 2021, Vol. 42
2021, Vol. 42



2. 重庆大学环境与生态学院, 重庆 400044;
3. 重庆大学煤矿灾害动力学与控制国家重点实验室, 重庆 400044
2. College of Environment and Ecology, Chongqing University, Chongqing 400044, China;
3. State Key Laboratory of Coal Mine Disaster Dynamics and Control, Chongqing University, Chongqing 400044, China
过去几十年里, 我国土壤重金属污染状况一直是备受关注的热点问题[1~4].第一次全国土壤环境质量调查报告显示, 19.4%的农业土壤样品存在汞、砷、铬、铅和镉超标的现象[5].土壤重金属污染具有累积性、潜伏性、长期性和不可逆性的特点, 直接威胁土地质量和动植物的生长发育, 还会通过食物链危及人类健康.我国农田土壤重金属污染分布广泛, 空间异质性高, 分人为和自然两种成因[3, 6].在过去, 人们对施肥、污水灌溉、采矿冶金和交通运输等人类活动引起的重金属污染已有深入地研究[1~4], 认为人为源的重金属污染生态风险较高.仅有部分研究关注自然成因导致的土壤重金属污染[7~9], 发现自然源的重金属污染对生态系统的危害较轻.然而以往评估生态风险都是基于指数法, 而忽略了直接影响土壤健康的微生物指标.
微生物对重金属胁迫的响应远比动植物要敏感[10], 细微的环境变化都能引起微生物群落的改变, 因此, 土壤微生物被认为是表征土壤环境质量的指标之一, 可能会更直观地反映不同污染源的生态效应, 及时反映土壤污染状况[11].土壤微生物是土壤生态系统的重要组成部分, 可以驱动物质循环和促进作物生长, 对吸收和转移污染物以及缓解重金属毒性起到关键作用[11~13].近年来人们开始关注土壤微生物对重金属污染的响应.有研究表明, 变形菌和放线菌等细菌在不同污染程度的土壤表现出不同的耐受性[11].Li等[13]的研究也发现高含量重金属会降低微生物多样性以及拟杆菌和硝化螺旋菌的相对丰度.于皓等[14]的研究则发现铬污染显著影响土壤细菌群落结构与构建机制.总的来说, 土壤重金属污染显著影响了微生物群落结构和构建过程.
然而目前较少研究关注土壤古菌群落对重金属污染的响应特征及群落构建机制.古菌是一类数量丰富、种类多样且分布广泛的微生物, 对全球气候和生物地球化学循环发挥重要作用[15, 16].之前, 古菌被认为只存在于极端环境, 但高通量测序技术揭示了自然界大量未培养古菌, 特别是氨氧化古菌的发现彻底改变了人们对细菌主导氮循环的认识.Li等[12]的研究认为古菌类群对重金属污染具有更高的抗性, 可以通过改变重金属价态降低其毒性, 从而利于其他生物类群的生存.因此, 土壤古菌在生态系统中起到举足轻重的地位.Jiao等[17]的研究认为pH、温度和地理距离等都可以显著影响土壤古菌群落构建.Zheng等[18]的研究发现古菌群落构建受pH和采样深度等影响.群落构建直接决定群落组成和结构, 进而影响群落功能, 因此揭示群落构建机制对深入理解土壤古菌群落如何适应重金属污染有重要意义.
西南地区地形复杂, 长期自然风化过程中, 成土母质中的重金属会被黏土矿物吸附, 在土壤中大量富集, 形成高地质背景带[9], 伴随着人类活动, 土壤中重金属不断累积和再分配, 加剧了污染程度.因此本文选取3种不同污染源的重金属污染区域周边的农田土壤为研究对象, 包括高地质背景区、工矿区和涉重金属生产企业, 采用高通量测序技术研究不同地区土壤古菌群落组成与结构, 深入挖掘不同程度重金属胁迫下土壤古菌群落构建机制和驱动因素, 以期为农业土壤的防控治理和可持续发展提供参考依据.
1 材料与方法 1.1 样品采集本研究分别在重庆市潼南区某工业园周边(TN、TJ)、秀山县西北部典型汞矿周边(LD、YM)和秀山-酉阳典型地质高背景带(CJ、HF)附近农田采集5~10个表层土壤样品. 3个研究区域位置分布见图 1, 均属于亚热带湿润季风气候, 气温正常, 降水充沛.采用200 m×200 m网格布样法布设点位, 每一个样点采用棋盘式采样法采集0~20 cm的表层土壤样品, 使用木铲直接采取样品, 去除植物根、石块等杂物, 混匀后组成一个复合样品, 装入样品袋.土壤样品风干、研磨后送实验室测试.另取一份于无菌离心管中, 放入装有冰袋的保温箱内, 迅速带回实验室, -80℃冰箱冷冻干燥保存, 用于DNA提取.

