 2021, Vol. 42
2021, Vol. 42



2. 中国科学院大学, 北京 100049
2. University of Chinese Academy of Sciences, Beijing 100049, China
近年来, 发现了一种新的微生物氨氧化过程, 即厌氧铁氨氧化.在2006年, 日本学者Sawayama[1]的研究发现, 在厌氧条件下, 微生物能利用Fe(Ⅲ)做电子受体将NH4+-N氧化成NO2--N、N2和NO3--N, 并将此过程称为厌氧铁氨氧化反应.已有研究发现在水稻土、湿地沉积物和湿地土壤中均有厌氧铁氨氧化现象发生[2~4], 厌氧铁氨氧化广泛发生在各种不同生态环境中, 在自然界氮循环中起着重要作用.同时, 厌氧铁氨氧化过程无需有机碳源和曝气等优点给生物脱氮带来了新思路, 也为氮铁循环的研究提供了新方向[5].然而厌氧铁氨氧化反应过程复杂, 如下列方程所示[6]:
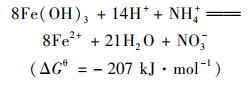
|
(1) |
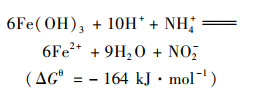
|
(2) |
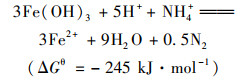
|
(3) |
反应产物有NO2-、NO3-和N2, 对FEAMMOX反应产物尚缺少调控的策略和方法[7].这些过程都是由微生物进行, 然而目前只分离纯化得到一株有铁还原和氨氧化功能的酸微菌A6[8], 对相关微生物群落结构、功能及相互作用等认识不深, 这些都是探明厌氧铁氨氧化微生物作用过程的主要阻碍.因此, 了解厌氧铁氨氧化过程微生物群落特征、相关功能及微生物间的相互作用关系, 对阐明厌氧铁氨氧化过程的生物学机制至关重要.
在微生物参与的铁循环过程中, 铁化合物固体-溶解状态的转换, 对微生物群落非常重要.Fe(Ⅲ)作为厌氧铁呼吸的末端电子受体, Fe(Ⅲ)矿物的溶解性能够影响微生物对Fe(Ⅲ)的利用[9].一般情况下, 微生物最易利用络合态Fe(Ⅲ), 其次是弱晶体Fe(Ⅲ)氧化物, 最难利用的是晶体Fe(Ⅲ)氧化物, 有些铁还原微生物比如Geothrix fermentans及Shewanella algae, 可分泌螯合物及电子穿梭体来增加Fe(Ⅲ)的利用率[10].因此对螯合铁的研究, 可以解决Fe(Ⅲ)的生物可利用性问题.厌氧铁氨氧化过程机制可通过富集培养相关菌种或驯化污泥的方式进行研究, 刘志文[11]将铁氨氧化菌制成磁性壳聚糖凝胶体, 吴悦溪等[12]将铁氨氧化污泥接种到厌氧反应器, 进行厌氧铁氨氧化脱氮研究.原核生物分类群的功能注释(FAPROTAX)方法能分析微生物群落中功能[13], 为阐明和预测生物地球化学过程提供重要的信息, 可以了解污泥中氮铁循环的联系及其相关功能注释强度[14].微生物之间相互作用是影响群落功能的重要因素, 微生物共现网络分析通过数学模型分析微生物互作关系[15], 可获得厌氧铁氨氧化过程群落的关键节点及规律.
本研究选取腐殖酸铁、柠檬酸铁、乙二胺四乙酸铁钠和氨三乙酸铁这4种类型螯合铁来研究螯合铁介导的厌氧铁氨氧化过程, 分析反应器水质变化, 并探讨污泥中微生物群落组成、功能及互作关系, 以期为厌氧铁氨氧化过程的微生物群落特征提供理论基础.
