水的50 mL离心管用于维持恒定湿度)和3个装有20 g鲜土和5 mg石英砂离心管被放置在培养桶中(1 L黑色塑料桶), 3个重复(n=3).另外将3个只装有二氧化碳的吸收装置和去离子水的的培养桶作为空白对照.培养7 d后, 以1%酚酞和甲基橙为pH指示剂, 用1 mol ·L-1 HCl滴定测定CO2排放量.土壤微生物呼吸(MR)单位采用mg ·(kg ·d)-1.
土壤水解酶活性参照Saiya-Cork等[20]描述的96微孔板荧光光度法测定, 其中BG酶底物为4-甲基伞形酮酰-β-D-吡喃葡萄糖苷(4-MUB-β-D-glucopyranoside), CBH酶底物为4-甲基伞形酮酰-β-D-纤维二糖糖苷(4-MUB-β-D-cellobioside), 并在培养结束后使用多功能酶标仪(Tecan Infinite M200, 奥地利)检测荧光值, 激发光和检测波长分别为365 nm和450 nm; 土壤氧化酶活性参考DeForest等[21]描述的方法测定, PO和PER酶底物均采用左旋多巴(L-dihydroxyphenylalanine)和乙二胺四乙酸二钠(EDTA-3Na)1 ∶2混合液, 并在培养结束后使用多功能酶标仪测定460 nm波长时的吸光值.酶活性单位采用nmol ·(g ·h)-1.
1.5 数据处理土壤活性有机碳含量(g ·kg-1)采用公式[22]:
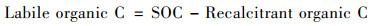
|
(1) |
式中, SOC为土壤有机碳含量(g ·kg-1); Recalcitrant organic C为土壤惰性有机碳含量(g ·kg-1).
土壤微生物熵qMB(mg ·g-1)计算公式[23]:
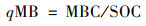
|
(2) |
式中, MBC为土壤微生物量碳含量(mg ·kg-1).
土壤微生物代谢熵qCO2(d-1)计算公式[23]:
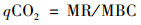
|
(3) |
式中, MR为土壤微生物呼吸[mg ·(kg ·d)-1].
利用Excel 2010和SPSS 23.0软件对试验数据进行统计分析.不同恢复年限土壤物理性质、碳库组分、碳降解酶活性、微生物呼吸及其熵值的差异采用单因素方差分析(one-way ANOVA)进行分析, 所有数据均采用最小显著差异(least significant difference, LSD)法进行多重比较(α=0.05).相关性分析(Correlation analysis)和偏最小二乘路径模型(Partial least squares-path model, PLS-PM)用于土壤物理性质、碳库组分、碳降解酶活性、微生物呼吸及其熵值之间的关系分析.相关性分析和偏最小二乘路径模型使用R软件(4.0.1版本)完成并用Photoshop完成作图.
2 结果与分析 2.1 不同恢复年限刺槐林土壤微生物呼吸及其熵值变化不同恢复年限刺槐林土壤微生物呼吸及其熵值(土壤微生物代谢熵和微生物熵)变化见表 2.随刺槐林龄增加, 土壤微生物量碳不断增加, 在恢复前期(0~14 a)增加并不明显, 但从恢复14~45 a显著增加了6.3倍(P < 0.05); 土壤微生物呼吸在恢复前期(0~14 a)显著增加了1.4倍(P < 0.05), 在20~30 a之间下降了0.4倍(P < 0.05), 在恢复14~20 a并无显著差异(P>0.05); 同时土壤微生物代谢熵随刺槐林龄增加呈先增后减的趋势(P < 0.05), 最大值和最小值出现在14 a和45 a, 从农田到恢复14 a显著增加了1.4倍(P < 0.05), 在恢复14~45 a之间显著下降了0.9倍(P < 0.05); 微生物熵总体上呈现随刺槐林龄增加不断增加的趋势, 但在农田到植被恢复14 a期间下降了0.3倍, 随后在各恢复年限显著增加(P < 0.05).
