 2021, Vol. 42
2021, Vol. 42


2. 吉首大学生物资源与环境科学学院, 吉首 416000;
3. 湘西土家族苗族自治州生态环境局, 吉首 416000
2. College of Biology and Environmental Sciences, Jishou University, Jishou 416000, China;
3. Ecology and Environment Bureau of Xiangxi Tujiazu & Miaozu Autonomous Prefecture, Jishou 416000, China
重金属尾矿是有色金属矿山开采后选矿过程中排放的固体废弃物, 通常以尾矿浆的形式排出, 排出后一般堆放在尾矿库内, 闭库后形成尾矿废弃地[1].重金属尾矿的治理是世界性的难题, 因为它的理化性质非常极端, 主要表现在物理结构不良、持水保肥能力差、养分贫乏、重金属毒性强和酸化潜力高等方面[2, 3].传统的尾矿治理方式主要包括物理固定法, 化学改良法等, 这些方法均存在费用高、工程量大和易造成二次污染等问题[4].基于植被重建的重金属尾矿生态恢复具有经济、环保等优点, 在世界范围内获得了广泛的认可.该技术的核心思想是通过辅助措施, 在重金属尾矿上重新建立植被, 并促使它们逐步演替成为能自维持的和稳定的生态系统, 从而实现将重金属污染长期固定在原地[5, 6].
过去20a, 生物多样性与生态系统功能的关系一直是生态学、土壤学和环境科学等学科共同关注的重大理论问题[7, 8].有试验证明物种多样性有利于提高生态系统的功能, 如增加植被生产力、改善土壤理化性质、提高保肥持水能力和减少重金属在土壤-植被系统的毒害作用等[9, 10].Tilman等[11~13]在美国Minnesota草地试验区建立不同物种组成的小区试验, 发现物种多样性导致更高的群落生产力、更高的系统稳定性和更高的抗入侵能力, 对生态系统稳定性有积极的影响.Li等[14, 15]和马頔[16]通过藻类的微宇宙(microcosm)试验发现, 物种多样性有利于提高受重金属污染生态系统的生物量(生产力), 构建包括重金属耐性种和非耐性种的多样性群落, 能更有效地去除重金属污染物.杨扬等[17]通过室内控制试验研究不同植物物种多样性对植物生长与土壤Cd污染修复的影响.结果表明, 植物物种多样性的提高不仅显著促进了植物地下生物量和总生物量的积累, 而且削弱Cd对植物的毒害作用, 还能通过改变植物根系形态和生物量来提高对土壤Cd污染的修复.虽然过往研究已经证明增加生物多样性能够提高生态系统功能; 但是这些理论研究成果并没有在退化系统的生态恢复方面产生广泛的影响[18].而且这些研究多处于模拟或室内阶段, 目前仍不能确定室内条件下的研究结果能否很好地反映野外生态修复的实际[19].在污染环境或退化的生态系统中, 增强生态系统的功能显得尤为重要.尾矿废弃地作为新形成的生态系统, 具有植被缺乏、重金属浓度高、养分不足、微生物群落结构简单和土壤团聚体尚未形成等特点[2, 3].对于研究植物物种多样性在生态系统恢复中的作用及其机制, 尾矿生态系统是非常理想的自然实验室[20].本文选取典型的重金属尾矿作为研究对象, 开展野外田间试验, 在重金属尾矿上建立具有物种多样性梯度的植被, 分析物种多样性对植物群落、重金属迁移和营养元素累积的影响, 揭示物种多样性影响重金属尾矿生态恢复效率的内在机制, 以期为物种多样性在重金属尾矿生态恢复中的应用提供科学依据.