|
图 1 采样区点位分布示意 Fig. 1 Map of the sampling sites |
土壤pH采用电位法测定, 水土比2.5∶1; 土壤有机质含量采用重铬酸钾法测定; 土壤重金属镉(Cd)、铅(Pb)、汞(Hg)、锌(Zn)、铜(Cu)、铬(Cr)、镍(Ni)和类金属砷(As)用电感耦合等离子体质谱仪(ICP-MS, Thermo Fisher, 美国)测定.
1.3 土壤DNA提取依托上海美吉生物医药科技有限公司进行土壤DNA提取与Illumina MiSeq高通量测序.利用Qiagen DNA Extraction Kits试剂盒(Qiagen GmbH, 德国)进行DNA提取, 根据说明书进行操作.选择引物524F(5′-TGYCAGCCGCCGCGGTAA-3′)和958R(5′-YCCGGCGTTGAVTCCAATT-3′)对16S rRNA的V4~V5区进行PCR扩增.
1.4 数据分析下机后得到的测序数据依托上海美吉生物医药科技有限公司的云平台进行处理和分析.简单来说, 利用FLASH(V1.2.11, http://ccb.jhu.edu/software/FLASH/)对每个样品的序列进行拼接, 根据QIIME(V1.9.1, http://qiime.org/)质控流程进行质量控制并去除嵌合体, 得到有效序列后利用Uparse 7.0按照97%的相似性聚类成操作分类单元(operational taxonomic units, OTUs), 并采用SILVA数据库(http://www.arb-silva.de/)对OTUs进行注释.运用Mothur计算群落α多样性, QIIME构建系统发育树、计算群落β多样性等.使用R软件(V 3.6.2)进行冗余分析(redundancy analysis, RDA)和Spearman相关性分析等.
根据Stegen[19]的研究方法, 使用R的“picante”包计算平均最近分类距离(mean nearest taxon distance, MNTD), 随后利用零模型将系统发育树分支末端的物种随机置换999次, 比较MNTD观测值与零模型生成的随机值之间的偏差, 得到最近分类指数(nearest taxon index, NTI).NTI表示群落内所有物种与其亲缘关系最近的种间距离的平均值, NTI>0表示组内样品聚类, NTI < 0表示组内样品存在过度分散.进一步计算平均最近种间距离(between-community mean nearest taxon distance, βMNTD)用来量化群落系统发育周转率, 最近β分类指数(between-community nearest taxon index, βNTI)量化βMNTD观测值和零模型生成的随机值之间偏差.βNTI>2表示随机过程, βNTI < 2表示确定过程, 计算公式如下.
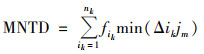
|
(1) |
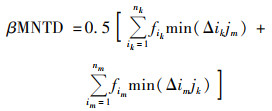
|
(2) |
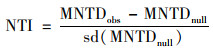
|
(3) |
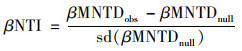
|
(4) |
式中, fik和fim是群落k和m中OTUi相对丰度; nk和nm是群落k和m中OTU数量; min(Δikjm)和min(Δimjk)分别是群落k中OTUi与m中OTUj和群落m中OTUi与k中OTUj之间的最小系统发育距离. MNTDobs和βMNTDobs是观察到的平均实际值, MNTDnull和βMNTDnull是零模型随机分配999次得到的平均预测值.
此外, 根据Stegen等[19]的研究方法计算RCbray(Bray-Curtis-based Raup-Crick)来量化基于Bray-Curtis相异性的Raup Crick指数观测值和零模型随机值之间差异.结合βNTI和RCbray结果, 确定性构建过程由同质性选择(βNTI < -2)和异质性选择(βNTI>2)组成, 随机性过程包括扩散限制(βNTI < 2, RCbray>0.95)和同质性扩散(βNTI < 2, RCbray < -0.95).如果βNTI < 2, RCbray < 0.95, 则群落构建过程由未知因素控制.
1.5 土壤重金属生态风险评价采用Hakanson[20]的潜在生态危害指数法评价研究区土壤重金属生态风险, 分级标准见表 1, 计算公式如下.