1 材料与方法 1.1 实验材料本实验所用模拟废水为:NH4Cl 1.0 g·L-1、NaHCO3 1.0 g·L-1、KH2PO4 0.20 g·L-1、MgSO4 0.25 g·L-1、维生素1.0 mL·L-1和微量元素溶液1.0 mL·L-1, 试剂均为分析纯(成都市科隆化学品有限公司).本实验所用微量元素包含:MnCl2·4H2O 0.10 g·L-1、H3BO3 0.10 g·L-1、FeSO4·7H2O 0.20 g·L-1、CoCl2·6H2O 0.10 g·L-1、CuCl2·2H2O 0.01 g·L-1、ZnCl2 0.01 g·L-1、Na2MoO4·2H2O 0.02 g·L-1和NiCl2·6H2O 0.02 g·L-1.本实验所用维生素为:维生素B1 0.05 g·L-1、维生素B12 0.05 g·L-1、烟酸0.05 g·L-1、对氨基苯甲酸0.03 g·L-1、吡哆酸0.01 g·L-1、生物素0.05 g·L-1、肌酸0.01 g·L-1、叶酸0.01 g·L-1、抗坏血酸0.01 g·L-1和柠檬酸0.01 g·L-1.铁源:Fe(Ⅲ)(30 mmol·L-1)分别由4种螯合铁提供, 乙二胺四乙酸铁钠30 mmol·L-1、柠檬酸铁30 mmol·L-1、氨三乙酸铁45 mmol·L-1和腐殖酸铁45 mmol·L-1.
本实验在1 000 mL广口瓶反应器中进行, 每个反应器中加入800 mL营养液和15 g稻田土, 通入氮气约15 min进行除氧操作, 最后用胶塞密封, 使装置保持在厌氧状态.置于33℃摇床(90 r·min-1)中放置77 d, 运行期间考察废水中氮的变化, 污泥微生物群落结构、功能及相互作用关系.
1.2 水质分析方法分析方法均采用国家水质检测标准测定[16].NH4+-N采用纳氏试剂分光光度法测定, NO2--N采用N-(1-萘基)-乙二胺比色法测定, NO3--N采用紫外分光光度法测定.测定时, 将样品溶液根据各指标的标准曲线稀释适宜的倍数, 使用紫外分光光度计(UV-1800, 岛津)测定.
1.3 微生物多样性分析16S rRNA高通量测序:反应初始时, 将要加入的污泥混匀取样并立即放入-80℃冰箱保存, 在反应结束后取污泥样品, 最后将样品送至上海美吉生物医药科技有限公司完成测序, PCR反应所用引物为515F/907R.
共现网络分析:基于微生物高通量测序结果, 在美国俄克拉荷马大学环境基因组学研究所网站(http://ieg2.ou.edu/MENA)进行网络构建, 通过MENA(molecular ecological network analysis)构建微生物共现网络并计算相关特征参数, 然后利用Cytoscape软件(3.6.1版)对网络进行可视化分析.
微生物功能预测分析:下载FAPROTAX(functional annotation of prokaryotic taxa)软件及其Python脚本[13], 在Python 2.7版本下运行软件, 结果使用R3.5.3进行分析绘图.
2 结果与讨论 2.1 反应过程水中氮素变化分析从图 1(a)可知, 经过77 d的厌氧反应过程, 不同螯合铁类型脱氮效果有显著差别.其中腐殖酸铁组的氨氮去除效果最好, 达到83.68%左右, 总氮去除率约83.32%; 乙二胺四乙酸铁钠组对氨氮的去除率达到55.59%, 总氮去除率约55.07%; 柠檬酸铁组氨氮去除率达到51.43%, 总氮去除率约43.67%; 氨三乙酸铁组在这个反应体系中对氨氮去除效果较差, 去除率约12.93%, 总氮去除率约12.65%. 4种螯合铁类型中, 腐殖酸铁脱氮效果最好.

|
图 1 反应过程中水质变化曲线 Fig. 1 Curve of change in water quality during the experiment |
图 1(b)是各组氨氮浓度曲线, 反应的初始阶段(0~7 d), 腐殖酸铁组的氨氮去除量约为42 mg·L-1, 高于其它实验, 可知在初始阶段腐殖酸铁组氨氮去除速度更快.在反应的8~77 d, 腐殖酸铁组、柠檬酸铁组、乙二胺四乙酸铁钠组中氨氮浓度均持续稳定下降.其中腐殖酸铁组氨氮去除效果略高于其它组, 并且各组亚硝态氮浓度[图 1(c)]都稳定保持在30 mg·L-1以内, 各组硝态氮浓度[图 1(d)]都处于较低水平(<6 mg·L-1).由于各实验组均处于厌氧状态, 且Fe(Ⅲ)作为唯一的电子受体, 因此可认为反应过程中NO2-和NO3-是由微生物利用Fe(Ⅲ)将氨氮氧化所生成[17].