1 材料与方法 1.1 研究地点概况本研究是在湖南省湘西州花垣县浩宇化工有限公司Pb-Zn尾矿库进行的(109°11′~109°15′E, 28°44′~28°47′N).湘西州地处湖南省西北部, 湘、渝和黔边陲, 境内具有丰富的矿产资源和植物资源, 铅锌矿和锰矿储量分别居全省第一位和全国第二位, 素有“有色金属之乡”的美称[21].矿区为丘陵地貌, 海拔高度为300~1 800 m, 属中亚热带山地气候, 年平均气温15~16.9℃, 年降水量为1 250~1 500 mm, 多年平均日照时数1 291~1 406 h.原始植被为中亚热带典型山地植被, 光热水基本同季, 有利于植物生长.浩宇铅锌尾矿库闭库时间为3 a, 面积约50 000 m2.对该尾矿库进行了生态调查和立地分析, 初步结果表明:尾矿弱碱性, pH值为8.5左右; 重金属Cd和Zn含量较高, 总Cd和总Zn含量范围分别为13.33~34.56 mg ·kg-1和841.60~2 441.40 mg ·kg-1, DTPA-Cd和DTPA-Zn含量范围分别为0.14~0.42mg ·kg-1和31.64~110.566 mg ·kg-1; 肥力差, 有机质、总氮和总磷含量均值分别为3.39 g ·kg-1、47.45 mg ·kg-1和16.40 mg ·kg-1; 土质疏松(黏粒13.05%、粉粒22.14%和沙粒65.33%), 未形成团粒结构, 植被无法定居而处于裸露状态(表 1).
|
|
表 1 尾矿的基本理化性质(n=10) Table 1 General physico-chemical properties of the Pb-Zn mine tailings(n=10) |
1.2 供试植物准备与功能群划分
以湘西浩宇化工铅锌尾矿库为中心, 从附近区域采集土著植物30种, 以采集的30种植物种子作为供试植物材料, 开展室内盆栽试验, 设置2种试验处理, 即将供试植物分别种在采自该尾矿库和附近未受重金属污染农用地两种土壤中; 每处理设置3个重复, 根据供试植物种子的大小, 每个重复的播种量设为30颗种子; 记录种子的萌发率, 种子萌发3个月以后, 收获所有存活植株, 测定植物的生物量, 并计算各个供试物种的重金属耐性指数(耐性指数=生长在尾矿废弃地土壤的植物生物量÷生长在未受重金属污染农用地土壤的植物生物量), 耐性指数大于或等于1的为重金属耐性种, 小于1的为非耐性种.根据种子的萌发率筛选出16种植物作为供试物种, 分别是狼尾草(Pennisetum alopecuroides)、画眉草(Eragrostis pilosa)、芒(Miscanthus sinensis)、粟草(Milium effusum)、硬杆子草(Capillipedium assimile)、狗牙根(Cynodon dactylon)、苍耳(Xanthium sibiricum)、魁蒿(Artemisia princeps)、黄花蒿(Artemisia annua)、马棘(Indigofera pseudotinctoria)、紫穗槐(Amorpha fruticosa)、胡枝子(Lespedeza bicolor)、刺槐(Robinia pseudoacacia)、金合欢(Acacia farnesiana)、斑花败酱(Patrinia punctiflora)和苎麻(Boehmeria nivea).其中, 狼尾草、芒、狗牙根、苍耳、刺槐和苎麻的耐性指数大于或等于1, 在本试验中认为是重金属耐性物种, 其余10种为非耐性物种.
1.3 试验设计、样品采集和分析在湘西浩宇化工有限公司铅锌尾矿库建立了具有不同物种多样性的植被试验小区.物种多样性以盆栽试验筛选出的16种土著植物作为物种库, 设置4个多样性梯度处理, 分别记为1、4、8和16.物种多样性为1的处理(单种)设置3个重复, 16个供试物种合计建立48个试验小区; 物种多样性为4和8的处理均设置20个重复, 每个重复的物种组成均是从物种库随机抽取的物种的组合, 且同一梯度处理下各个重复的物种组成不完全相同, 合计建立40个试验小区; 物种多样性为16的处理设置10个重复, 合计建立10个试验小区; 另外建立4个不种植物的对照试验小区; 总计建立了102个试验小区.田间试验按完全随机设计, 每个试验小区面积为4 m2(2 m×2 m).播种前将中药渣和鸡粪[15 t ·hm-2, 中药渣∶鸡粪=2 ∶1(质量比)]作为改良剂平铺在试验小区的地表, 采用犁耕法将其与0~20 cm的尾矿混匀.播种量根据上述室内盆栽试验结果确定, 采用撒播法, 播种于试验小区.试验期间植物在自然条件下生长, 不采取灌溉或其他农艺措施.
2 a后对植被小区进行全面调查, 采用数码照相法测算每个试验小区的植被盖度.在各个试验小区收获部分植株(0.5 m×2 m条带)的地上部分, 测定各目标物种的生物量(干重), 并据此计算每个试验小区植被总的生物量(生产力)以及各目标物种的生物量.各目标物种在完成生物量测定后, 将每个植物样品粉碎后, 过60目尼绒筛, 用于植物重金属含量的分析, 植物样品共计223个.
