 2021, Vol. 42
2021, Vol. 42



2. 中国科学院生态环境研究中心, 北京 100085
2. Research Center for Eco-Environmental Sciences, Chinese Academy of Sciences, Beijing 100085, China
磷是植物生长发育所必需的营养元素, 同时也是水体富营养化的限制因子, 其含量及赋存形态的变化对植物初级生产力和生态系统循环有着重要影响[1].滨海湿地是典型海陆交互作用地带, 对磷具有较强的过滤和截留作用, 是磷输送累积和再生的重要场所[2].目前, 国内外对于滨海湿地土壤磷开展了不少研究, 主要集中于土壤磷形态分布[3]、磷吸附-解吸及其影响因素[4]、不同磷营养水平对湿地植物生态影响[5]和不同活性磷组分转化动态与生物可利用性等方面[6].
生物入侵可能改变生态系统的功能和进程, 对生态系统的碳、氮、磷循环有潜在影响[7].其中, 互花米草是我国滨海湿地最主要的入侵植物之一, 对滨海湿地系统产生了重大影响[8].国内关于互花米草入侵对滨海湿地磷的相关研究虽已涉及闽江口湿地[9]、江苏盐城[10]、漳江口湿地[11]、杭州湾湿地[12]和九段沙湿地[13]等地, 但关于互花米草入侵对湿地土壤磷循环的影响尚无共识, 互花米草早期入侵增加了闽江河口鳝鱼滩湿地土壤TP含量[9], 而入侵漳江河口湿地后土壤TP含量降低[11], 也有研究者发现互花米草入侵增加了滩涂区土壤TP含量, 而入侵的红树林湿地土壤磷含量并没有明显差异[14].此外, 以上研究大多侧重于湿地土壤总磷和有效磷等方面, 对于无机磷的研究较少.
胶州湾湿地是山东半岛南部面积最大的近海与海岸湿地资源, 自20世纪末引种互花米草以来, 该群落出现了快速扩展的趋势.目前整个胶州湾滩涂不同程度分布着互花米草群落, 其中河口区域分布最为集中.胶州湾洋河口是胶州湾典型的受互花米草干扰的河口湿地, 其面积从2009年的1.7 hm2增加到2016年的78.4 hm2[15].当前关于互花米草入侵对胶州湾湿地土壤磷的研究主要集中于全磷、速效磷和有机磷等[16, 17], 而对于无机磷及其赋存形态的关注不足.本文中, 基于时空互易原则选取胶州湾湿地光滩区和互花米草区为研究靶区, 对比分析了两个采样区总磷和无机磷各组分含量及分布特征差异, 探讨了互花米草入侵对湿地土壤不同无机磷赋存形态变化特征的影响, 以期揭示在互花米草入侵影响下湿地土壤磷的转化机制.
1 材料与方法 1.1 研究区概况胶州湾(36°01′~36°15′N, 120°03′E~120°25′E)位于中国山东省青岛市, 延伸到山东半岛并向黄海开放, 属于半封闭型海湾(图 1), 是山东半岛南部面积最大的海湾.胶州湾沿岸潮汐为典型半日潮, 平均潮差2.71 m, 最大潮差6.87 m, 属于暖温带季风气候区, 年平均气温为12.2℃, 无霜期220 d左右[15].研究区位于胶州湾洋河入湾口潮滩湿地, 土壤质地主要是黏质土和砂质土.研究区内覆盖的优势植被为互花米草, 其茎秆坚韧直立、地下根茎发达, 常密布于地下0~40 cm深的土壤中, 有时可深达50 cm[18].

|
图 1 研究区及采样点示意 Fig. 1 Study area and sampling site |
依据典型性、代表性、一致性和可行性原则, 于2017年互花米草生长初期(5月)、生长旺盛期(7月)、生长成熟期(9月)和生长末期(11月), 选取胶州湾洋河口湿地互花米草区(SA区)和光滩区(MF区)为研究靶区, SA区互花米草覆盖率在90%以上, 光滩区无植被覆盖, 每个研究区内设置3个平行采样点(图 1).在对应样方内利用土钻自上而下分层分别采集0~10、10~20、20~30、30~40、40~50和50~60 cm的土样.取土后迅速将土壤装入密封袋中带回实验室, 去除杂质, 测定含水量, 自然风干后磨细过100目筛, 实验备用.4个月土壤样品为144个.