|
|
表 2 不同恢复年限刺槐林土壤微生物呼吸及其熵值变化 Table 2 Changes in soil microbial respiration and its entropy in Robinia pseudoacacia forests with different restoration years |
2.2 不同恢复年限刺槐林土壤碳库组分变化
不同恢复年限刺槐林土壤的碳库组分变化如图 1所示.土壤有机碳含量随着刺槐林龄的增加而增加, 但在恢复前期(14~20 a)并无显著差异(P>0.05), 在恢复30~45 a之间显著增加了0.3倍(P < 0.05); 土壤活性有机碳总体上呈现增加的趋势, 但在恢复期间(14~30 a)并无显著差异(P>0.05); 土壤惰性有机碳在恢复期间与土壤有机碳呈现相同的趋势, 而在恢复前期(14~20 a)无显著差异(P>0.05).

|
小写字母表示不同林龄刺槐林之间惰性和活性有机碳的差异(P < 0.05), 大写字母表示不同林龄刺槐林之间土壤有机碳的差异(P < 0.05), n=3 图 1 不同恢复年限刺槐林土壤碳库组分变化 Fig. 1 Changes in soil carbon pool components in Robinia pseudoacacia forests with different restoration years |
不同恢复年限刺槐林土壤碳降解酶活性如图 2所示, BG、CBH、PO和PER这4种酶活性的变化范围分别为26.68~91.46、5.06~11.01、1.86~3.15和2.12~3.05 nmol ·(g ·h)-1.农田退耕还林后, BG酶的活性随刺槐林龄的增加显著增加(P < 0.05), 恢复14、20、30和45 a土壤BG酶活性较农田土壤分别增加了0.7、0.9、1.2和2.4倍(P < 0.05); 土壤CBH酶随刺槐林龄的增加表现出先增加后稳定的趋势, 从农田到恢复20 a显著增加了1.1倍(P < 0.05), 随后各恢复年限并无显著差异(P>0.05); PO和PER酶活性均随恢复年限呈先增加后降低趋势, 最大值分别在20 a和14 a, 相比较农田分别增加了0.7倍和0.4倍.

|
小写字母表示不同林龄刺槐林之间的差异(P < 0.05), n=3 图 2 不同恢复年限刺槐林土壤碳降解酶活性的变化 Fig. 2 Changes in soil carbon-degrading enzyme activities in Robinia pseudoacacia forests with different restoration years |
同样, 在植被恢复的过程中这2种水解酶(BG+CBH)和2种氧化酶(PO+PER)呈现不同的变化趋势(图 3).水解酶随刺槐林龄的增加而显著增加(P < 0.05), 并且恢复14、20、30和45 a土壤水解酶相较于农田分别增加了0.7、1.0、1.2和2.2倍(P < 0.05); 而氧化酶在植被恢复期间, 总体上呈现先增加后降低的变化趋势, 其中最大值和最小值分别出现在恢复20 a和农田.

|
小写字母表示不同林龄刺槐林之间的差异(P < 0.05), n=3 图 3 不同恢复年限刺槐林土壤碳降解酶活性的变化 Fig. 3 Changes in soil carbon-degrading enzyme activities in Robinia pseudoacacia forests with different restoration years |
为探讨土壤碳降解酶活性和碳库组分与微生物呼吸及其熵值之间的关系, 进行了相关性分析(图 4).如图 4所示, 刺槐林的恢复对土壤碳库组分、碳降解酶活性和微生物呼吸影响显著.从中可知BG、CBH、PO和PER酶均与黏粒呈显著正相关(P < 0.05), 其中Hy-EEA(BG+CBH)水解酶与土壤有机碳、微生物量碳、微生物熵、惰性和活性有机碳呈显著正相关; 对Oy-EEA(PO+PER)氧化酶而言, 与微生物呼吸呈极显著正相关(P < 0.01).Hy-EEA(BG+CBH)水解酶与黏粒呈显著负相关(P < 0.05), 与容重呈极显著负相关(P < 0.01); PER酶与砂粒呈极显著负相关(P < 0.01).