收获植物后, 分别从各个试验小区(包括对照小区)按梅花布点法采集5个亚土样混合成1个土壤样品, 采集深度0~20 cm, 采集量1 kg左右, 合计采集土壤样品102个.样品采集后立即装入塑料密封袋, 做好标记, 运回实验室, 在室温下自然风干, 用于土壤营养元素与重金属含量分析.
总有机碳和水溶性碳采用重铬酸钾氧化容量法[22], 总氮采用半微量凯氏定氮法, 铵态氮采用靛酚蓝比色法[23], 总磷和有效磷采用钼锑抗比色法[24], 有效钾采用乙酸铵浸提法[25].植物重金属含量采用浓硝酸-高氯酸消解法[26], 土壤重金属总量和总钾采用王水消解法[27], 土壤重金属有效态含量采用DTPA浸提法[28], 植物、土壤消解液和浸提液中的重金属元素(Cd、Cu、Pb和Zn)含量采用iCAP6300型电感耦合等离子体发射光谱仪(ICP-AES美国, 热电)测定.
1.4 数据统计与分析数据统计分析采用word 2010和SPSS 19.0, 作图使用Origin 8.0和Sigmaplot 11.
物种多样性效应机制采用下列公式计算, 物种多样性净效应分割求出选择效应和互补效应[29].
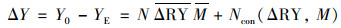
|
式中, ΔY是净效应, Y0是混种群落的总产量, YE是混种群落的期望总产量, NΔRY M是互补效应, Ncon(ΔRY, M)是选择效应, N是混种样方的物种数, ΔRY是物种在混种时相对观察值与期望值间的偏差, M是单种平均值, ΔRY是观察值和期望值偏差与单种观察值的协方差.
2 结果与分析 2.1 物种多样性对尾矿生态恢复中植被盖度和生产力的影响物种多样性对尾矿生态恢复中植被盖度和生产力的影响见图 1.物种多样性植被群落小区在铅锌尾矿上建立2 a后, 1-物种、4-物种、8-物种和16-物种植被盖度均值分别为33.4%、59.1%、66.6%和78.4%; 生物量均值分别为66.7、142.1、152.4和183.8 g ·m-2.对植被盖度和生产力随物种多样性的增加进行线性回归发现, 随着植物物种多样性的提高(也就是物种丰富度从1提高到16), 植物群落的植被盖度(r=0.53, P < 0.001)和生产力(r=0.52, P < 0.001)都是显著提高的.具体地说, 物种丰富度为16的时候, 试验小区的植被覆盖度和生物量均值分别为78.4%和183.8 g ·m-2(干重), 是物种丰富度为1的试验小区均值的2.4倍和2.8倍.这说明物种多样性的增加显著增加了植物群落的植被盖度和生产力, 初步确定植物物种多样性对尾矿生态恢复有促进作用.

|
图 1 植物物种多样性对植被盖度和生产力的影响(n=102) Fig. 1 Relationship between species diversity and plant coverage and biomass of the plots (n=102) |
物种多样性对尾矿生态恢复中植被重金属含量的影响见图 2.在尾矿上建立的植物群落地上部分重金属(Cd、Cu、Pb和Zn)含量的均值分别为0.36~11.62、3.36~36.69、0.77~30.04和103.56~1 384.62 mg ·kg-1, 对植物地上部分重金属含量随物种多样性的增加进行线性回归发现, 随着植物物种多样性的提高, 在尾矿上建立的植物地上部分重金属Cd(r=-0.16, P < 0.01)、Cu(r=-0.23, P < 0.001)、Pb(r=-0.15, P < 0.05)和Zn(r=-0.18, P < 0.001)的含量是逐渐下降的.这说明物种多样性的增加显著降低了植物对重金属的累积能力, 这将有利于降低由于动物取食以及枯枝落叶带来的生态风险.