本实验中, 由于NO2-和NO3-浓度远低于氨氮去除量, 氨氮去除率与总氮去除率接近, 因此推测反应过程中大部分氮素转化为气体.在能量方面上, FEAMMOX反应更容易产生N2[18], 从而推测本实验NH4+转化的产物主要为N2.表 1是铁氨氧化在污水脱氮方面的研究, 在已有报道的厌氧铁氨氧化研究中, 大部分以铁氧化物进行实验, 产物以NO3-和NO2-为主, 总氮去除率在11.30%~71.80%[11, 12, 19~22], 同时反应pH值处于4~5之间, 这是因为在中性条件下, Fe(OH)3溶解度比较低, 会阻碍微生物对Fe(Ⅲ)的利用[23]; 由于螯合铁在中性条件下能保持络合状态, 可促进微生物对Fe(Ⅲ)的利用, 因此螯合铁能在中性条件下有较好的脱氮效果, 这些可推断出铁形态会对厌氧铁氨氧化产物有一定的影响.本实验结果表明, 腐殖酸铁对于厌氧铁氨氧化的作用效果最好, 腐殖酸铁中有醌类结构, 目前含有醌类结构物质已被证明可以充当电子中介体, 能提高铁还原细菌对Fe(Ⅲ)的利用, 从而促进厌氧铁氨氧化反应的进行[24].
|
|
表 1 厌氧铁氨氧化污水脱氮 Table 1 Denitrification of wastewater by FEAMMOX |
2.2 微生物α多样性分析
首先对微生物群落进行α多样性分析, 主要是分析每个样本中微生物群落本身的多样性, Chao指数和Ace指数体现样品中微生物群落物种丰富度, 在样品中, 这两个指数数值越高, 则代表该样品群落物种丰富度越高; Shannon指数和Simpson指数能体现样品中微生物群落多样性, Shannon指数越高, 样品中微生物群落多样性越高, Simpson指数是优势度指数, 其值越低, 则说明该样品微生物多样性越高.表 2是Chao指数、Ace指数、Shannon指数、Simpson指数以及群落覆盖度, 从中可知, 氨三乙酸铁组中OTU数、Ace指数、Chao指数和Shannon指数均低于其它组, 并且Simpson指数最高, 这些说明氨三乙酸铁组中物种丰富度和群落多样性都低于其它实验组; 乙二胺四乙酸铁钠组中Ace指数和Chao指数均高于其它实验组, 表明该组中微生物群落中物种丰富度较其它实验组高; 腐殖酸铁组中Shannon指数高于其它实验组, 同时其Simpson指数最小, 这些表明腐殖酸铁组中微生物群落多样性较其它实验组高.本次测序的覆盖度均在99.15%以上, 说明此次测序结果能较好地代表污泥样品中微生物群落组成的真实情况.
|
|
表 2 污泥微生物α多样性指数 Table 2 The α diversity index table of microbial communities |
2.3 微生物群落组成分析
对污泥样品微生物进行科水平上分析, 图 2(a)是各污泥样品群落在科水平上的韦恩图, 可以清晰了解各样品微生物群落之间的异同情况, 初始污泥微生物中有56个特有的科, 在经过厌氧铁氨氧化过程后, 污泥中特有科逐渐减少, 说明有些微生物类群不适合该反应体系, 螯合铁氨氧化体系中的微生物表现出趋简性[25].由图 2(b)可知, 在反应结束后, 各实验组污泥中Steroidobacteraceae和Rhodocyclaceae含量较初始污泥低, Clostridiaceae、Comamonadaceae和Anaerolineaceae含量增加, Anaerolineaceae是厌氧消化核心微生物种群[26].因此可以推断出, 在反应过程中, 污泥中有特定功能菌群成为优势菌群, 相对丰度提高, 有些菌群则被淘汰, 体系中形成了相对单一稳定的微生物群落.