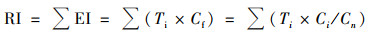
|
(5) |
|
|
表 1 Hakanson潜在生态风险分级标准 Table 1 Indices used to assess the potential ecological risks |
式中, RI为综合潜在生态风险指数; EI为单项潜在生态风险指数; Cf为某重金属的单因子污染指数; Ci为土壤中某重金属的实测值; Cn为某重金属的评价标准, 本文采用筛选值作为评价标准; Ti为重金属的毒性响应系数, 即Zn(1)<Cr(2)<Cu、Ni、Pb(5)<As(10)<Cd(30)<Hg(40).
2 结果与讨论 2.1 土壤理化特征6个地区土壤样品pH和有机质(SOM)含量如表 2所示.可以看出, 不同地区理化性质差异较大.pH的顺序为TJ>TN>LD>HF>CJ>YM, 除TN和TJ外, 其他4个地区均为酸性土壤.TN和TJ土壤样品SOM含量在10.0~47.5 g·kg-1之间, 平均值显著高于其他采样区(P < 0.01), 可能与较高的肥料投入有关.整体上, 小空间尺度下的土壤样品间理化性质差异较小, 灌溉方式、肥料用量和土壤质地等因素都会导致理化因子的异质性.然而矿区周边农田人类活动干扰最大, 样品间土壤理化性质差异显著(P < 0.01), YM周边矿业开采时间久, 土壤酸化严重.
|
|
表 2 土壤重金属含量及污染指数1) Table 2 Heavy metal content of soil and pollution index |
2.2 土壤重金属分布特征及污染评价
如表 2所示, 不同地区重金属含量差异明显, 在土壤中分布不均匀.整体上, 各重金属含量按平均值由大到小依次为:Zn>Cr>Ni>Pb>Cu>As>Hg>Cd.对比《土壤环境质量农用地土壤污染风险管控标准》(GB 15618-2018), 所有土壤样品Cu、Pb、Cr、Ni和Zn含量均没有超过风险筛选值, 而Cd和Hg超标率分别为56.6%和28.3%.仅有YM地区部分样品As含量超标. 6个地区土壤样品的Cd均有超标, CJ样品的Cd超标率最高(88.9%), TJ样品的Cd超标率最低(20.0%).只有YM和LD周边样品存在Hg超标.如表 2所示, 重金属综合潜在生态危害(RI)从大到小依次为YM>LD>CJ>HF>TJ>TN.结果表明矿区周边土壤污染RI值最高, 存在轻微到强生态风险.高地质背景区和涉重金属企业周边农田土壤的RI值都低于150, 存在轻微生态风险, 但CJ有55.6%的样品Cd的潜在生态风险指数大于40, 存在轻微至强生态风险.
以上结果表明, 本研究土壤重金属空间异质性高, 与成土母质、农业管理方式及周边人类活动都有关.污染最严重且分布广泛的重金属元素是Cd, 其次是Hg.有研究表明, Cd和Hg是农田土壤最普遍的污染物[2~4].Cd来源广泛, 自然成土过程中的迁移与再分配、污水灌溉和化肥农药的投入, 都能造成土壤Cd污染.酸性土壤中Cd活性高且迁移性强, 易在植株中累积, 对人类健康产生威胁[7].因此, Cd在YM样品中存在强生态危害, TN样品中生态危害低.Hg污染主要来源于矿业开采活动, 大气沉降、地面径流和地下径流等使采矿冶炼释放出来的Hg在周边农田土壤严重富集[3, 4]. Hg会被生物转化为毒性强的有机Hg, 抑制生物细胞代谢, 降低机体功能[3, 4]. Hakanson[20]的评价尽管考虑了生态风险, 但是基于重金属总量, 未考虑重金属元素多种赋存形态和价态产生的生物毒性, 因此研究土壤微生物对重金属胁迫的响应十分必要.
2.3 土壤古菌多样性和群落组成测序数据处理后共得到508个OTUs, 分类获得12门25纲37目51科69属146种.不同地区样品OTU数目平均值从大到小依次为TN(82)>TJ(78)>YM(30)>LD(26)>CJ(25)>HF(22), TN和TJ样品土壤古菌多样性显著高于其他地区, 而高地质背景区样品多样性最低.门水平上(图 2), 泉古菌门(Crenarchaeota)在6个地区土壤古菌群落都占据明显的优势, 其在TN和TJ样品中占比61.2%和64.6%, 在其他4个地区占比90%以上.Halobacterota在TN样品中平均相对丰度最高, 为22.8%.此外, 还检测到了广古菌门(Euryarchaeota)、Thermoplasmatota及其它未分类的古菌门类等.