|
1.黏粒, 2.粉粒, 3.砂粒, 4.容重, 5.土壤有机碳, 6.活性有机碳, 7.惰性有机碳, 8.土壤微生物量碳, 9.土壤微生物呼吸, 10.微生物代谢熵, 11.微生物熵, 12.BG(β-1, 4-葡萄糖苷酶), 13.CBH(纤维二糖水解酶), 14.PO(酚氧化酶), 15.PER(过氧化物酶), 16.Hy-EEA(水解酶), 17.Oy-EEA(氧化酶); 通过曼特尔检验刺槐林造林对土壤碳库组分、碳降解酶活性和微生物呼吸及其熵值相关, 边缘宽度对应曼特尔R统计量, 边缘颜色表示其显著性; 皮尔逊R值表示两个指标之间的相关系数, 其中*表示0.05水平差异显著, **表示0.01水平差异显著, ***表示0.001水平差异显著, n=12 图 4 土壤碳降解酶活性和碳库组分与微生物呼吸及其熵值的相关性分析 Fig. 4 Correlation analysis of soil microbial carbon metabolism characteristics, carbon pool components, and carbon metabolism enzyme activities |
为进一步探究土壤碳降解酶活性和碳库组分与微生物呼吸及其熵值的响应关系, 进行了偏最小二乘路径模型分析, 模型拟合优度为0.770, 且拟合结果显著[P < 0.05, 图 5(a)].结果表明, 人工刺槐林林龄的增加对土壤水解酶和惰性有机碳有着极显著的正相关效应[P < 0.001, 图 5(a)], 与氧化酶呈极显著的负相关关系[P < 0.001, 图 5(a)].在该模型中, 人工刺槐林恢复可以通过影响氧化酶活性(β=-0.876, P < 0.001)或惰性有机碳含量(β=1.12, P < 0.001)来间接影响土壤微生物代谢, 而且惰性有机碳是植被恢复影响土壤微生物代谢的关键环节[图 5(b)].此外, 刺槐林恢复还可以通过改变土壤物理性质(主要为粉粒和容重)来间接影响水解酶惰性和活性有机碳组分含量, 从而改变微生物代谢; 同时亦可直接改变土壤物理性质来间接影响微生物, 但刺槐林恢复并未对土壤物理性质造成显著影响(β=-0.377, P>0.05).总而言之, 人工刺槐林恢复并未直接对微生物造成影响, 而主要是通过改变碳降解酶活性或者土壤惰性有机碳库组分含量, 从而间接影响了微生物的代谢和生长.

|
(a)不同恢复年限、土壤物理性质、碳降解酶(水解酶和氧化酶)和土壤碳库(惰性和活性有机碳)作为影响微生物代谢驱动因子的偏小二乘路径模型; 红色和黑色箭头显示的是影响路径, 数字为路径系数, 表示关系的影响大小, 即β值, 红色箭头表示正相关, 黑色箭头表示负相关; 箭头宽度与路径系数的强度成正比, 其中*表示0.05水平差异显著, **表示0.01水平差异显著, ***表示0.001水平差异显著; (b)1.造林, 2.土壤物理性质, 3.水解酶, 4.氧化酶, 5.惰性有机碳, 6.活性有机碳; 各类驱动因素对微生物代谢的影响的总效应, 最终模型只显示共线性偏差(方差膨胀系数 & 10)的指标, n=12 图 5 土壤微生物代谢影响因素的偏最小二乘路径模型和各变量对微生物代谢的总效应 Fig. 5 Partial least squares-path model(PLS-PM) for influencing factors of soil microbial metabolism and the total effect of each variable on microbial metabolism |
土壤酶参与土壤碳库分解转化、影响微生物呼吸及生长代谢, 进而加速了物质转化过程与能量循环变化, 是评价植被恢复过程中土壤质量的重要生物活性指标[24].本研究发现农田退耕后, 土壤酶活性在人工刺槐林种植前期显著增加, 说明植被恢复后土壤生物学性质得到改善, 这可能与植物凋落物、根系分解以及植物根系分泌物等植物来源碳含量增加有关[25].但在刺槐林恢复后期, 氧化酶(PO+PER)和CBH酶活性均会有不同程度地降低[图 2(b)和图 3(b)], 这可能与刺槐林恢复过程中土壤物理组分如粉粒和砂粒没有发生明显的变化有关(表 1), 前人研究认为土壤物理组分表面能提供酶分解过程所需的反应位点, 土壤物理组分含量的增加可能增加酶活性[26~28].
