 2021, Vol. 42
2021, Vol. 42

 , TAN Meng-yi , WANG Zi-jun , FU Pei-jiao , LI Jin-qiu , TANG Shui-rong , WU Yan-zheng
, TAN Meng-yi , WANG Zi-jun , FU Pei-jiao , LI Jin-qiu , TANG Shui-rong , WU Yan-zheng , MENG Lei
, MENG Lei
全球气候变化对人类产生的影响已经逐步显现, 随着温室气体(greenhouse gas, GHG)排放量的逐年增加, 全球变暖已经成为当前的事实[1].据IPCC(Intergovernmental Panel on Climate Change)统计, 与第一次工业革命时期相比, 目前全球平均气温已升高0.61℃, 若不采取措施减少GHG的排放, 预计到本世纪末, 全球平均气温将升高1.1℃[2]. N2O作为重要的温室气体, 其在百年尺度上的全球增温潜势(global warming potential, GWP)是CO2的265倍[2].农田作为N2O产生的重要场所, 每年产生的N2O(以N计)达到了3.3Tg, 占全球N2O总排放量的43%以上[3, 4].农业源产生的N2O受多方面因素的影响, 其中主要影响因素包括田间管理模式、土地利用方式和施肥量等[5~7].在热区外水分管理良好的稻田中N2O排放量可能较小, 而海南地区年均蒸发量和降雨量大, 农田水利设施欠缺, 土壤频繁干湿交替过程十分常见, 现有田间试验结果表明干湿交替会改善土壤通气环境, 导致土壤N2O排放的增加[8, 9].但在培养过程中干湿交替影响下的温室气体排放规律尚不明确, 这种干湿交替过程可能会导致土壤N2O排放量的增加.
有研究表明多种土壤改良剂如硝化抑制剂和有机物料等在减少土壤N2O排放方面具有良好的作用[10~12].生物炭作为一种新型土壤改良剂近年来在土壤修复、农田改良和作物增产方面效果显著[13, 14].但由于其制备原料、分解温度和分解时间等诸多因素的差异, 不同生物炭的理化性质如比表面积、含氧官能团、C/N、pH和CEC等差异较大, 其对N2O的减排效果也有一定的差异, 同时研究表明生物炭添加量与N2O排放量相关[15, 16].水分同样是影响生物炭减排效果的重要因素, 不同水分条件下, 生物炭的减排效果存在较大差异[17~19].但目前对于何种生物炭在不同水平添加量下以及何种水分条件下对温室气体的减排效果最佳存在一定争议.多数研究表明, 添加生物炭可以增加土壤C/N, 降低NO3--N浓度, 显著降低稻田土壤反硝化过程的N2O排放[20, 21], 相反的观点认为添加尿素和生物炭会减少土壤中矿质氮的淋溶, 导致水稻土产生较高的N2O排放量[22].因此, 在干湿交替频繁的水稻土中添加C/N较高的玉米秸秆生物炭可能会减少N2O的排放.同时, 当前多数水稻土中添加生物炭的培养试验均在固水分条件下进行, 无法较好地模拟田间动态水分变化下生物炭对N2O的减排效果, 进行更为合理的动态水分管理可以更好地模拟热带地区水稻土频繁干湿交替条件下添加生物炭的N2O排放规律.
海南岛地处热带北缘地区, 水稻土成土母质主要以花岗岩、玄武岩、砂页岩、滨海沉积物以及河流沉积物为主.其中土壤有机质含量多在2%左右.由于脱硅富铝作用, 其土壤pH多在7以下, 呈酸性或弱酸性.同时海南地区雨热同期, 土壤矿化过程迅速, 水稻土CEC较低, 保肥能力差, 但雨热条件丰富为其独特的水稻-冬季瓜菜轮作模式提供了气候基础[23, 24].水稻种植使田面淹水, 形成强还原条件, 从而减少冬季瓜菜种植过程中病虫害的发生[25].据统计, 2018年海南水稻种植面积达到了24.6万hm2, 是海南主要粮食作物[26], 由于水利设施的落后, 同时在水稻种植过程中管理较为粗犷, 集雨灌溉为主要的灌溉方式, 田面干湿交替过程十分常见[27].同时由于独特的热带季风气候和海相沉积物土壤母质, 雨季旱季差异明显, 土壤保水保肥能力较差, 早稻种植处于旱季, 这种干湿交替现象更为常见.田伟等[23]和胡玉麟等[24]的研究表明, 热区水稻田N2O排放量不可忽视.在水分管理良好的水稻土中N2O产生多以反硝化过程为主, 而琼北地区频繁的干湿交替过程可能会增加土壤矿化速率, 促进硝化作用, 导致N2O排放风险的增加[28, 29].目前, 关于施用生物炭对我国琼北地区稻田频繁干湿交替条件下温室气体排放的研究鲜见报道.
因此, 本文假设频繁的干湿交替条件会导致N2O排放量的增加, 添加玉米秸秆生物炭会减少N2O的排放, 并采用室内培养试验的方法, 以琼北地区海相沉积物发育而来的水稻土作为研究对象, 模拟海南稻田水分管理模式, 探究在两种水分管理模式下添加不同量的生物炭对土壤N2O排放及理化性质和相关功能基因产生的影响, 以期为生物炭对我国琼北地区稻田温室气体减排提供理论基础.
1 材料与方法 1.1 供试土壤基本性质供试土壤取自海南省澄迈县西岸村(19°55′N, 109°57′E)的稻菜轮作田地水稻种植季土壤, 当地属于热带季风气候, 雨热条件丰富, 年均降雨量1 786.1 mm, 年均气温23.8℃, 土壤类型为海相沉积物发育而来的水稻土.土壤经室内风干后去除石砾和植物根系并过2 mm筛.土壤基本理化性质见表 1.
|
|
表 1 土壤背景值 Table 1 Background value of soils |