|
图 2 不同地区门水平土壤古菌群落结构 Fig. 2 Archaeal community composition of different typical areas at the phylum level |
将所有样品根据RI值分为3个污染水平, OTU数目平均值从大到小依次为轻微(44)>强(30)>中等(26), 表明重金属轻微生态风险条件下土壤古菌呈现出最高的多样性.目水平上(图 3), 随生态风险等级的增加, 亚硝化球菌(Nitrososphaerales)平均相对丰度从72.0%降低到53.4%, 而亚硝化杆菌(Nitrosotaleales)平均相对丰度从5.9%增加到33.5%.此外还有少量的Methanocellales等产甲烷菌分布在各个样品中.Nitrososphaerales相对丰度随pH的升高先升高后降低, Nitrosotaleales则相反.本研究结果表明古菌分布可能与pH有关.

|
潜在生态风险等级:A.轻微, B.中等, C.强; pH:D < 5.5, E 5.5~6.5, F 6.5~7.5, G >7.5 图 3 目水平土壤古菌群落结构 Fig. 3 Archaeal community composition at the order level |
Crenarchaeota在本研究的农田土壤中占主导地位.Crenarchaeota广泛存在于海洋、热泉和土壤等环境, 可以将氨氧化为亚硝酸盐进行化能自养生长, 是自然界数量最多、分布最广泛的氨氧化古菌类群[15, 21, 22].氨氧化过程是硝化作用的第一步, 是氮肥利用的关键, 可以控制温室气体N2O排放速率, 进而影响氮循环[21].此外, 这类古菌还被报道具有无机碳固定的功能[23~25].Nitrososphaerales和Nitrosotaleales为Crenarchaeota的优势类群, 除了氨氧化, 还具有生物膜形成、细胞表面修饰、糖类及芳香族化合物代谢等能力, 以此应对环境胁迫[23, 24].Halobacterota是从广古菌门分出的新古菌门, 分类到其门下的Methanocellales等产甲烷古菌, 可以进行厌氧发酵生成甲烷, 参与全球碳循环.低污染的TN和TJ样本中有机质丰富, pH偏碱性, 需要氨氧化古菌和产甲烷古菌协同进行物质循环, 因此群落多样性最高.中等污染的CJ和HF样本中, pH接近中性, 低浓度的重金属可以提供细胞生长和代谢所需的微量元素, 适宜微生物生长, 古菌多样性最低可能是因为细菌的大量繁殖.YM样本中, 极端酸性和高浓度的重金属胁迫抑制细胞生长, 使得耐受菌生存下来获得生态位, 导致具有相同功能的优势菌Nitrososphaerales占比减少而Nitrosotaleales占比增加.
总而言之, 本研究典型重金属污染农田土壤古菌群落, 由氨氧化古菌和产甲烷古菌主导, 其相对贡献取决于土壤环境.这一发现与Jiao等[17]和Liu等[26]的研究结果相似, 古菌在农田土壤主要促进硝化作用, 从而驱动农田生态系统碳氮循环.
2.4 土壤古菌群落与环境因子的关系RDA结果显示(图 4), CJ、HF、YM和LD样品由于地理位置较近而聚集, 群落结构较为相似, TN和TJ样品较分散.第1轴(RDA1)和第2轴(RDA2)共贡献67%, 表明环境因子可以很好地解释群落结构差异.Envfit检验表明, SOM(R2=0.63, P=0.001)、pH(R2=0.37, P=0.001)、Pb(R2=0.36, P=0.001)和As(R2=0.33, P=0.001)对土壤古菌群落结构具有显著影响.总体上, SOM贡献最大, 与Crenarchaeota呈负相关, 与Halobacterota呈正相关, 对TN和TJ样品群落有显著的驱动作用.Pb和As等重金属箭头方向与CJ、HF、YM和LD样品排列方向一致, 且对Crenarchaeota有正面影响.图 5显示pH与Nitrosotaleales、Group_1.1c呈显著负相关(P < 0.01), SOM与Nitrososphaerales呈显著负相关(P < 0.01), 而二者与Methanobacteriales等呈显著正相关(P < 0.01), Hg、As和Pb对Methanobacteriales等产甲烷菌呈显著负相关(P < 0.01), Pb与Nitrososphaerales呈显著正相关(P < 0.01).