|
(a)基于科水平微生物群落韦恩图, (b)污泥中优势菌的科水平对比; A1:初始污泥, A2:腐殖酸铁组, A3:柠檬酸铁组, A4:氨三乙酸铁组, A5:乙二胺四乙酸铁钠组, 下同; ①Rhizobiaceae, ②Clostridiaceae, ③Xanthomonadaceae, ④Comamonadaceae, ⑤Anaerolineaceae, ⑥Steroidobacteraceae, ⑦Rhodocyclaceae 图 2 反应结束后污泥在科水平上微生物群落 Fig. 2 Microbial community analysis of sludge at the family level |
图 3是在属水平上进行微生物群落结构分析, 初始污泥中的优势菌主要有norank_f_Steroidobacteraceae、norank_f_Caldilineaceae和Candidatus_Competibacter属, 丰度分别为7.26%、7.56%和4.76%, 其中Candidatus_Competibacter属对于脱氮除磷均有一定的作用[27].初始污泥中还存在极少量的unclassified_f_Comamonadaceae, 含量约为0.40%.腐殖酸铁组微生物群落组成主要有unclassified_f_Comamonadaceae, 丰度由初始污泥中0.40%增加到17.57%, Thermomonas含量约占1.76%; 目前有研究报道Comamonadaceae能够进行反硝化作用[28]并参与氨氧化为亚硝酸盐的过程[29], Thermomonas可以利用硝酸盐作为电子受体进行硝酸盐还原[30], 这表明腐殖酸铁组具有一定的脱氮能力, 并且添加腐殖酸铁可促进污泥中脱氮功能菌群的生长, 增强该体系的脱氮能力.柠檬酸铁组中优势菌群主要有Clostridium和Thiobacillus, 含量分别为47.70%和10.37%; 已经有相关文献报道Clostridium为铁还原菌[31], 因此, 柠檬酸铁组微生物能够进行铁代谢相关活动, 添加柠檬酸铁能促进相关铁还原菌的生长, 并有研究表明铁还原菌在厌氧铁氨氧化过程中起着重要作用, 其含量增加可能会促进厌氧铁氨氧化作用[32], 氨三乙酸铁组优势菌有Aquamicrobium, 丰度为62.33%, 该组中unclassified_f_Comamonadaceae的丰度只占0.08%, 这表明该组参与氮循环作用的菌群较少.乙二胺四乙酸铁钠组优势菌属主要是Thermomonas, 含量约20.11%, Shinella含量占18.71%, unclassified_f_Comamonadaceae丰度约0.81%, 其中Thermomonas能进行硝酸盐还原作用, 是参与脱氮作用的菌群[30].对各实验组污泥中微生物群落的变化分析可知, 在各组实验污泥中参与氮循环的微生物菌群丰度高于初始污泥, 其中腐殖酸铁组中氨氧化菌群Comamonadaceae显著增加[28, 29], 并远高于其它实验组, 这表明腐殖酸铁可能比其它螯合铁更适合厌氧铁氨氧化过程.

|
图 3 反应结束后污泥在属水平上微生物群落 Fig. 3 Microbial community analysis of sludge at the genus level |
FAPROTAX是基于微生物测序结果对微生物群落功能表型进行预测的方法, 该方法适合于对环境样品进行生物地球化学循环功能预测分析, 特别是碳、氢、氮、磷和硫的循环的功能注释和预测.图 4是FAPROTAX分析, 主要有14个与氮循环相关的功能注释, 包括硝酸盐还原、硝酸盐反硝化和亚硝酸盐反硝化等.各实验组功能预测中好氧氨氧化作用均为0, 这说明反应体系中没有发生好氧氨氧化过程, 进而可推断此体系中氨氧化是以Fe(Ⅲ)为电子受体进行的, 也表明该体系中存在厌氧铁氨氧化过程.根据聚类分析关系可知, 铁呼吸、硫呼吸、硫化物呼吸、硝酸盐反硝化及亚硝酸盐反硝化联系紧密, 也说明了氮循环、铁循环和硫循环关系密切, 铁硫代谢对脱氮有重要作用.目前也有报道硫铁反硝化脱氮方面的研究[33], 根据微生物群落组成进行分析.根据图 4中各功能注释强度可知, 腐殖酸铁组中铁呼吸强度达到578, 初始污泥中铁呼吸强度为40, 柠檬酸铁组中铁呼吸强度为12, 氨三乙酸铁组中铁呼吸强度为27, 乙二胺四乙酸铁钠组中铁呼吸为2, 腐殖酸铁组中铁呼吸强度远高于其它实验组, 表明在腐殖酸铁组中, 微生物发生铁代谢强度更高, 更易利用腐殖酸铁中Fe(Ⅲ).