1.3 样品分析参照文献[19]的相关方法, 分析土壤样品中各环境因子: pH(电位法)、可溶性盐(电导分析法)和含水率(烘干法).有机质测定采用K2Cr2O7-H2SO4外加热法, TP测定采用磷钼蓝比色法, 无机磷(IP)组分测定参考顾益初等[20]提出的浸提流程, 将IP组分分为钙磷(Ca-P)、铝磷(Al-P)、铁磷(Fe-P)和闭蓄态磷(O-P)进行分析.所有测定均采用3份重复样品, 取平均值以减少误差.表 1为研究区土壤理化性质.
|
|
表 1 不同采样区土壤理化性质1) Table 1 Physical and chemical properties of the soils in the different sampling areas |
1.4 数据处理与分析
磷富集系数(enrichment coefficient, EC)可以用植物体内磷元素的含量与对应土壤的总磷含量的比值来表示, 计算公式如下:
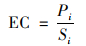
|
(1) |
式中, EC为植物对土壤磷富集系数, Pi为植物体内磷含量(mg·kg-1), Si为植物对应点土壤的总磷含量(mg·kg-1).
磷吸收量(PA)单位为g·m-2, 计算公式如下:

|
(2) |
式中, PA为植物养分总吸收量(g·m-2), PB为植物体内磷含量(g·kg-1), C为植物烘干后生物量(kg·m-2).
运用Origin 2017软件绘图, 采用SPSS 25.0(SPSS for windows, SPSS, Inc.)软件对数据进行单因素方差分析(ANOVA)、重复方差检验和采用最小二乘法(LSD)进行检验, P < 0.05为显著性, 数据在统计分析之前进行对数转换.最后采用IBM SPSS AMOS构建了结构方程模型(SEM).
2 结果与分析 2.1 互花米草入侵对土壤TP含量的影响胶州湾滨海湿地光滩和互花米草湿地土壤TP含量分别介于335.36~454.83mg·kg-1和374.31~549.86mg·kg-1之间, 其平均含量分别为386.19mg·kg-1和472.70mg·kg-1, 互花米草入侵后TP含量增幅为22.40%.采样期间, SA区土壤TP含量均显著高于MF区(P < 0.05), 尤其是9月, 互花米草区TP含量达477.18mg·kg-1, 相比于光滩区的363.13mg·kg-1, 增幅达到31.41%. SA区土壤TP含量在7月达到最大值, 平均含量为505.46mg·kg-1, 并在10~30 cm土层出现明显磷富集现象, 土壤总磷含量高达549.96mg·kg-1(图 2).垂直方向上, 每一土壤层, SA区TP含量均显著高于MF区(P < 0.05), 0~30 cm土壤层差异最为明显.两区0~10 cm土壤层TP含量均高于50~60 cm层(P < 0.05).此外, 经过重复方差检验, 月份变化、土壤深度对土壤TP含量的影响显著(P < 0.05), 但两者的交互作用不显著.

|
图 2 湿地土壤TP含量月份特征 Fig. 2 Monthly characteristics of TP content in wetland soil |
湿地土壤中的TP可分为无机磷和有机磷两大部分, 其中无机磷是土壤磷素的一个极其重要的组成成分, 是植物所需磷的主要来源[9].SA区和MF区土壤IP含量分别为212.22~283.42mg·kg-1和187.67~238.29mg·kg-1, 其平均含量分别为239.00mg·kg-1和212.68mg·kg-1.从图 3可得月份变化对土壤IP存在显著影响(P < 0.01), 而土层深度、两者之间的交互作用对土壤IP影响不显著.从月份变化上看, SA区土壤IP含量在9月达到最低, 平均含量220.40mg·kg-1, 显著低于5月和11月(P < 0.05).对比两区IP含量发现, 除7月外, SA区土壤IP含量显著高于MF区(P < 0.05).