在人工刺槐林植被恢复过程中, 土壤有机碳和惰性有机碳含量随着刺槐林龄的增加而显著增加, 同前人研究结果相似, 人工植被恢复能显著提高土壤有机碳及组分碳库[29].在薛萐等[30]的研究中, 分别对黄土丘陵区人工刺槐林总有机碳、活性有机碳与其生长年限进行耦合分析发现, 随林龄增加, 总有机碳与活性有机碳均线性增加.而在本研究中土壤活性有机碳含量在刺槐林各恢复年限并无显著差异, 其原因可能是黄土高原具有特殊的地理气候, 存在一定的干湿交替现象, 再加上样地具有特殊的小气候, 多次的干湿交替激发土壤有机碳的矿化, 提高了土壤活性碳库的降解速率和大小, 降低了惰性有机碳的降解[31].董扬红等[32]在对黄土高原不同植被类型下土壤活性有机碳组成特性研究得出, 不同植被类型恢复下土壤碳库以惰性有机碳为主的结论.同样在本研究中, 刺槐林各恢复年限土壤有机碳的平均增量有78.92%来自于惰性有机碳, 也证明了这一点(图 1).
3.2 人工林恢复模式下土壤碳降解酶活性和碳库组分与微生物代谢的关系土壤酶参与一系列的土壤生化过程, 与土壤有机碳转化密切相关[33].在植被恢复过程中, 大量的外源碳进入土壤会被土壤碳降解酶分解释放低分子量的糖, 为微生物生长代谢提供重要的碳源和能量源[34], 对土壤微生物呼吸具有一定的激发效应[35].偏最小二乘路径模型显示, 人工刺槐林恢复可以通过影响土壤氧化酶(PO+PER)活性来间接影响土壤微生物代谢, 而且微生物呼吸和氧化酶活性随刺槐林龄表现出相同的变化趋势, 这与Xiong等[36]的研究结果相似, 即: 土壤呼吸和转化酶活性变化具有一致性, 两者具有正相关关系.但是水解酶(BG+CBH)活性却和土壤微生物呼吸呈现出不同的显著结果(图 4), 其原因可能是一方面不同酶具有不同的功能[37], 在植被恢复不同阶段微生物所利用的倾向不同; 另一方面刺槐林在植被恢复过程中植被-土壤系统中可能会出现N或者P限制[38], 进而促进或抑制了不同碳降解酶活性的表达, 影响微生物活性.有研究得出β-1, 4-葡萄糖苷酶等水解酶对土壤有机碳的累积有着直接的正效应[39].在本研究中, 水解酶(BG+CBH)活性与qMB成正比(P < 0.05), 与qCO2成反比(图 4), 说明BG和CBH酶活性越高, 微生物呼吸消耗的碳比例越小, 生成微生物量碳的比例就越高, 这意味着BG和CBH酶对土壤的碳汇具有积极的正效应.