|
图 4 反应结束后污泥微生物功能预测图 Fig. 4 Function prediction of microbial at the end of experiment |
微生物共现网络分析通常用于识别菌群之间的相互作用[34], MENA是基于随机矩阵原理(random matrix theory, RMT)的生态网络构建方法.表 3是网络的属性表, 可知网络是由122个节点和505条边组成, 其中节点表示不同菌群, 边表示不同菌群之间的连接关系.
|
|
表 3 网络的拓扑学指数 Table 3 Topological indices of network |
节点之间的边数量水平称为度, 度越强则相互关系越强[35], 因此网络中度较高的节点在网络中非常重要.此次网络节点度为1~27(图 5), 筛选度大于20的节点进行微生物门水平分析, 发现主要是Acidimicrobiia和Actinobacteria, 已有研究在Acidimicrobiia分离出厌氧铁氨氧化菌A6[8], 推测Acidimicrobiia和Actinobacteria在厌氧铁氨氧化螯合铁体系中有重要作用.微生物共现网络还可用于识别关键物种, 微生物生态系统中关键物种是高度关联的菌群, 它们可以单独或共同对微生物生态系统的结构和功能产生相当大的影响, 关键物种对于维持生态系统平衡有重要作用[36].在共现网络中, 通常根据中介中心性来确定关键物种, 具有最大中介中心性的菌群可以推断为关键物种[37, 38], 此次共现网络表明Tessaracoccus是螯合铁实验中微生物群落的关键物种, Tessaracoccus属于丙酸杆菌科, 能在厌氧条件下生长, Puente-Sánchez等[39]在黄铁矿地带分离出Tessaracoccus菌株, 该菌可以在厌氧条件下进行硝酸盐还原, 这表明Tessaracoccus也能参与氮循环过程, 并可能在螯合铁铁氨氧化体系中起着关键作用.

|
图 5 反应结束后污泥微生物共现网络分析 Fig. 5 Co-occurrence network analysis of microbial community at the end of the experiment |
(1) 经过77 d的厌氧反应, 4种类型的螯合铁中, 腐殖酸铁组脱氮效果最好, 氨氮去除率约83.68%, 总氮去除率约83.32%, 推测螯合铁进行厌氧铁氨氧化反应的产物主要是N2.
(2) 腐殖酸铁组中微生物群落多样性较其它实验组高, 腐殖酸铁组微生物脱氮菌群有unclassified_f_Comamonadaceae和Thermomonas, 菌群丰度分别为17.57%、1.76%, 其含量均高于其它实验组, 表明腐殖酸铁有利于脱氮菌群生长.总体上, 螯合铁氨氧化体系中的微生物表现出趋简性.
(3) FAPROTAX功能预测结果中, 铁循环、硫循环和氮循环关系密切, 铁硫代谢对于脱氮有重要作用; 腐殖酸铁组中铁呼吸功能注释强度为578, 腐殖酸铁组中微生物参与铁代谢循环较其它组活跃, 微生物更易利用腐殖酸铁中的Fe(Ⅲ).