|
图 4 冗余分析 Fig. 4 Redundancy analysis |

|
***表示P < 0.001,**表示P < 0.01,*表示P < 0.05 图 5 古菌群落与土壤理化性质的相关性热力图 Fig. 5 Spearman's correlation of archaeal communities and environmental factors |
整体上, 土壤SOM是本研究农田土壤古菌群落结构分异的主要驱动因素, 然而有研究表明pH主导古菌群落结构[26~28].据报道, SOM对产甲烷古菌类群影响较大, 而pH对氨氧化古菌影响显著[17, 28].TN和TJ样品较高的pH导致氨氧化古菌活性下降, 而丰富的有机质使产甲烷的Halobacterota类群占据生态位[28], 导致该地区截然不同的群落结构空间格局.此外, Crenarchaeota类群对重金属耐受性更强, Nitrososphaerales表现出较高的耐Pb和As性, 而Nitrosotaleales表现出较高的耐Hg性.尽管Pb和As未构成污染, 但其含量与大多优势菌有显著的负相关关系, 可能是Pb2+和As3+的累积破坏细胞结构从而影响土壤古菌群落结构.相反, Cd含量对古菌群落作用较小, 可能是地质高背景区和中性土壤样品中, Cd活性较低.先前研究表明, Crenarchaeota类群丰度与Cd、Pb和As等重金属含量呈正相关, 与本研究的结果相似[12, 29].因此, 重金属等环境因子的空间异质性使得功能相似的古菌类群产生不同的响应, 从而导致群落结构的差异.
本研究中pH没有形成明显的环境梯度, 这可能是pH不是主导因素的原因, 但pH通过调节有毒金属的浸出和土壤养分的转化[27], 间接影响古菌群落组成和构建.Cao等[28]的研究发现除pH外, 土壤养分含量与古菌丰度呈正相关, 此外古菌群落分布还受盐度、温度、海拔和纬度等因素影响[17, 26, 27].由于古菌群落之间、古菌与细菌之间和古菌与环境之间均存在复杂的互作网络, 本研究采样范围有限, 今后还需配合更大时空尺度的土壤污染调查, 才能更好地揭示污染农田土壤古菌群落组成和驱动因素.
2.5 古菌群落构建机制分析如图 6所示, 所有样本NTI指数平均值显著大于0, 表明各样本古菌群落在系统发育上聚类, 聚类强度从大到小依次为TJ>TN>LD>CJ>HF>YM, TJ和TN样本古菌群落有明显高于预期的系统发育聚类, 表现出较强的确定性过程.根据不同风险等级来看, NTI值在轻微风险样品中最高, 中等风险样品中最低, 与OTU数量变化一致, 表明重金属轻微污染刺激古菌群落表现出较高的系统发育聚类.单因素方差分析结果表明pH>7.5的样品中, NTI平均值显著高于其他样品(P < 0.05), 表明碱性土壤中古菌群落种间亲缘关系更紧密, 而在中酸性土壤, NTI随着pH的降低先降低后升高, 这与图 3群落组成的差异类似, 表明重金属污染和pH可能是影响群落构建的重要因素.此外, 本研究结合βNTI和RCbray指数分析不同地区土壤古菌群落构建过程, 结果如图 7所示.CJ、HF、YM和LD周边农田土壤古菌群落构建主要是异质过程(异质选择+扩散限制)占主导, 占比80.0%~93.9%, 其中异质选择(确定过程)占53.3%~71.2%, 扩散限制(随机过程)占20.5%~26.7%, 但均未发现同质选择过程.TJ和TN农田土壤古菌群落构建过程中, 确定过程占比72.8%和70.0%, 其中异质选择分别占54.3%和50.0%, 同质选择分别占18.5%和20.0%, 而随机过程由扩散限制主导, 分别占比14.8%和20.0%, 相比其他地区, 同质选择和确定过程的相对重要性增加.整体上, 本研究确定过程的相对贡献高于随机过程, 二者共同驱动农田土壤古菌群落构建.

|
风险等级: A轻微, B中等, C强; pH: D < 5. 5, E 5.5~6.5, F6.5~7.5, G >7.5 图 6 NTI值 Fig. 6 Nearest taxon index |

|
图 7 不同地区古菌群落构建过程占比 Fig. 7 Archaeal community assembly of different areas |
越来越多证据表明, 随机过程和确定过程会同时发生在群落构建中, 二者相对作用大小随物种、研究区域和作物类型的不同而有所差异[30, 31].一般来说, 小时空尺度下环境选择起主要作用, 大时空尺度下, 扩散限制起主要作用[30].而Jiao等[17]的研究中, 农田土壤古菌群落构建由环境过滤选择, 古菌表现出强烈的生境偏好, 如极端pH和温度等.本研究采样区空间尺度较小, 频繁的人类活动和持续的外源重金属输入等使环境剧烈变化, 土壤环境空间异质性高, 古菌群落构建过程主要受异质选择影响, 地理距离引起的扩散限制作用较小.RDA结果表明(图 4), pH、SOM、Pb和As等都是本研究中用于筛选物种的环境过滤条件, 只有具有特定功能性状的古菌, 才能被环境过滤选入本地群落.图 5显示, Nitrososphaerales这类自养菌在低营养的土壤中竞争力更强, 因此在不同地区群落中都占优势, 而在高营养环境中, Halobacterota菌被丰富的SOM筛选入TN和TJ群落进行有机物代谢, 因此SOM的空间异质性导致了不同营养型古菌群落构建的差异; 而pH对古菌的选择作用更为直接, 低pH将嗜酸的Nitrosotaleales和Group_1.1c类群筛入YM群落中生存下来获得生态位, Halobacterota类群不能适应则失去竞争优势; Nitrososphaerales与Pb和As之间以及Nitrosotaleales与Hg之间呈显著的正相关, 体现了重金属对古菌的正向选择作用, 低重金属污染下这些古菌可能会改变Pb、As和Hg的价态降低其毒性从而成功定殖[12].因此, 环境选择最终驱动本研究古菌群落的构建, 使得抗性高、营养需求低和竞争力强的Crenarchaeota成为优势类群, 而环境的异质性增加了异质选择对群落构建的贡献, 使古菌群落空间分布格局因地而异.