偏最小二乘路径模型表明, 人工刺槐林恢复主要是通过影响土壤惰性有机碳含量来间接影响土壤微生物代谢[图 5(a)].究其原因可能是, 从微生物生存策略角度来说, 微生物更易利用活性碳库(如刚进入土壤的新鲜有机质), 随着恢复的进行, 会有大量惰性有机碳库积累, 这也就刺激了微生物会分泌大量的水解酶分解活性物质来提供满足微生物自身代谢和生长所需的能源, 但随着人工刺槐林恢复年限的增加, 惰性有机碳的占比增加致使微生物可直接利用的碳源不断减少, 从而在刺槐林恢复后期在一定程度上抑制了微生物活性, 微生物呼吸降低(表 2).通常来说, 土壤有机碳大部分都保存在黏粒和粉粒中[40], 土壤矿物组分含量对土壤有机碳的分解速率有一定的影响, 质地越重的土壤分解速率越小, 有机碳含量就越高[41], 这表明土壤物理组分可以直接影响微生物代谢[图 5(a)].其可能的原因是土壤物理组分如粉粒和黏粒, 具有较大的表面积, 更易与土壤中的有机质结合形成土壤团聚体, 对有机质形成一种物理保护作用, 减缓了微生物对底物的利用[42], 进而影响了微生物代谢.有研究发现, 在植被恢复过程中, 较高的qMB和较低的qCO2说明微生物可以满足自己生态系统的平衡, 无需从外界获取更多的能量来维持生态系统的稳定性[30, 43].本研究中qMB和qCO2随人工刺槐林恢复过程表现出同薛萐等[30]的研究结果相似的趋势(表 2), 即: 在人工刺槐林恢复过程中其生态系统可能会趋于平衡稳定的状态.
总而言之, 在人工刺槐林植被恢复过程中, 大量的外源碳进入土壤给微生物提供足够的碳源, 微生物需要通过分泌更多水解酶(BG+CBH)来促进木质素的分解, 加快被木质素包裹的纤维素释放[21], 从而能够为参与纤维素分解的酶提供充足的底物, 促进活性有机碳的形成.而微生物分泌的氧化酶(PO+PER)主要是通过将土壤中的简单芳香族化合物氧化成醌类物质, 醌类物质能够与土壤中蛋白质和糖类等物质反应生成复杂的有机物质[44], 从而能够有效减少微生物对有机物质的利用, 利于活性碳库的转化保留, 进而增加惰性有机碳含量, 达到了固碳的效果.但是醌类物质并不能直接为微生物提供降解所需的碳源[25], 因此在恢复后期, 微生物并未增加氧化酶(PO+PER)的分泌, 而是通过增加水解酶(BG+CBH)的活性来分解足够多的活性碳物质来满足自身的生长和代谢.
4 结论(1) 在本研究中, 人工刺槐林恢复初期显著提升土壤微生物呼吸, 而且微生物熵(qMB)随刺槐林龄的增加显著增加, 微生物代谢熵(qCO2)则相反.
(2) 随着人工刺槐林植被恢复, 土壤有机碳和惰性有机碳含量显著积累, 活性有机碳无明显变化.
(3) 刺槐造林前期显著增加了土壤碳降解酶活性, 但在恢复后期氧化酶(PO+PER)和CBH酶活性呈现不同程度的降低趋势.
(4) 相关性分析表明土壤微生物碳降解酶活性与土壤碳库组分存在显著的相关关系; 偏最小二乘路径模型表明惰性有机碳组分是植被恢复影响土壤微生物代谢的关键环节.
(5) 总之, 人工刺槐林种植对土壤的碳汇具有积极的正效应, 而且随着刺槐林的恢复年限的增长其生态系统可能会趋向平衡稳定的状态.