|
图 2 植物物种多样性对植物地上部分重金属含量的影响(n=223) Fig. 2 Relationship between species diversity and heavy metal concentrations in the aboveground parts of the plants (n=223) |
物种多样性对尾矿生态恢复中铅锌尾矿营养元素的影响见图 3.对尾矿营养元素含量随物种多样性的增加进行线性回归发现, 随着植物物种多样性的提高, 总有机碳(TOC)、总氮(TN)和总磷(TP)累积量显著性增加(TOC: r=0.30, P < 0.001; TN: r=0.24, P < 0.05; TP: r=0.20, P < 0.05), 全钾(TK)累积量没有显著性变化(TK: r=0.03, P=0.72).此外, 水溶性有机碳的累积量随着植物物种多样性增加显著性增加(SOC: r=0.20, P < 0.11), 铵态氮(NH4+-N), 有效磷(AP)和有效钾(AK)的累积量随物种多样性的增加没有显著性变化(NH4+-N: r=0.06, P=0.50; AP: r=0.09, P=0.38; AK: r=-0.03, P=0.76).

|
图 3 植物物种多样性对尾矿营养元素的影响(n=102) Fig. 3 Relationship between species diversity and nutrient parameters in the mine tailings (n=102) |
物种多样性对尾矿生态恢复中铅锌尾矿重金属含量的影响见图 4.铅锌尾矿重金属(Cd、Cu、Pb和Zn)含量均值分别为19.5、15.9、48.0和1 294.4 mg ·kg-1; 重金属有效态(DTPA-Cd、DTPA-Cu、DTPA-Pb和DTPA-Zn)含量均值分别为0.27、1.08、2.51和57.8 mg ·kg-1.对尾矿重金属总量和重金属有效态含量随物种多样性的增加进行线性回归发现, 随着植物物种多样性的提高, 尾矿重金属总量没有显著性变化(Cd: r=0.02, P=0.87; Cu: r=-0.01, P=0.92; Pb: r=-0.02, P=0.83和Zn: r=-0.12, P=0.22)[图 4 (a)~4(d)], 重金属有效态含量随物种多样性的增加显著下降: DTPA-Cd(r=-0.20, P < 0.05)、DTPA-Cu(r=-0.19, P < 0.05)、DTPA-Pb(r=-0.23, P < 0.05)和DTPA-Zn(r=-0.23, P < 0.05)[图 4 (e)~4(h)].

|
图 4 植物物种多样性对尾矿重金属元素的影响(n=102) Fig. 4 Relationship between species diversity and heavy metal concentrations in the mine tailings (n=102) |
一个成功的植被恢复可以稳定土壤、控制污染、改善景观和减轻污染对人类的健康威胁[5, 30].近年来, 生物多样性与生态系统功能的关系成为当前生态学领域内的一个重大科学问题[7, 8].本研究在重金属尾矿废弃地上建立了不同物种多样性的植被群落(1-物种、4-物种、8-物种和16-物种), 物种多样性的增加显著提高了植物群落的植被盖度和生产力(图 1), 初步确定了植物物种多样性对尾矿生态恢复的促进作用.这与前人研究结果一致, 多样性导致更高的群落生产力、更高的系统稳定性和更高的抗入侵能力[31~33].对植物生产力的多样性效应分析表明, 4-物种、8-物种和16-物种净效应没有显著差异(图 5).有研究表明, 植物物种多样性的增加会通过“选择效应”(随着物种数增加, 随机带入高产物种的可能性随之增加, 最终使整个群落的均产值提高)或“互补效应”(群落中各物种通过互补利用资源, 或者种间促进作用来提高产值)提高生态系统生产力[14, 34, 35].对净效应分割发现, 16-物种的选择效应显著高于4-物种和8-物种; 16-物种的互补效应高于4-物种和8-物种, 但在数据统计上差异不显著(图 5).其原因可能与本研究的调查时间区段有关, 植被调查时间仅为植被建立后的两年, 属群落建立初期, 在尾矿如此恶劣的环境基质中, 耐性较强的物种, 如禾本科物种(特别是芒草、狼尾草、狗牙根)、苎麻、苍耳和黄花蒿等在相对较短的时间内发芽、生长和定居, 故包含有这些物种的组合植被盖度较高, 生物量也较大.而非耐性物种, 如金合欢、紫穗槐、画眉草和斑花败酱等发芽率较低, 在恶劣的环境和竞争压力下处弱势地位, 生产力极其低下.有研究表明, 群落建立初期, 多样性作用机制主要是抽样效应, 但是生态位互补的作用会随时间推移而加强, 并逐步成为主要的多样性作用机制[17, 36, 37].