于皓等[14]的研究认为铬污染土壤中细菌群落构建是同质选择占主导[14], 随着铬含量的升高, 环境选择对群落的确定性逐渐增加.本研究TN和TJ区域也存在一定比例的同质选择作用, 可能因为该地区受工业活动影响较小, 污染较轻, 而长期固定的耕作和施肥模式使得局域内土壤环境变化不大, 环境选择的压力也是均一的, 因此同质选择的相对重要性增加, 群落周转率降低, 缩小了样点间群落结构的差异.
有研究表明, 随机过程在资源丰富和环境压力较小的生境下起主导作用[32, 33], 由于微生物易于迁移, 通过扩散迁移到新生境进行定殖, 从而导致空间上均匀的群落结构.本研究区域内土壤古菌群落空间分布尽管差异很大, 但存在一定比例的扩散限制作用.各采样点与工业区之间有着不同的距离, 形成一定的污染梯度, 污染达到某个程度, 古菌向新生境的扩散受到限制, 使古菌在特定的生态位定殖下来, 从而产生更多的物种多样性以及生态系统功能, 加大了群落空间异质性[32].Jiao等[34]的研究发现, 随机过程与确定过程之间的平衡受到环境因子的调节, 环境异质性会改变不同生态过程对古菌群落构建的相对贡献.总的来说, 本研究环境的异质性驱动了不同生态位偏好的古菌群落构建, 异质选择强化了古菌群落在特定生境下的功能, 从而增强了古菌抵御环境胁迫和参与碳氮循环的能力, 有助于维持农田生态系统稳定性和可持续性.
本研究揭示了古菌在不同类型重金属污染地区农田土壤中的群落组成与响应机制, 证明研究古菌群落构建对阐明农田生态系统物质循环和应对胁迫的机制有重要意义.然而确定性过程除了环境选择, 还包括种间关系等生物因素, 本文未考虑土壤微生物群落物种间的共存互作网络, 将在下一步工作中深入研究.
3 结论(1) 不同类型重金属污染地区土壤理化差异明显, 重金属空间分布异质性高, 镉是最主要的污染物, 除工矿区部分样点土壤存在强生态风险, 其他样品的潜在生态风险为轻微~中等.
(2) 不同地区古菌群落结构差异较大, 优势门为泉古菌门, 优势目是亚硝化球菌和亚硝化杆菌, 其相对丰度取决于土壤环境.
(3) 土壤有机质、pH通过影响不同优势菌相对丰度驱动本研究农田土壤古菌群落结构分异, 铅和砷是对群落结构差异贡献较高的重金属元素.
(4) 古菌群落构建过程主要受确定性过程影响, 异质选择主导本研究古菌群落构建.