| [1] | Jobbágy E G, Jackson R B. The vertical distribution of soil organic carbon and its relation to climate and vegetation[J]. Ecological Applications, 2000, 10(2): 423-436. DOI:10.1890/1051-0761(2000)010[0423:TVDOSO]2.0.CO;2 |
| [2] | Lal R. Soil carbon sequestration to mitigate climate change[J]. Geoderma, 2004, 123(1-2): 1-22. DOI:10.1016/j.geoderma.2004.01.032 |
| [3] | Stockmann U, Adams M A, Crawford J W, et al. The knowns, known unknowns and unknowns of sequestration of soil organic carbon[J]. Agriculture, Ecosystems & Environment, 2013, 164: 80-89. |
| [4] | Kirkby C A, Richardson A E, Wade L J, et al. Carbon-nutrient stoichiometry to increase soil carbon sequestration[J]. Soil Biology and Biochemistry, 2013, 60: 77-86. DOI:10.1016/j.soilbio.2013.01.011 |
| [5] | Knorr W, Prentice I C, House J I, et al. Long-term sensitivity of soil carbon turnover to warming[J]. Nature, 2005, 433(7023): 298-301. DOI:10.1038/nature03226 |
| [6] | Chang R Y, Fu B J, Liu G H, et al. Soil carbon sequestration potential for "Grain for Green" project in Loess Plateau, China[J]. Environmental Management, 2011, 48(6): 1158-1172. DOI:10.1007/s00267-011-9682-8 |
| [7] | Shao S, Zhao Y, Zhang W, et al. Linkage of microbial residue dynamics with soil organic carbon accumulation during subtropical forest succession[J]. Soil Biology and Biochemistry, 2017, 114: 114-120. DOI:10.1016/j.soilbio.2017.07.007 |
| [8] | Jones D L, Nguyen C, Finlay R D. Carbon flow in the rhizosphere: carbon trading at the soil-root interface[J]. Plant and Soil, 2009, 321(1-2): 5-33. DOI:10.1007/s11104-009-9925-0 |
| [9] | Kato T, Tang Y H, Gu S, et al. Carbon dioxide exchange between the atmosphere and an alpine meadow ecosystem on the Qinghai-Tibetan Plateau, China[J]. Agricultural and Forest Meteorology, 2004, 124(1-2): 121-134. DOI:10.1016/j.agrformet.2003.12.008 |
| [10] | Fuchs R, Schulp C J E, Hengeveld G M, et al. Assessing the influence of historic net and gross land changes on the carbon fluxes of Europe[J]. Global Change Biology, 2016, 22(7): 2526-2539. DOI:10.1111/gcb.13191 |
| [11] | Borie F, Rubio R, Rouanet J L, et al. Effects of tillage systems on soil characteristics, glomalin and mycorrhizal propagules in a Chilean Ultisol[J]. Soil and Tillage Research, 2006, 88(1-2): 253-261. DOI:10.1016/j.still.2005.06.004 |
| [12] | Geisseler D, Horwath W R, Scow K M. Soil moisture and plant residue addition interact in their effect on extracellular enzyme activity[J]. Pedobiologia, 2011, 54(2): 71-78. DOI:10.1016/j.pedobi.2010.10.001 |
| [13] | Wei M M, Xu G P, Guo H R, et al. Effect of vinasse application on soil in sugarcane fields[J]. Applied Mechanics and Materials, 2014, 700: 270-275. DOI:10.4028/www.scientific.net/AMM.700.270 |
| [14] | Kardol P, Cregger M A, Campany C E, et al. Soil ecosystem functioning under climate change: plant species and community effects[J]. Ecology, 2010, 91(3): 767-781. DOI:10.1890/09-0135.1 |
| [15] | Wang H Y, Wu J Q, Li G, et al. Changes in soil carbon fractions and enzyme activities under different vegetation types of the northern Loess Plateau[J]. Ecology and Evolution, 2020, 10(21): 12211-12223. DOI:10.1002/ece3.6852 |
| [16] | 鲍士旦. 土壤农化分析[M]. ((第三版)). 北京: 中国农业出版社, 2000. |
| [17] | Rovira P, Vallejo V R. Examination of thermal and acid hydrolysis procedures in characterization of soil organic matter[J]. Communications in Soil Science and Plant Analysis, 2000, 31(1-2): 81-100. DOI:10.1080/00103620009370422 |