|
不同的大写字母表示相同月份不同样地差异显著, 不同小写字母表示同一样地不同月份差异显著(P < 0.05), 误差棒表示均值的标准误差 图 3 采样区土壤IP含量分布特征 Fig. 3 Distribution characteristics of soil IP content in the sampling area |
土壤中磷素的形态决定磷素有效性, 恰当的磷素分级方法能较好地评价土壤有效磷库的大小和磷素的供应状况[21].研究区不同湿地类型土壤剖面各形态无机磷占IP总量的百分比见图 4, 其垂直分布随月份的变化见图 5.研究区湿地土壤钙磷(Ca-P)的均值为120.15mg·kg-1, 铁磷(Fe-P)的均值为88.65 mg·kg-1, 铝磷(Al-P)为11.91mg·kg-1, 闭蓄态磷(O-P)为5.25 mg·kg-1.胶州湾湿地土壤IP主要以Ca-P为主.Ca-P主要包括自生磷灰石磷及磷酸钙矿物, 如羟基磷灰石和过磷酸钙等[22].在磷形态分级中, 通常认为Ca-P属于惰性磷的一种, 难以被生物利用, 但互花米草入侵后, 10~30 cm层土壤Ca-P含量显著降低(P < 0.05), 7月尤为明显(图 5).铁磷(Fe-P)主要是指铁氧化物和氢氧化物胶体表面上吸附的磷, 该吸附作用易受环境因子的影响[16].互花米草入侵后土壤Fe-P含量显著增加(P < 0.05), 0~40 cm土壤层含量高于40~60 cm层, 并且7月份在10~40 cm土壤表现出明显富集现象(图 5).时间上, SA区Ca-P随着月份推移表现出先降低后增加再降低的趋势, 而Fe-P变化趋势与之相反(图 4).在本研究区, Al-P和O-P含量远低于Ca-P和Fe-P, 其含量变化主要体现在月份变化上, 在土壤剖面上没有明显差异(图 5), 5月SA区土壤Al-P含量显著高于MF区, 7月显著低于MF区; 9月SA区土壤O-P含量显著高于MF区(P < 0.05).

|
图中所示为平均含量袁误差棒表示平均值的标准差 图 4 采样区土壤IP组分含量变化及百分比 Fig. 4 Changes and percentages of soil IP components in the sampling area |

|
等高线数字对应无机磷各组分含量 图 5 采样区无机磷组分垂直分布 Fig. 5 Vertical distribution of inorganic phosphorus components in the sampling area |
湿地土壤中的磷生物地球化学过程涉及磷组分之间的转化, 土壤环境强烈影响这些转化[21].相关分析确定了互花米草入侵前后不同磷组分与土壤环境因子之间的关系[图 6(a)和6(b)].互花米草入侵前TP与Ca-P呈显著负相关(P < 0.05, R=-0.55), 与Fe-P呈显著正相关(P < 0.001, R=0.83).互花米草入侵降低了TP与Fe-P的相关性(P < 0.01, R=0.45), Fe-P与Ca-P呈显著负相关(P < 0.05, R=-0.36).比较图 6(a)和图 6(b), 发现互花米草入侵前后OM与TP和Fe-P始终保持高度正相关(P < 0.001).值得注意的是, 互花米草入侵后, pH值与Fe-P呈显著负相关(P < 0.01, R=-0.41), 土壤深度与TP和Fe-P呈显著负相关(P < 0.001).

|
(a)和(b)分别为互花米草入侵前(MF区)和入侵后(SA区)磷组分相关性分析, 数字表示相关系数, 紫色表示正相关, 蓝色表示负相关, 椭圆越小相关系数越大; (c)和(d)分别为互花米草入侵前(MF区)和入侵后(SA区)结构方程模型, 数字表示标准化路径系数, 带箭头的实线表示有重要影响的路径, 红色实线表示正向路径, 蓝色实线表示负向路径, 灰色虚线表示无意义的路径; 经Fisher检验, 模型拟合良好:(c)χ2=15.01, χ2/df=1.67, GFI=0.980, CFI=0.970; (d)χ2=50.12, χ2/df=2.64, GFI=0.910, CFI=0.933. *表示显著水平: *表示P < 0.05; **表示P < 0.01; ***表示P < 0.001 图 6 湿地土壤磷组分相关性分析和结构方程模型 Fig. 6 Correlation analysis and structural equation model of the phosphorus components in wetland soils |
利用结构方程模型进一步确定了互花米草入侵前后土壤磷含量的主要环境驱动因子[图 6(c)和6(d)].剔除不显著路径后, 模型具有显著性(卡方P>0.05), 拟合优度指数(GFISA=0.910, GFIMF=0.980)和卡方自由度比(χ2/df)可接受.SEM分析表明, OM、含水率、EC和pH对IP有显著影响, OM对Fe-P的影响最大(P < 0.01), 互花米草入侵前后的标准化系数分别为0.804和0.724, 说明互花米草入侵并没有改变OM与Fe-P的关系, 土壤Fe-P含量随有机质含量的增加而增加.互花米草入侵前, 土壤pH对Fe-P、Al-P和O-P含量均具有显著正向影响, 而Ca-P作为土壤的不溶性组分, 对土壤环境变化没有明显响应.与入侵前不同, 入侵后IP组分之间存在关联特征, Fe-P对Ca-P有显著负效应(P < 0.01), 标准化路径系数为-0.435, Al-P对Ca-P有显著正效应(P < 0.05).值得注意的是, Ca-P对土壤养分状况的变化没有反应.这个模型分别解释了71%(入侵前)和52%(入侵后)的Fe-P变异.