(4) 共现网络分析表明, Acidimicrobiia和Actinobacteria在厌氧铁氨氧化螯合铁体系中具有重要作用, Tessaracoccus是螯合铁进行厌氧铁氨氧化反应的关键物种.
| [1] | Sawayama S. Possibility of anoxic ferric ammonium oxidation[J]. Journal of Bioscience and Bioengineering, 2006, 101(1): 70-72. DOI:10.1263/jbb.101.70 |
| [2] | Ding B J, Chen Z H, Li Z K, et al. Nitrogen loss through anaerobic ammonium oxidation coupled to iron reduction from ecosystem habitats in the Taihu estuary region[J]. Science of the Total Environment, 2019, 662: 600-606. DOI:10.1016/j.scitotenv.2019.01.231 |
| [3] | Clement J C, Shrestha J, Ehrenfeld J G, et al. Ammonium oxidation coupled to dissimilatory reduction of iron under anaerobic conditions in wetland soils[J]. Soil Biology and Biochemistry, 2005, 37(12): 2323-2328. DOI:10.1016/j.soilbio.2005.03.027 |
| [4] | Ding L J, An X L, Li S, et al. Nitrogen loss through anaerobic ammonium oxidation coupled to iron reduction from paddy soils in a chronosequence[J]. Environmental Science & Technology, 2014, 48(18): 10641-10647. |
| [5] | Ren Y, Ngo H H, Guo W S, et al. New perspectives on microbial communities and biological nitrogen removal processes in wastewater treatment systems[J]. Bioresource Technology, 2020, 297. DOI:10.1016/j.biortech.2019.122491 |
| [6] |
钟小娟, 王亚军, 唐家桓, 等. 铁氨氧化: 新型的厌氧氨氧化过程及其生态意义[J]. 福建农林大学学报(自然科学版), 2018, 47(1): 1-7. Zhong X J, Wang Y J, Tang J H, et al. Feammox: a novel anammox process and ecological significance[J]. Journal of Fujian Agriculture and Forestry University (Natural Science Edition), 2018, 47(1): 1-7. |
| [7] |
李祥, 林兴, 杨朋兵, 等. 活性污泥厌氧Fe(Ⅲ)还原氨氧化现象初探[J]. 环境科学, 2016, 37(8): 3114-3119. Li X, Lin X, Yang P B, et al. Simultaneous ferric reduction with ammonia oxidation phenomena in activated sludge in anaerobic environment[J]. Environmental Science, 2016, 37(8): 3114-3119. |
| [8] | Huang S, Jaffé P R. Characterization of incubation experiments and development of an enrichment culture capable of ammonium oxidation under iron-reducing conditions[J]. Biogeosciences, 2015, 12(3): 769-779. DOI:10.5194/bg-12-769-2015 |
| [9] |
覃海华. 异化铁还原对重金属形态转化及其产氢过程研究[D]. 天津: 天津科技大学, 2018. Tan H H. Research on transformation of heavy metals and hydrogen-producing process by dissimilatory Fe(Ⅲ) reduction[D]. Tianjin: Tianjin University of Science and Technology, 2018. |
| [10] |
司友斌, 王娟. 异化铁还原对土壤中重金属形态转化及其有效性影响[J]. 环境科学, 2015, 36(9): 3533-3542. Si Y B, Wang J. Influence of dissimilatory iron reduction on the speciation and bioavailability of heavy metals in soil[J]. Environmental Science, 2015, 36(9): 3533-3542. |
| [11] |
刘志文. 厌氧铁氨氧化菌的固定化及其处理氨氮废水的研究[D]. 广州: 广东工业大学, 2018. Liu Z W. Study on the treatment of ammonium in wastewater by immobilized Feammox bacteria[D]. Guangzhou: Guangdong University of Technology, 2018. |
| [12] |
吴悦溪, 曾薇, 刘宏, 等. Feammox系统内氮素转化途径的研究[J]. 化工学报, 2020, 71(5): 2265-2272. Wu Y X, Zeng W, Liu H, et al. Exploration of nitrogen transformation pathway in Feammox[J]. CIESC Journal, 2020, 71(5): 2265-2272. |
| [13] | Louca S, Jacques S M S, Pires A P F, et al. High taxonomic variability despite stable functional structure across microbial communities[J]. Nature Ecology & Evolution, 2017, 1(1). DOI:10.1038/s41559-016-0015 |