|
同一组中不同小写字母表示各处理间差异显著P < 0.05 图 5 物种多样性对尾矿生态恢复过程中植被群落的多样性效应 Fig. 5 Effect of species diversity on vegetation community in ecological restoration of mine tailings |
植被恢复是控制尾矿中重金属毒性及其传播的有效途径之一, 植被恢复的目的不是清除土壤中的重金属, 而是通过植物根际作用将重金属离子固定下来, 以降低其生物有效性和移动性, 并防止其进入地下水和食物链, 减少对环境和人类健康的危害[38, 39].本研究表明, 随着植物物种多样性的增加, 尾矿重金属总量没有显著性变化[图 4(a)~4(d)], 重金属有效态含量随物种多样性的增加呈显著下降的趋势[图 4(e)~4(h)]. Bes等[40]的研究表明, 根系分泌物和土壤微生物活动可通过氧化还原反应、络合/螯合作用有效固定重金属离子.此外, 重金属在土壤-植物系统中的迁移和累积一直是矿业废弃地植被重建过程中人们非常关心的问题.从植物稳定的角度来看, 植物应转移尽可能少的重金属到地上部分, 以防止重金属进入食物链[38, 39].本研究发现, 随着植物物种多样性的提高, 植物地上部分重金属Cd、Cu、Pb和Zn的含量均值是逐渐下降的(图 2).这与尾矿中重金属有效态含量随物种多样性的增加而下降有关[图 4 (e)~4(h)].这种相关关系通过植被盖度、生物量与尾矿中重金属总量及有效态含量之间的负相关关系得到进一步的验证(表 2).保险假说认为, 在恶劣环境中物种间生态位差异可以使不同物种“风险分摊”, 物种丰富的系统会表现出“缓冲效应”[17, 41].可以推测, 本试验中多物种处理植物间可能存在互补效应, 物种之间的正相互关系在一定程度上分摊了重金属带来的不利影响, 毒害作用得以被“缓冲”.同时, 物种多样性的增加显著降低了植物对重金属的累积能力, 这将有利于降低由于动物取食以及枯枝落叶带来的生态风险.
|
|
表 2 植被参数与尾矿理化性质Pearson相关性分析1)(n=102) Table 2 Pearson's correlation coefficients between vegetation parameters and physico-chemical properties of mine tailings (n=102) |
3.3 物种多样性对尾矿生态恢复中营养元素累积的生态效应
土壤熟化过程在很大程度上依赖于生长在其上的植被.有研究表明, 植物在促进营养元素的累积、有机物质的分解及接下来的土壤形成和发展过程起着非常重要的作用[42, 43].尽管本试验植被建立的时间相对较短, 只有两年的时间, 但随着植物物种多样性的提高, 总有机碳(TOC)、总氮(TN)、总磷(TP)和水溶性有机碳(SOC)累积量显著性增加[图 3(a)、3(b)、3(c)和3(e)], 这与前人的研究结果一致[44, 45].Yang等[44]在广东大宝山多金属矿排土场建立了Cynodon dactylon (Plot Ⅰ)、Pennisetum purpureum (Plot Ⅱ)、Eucalyptus robusta (Plot Ⅲ)、Neyraudia reynaudiana+Panicum repens (Plot Ⅳ)和E. robusta+N. reynaudiana +C. dactylon (Plot Ⅴ) 这5种不同植被恢复试验小区, 对不同植物物种及组合的生态效应分析表明, 物种多样性最高的Plot V显示最高的有机质、总氮、铵态氮、有效磷和有效钾累积率. Pérez-de-Mora等[45]在西班牙Aznalcóllar矿区建立新的植物群落, 对植被建立后4~6 a植被发展情况进行了跟踪调查, 结果发现植物物种数从33种增加到40种, 植被盖度从40%提高到80%, 部分区域达到接近100%, 伴随着物种数与盖度的增加土壤有机碳积累量显著提高.本研究中, 对植被盖度、生物量与尾矿营养元素指标进行相关性分析表明, 植被盖度和生物量与尾矿有机碳、总氮、总磷和有效磷呈显著正相关(表 2).这也进一步说明, 尾矿土壤营养状况的改善会促进植物的生长, 接下来的养分累积量也会逐渐增加, 尾矿生态系统会进入一个养分逐渐累积的良性循环.