3 讨论 3.1 研究区土壤磷含量水平本研究区湿地土壤TP含量平均值是429.45mg·kg-1, 其值低于长江河口[23]、闽江河口[24]和珠江河口[25]等(表 2).P主要来自于土壤的天然母体, 本研究区湿地土壤主要发育于河海相沉积母质[26], 与河水冲淤的泥沙沉积形成的土壤母质相比肥力较低, 含磷量低.此外, 滨海湿地是典型的海陆交接地带, 周边径流的贡献将直接影响土壤P含量, 与长江和渤海相比, 胶州湾径流量低, 外源不足.本研究区土壤无机磷中Ca-P含量最高(图 4), 与河海相沉积物中Ca-P占主要优势一致[4].有研究表明, 有机质矿化也会增加土壤中Ca-P丰度[22], 然而, 在本研究中, SEM结果表示OM对Ca-P含量没有明显影响, 因此土壤中Ca-P的滞留也可能是钙与土壤中的矿物石英和方解石结合沉淀而来.此外, 钙磷含量可以较好地反映海陆相互作用的强弱.有研究发现, 河口或滨海湿地因受海洋作用较强, 土壤中钙磷含量往往占较大比重[27].胶州湾河口湿地土壤中除Ca-P外, Fe-P也占有很大比重, 占总无机磷含量的31.09%~48.74%, Fe-P与有机质含量呈显著正相关关系(P < 0.01), 说明有机质的累积和氧化降解会影响土壤中Fe-P的丰度.本研究区土壤Fe-P含量远低于印度Cochin河口[28], 胶州湾湿地属于温带, 温度和降水量均低于属于热带的Cochin河口湿地, 土壤风化作用不强烈, 土壤铁含量(35.325g·kg-1)低于Cochin湿地铁含量(74.531g·kg-1)[18], 进而造成了湿地Fe-P含量的差异.本研究区土壤Al-P含量为7.70~20.48mg·kg-1, 占IP的4.33%~6.75%, 远低于Fe-P含量, 这是由于磷酸盐在铝氧化物表面的结合力小于在铁氧化物表面的结合力[27].闭蓄态磷(O-P)也称为惰性磷或者不溶性磷, 主要来自表面水合铁氧化物包裹的结合态磷盐和自然岩石状态磷[29], 本研究区O-P含量较低, 平均含量只有5.25mg·kg-1, 占IP的1.43%~2.93%.
|
|
表 2 不同研究区磷素含量对比 Table 2 Comparison of P morphological contents in different research areas |
3.2 互花米草入侵对于湿地磷含量的影响 3.2.1 互花米草入侵对湿地总磷含量的影响
植物入侵对土壤磷具有重要影响, 它通过对磷的吸收、累积、归还以及根系作用等方式直接或间接改变土壤磷含量的高低及分布[24, 27, 29].本研究发现, 互花米草入侵在一定程度上改变了胶州湾湿地土壤磷的分布格局, 同时经重复方差检验得出月份变化和土壤深度对土壤TP均存在显著影响(P < 0.01).从图 2可知, 互花米草入侵后土壤TP在7月含量最高, 此时胶州湾正值雨季, 较好的水热条件可提高微生物活性, 促进地表凋落物的分解和土壤磷的释放, 尽管此时互花米草处于生长旺季, 对养分需求较高, 磷总吸收量PA达到全年峰值8.94 g·m-2(表 3), 但其与地表残体的P养分归还量相比较低.就垂直方向而言, 0~60 cm每一土壤层TP含量均表现为互花米草湿地>光滩区, 特别是0~30 cm土层.互花米草地下生物量多集中于0~40 cm土壤层, 其发达的根系形成于土壤剖面的根孔可显著增加土壤的水分迁移能力, 增加土壤TP的垂直迁移能力[17], 这也是7月SA区在10~30 cm出现P明显富集现象的原因, 此时互花米草根系生物量为全年最高值762.33 g·m-2.