| [18] | Stotzky G, Goos R D. Effect of high CO2 and low O2tensions on the soil microbiota[J]. Canadian Journal of Microbiology, 1965, 11(5): 853-868. DOI:10.1139/m65-115 |
| [19] | Yuan X B, Niu D C, Gherardi L A, et al. Linkages of stoichiometric imbalances to soil microbial respiration with increasing nitrogen addition: evidence from a long-term grassland experiment[J]. Soil Biology and Biochemistry, 2019, 138. DOI:10.1016/j.soilbio.2019.107580 |
| [20] | Saiya-Cork K R, Sinsabaugh R L, Zak D R. The effects of long term nitrogen deposition on extracellular enzyme activity in an Acer saccharum forest soil[J]. Soil Biology and Biochemistry, 2002, 34(9): 1309-1315. DOI:10.1016/S0038-0717(02)00074-3 |
| [21] | DeForest J L. The influence of time, storage temperature, and substrate age on potential soil enzyme activity in acidic forest soils using MUB-linked substrates and L-DOPA[J]. Soil Biology and Biochemistry, 2009, 41(6): 1180-1186. DOI:10.1016/j.soilbio.2009.02.029 |
| [22] | Maia S M F, Xavier F A S, Oliveira T S, et al. Organic carbon pools in a Luvisol under agroforestry and conventional farming systems in the semi-arid region of Ceará, Brazil[J]. Agroforestry Systems, 2007, 71(2): 127-138. DOI:10.1007/s10457-007-9063-8 |
| [23] | Liu W X, Xu W H, Hong J P, et al. Interannual variability of soil microbial biomass and respiration in responses to topography, annual burning and N addition in a semiarid temperate steppe[J]. Geoderma, 2010, 158(3-4): 259-267. DOI:10.1016/j.geoderma.2010.05.004 |
| [24] | Keiluweit M, Nico P S, Kleber M, et al. Are oxygen limitations under recognized regulators of organic carbon turnover in upland soils?[J]. Biogeochemistry, 2016, 127(2): 157-171. |
| [25] |
钟泽坤, 杨改河, 任成杰, 等. 黄土丘陵区撂荒农田土壤酶活性及酶化学计量变化特征[J]. 环境科学, 2021, 42(1): 411-421. Zhong Z K, Yang G H, Ren C J, et al. Effects of farmland abandonment on soil enzymatic activity and enzymatic stoichiometry in the Loess Hilly Region, China[J]. Environmental Science, 2021, 42(1): 411-421. |
| [26] | Allison S D. Soil minerals and humic acids alter enzyme stability: implications for ecosystem processes[J]. Biogeochemistry, 2006, 81(3): 361-373. DOI:10.1007/s10533-006-9046-2 |
| [27] | Zhong Z K, Han X H, Xu Y D, et al. Effects of land use change on organic carbon dynamics associated with soil aggregate fractions on the Loess Plateau, China[J]. Land Degradation & Development, 2019, 30(9): 1070-1082. |
| [28] | Naidja A, Huang P M, Bollag J M. Enzyme-clay interactions and their impact on transformations of natural and anthropogenic organic compounds in soil[J]. Journal of Environmental Quality, 2000, 29(3): 677-691. |
| [29] | Pongratz J, Reick C H, Raddatz T, et al. Effects of anthropogenic land cover change on the carbon cycle of the last millennium[J]. Global Biogeochemical Cycles, 2009, 23(4). DOI:10.1029/2009GB003488 |
| [30] |
薛萐, 刘国彬, 潘彦平, 等. 黄土丘陵区人工刺槐林土壤活性有机碳与碳库管理指数演变[J]. 中国农业科学, 2009, 42(4): 1458-1464. Xue S, Liu G B, Pan Y P, et al. Evolution of soil labile organic matter and carbon management index in the artificial robinia of Loess Hilly area[J]. Scientia Agricultura Sinica, 2009, 42(4): 1458-1464. DOI:10.3864/j.issn.0578-1752.2009.04.042 |
| [31] | Meisner A, Snoek B L, Nesme J, et al. Soil microbial legacies differ following drying-rewetting and freezing-thawing cycles[J]. The ISME Journal, 2021, 15(4): 1207-1221. DOI:10.1038/s41396-020-00844-3 |