| [14] | Hochstein R, Zhang Q, Sadowsky M J, et al. The deposit feeder Capitella teleta has a unique and relatively complex microbiome likely supporting its ability to degrade pollutants[J]. Science of the Total Environment, 2019, 670: 547-554. DOI:10.1016/j.scitotenv.2019.03.255 |
| [15] |
朱瑞芬, 刘杰淋, 王建丽, 等. 基于分子生态学网络分析松嫩退化草地土壤微生物群落对施氮的响应[J]. 中国农业科学, 2020, 53(13): 2637-2646. Zhu R F, Liu J L, Wang J L, et al. Molecular ecological network analyses revealing the effects of nitrogen application on soil microbial community in the degraded grasslands[J]. Scientia Agricultura Sinica, 2020, 53(13): 2637-2646. DOI:10.3864/j.issn.0578-1752.2020.13.012 |
| [16] | 国家环境保护总局. 水和废水监测分析方法[M]. 北京: 中国环境科学出版社, 1989. |
| [17] | Yin S Y, Li J, Dong H Y, et al. Enhanced nitrogen removal through marine anammox bacteria (MAB) treating nitrogen-rich saline wastewater with Fe(Ⅲ) addition: nitrogen shock loading and community structure[J]. Bioresource Technology, 2019, 287. DOI:10.1016/j.biortech.2019.121405 |
| [18] | Yang W H, Weber K A, Silver W L. Nitrogen loss from soil through anaerobic ammonium oxidation coupled to iron reduction[J]. Nature Geoscience, 2012, 5(8): 538-541. DOI:10.1038/ngeo1530 |
| [19] | Li X, Huang Y, Liu H W, et al. Simultaneous Fe(Ⅲ) reduction and ammonia oxidation process in Anammox sludge[J]. Journal of Environmental Sciences, 2018, 64(2): 42-50. |
| [20] |
吴胤, 陈琛, 毛小云, 等. 基于Feammox的生物膜反应器性能研究[J]. 中国环境科学, 2017, 37(9): 3353-3362. Wu Y, Chen C, Mao X Y, et al. Study on performance of the Feammox biofilm-reactor[J]. China Environmental Science, 2017, 37(9): 3353-3362. DOI:10.3969/j.issn.1000-6923.2017.09.019 |
| [21] |
吴彦成, 顾鑫, 朱继涛, 等. 铁氨氧化污水生物脱氮技术的研究进展[J]. 中国给水排水, 2020, 36(18): 38-44. Wu Y C, Gu X, Zhu J T, et al. Research advances of biological nitrogen removal from wastewater via Fe(Ⅲ) reduction coupled to anaerobic ammonium oxidation (Feammox) process[J]. China Water & Wastewater, 2020, 36(18): 38-44. |
| [22] | Yang Y F, Zhang Y B, Li Y, et al. Nitrogen removal during anaerobic digestion of wasted activated sludge under supplementing Fe(Ⅲ) compounds[J]. Chemical Engineering Journal, 2018, 332: 711-716. DOI:10.1016/j.cej.2017.09.133 |
| [23] |
苏润华, 丁丽丽, 任洪强. 纳米零价铁(NZVI)对厌氧产甲烷活性、污泥特性和微生物群落结构的影响[J]. 环境科学, 2018, 39(7): 3286-3296. Su R H, Ding L L, Ren H Q. Impact of nano zero-valent iron (NZVI) on methanogenic activity, physiological traits, and microbial community structure in anaerobic digestion[J]. Environmental Science, 2018, 39(7): 3286-3296. |
| [24] | Dunham-Cheatham S, Mishra B, Myneni S, et al. The effect of natural organic matter on the adsorption of mercury to bacterial cells[J]. Geochimica Et Cosmochimica Acta, 2015, 150: 1-10. DOI:10.1016/j.gca.2014.11.018 |
| [25] | Goldford J E, Lu N X, Baji Dc' D, et al. Emergent simplicity in microbial community assembly[J]. Science, 2018, 361(6401): 469-474. DOI:10.1126/science.aat1168 |
| [26] |
宋壮壮, 吕爽, 刘哲, 等. 厌氧氨氧化耦合反硝化工艺的启动及微生物群落变化特征[J]. 环境科学, 2019, 40(11): 5057-5065. Song Z Z, Lv S, Liu Z, et al. Start-up of simultaneous ANAMMOX and denitrification process and changes in microbial community characteristics[J]. Environmental Science, 2019, 40(11): 5057-5065. |