4 结论植物物种多样性显著提高了尾矿新建植物群落的盖度和生产力, 促进了尾矿土壤营养元素的累积, 减少了尾矿重金属有效态含量及重金属在尾矿土壤-植物系统中的迁移和累积, 说明植物物种多样性对尾矿生态恢复具有促进作用.对植物生产力的多样性效应分析表明, 4-物种、8-物种和16-物种净效应没有显著差异, 但16-物种的选择效应显著高于4-物种和8-物种, 其主要作用机制可能是随着植物物种多样性的增加, 随机带入高产物种的可能性随之增加, 最终使整个植物群落的生产力产值提高.对植物参数与尾矿理化性质相关分析表明, 植被盖度、生物量与尾矿有机碳、总氮、总磷和有效磷呈显著正相关, 其主要作用机制可能是植物物种间的正相关关系促进了尾矿营养元素的累积以及接下来的土壤形成、发展和熟化过程.对重金属在土壤-植物系统中的迁移和累积分析表明, 物种多样性加快了重金属离子的固定, 物种之间的相互关系在一定程度上分摊了重金属带来的不利影响, 其主要作用机制可能来自两个方面: 一方面植物物种多样性增加了根系分泌物的数量和种类, 加快了重金属离子的固定; 另一方面多物种间存在互补效应, 在一定程度上分摊了重金属带来的不利影响, 毒害作用得以被“缓冲”.
| [1] | Karaca O, Cameselle C, Reddy K R. Mine tailing disposal sites: contamination problems, remedial options and phytocaps for sustainable remediation[J]. Reviews in Environmental Science and Bio/Technology, 2018, 17: 205-228. DOI:10.1007/s11157-017-9453-y |
| [2] | Liu K H, Zhang H C, Liu Y F, et al. Investigation of plant species and their heavy metal accumulation in manganese mine tailings in Pingle Mn mine, China[J]. Environmental Science and Pollution Research, 2020, 27(16): 19933-19945. DOI:10.1007/s11356-020-08514-9 |
| [3] | Wang P, Sun Z H, Hu Y N, et al. Leaching of heavy metals from abandoned mine tailings brought by precipitation and the associated environmental impact[J]. Science of the Total Environment, 2019, 695. DOI:10.1016/j.scitotenv.2019.133893 |
| [4] | Luo Z T, Tang C B, Hao Y H, et al. Solidification/stabilization of heavy metals and its efficiency in lead-zinc tailings using different chemical agents[J]. Environmental Technology, 2020. DOI:10.1080/09593330.2020.1845817 |
| [5] | Wang L, Ji B, Hu Y H, et al. A review on in situ phytoremediation of mine tailings[J]. Chemosphere, 2017, 184: 594-600. DOI:10.1016/j.chemosphere.2017.06.025 |
| [6] | Gastauer M, Caldeira C F, Ramos S J, et al. Active rehabilitation of Amazonian sand mines converges soils, plant communities and environmental status to their predisturbance levels[J]. Land Degradation & Development, 2020, 31(5): 607-618. |
| [7] | Midgley G F. Biodiversity and ecosystem function[J]. Science, 2012, 335(6065): 174-175. DOI:10.1126/science.1217245 |
| [8] |
李奇, 朱建华, 肖文发. 生物多样性与生态系统服务——关系、权衡与管理[J]. 生态学报, 2019, 39(8): 2655-2666. Li Q, Zhu J H, Xiao W F. Relationships and trade-offs between, and management of biodiversity and ecosystem services[J]. Acta Ecologica Sinica, 2019, 39(8): 2655-2666. |
| [9] | Odenbaugh J. Biodiversity, ecosystem functioning, and the environmentalist agenda[J]. Biology & Philosophy, 2020, 35. DOI:10.1007/s10539-019-9723-x |
| [10] | Yang X, Tan J Q, Sun K H, et al. Experimental demonstration of the importance of keystone communities for maintaining metacommunity biodiversity and ecosystem functioning[J]. Oecologia, 2020, 193(2): 437-447. DOI:10.1007/s00442-020-04693-x |
| [11] | Tilman D, Downing J A. Biodiversity and stability in grasslands[J]. Nature, 1994, 367(6461): 363-365. DOI:10.1038/367363a0 |
| [12] | Tilman D, Reich P B, Knops J, et al. Diversity and productivity in a long-term grassland experiment[J]. Science, 2001, 294(5543): 843-845. DOI:10.1126/science.1060391 |
| [13] | Tilman D, Reich P B, Knops J M H. Biodiversity and ecosystem stability in a decade-long grassland experiment[J]. Nature, 2006, 441(7039): 629-632. |
| [14] | Li J T, Duan H N, Li S P, et al. Cadmium pollution triggers a positive biodiversity-productivity relationship: evidence from a laboratory microcosm experiment[J]. Journal of Applied Ecology, 2010, 47(4): 890-898. DOI:10.1111/j.1365-2664.2010.01818.x |