|
|
表 3 互花米草不同器官生物量、磷含量、磷吸收量和磷富集系数 Table 3 Biomass, TP content, PA, and EC of different organs in S. alterniflora |
3.2.2 互花米草入侵对胶州湾河口湿地无机磷赋存形态的影响
已有研究表明植被群落是影响湿地土壤磷素形态及其在土壤剖面上分布差异的主要因素[21].在本研究中, 互花米草入侵使得湿地土壤IP含量增加(增幅为21.32%).作为胶州湾湿地土壤中IP的主要存在形式, Ca-P和Fe-P含量的变化最能体现磷素在互花米草入侵下的变化趋势.互花米草入侵后, 研究区表现出0~40 cm土壤Fe-P相对富集, Ca-P相对减少的现象, SEM模型结果显示, 互花米草入侵后Fe-P含量对Ca-P有显著负向影响(R=-0.435, P < 0.001).植被根系分泌的有机酸活化土壤的Al-P、Ca-P和Fe-P, 其中低分子有机酸可将有效性低的Ca-P转化为其他形态无机磷供自身利用[21, 30].此外, 根系分泌的其他无机物质, 如H+、OH-和CO2等, 改变环境的pH值, 从而影响湿地中磷的赋存形态[31].本研究表明, 互花米草入侵后SA区土壤pH低于MF区(表 1), 低pH促进不溶性Ca-P的溶解和羟基磷灰石(HAP)的释放, 一旦不稳定的PO43-从HAP释放到土壤中, 由于PO43-对氧化铁的高亲和力, 它可能与氧化铁结合形成Fe-P, 随后储存在土壤中[32].作为稳定态磷的O-P实质上是被铁氧化物Fe2O3胶膜包被的还原性磷酸铁铝盐, 其在还原条件下将被分解[29], 而0~40 cm是互花米草根系主要分布区域, 在根毛呼吸作用下土壤氧化还原电位(Eh)降低, 部分O-P转化成为Fe-P. SEM模型结果表示OM是影响Fe-P含量分布的主要因素, 这是由于互花米草的大量凋落物为土壤提供了丰富的有机质来源.其凋落物由于腐殖质化产生的氧化膜阻碍磷酸根与矿物的结合, 并且降解时产生的有机酸能溶解固定态磷[33], 促使Ca-P释放, Fe-P含量增加.在本研究中, 土壤含水率与Fe-P含量呈极显著正相关(P < 0.01), 表明淹水条件也是影响土壤磷素组分的关键因素.淹水前期, 土壤产生大量Fe2+, 但在持续淹水的过程中, Fe2+和Fe3+能够形成Fe2+-Fe3+无定形混合氢氧化物沉淀, 其表面积大于三价铁氧化物, 对磷酸根的吸附位点更多, Fe-P含量增加[21].
已有研究指出, Ca-P属于滨海环境中重要的磷素蓄积库, 而Fe-P具有潜在的生物有效性, 土壤磷素的迁移和转化与土壤Ca-P活性增加显著相关, 其中植被群落对无机磷形态有至关重要的作用, 如梁威等[34]的研究发现互花米草可将0~5 cm沉积物Ca-P向可溶性松散态磷转变.赖月婷等[3]的研究发现, 互花米草湿地Ca-P远低于短叶茳芏和扁穗莎草, 铁铝磷含量较高.此外, 其他湿地植物也可明显改善湿地沉积物的物理化学特性, 促进Ca-P向有效性高的形态转化.如包先明等[21]的研究指出苔草、芦苇和酸模可将土壤Ca-P活化, 造成Fe-P等形成土壤层表聚; 刘晶晶等[30]的研究通过对比扁秆藨草、芦苇和茭白根际和非根际土壤无机磷组分含量, 发现在3种植物根系的作用下, 某些有效性较低的Ca-P形态被大量转化成Fe-P等.综上, 相对于光滩, 互花米草入侵后胶州湾滨海湿地土壤中Ca-P含量降低, Fe-P含量增加, 提高了胶州湾湿地土壤磷的有效性.
4 结论(1) 互花米草入侵后湿地土壤TP和IP平均含量(472.70 mg·kg-1和239.00 mg·kg-1)明显高于入侵前光滩湿地(386.19 mg·kg-1和212.68 mg·kg-1).TP和IP含量的明显改变, 与入侵过程中土壤有机质含量的变化有关.