| [27] | Rubio-Rincón F J, Lopez-Vazquez C M, Welles L, et al. Cooperation between Candidatus competibacter and Candidatus accumulibacter clade Ⅰ, in denitrification and phosphate removal processes[J]. Water Research, 2017, 120: 156-164. DOI:10.1016/j.watres.2017.05.001 |
| [28] | Wu Y C, Shukal S, Mukherjee M, et al. Involvement in denitrification is beneficial to the biofilm lifestyle of Comamonas testosteroni: a mechanistic study and its environmental implications[J]. Environmental Science & Technology, 2015, 49(19): 11551-11559. |
| [29] | Bao P, Li G. Sulfur-driven iron reduction coupled to anaerobic ammonium oxidation[J]. Environmental Science and Technology, 2017, 51(12): 6691-6698. DOI:10.1021/acs.est.6b05971 |
| [30] | Chen H, Zhao X H, Cheng Y Y, et al. Iron robustly stimulates simultaneous nitrification and denitrification under aerobic conditions[J]. Environmental Science & Technology, 2018, 52(3): 1404-1412. |
| [31] | Reyes C, Schneider D, Thürmer A, et al. Potentially active iron, sulfur, and sulfate reducing bacteria in Skagerrak and Bothnian Bay Sediments[J]. Geomicrobiology Journal, 2017, 34(10): 840-850. DOI:10.1080/01490451.2017.1281360 |
| [32] | Peng Q A, Shaaban M, Wu Y P, et al. The diversity of iron reducing bacteria communities in subtropical paddy soils of China[J]. Applied Soil Ecology, 2016, 101: 20-27. DOI:10.1016/j.apsoil.2016.01.012 |
| [33] |
付炳炳, 潘建新, 马景德, 等. 采用含硫铁化学污泥作为反硝化电子供体进行焦化废水中总氮深度去除[J]. 环境科学, 2018, 39(7): 3262-3270. Fu B B, Pan J X, Ma J D, et al. Evaluation of advanced nitrogen removal from coking wastewater using sulfide iron-containing sludge as a denitrification electron donor[J]. Environmental Science, 2018, 39(7): 3262-3270. |
| [34] | Berry D, Widder S. Deciphering microbial interactions and detecting keystone species with co-occurrence networks[J]. Frontiers in Microbiology, 2014, 5. DOI:10.3389/fmicb.2014.00219 |
| [35] |
闫冰, 夏嵩, 桂双林, 等. 厌氧氨氧化菌富集培养过程微生物群落结构及多样性[J]. 环境科学, 2020, 41(12): 5535-5543. Yan B, Xia S, Gui S L, et al. Microbial community structure and diversity during the enrichment of anaerobic ammonium oxidation bacteria[J]. Environmental Science, 2020, 41(12): 5535-5543. |
| [36] | Banerjee S, Schlaeppi K, Van Der Heijden M G A. Keystone taxa as drivers of microbiome structure and functioning[J]. Nature Reviews Microbiology, 2018, 16(9): 567-576. DOI:10.1038/s41579-018-0024-1 |
| [37] | Banerjee S, Kirkby C A, Schmutter D, et al. Network analysis reveals functional redundancy and keystone taxa amongst bacterial and fungal communities during organic matter decomposition in an arable soil[J]. Soil Biology and Biochemistry, 2016, 97: 188-198. DOI:10.1016/j.soilbio.2016.03.017 |
| [38] | Jiang Y J, Li S Z, Li R P, et al. Plant cultivars imprint the rhizosphere bacterial community composition and association networks[J]. Soil Biology and Biochemistry, 2017, 109: 145-155. DOI:10.1016/j.soilbio.2017.02.010 |
| [39] | Puente-Sánchez F, Sánchez-Román M, Amils R, et al. Tessaracoccus lapidicaptus sp. nov., an actinobacterium isolated from the deep subsurface of the Iberian Pyrite Belt[J]. International Journal of Systematic and Evolutionary Microbiology, 2014, 64(10): 3546-3552. |