| [15] | Li S P, Li J T, Kuang J L, et al. Effects of species richness on cadmium removal efficiencies of algal microcosms[J]. Journal of Applied Ecology, 2012, 49(1): 261-267. DOI:10.1111/j.1365-2664.2011.02091.x |
| [16] |
马頔. 镉胁迫对藻类生物多样性影响的微宇宙实验研究[D]. 西安: 西北大学, 2018. Ma D. The relationship between algae biodiversity and biomass under different cadmium stress: a microcosm experiment[D]. Xi'an: Northwest University, 2018. |
| [17] |
杨扬, 王晓燕, 王江, 等. 物种多样性对植物生长与土壤镉污染修复的影响[J]. 环境科学学报, 2016, 36(6): 2103-2113. Yang Y, Wang X Y, Wang J, et al. Effects of species diversity on plant growth and remediation of Cd contamination in soil[J]. Acta Scientiae Circumstantiae, 2016, 36(6): 2103-2113. |
| [18] | Bardgett R D, Van Der Putten W H. Belowground biodiversity and ecosystem functioning[J]. Nature, 2014, 515(7528): 505-511. DOI:10.1038/nature13855 |
| [19] | Rohr J R, Bernhardt E S, Cadotte M W, et al. The ecology and economics of restoration: when, what, where, and how to restore ecosystems[J]. Ecology and Society, 2018, 23. DOI:10.5751/ES-09876-230215 |
| [20] | Carpio I E M, Ansari A, Rodrigues D F. Relationship of biodiversity with heavy metal tolerance and sorption capacity: a meta-analysis approach[J]. Environmental Science & Technology, 2017, 52(1): 184-194. |
| [21] |
陈明辉, 孙际茂, 付益平, 等. 湘西州矿产资源现状及找矿方向[J]. 矿产与地质, 2008, 22(2): 93-96. Chen M H, Sun J M, Fu Y P, et al. Current status of the mineral resouces in Xiangxi Autonomous Prefecture and the prospecting direction in this area[J]. Mineral Resources and Geology, 2008, 22(2): 93-96. DOI:10.3969/j.issn.1001-5663.2008.02.001 |
| [22] | Nelson D W, Sommers L E. Total carbon, organic carbon, and organic matter[A]. In: Page A L (Ed. ). Methods of Soil Analysis[M]. Madison: American Society of Agronomy and Soil Science Society of America, 1982. |
| [23] | Maynard D G, Kalra Y P, Crumbaugh J A. Soil sampling and methods of analysis[M]. Taylor and Francis, Boca Raton: CRC Press, 2008. |
| [24] | Bray R H, Kurtz L T. Determination of total, organic, and available forms of phosphorus in soil[J]. Soil Science, 1945, 59(1): 39-46. DOI:10.1097/00010694-194501000-00006 |
| [25] | 鲁如坤. 土壤农业化学分析方法[M]. 北京: 中国农业科技出版社, 1999. |
| [26] | Allen S E. Chemical analysis of ecological materials[M]. (2nd ed.). Oxford: Blackwell Scientific Publications, 1989. |
| [27] | McGrath S P, Cunliffe C H. A simplified method for the extraction of the metals Fe, Zn, Cu, Ni, Cd, Pb, Cr, Co and Mn from soils and sewage sludges[J]. Journal of the Science of Food and Agriculture, 1985, 36(9): 794-798. DOI:10.1002/jsfa.2740360906 |
| [28] | Lindsay W L, Norvell W A. Development of a DTPA soil test for zinc, iron, manganese, and copper[J]. Soil Science Society of America Journal, 1978, 42(3): 421-428. DOI:10.2136/sssaj1978.03615995004200030009x |
| [29] | Loreau M, Hector A. Partitioning selection and complementarity in biodiversity experiments[J]. Nature, 2001, 412(6842): 72-76. DOI:10.1038/35083573 |
| [30] | Hou X Y, Liu S L, Zhao S, et al. Selection of suitable species as a key factor for vegetation restoration of degraded areas in an open-pit manganese-ore mine in Southern China using multivariate-analysis methods[J]. Land Degradation & Development, 2019, 30(8): 942-950. |
| [31] | Tilman D. Biodiversity: population versus ecosystem stability[J]. Ecology, 1996, 77(2): 350-363. |