| [1] | Zhao F J, Ma Y B, Zhu Y G, et al. Soil contamination in China: current status and mitigation strategies[J]. Environmental Science & Technology, 2015, 49(2): 750-759. |
| [2] | Duan Q N, Lee J, Liu Y S, et al. Distribution of heavy metal pollution in surface soil samples in China: a graphical review[J]. Bulletin of Environmental Contamination and Toxicology, 2016, 97(3): 303-309. DOI:10.1007/s00128-016-1857-9 |
| [3] | Hu B F, Shao S, Ni H, et al. Current status, spatial features, health risks, and potential driving factors of soil heavy metal pollution in China at province level[J]. Environmental Pollution, 2020, 266. DOI:10.1016/j.envpol.2020.114961 |
| [4] |
陈文轩, 李茜, 王珍, 等. 中国农田土壤重金属空间分布特征及污染评价[J]. 环境科学, 2019, 41(6): 2822-2833. Chen W X, Li Q, Wang Z, et al. Spatial distribution characteristics and pollution evaluation of heavy metals in arable land soil of China[J]. Environmental Science, 2019, 41(6): 2822-2833. |
| [5] | 环境保护部, 国土资源部. 全国土壤污染状况调查公报[R]. 北京: 环境保护部, 国土资源部, 2014. |
| [6] | Zhang Q C, Wang C C. Natural and human factors affect the distribution of soil heavy metal pollution: a review[J]. Water, Air, & Soil Pollution, 2020, 231(7). DOI:10.1007/s11270-020-04728-2 |
| [7] |
刘意章, 肖唐付, 熊燕, 等. 西南高镉地质背景区农田土壤与农作物的重金属富集特征[J]. 环境科学, 2019, 40(6): 2877-2884. Liu Y Z, Xiao T F, Xiong Y, et al. Accumulation of heavy metals in agricultural soils and crops from an area with a high geochemical background of cadmium, Southwestern China[J]. Environmental Science, 2019, 40(6): 2877-2884. |
| [8] |
唐瑞玲, 王惠艳, 吕许朋, 等. 西南重金属高背景区农田系统土壤重金属生态风险评价[J]. 现代地质, 2020, 34(5): 917-927. Tang R L, Wang H Y, Lü X P, et al. Ecological risk assessment of heavy metals in farmland system from an area with high background of heavy metals, Southwestern China[J]. Geoscience, 2020, 34(5): 917-927. |
| [9] |
孙斌, 魏志敏, 张力浩, 等. 地质高背景土壤重金属赋存特征及微生物群落结构差异[J]. 土壤学报, 2020. Sun B, Wei Z M, Zhang L H, et al. Distribution of heavy metals and microbial community structure in soils high in geological background value[J]. Acta Pedologica Sinica, 2020. DOI:10.11766/trxb202001030003 |
| [10] | Gómez-Sagasti M T, Alkorta I, Becerril J M, et al. Microbial monitoring of the recovery of soil quality during heavy metal phytoremediation[J]. Water, Air, & Soil Pollution, 2012, 223(6): 3249-3262. |
| [11] | Lin Y B, Ye Y M, Hu Y M, et al. The variation in microbial community structure under different heavy metal contamination levels in paddy soils[J]. Ecotoxicology and Environmental Safety, 2019, 180: 557-564. DOI:10.1016/j.ecoenv.2019.05.057 |
| [12] | Li X Q, Meng D L, Li J, et al. Response of soil microbial communities and microbial interactions to long-term heavy metal contamination[J]. Environmental Pollution, 2017, 231: 908-917. DOI:10.1016/j.envpol.2017.08.057 |
| [13] | Li C C, Quan Q, Gan Y D, et al. Effects of heavy metals on microbial communities in sediments and establishment of bioindicators based on microbial taxa and function for environmental monitoring and management[J]. Science of the Total Environment, 2020, 749. DOI:10.1016/j.scitotenv.2020.141555 |
| [14] |
于皓, 安益君, 金德才, 等. 铬污染对土壤细菌群落结构及其构建机制的影响[J]. 环境科学, 2021, 42(3): 1197-1204. Yu H, An Y J, Jin D C, et al. Effects of chromium pollution on soil bacterial community structure and assembly processes[J]. Environmental Science, 2021, 42(3): 1197-1204. DOI:10.13227/j.hjkx.202010209 |
| [15] | Baker B J, De Anda V, Seitz K W, et al. Diversity, ecology and evolution of Archaea[J]. Nature Microbiology, 2020, 5(7): 889-900. |
| [16] |
邓霏. 古菌研究进展[J]. 安徽农业科学, 2018, 46(28): 11-14, 47. Deng F. Research progress of archaea[J]. Journal of Anhui Agricultural Sciences, 2018, 46(28): 11-14, 47. DOI:10.3969/j.issn.0517-6611.2018.28.004 |
| [17] | Jiao S, Xu Y Q, Zhang J, et al. Environmental filtering drives distinct continental atlases of soil archaea between dryland and wetland agricultural ecosystems[J]. Microbiome, 2019, 7(1). DOI:10.1186/s40168-019-0630-9 |