| [32] |
董扬红, 曾全超, 安韶山, 等. 黄土高原不同林型植被对土壤活性有机碳及腐殖质的影响[J]. 水土保持学报, 2015, 29(1): 143-148. Dong Y H, Zeng Q C, An S S, et al. Effects of different forest types on soil active organic carbon and soil humus composition in the Loess Plateau[J]. Journal of Soil and Water Conservation, 2015, 29(1): 143-148. |
| [33] | Liyanage L R M C, Sulaiman M F, Ismail R, et al. Carbon mineralization dynamics of organic materials and their usage in the restoration of degraded tropical tea-growing soil[J]. Agronomy, 2021, 11(6): 1191. DOI:10.3390/agronomy11061191 |
| [34] | Davidson E A, Ishida F Y, Nepstad D C. Effects of an experimental drought on soil emissions of carbon dioxide, methane, nitrous oxide, and nitric oxide in a moist tropical forest[J]. Global Change Biology, 2004, 10(5): 718-730. DOI:10.1111/j.1365-2486.2004.00762.x |
| [35] | Sinsabaugh R L, Gallo M E, Lauber C, et al. Extracellular enzyme activities and soil organic matter dynamics for northern hardwood forests receiving simulated nitrogen deposition[J]. Biogeochemistry, 2005, 75(2): 201-215. DOI:10.1007/s10533-004-7112-1 |
| [36] | Xiong D, Gao Z Z, Fu B, et al. Effect of pyrimorph on soil enzymatic activities and respiration[J]. European Journal of Soil Biology, 2013, 56: 44-48. DOI:10.1016/j.ejsobi.2013.02.005 |
| [37] | Tischer A, Blagodatskaya E, Hamer U. Extracellular enzyme activities in a tropical mountain rainforest region of southern Ecuador affected by low soil P status and land-use change[J]. Applied Soil Ecology, 2014, 74: 1-11. DOI:10.1016/j.apsoil.2013.09.007 |
| [38] |
许淼平, 任成杰, 张伟, 等. 土壤微生物生物量碳氮磷与土壤酶化学计量对气候变化的响应机制[J]. 应用生态学报, 2018, 29(7): 2445-2454. Xu M P, Ren C J, Zhang W, et al. Responses mechanism of C: N: P stoichiometry of soil microbial biomass and soil enzymes to climate change[J]. Chinese Journal of Applied Ecology, 2018, 29(7): 2445-2454. |
| [39] | Xu H W, Qu Q, Chen Y H, et al. Responses of soil enzyme activity and soil organic carbon stability over time after cropland abandonment in different vegetation zones of the Loess Plateau of China[J]. CATENA, 2021, 196. DOI:10.1016/j.catena.2020.104812 |
| [40] | Freixo A A, de A Machado P L O, dos Santos H P, et al. Soil organic carbon and fractions of a rhodic ferralsol under the influence of tillage and crop rotation systems in southern Brazil[J]. Soil and Tillage Research, 2002, 64(3-4): 221-230. DOI:10.1016/S0167-1987(01)00262-8 |
| [41] |
姜勇, 庄秋丽, 梁文举. 农田生态系统土壤有机碳库及其影响因子[J]. 生态学杂志, 2007, 26(2): 278-285. Jiang Y, Zhuang Q L, Liang W J. Soil organic carbon pool and its affecting factors in farmland ecosystem[J]. Chinese Journal of Ecology, 2007, 26(2): 278-285. DOI:10.3321/j.issn:1000-4890.2007.02.023 |
| [42] | Lucas-Borja M E, Candel D, Jindo K, et al. Soil microbial community structure and activity in monospecific and mixed forest stands, under Mediterranean humid conditions[J]. Plant and Soil, 2012, 354(1-2): 359-370. DOI:10.1007/s11104-011-1072-8 |
| [43] | Zhou Z H, Wang C K, Jin Y. Stoichiometric responses of soil microflora to nutrient additions for two temperate forest soils[J]. Biology and Fertility of Soils, 2017, 53(4): 397-406. DOI:10.1007/s00374-017-1188-y |
| [44] | Morris S J, Bohm S, Haile-Mariam S, et al. Evaluation of carbon accrual in afforested agricultural soils[J]. Global Change Biology, 2007, 13(6): 1145-1156. DOI:10.1111/j.1365-2486.2007.01359.x |