| [32] | Weis J J, Cardinale B J, Forshay K J, et al. Effects of species diversity on community biomass production change over the course of succession[J]. Ecology, 2007, 88(4): 929-939. DOI:10.1890/06-0943 |
| [33] | Brassard B W, Chen H Y H, Cavard X, et al. Tree species diversity increases fine root productivity through increased soil volume filling[J]. Journal of Ecology, 2013, 101(1): 210-219. DOI:10.1111/1365-2745.12023 |
| [34] | Cardinale B J, Wright J P, Cadotte M W, et al. Impacts of plant diversity on biomass production increase through time because of species complementarity[J]. Proceedings of the National Academy of Science of the United States of America, 2007, 104(46): 18123-18128. DOI:10.1073/pnas.0709069104 |
| [35] | Gross N, Suding K N, Lavorel S, et al. Complementarity as a mechanism of coexistence between functional groups of grasses[J]. Journal of Ecology, 2007, 95(6): 1296-1305. DOI:10.1111/j.1365-2745.2007.01303.x |
| [36] | Hooper D U, Chapin III F S, Ewel J J, et al. Effects of biodiversity on ecosystem functioning: a consensus of current knowledge[J]. Ecological Monographs, 2005, 75(1): 3-35. DOI:10.1890/04-0922 |
| [37] | Weisser W W, Roscher C, Meyer S T, et al. Biodiversity effects on ecosystem functioning in a 15-year grassland experiment: Patterns, mechanisms, and open questions[J]. Basic and Applied Ecology, 2017, 23: 1-73. DOI:10.1016/j.baae.2017.06.002 |
| [38] | Yang S X, Liao B, Yang Z H, et al. Revegetation of extremely acid mine soils based on aided phytostabilization: a case study from southern China[J]. Science of the Total Environment, 2016, 562: 427-434. DOI:10.1016/j.scitotenv.2016.03.208 |
| [39] | Acosta J A, Abbaspour A, Martínez G R, et al. Phytoremediation of mine tailings with Atriplex halimus and organic/inorganic amendments: a five-year field case study[J]. Chemosphere, 2018, 204: 71-78. DOI:10.1016/j.chemosphere.2018.04.027 |
| [40] | Bes C M, Pardo T, Bernal M P, et al. Assessment of the environmental risks associated with two mine tailing soils from the La Unión-Cartagena (Spain) mining district[J]. Journal of Geochemical Exploration, 2014, 147: 98-106. DOI:10.1016/j.gexplo.2014.05.020 |
| [41] | Loreau M. Biodiversity and ecosystem functioning: recent theoretical advances[J]. Oikos, 2000, 91(1): 3-17. DOI:10.1034/j.1600-0706.2000.910101.x |
| [42] | Mpai T, Jaiswal S K, Dakora F D. Accumulation of phosphorus and carbon and the dependency on biological N2 fixation for nitrogen nutrition in Polhillia, Wiborgia and Wiborgiella species growing in natural stands in cape fynbos, South Africa[J]. Symbiosis, 2020, 81: 65-78. DOI:10.1007/s13199-020-00683-y |
| [43] | Van Bellen S, Shotyk W, Magnan G, et al. Carbon and nitrogen accumulation rates in ombrotrophic peatlands of central and northern Alberta, Canada, during the last millennium[J]. Biogeochemistry, 2020, 151(2-3): 251-272. DOI:10.1007/s10533-020-00724-0 |
| [44] | Yang S X, Liao B, Li J T, et al. Acidification, heavy metal mobility and nutrient accumulation in the soil-plant system of a revegetated acid mine wasteland[J]. Chemosphere, 2010, 80(8): 852-859. DOI:10.1016/j.chemosphere.2010.05.055 |
| [45] | Pérez-de-Mora A, Madejón P, Burgos P, et al. Phytostabilization of semiarid soils residually contaminated with trace elements using by-products: sustainability and risks[J]. Environmental Pollution, 2011, 159(10): 3018-3027. DOI:10.1016/j.envpol.2011.04.015 |