(2) 互花米草区和光滩区各形态无机磷含量整体均表现为: Ca-P>Fe-P>Al-P>O-P, Ca-P和Fe-P是土壤IP的主要成分, 分别占IP的45%~61%和31%~49%.互花米草入侵后, 10~30 cm层土壤Ca-P含量显著降低, Fe-P含量显著增加并在10~40 cm土壤表现出明显富集现象, 尤其是7月, 表现最为明显.
(3) SEM模型结果显示, 互花米草入侵后, Fe-P对Ca-P有显著负效应, 进而说明互花米草入侵不仅对湿地土壤磷含量有明显影响, 也改变不同磷形态间的转化, 提高土壤磷的有效性.
| [1] | Vitousek P M, Porder S, Houlton B Z, et al. Terrestrial phosphorus limitation: mechanisms, implications, and nitrogen-phosphorus interactions[J]. Ecological Applications, 2010, 20(1): 5-15. DOI:10.1890/08-0127.1 |
| [2] |
刘景双. 湿地生物地球化学研究[J]. 湿地科学, 2005, 3(4): 302-309. Liu J S. Wetland biogeochemistry research[J]. Wetland Science, 2005, 3(4): 302-309. DOI:10.3969/j.issn.1672-5948.2005.04.010 |
| [3] |
赖月婷, 吴如林, 龚开晟, 等. 闽江口湿地沉积物各形态磷分布特征及风险评价[J]. 水土保持学报, 2019, 33(1): 171-177, 185. Lai Y T, Wu R L, Gong K S, et al. Distribution characteristics and risk assessment of phosphorus speciation in the sediments of Min River Estuary wetland[J]. Journal of Soil and Water Conservation, 2019, 33(1): 171-177, 185. |
| [4] |
宋佳伟, 徐刚, 张扬, 等. 环渤海芦苇湿地磷的吸附容量及释放风险评估[J]. 环境科学, 2020, 41(2): 728-733. Song J W, Xu G, Zhang Y, et al. Phosphorus storage capacity and loss risk in coastal reed wetland surrounding Bohai Sea[J]. Environmental Science, 2020, 41(2): 728-733. |
| [5] | Zeng Q C, Liu Y, Xiao L, et al. How fencing affects the soil quality and plant biomass in the grassland of the loess plateau[J]. International Journal of Environmental Research and Public Health, 2017, 14(10). DOI:10.3390/ijerph14101117 |
| [6] | Coelho J P, Flindt M R, Jensen H S, et al. Phosphorus speciation and availability in intertidal sediments of a temperate estuary: relation to eutrophication and annual P-fluxes[J]. Estuarine, Coastal and Shelf Science, 2004, 61(4): 583-590. DOI:10.1016/j.ecss.2004.07.001 |
| [7] |
童晓雨, 孙志高, 曾阿莹, 等. 闽江河口互花米草海向入侵对湿地土壤无机硫赋存形态的影响[J]. 应用生态学报, 2019, 30(10): 3518-3526. Tong X Y, Sun Z G, Zeng A Y, et al. Effects of Spartina alterniflora invasion in a seaward direction on variations of inorganic sulfur forms in marsh soils of the Minjiang River Estuary, China[J]. Chinese Journal of Applied Ecology, 2019, 30(10): 3518-3526. |
| [8] |
解雪峰, 孙晓敏, 吴涛, 等. 互花米草入侵对滨海湿地生态系统的影响研究进展[J]. 应用生态学报, 2020, 31(6): 2119-2128. Xie X F, Sun X M, Wu T, et al. Impacts of Spartina alterniflora invasion on coastal wetland ecosystem: advances and prospects[J]. Chinese Journal of Applied Ecology, 2020, 31(6): 2119-2128. |
| [9] | Xie R R, Zhu Y C, Li J B, et al. Changes in sediment nutrients following Spartina alterniflora invasion in a subtropical estuarine wetland, China[J]. CATENA, 2019, 180: 16-23. DOI:10.1016/j.catena.2019.04.016 |
| [10] |
黄翊兰, 崔丽娟, 李春义, 等. 滨海滩涂湿地不同植被土壤磷的生物有效性及其影响因子分析[J]. 生态环境学报, 2019, 28(10): 1999-2005. Huang Y L, Cui L J, Li C Y, et al. Biologically-based availability and influencing factors of soil phosphorus under different vegetation in coastal beach wetlands[J]. Ecology and Environmental Sciences, 2019, 28(10): 1999-2005. |