1.2 供试生物炭基本性质
供试生物炭为玉米秸秆在400℃厌氧条件下裂解24 h形成, 由中国科学院南京土壤研究所提供, 具体理化性质见表 2.
|
|
表 2 生物炭理化性质 Table 2 Physical and chemical properties of biochar |
1.3 试验设计
本试验采用室内恒温培养法, 共设置6个处理, 每个处理设置6个重复, 其中3个重复用于采集N2O气体, 另外3个重复用于土壤理化和微生物样品采集.处理分别为:①不加生物炭并进行干湿交替处理(AWD1); ②添加2%的生物炭(体积比, 下同)并进行干湿交替处理(AWD2); ③添加4%的生物炭并进行干湿交替处理(AWD3); ④不加生物炭并持续淹水处理(CF1); ⑤添加2%的生物炭并持续淹水处理(CF2); ⑥添加4%的生物炭并持续淹水(CF3)处理.持续淹水处理的土壤充水孔隙度(water filled pore space, WFPS)维持在150%左右, 淹水深度2~3 cm左右, 用称重法对持续淹水处理每天补水, 而干湿交替处理同样从初始的150% WFPS放入培养箱内自然风干6 d后至约40% WFPS后补水至150% WFPS, 整个过程循环5次.试验开始前一周称取200 g风干土壤与各处理定量的玉米秸秆生物炭混匀放入760 mL培养瓶中并加水至60% WFPS进行活化, 通过称重法每天补水.活化完成后, 向每个处理中加入N 200 mg ·kg-1进行为期30 d的25℃恒温避光培养, 其中N由尿素提供(含N 46%).培养过程中添加生物炭的处理在瓶内壁外围存在微量生物炭漂浮.
1.4 样品采集与测定气体样品采集频率为2d ·次-1, 气体排放峰值期1 d ·次-1, 采气时间为当天08:00~09:00, 用10 mL针筒在培养瓶盖紧后的第0和30 min采集瓶中气体, 并收集6 mL气体于带三通阀的针筒中, 当日分析.收集的N2O气体采用气相色谱仪(岛津GC- 2014)进行分析.检测所用标准气体由中国计量科学研究院提供.
土壤pH采用pH计(雷磁PHS- 3C)进行测定, Eh采用便携式Eh计(Bante 220)进行测定.土壤样品中的NH4+-N和NO3--N分别用靛蓝比色法(625 nm)和紫外双波长比色法(220 nm和275 nm)进行测定.
采集培养18 d土壤样品用于微生物分析.土壤样品DNA的提取采用MOBIL公司的土壤快速提取试剂盒(PowerSoilTM DNA Isolation Kit, Mo Bio Laboratories Inc., CA)进行提取, 提取步骤参考试剂盒说明书, 同时用超微量紫外分光光度计(Nano Drop 2000)测定DNA浓度和质量.采用1%的琼脂糖凝胶电泳检测提取出的DNA片段大小, DNA样品存放于-20℃保存待用.
实时荧光定量PCR分析:本试验采用土壤中氨氧化细菌amoA基因、亚硝酸还原酶nirK、nirS基因和一氧化氮还原酶nosZ基因的拷贝数分别表示氨氧化细菌(AOB)、nirK、nirS和nosZ基因型反硝化菌, 所用引物分别为AOB-1F/AOB-2R、nirK-1F/nirK-5R、nirS-cd3aF/nirS-R3cd和nosZ-F/nosZ- 1662R[30].定量分析采用SYBR GREEN法, 反应体系为20 μL, 其中包括10 μL 2×T5 Fast qPCR Mix(SYBR Green I), 10 μmol ·L-1正反向引物各1 μL, 1 μL DNA模板, 7.4 μL无菌水.并使用10-1~10-8浓度梯度的标准质粒作为模板进行荧光定量PCR扩增, 最后根据扩增曲线计算基因丰度.
1.5 数据处理与分析N2O排放速率计算公式如下:
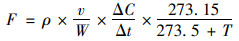
|
式中, F为N2O排放通量[μg ·(kg ·h)-1]; ρ为标准状态下的气体密度(kg ·m-3); v为培养瓶内气体体积(mL); W为风干土质量(kg); ΔC/Δt为采气过程瓶内气体摩尔分数变化速率; T为采样时瓶内平均温度(℃).
累积排放量计算公式如下:
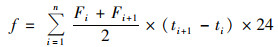
|
式中, f为累计排放量(mg ·kg-1), n和i为采样次数, ti+1-ti为两次采样间隔天数(d).
采用Microsoft Excel 2016对数据进行初步统计整理, 采用SPSS 25.0对数据进行方差分析与相关性分析, 采用Origin 2018进行图形制作.
2 结果与分析 2.1 不同处理N2O排放通量及总量各处理N2O排放高峰均出现在培养9 d后.其中, AWD条件下各处理N2O排放峰值出现在复水落干过程中, 排放最高峰出现在16~20 d[图 1(a)]. CF条件下各处理N2O排放峰值出现时间与AWD条件下各处理相近, 但除峰值期外, CF条件下各处理排放通量均高于AWD处理[图 1(b)].从培养30 d的N2O排放总量看, 各处理N2O排放总量由大到小依次为:CF1>CF2>CF3>AWD1>AWD2>AWD3, AWD各处理的N2O排放总量均显著低于CF各处理(P < 0.05, 下同), 其中CF1处理排放总量最高为2.88mg ·kg-1, 而AWD3处理排放总量最低为0.43mg ·kg-1(图 2).