| [18] | Zheng Y M, Cao P, Fu B J, et al. Ecological drivers of biogeographic patterns of soil archaeal community[J]. PLoS One, 2013, 8(5). DOI:10.1371/journal.pone.0063375 |
| [19] | Stegen J C, Lin X J, Fredrickson J K, et al. Quantifying community assembly processes and identifying features that impose them[J]. The ISME Journal, 2013, 7(11): 2069-2079. DOI:10.1038/ismej.2013.93 |
| [20] | Hakanson L. An ecological risk index for aquatic pollution control.a sedimentological approach[J]. Water Research, 1980, 14(8): 975-1001. DOI:10.1016/0043-1354(80)90143-8 |
| [21] | Brochier-Armanet C, Boussau B, Gribaldo S, et al. Mesophilic Crenarchaeota: proposal for a third archaeal phylum, the Thaumarchaeota[J]. Nature Reviews Microbiology, 2008, 6(3): 245-252. DOI:10.1038/nrmicro1852 |
| [22] | Dang H Y, Zhang X X, Sun J, et al. Diversity and spatial distribution of sediment ammonia-oxidizing Crenarchaeota in response to estuarine and environmental gradients in the Changjiang Estuary and East China Sea[J]. Microbiology, 2008, 154(7): 2084-2095. DOI:10.1099/mic.0.2007/013581-0 |
| [23] | Kerou M, Offre P, Valledor L, et al. Proteomics and comparative genomics of Nitrososphaera viennensis reveal the core genome and adaptations of archaeal ammonia oxidizers[J]. Proceedings of the National Academy of Sciences of the United States of America, 2016, 113(49): E7937-E7946. DOI:10.1073/pnas.1601212113 |
| [24] | Alves R J E, Minh B Q, Urich T, et al. Unifying the global phylogeny and environmental distribution of ammonia-oxidising archaea based on amoA genes[J]. Nature Communications, 2018, 9(1). DOI:10.1038/s41467-018-03861-1 |
| [25] | Zhang L M, Offre P R, He J Z, et al. Autotrophic ammonia oxidation by soil thaumarchaea[J]. Proceedings of the National Academy of Sciences of the United States of America, 2010, 107(40): 17240-17245. DOI:10.1073/pnas.1004947107 |
| [26] | Liu J J, Yu Z H, Yao Q, et al. Biogeographic distribution patterns of the archaeal communities across the black soil zone of northeast China[J]. Frontiers in Microbiology, 2019, 10. DOI:10.3389/fmicb.2019.00023 |
| [27] | Tripathi B M, Kim M, Tateno R, et al. Soil pH and biome are both key determinants of soil archaeal community structure[J]. Soil Biology and Biochemistry, 2015, 88: 1-8. DOI:10.1016/j.soilbio.2015.05.004 |
| [28] | Cao P, Zhang L M, Shen J P, et al. Distribution and diversity of archaeal communities in selected Chinese soils[J]. FEMS Microbiology Ecology, 2012, 80(1): 146-158. DOI:10.1111/j.1574-6941.2011.01280.x |
| [29] | Gough H L, Stahl D A. Microbial community structures in anoxic freshwater lake sediment along a metal contamination gradient[J]. The ISME Journal, 2011, 5(3): 543-558. DOI:10.1038/ismej.2010.132 |
| [30] |
贺纪正, 王军涛. 土壤微生物群落构建理论与时空演变特征[J]. 生态学报, 2015, 35(20): 6575-6583. He J Z, Wang J T. Mechanisms of community organization and spatiotemporal patterns of soil microbial communities[J]. Acta Ecologica Sinica, 2015, 35(20): 6575-6583. |
| [31] |
褚海燕, 冯毛毛, 柳旭, 等. 土壤微生物生物地理学: 国内进展与国际前沿[J]. 土壤学报, 2020, 57(3): 515-529. Chu H Y, Feng M M, Liu X, et al. Soil microbial biogeography: recent advances in China and research frontiers in the world[J]. Acta Pedologica Sinica, 2020, 57(3): 515-529. |
| [32] | Chase J M, Myers J A. Disentangling the importance of ecological niches from stochastic processes across scales[J]. Philosophical Transactions of the Royal Society B: Biological Sciences, 2011, 366(1576): 2351-2363. DOI:10.1098/rstb.2011.0063 |
| [33] | Dini-Andreote F, Stegen J C, van Elsas J D, et al. Disentangling mechanisms that mediate the balance between stochastic and deterministic processes in microbial succession[J]. Proceedings of the National Academy of Sciences of the United States of America, 2015, 112(11): E1326-E1332. DOI:10.1073/pnas.1414261112 |
| [34] | Jiao S, Yang Y F, Xu Y Q, et al. Balance between community assembly processes mediates species coexistence in agricultural soil microbiomes across Eastern China[J]. The ISME Journal, 2020, 14(1): 202-216. DOI:10.1038/s41396-019-0522-9 |