| [11] | Feng J X, Zhou J, Wang L M, et al. Effects of short-term invasion of Spartina alterniflora and the subsequent restoration of native mangroves on the soil organic carbon, nitrogen and phosphorus stock[J]. Chemosphere, 2017, 184: 774-783. DOI:10.1016/j.chemosphere.2017.06.060 |
| [12] |
邵学新, 梁新强, 吴明, 等. 杭州湾潮滩湿地植物不同分解过程及其磷素动态[J]. 环境科学, 2014, 35(9): 3381-3388. Shao X X, Liang X Q, Wu M, et al. Decomposition and phosphorus dynamics of the litters in standing and litterbag of the Hangzhou Bay coastal wetland[J]. Environmental Science, 2014, 35(9): 3381-3388. |
| [13] |
刘长娥, 杨永兴, 杨杨. 九段沙中沙湿地植物磷元素的分布积累与动态[J]. 同济大学学报(自然科学版), 2008, 36(11): 1537-1541. Liu C E, Yang Y X, Yang Y. Distribution, accumulation and development of element P in middle shoal plants of Jiuduansha Wetlands[J]. Journal of Tongji University(Natural Science), 2008, 36(11): 1537-1541. DOI:10.3321/j.issn:0253-374X.2008.11.016 |
| [14] | Yu X Q, Yang J, Liu L M, et al. Effects of Spartina alterniflora invasion on biogenic elements in a subtropical coastal mangrove wetland[J]. Environmental Science and Pollution Research, 2015, 22(4): 3107-3115. DOI:10.1007/s11356-014-3568-2 |
| [15] | Wang X T, Jiang Z X, Li Y, et al. Inorganic carbon sequestration and its mechanism of coastal saline-alkali wetlands in Jiaozhou Bay, China[J]. Geoderma, 2019, 351: 221-234. DOI:10.1016/j.geoderma.2019.05.027 |
| [16] |
杜云鸿, 徐振, 谢文霞, 等. 胶州湾盐沼不同植物群落下土壤磷的分布特征[J]. 湿地科学, 2016, 14(3): 415-420. Du Y H, Xu Z, Xie W X, et al. Distribution characteristics of phosphorus under different vegetation communities in salt marshes of Jiaozhou Bay[J]. Wetland Science, 2016, 14(3): 415-420. |
| [17] |
苗萍, 谢文霞, 于德爽, 等. 胶州湾互花米草湿地氮、磷元素的垂直分布及季节变化[J]. 应用生态学报, 2017, 28(5): 1533-1540. Miao P, Xie W X, Yu D S, et al. Vertical distribution and seasonal variation of nitrogen, phosphorus elements in Spartina alterniflora wetland of Jiaozhou Bay, Shandong, China[J]. Chinese Journal of Applied Ecology, 2017, 28(5): 1533-1540. |
| [18] |
燕倩, 谢文霞, 沙梦乔, 等. 互花米草入侵对胶州湾河口湿地土壤总铁分布的影响[J]. 生态学报, 2020, 40(12): 3991-3999. Yan Q, Xie W X, Sha M Q, et al. Effects of Spartina alterniflora invasion on soil total iron distribution in estuary wetland of Jiaozhou Bay[J]. Acta Ecologica Sinica, 2020, 40(12): 3991-3999. |
| [19] | 鲍士旦. 土壤农化分析[M]. ((第三版)). 北京: 中国农业出版社, 2000. |
| [20] | 顾益初, 蒋柏蕃. 石灰性土壤无机磷分级的测定方法[J]. 土壤, 1990, 22(2): 101-102. |
| [21] |
包先明, 崔宏. 退耕还湖后不同植被群落湿地土壤剖面磷素形态分布特征[J]. 土壤, 2019, 51(1): 25-31. Bao X M, Cui H. Distribution of phosphorus fractions in wetland soil profiles under different vegetation communities after returning farmland to lake[J]. Soil, 2019, 51(1): 25-31. |
| [22] | Kang X M, Song J M, Yuan H M, et al. Phosphorus speciation and its bioavailability in sediments of the Jiaozhou Bay[J]. Estuarine, Coastal and Shelf Science, 2017, 188: 127-136. DOI:10.1016/j.ecss.2017.02.029 |
| [23] |
侯立军, 陆健健, 刘敏, 等. 长江口沙洲表层沉积物磷的赋存形态及生物有效性[J]. 环境科学学报, 2006, 26(3): 488-494. Hou L J, Lu J J, Liu M, et al. Species and bioavailability of phosphorus in surface sediments from the shoals in the Yangtze Estuary[J]. Acta Scientiae Circumstantiae, 2006, 26(3): 488-494. DOI:10.3321/j.issn:0253-2468.2006.03.022 |
| [24] | Li J B, Lai Y T, Xie R R, et al. Sediment phosphorus speciation and retention process affected by invasion time of Spartina alterniflora in a subtropical coastal wetland of China[J]. Environmental Science and Pollution Research, 2018, 25(35): 35365-35375. DOI:10.1007/s11356-018-3447-3 |
| [25] | Wang L L, Ye M, Li Q S, et al. Phosphorus speciation in wetland sediments of Zhujiang(Pearl) River Estuary, China[J]. Chinese Geographical Science, 2013, 23(5): 574-583. DOI:10.1007/s11769-013-0627-4 |
| [26] |
朱媛媛, 刘素美. 胶州湾沉积物中各形态P的分布及其生物地球化学研究[J]. 海洋环境科学, 2010, 29(6): 785-789. Zhu Y Y, Liu S M. Study on distribution and biogeochemistry of various phosphorus forms in sediments of Jiaozhou Bay[J]. Marine Environmental Science, 2010, 29(6): 785-789. DOI:10.3969/j.issn.1007-6336.2010.06.003 |
| [27] |
章文龙, 曾从盛, 仝川, 等. 闽江河口芦苇和短叶茳芏沼泽土壤磷分级特征比较[J]. 湿地科学, 2014, 12(6): 683-689. Zhang W L, Zeng C S, Tong C, et al. Comparison of characteristics of phosphorus speciation in soils of Phragmites australis and Cyperus malaccensis marshes in Min River Estuary[J]. Wetland Science, 2014, 12(6): 683-689. |
| [28] | Gireeshkumar T R, Deepulal P M, Chandramohanakumar N. Phosphorous speciation in surface sediments of the Cochin Estuary[J]. Environmental Monitoring and Assessment, 2013, 185(3): 2535-2552. DOI:10.1007/s10661-012-2729-3 |
| [29] | Tang W Z, Zhang H, Zhang W Q, et al. Biological invasions induced phosphorus release from sediments in freshwater ecosystems[J]. Colloids and Surfaces A: Physicochemical and Engineering Aspects, 2013, 436: 873-880. DOI:10.1016/j.colsurfa.2013.08.030 |
| [30] |
刘晶晶, 李敏, 曲博, 等. 湿地挺水植物根系土壤中的磷形态变化与分析[J]. 中国环境科学, 2013, 33(11): 2040-2046. Liu J J, Li M, Qu B, et al. The morphological changes and analysis of phosphorus in the root soil of emergent aquatic plant in the wetland[J]. China Environmental Science, 2013, 33(11): 2040-2046. |
| [31] | Joop-Jakobsen K, Mueller P, Meier R J, et al. Plant-sediment interactions in salt marshes-an optode imaging study of O2, pH, and CO2 gradients in the rhizosphere[J]. Frontiers in Plant Science, 2018, 9. DOI:10.3389/fpls.2018.00541 |
| [32] | Andersson K O, Tighe M K, Guppy C N, et al. XANES demonstrates the release of calcium phosphates from alkaline vertisols to moderately acidified solution[J]. Environmental Science & Technology, 2016, 50(8): 4229-4237. |
| [33] |
张晶. 北京野鸭湖湿地土壤中磷的形态分布和转化行为研究[D]. 北京: 北京林业大学, 2012. Zhang J. Distribution and transformation behaviors of phosphorus fractions in soil of Yeyahu Wetland in Beijing city[D]. Beijing: Beijing Forestry University, 2012. |
| [34] |
梁威, 邵学新, 吴明, 等. 杭州湾滨海湿地不同植被类型沉积物磷形态变化特征[J]. 生态学报, 2012, 32(16): 5025-5033. Liang W, Shao X X, Wu M, et al. Phosphorus fraction in the sediments from different vegetation type in Hangzhou Bay coastal wetlands[J]. Acta Ecologica Sinica, 2012, 32(16): 5025-5033. |