|
↓表示当日复水至150% WFPS 图 1 不同处理N2O排放通量变化 Fig. 1 Variation in N2O emissions in different treatments |

|
Δ表示各处理与AWD1处理N2O排放总量的比较; 虚线表示以AWD1处理N2O排放总量作为参考物设置的参考线; 不同小写字母表示不同处理间差异显著(P < 0.05), 下同 图 2 不同处理N2O排放总量 Fig. 2 Cumulative emissions of N2O in different treatments |
添加N源后, 土壤pH相对培养前均升高1个单位以上.其中, 在培养第12 d, 添加4%生物炭的AWD3处理pH最高, 达到6.98.除CF1处理外, 所有处理在培养0~6 d, pH均呈上升趋势. 12~30 d, 所有处理pH均呈下降趋势.培养30 d后, 在相同生物炭添加量下, AWD处理的土壤pH显著高于CF处理, 其中AWD2和AWD3处理的土壤pH相比培养前分别提高了0.27和0.47, 而未添加生物炭的淹水处理CF1在培养结束后pH最低, 为5.36(图 3).

|
图 3 不同处理pH变化 Fig. 3 Variation in pH in different treatments |
培养过程中, AWD处理与CF处理的Eh变化趋势相反. 0~18 d, AWD处理的Eh呈先快后慢上升趋势, 在18 d后, Eh趋于稳定.其中AWD1处理的Eh显著高于AWD2和AWD3处理, 达到308 mV.而在CF处理中, 0~18 d, Eh均呈下降趋势, 随后趋于稳定, 最终不添加生物炭的CF1处理其Eh显著高于添加生物炭的CF2和CF3处理. 30 d后, CF3处理Eh显著低于其他处理, 达到-125 mV(图 4).

|
图 4 不同处理Eh变化 Fig. 4 Variation in Eh in different treatments |
培养过程中, 土壤NH4+-N与NO3--N呈现此消彼长的变化趋势.整个培养期内, NH4+-N均呈下降趋势, 加入氮源后第6 d, 各处理NH4+-N含量均在190mg ·kg-1左右, 并没有表现出显著差异.其中AWD3处理在整个培养过程中NH4+-N均高于其它各处理.培养30 d后, 同等生物炭添加量下, AWD各处理NH4+-N含量均显著高于CF各处理[图 5(a)].土壤NO3--N在培养初期含量趋近于0, 6~25 d内各处理NO3--N含量不断升高, 并在25 d时达到最大, 其中CF1和CF2处理的NO3--N要显著高于其他处理. 25~30 d, 各处理NO3--N均呈现下降趋势, 最终CF1和CF2处理的NO3--N含量显著高于其他各处理[图 5(b)].

|
图 5 不同处理NH4+-N和NO3--N变化 Fig. 5 Variation in NH4+-N and NO3--N in different treatments |
不同处理AOB在培养第18 d表现出明显的差异, 其中AWD1和CF1处理具有较高的AOB-amoA基因丰度, 分别为2.28×105 copies ·g-1和2.04×105copies ·g-1, 在同等生物炭添加量下, CF处理的AOB-amoA基因丰度显著高于AWD处理[图 6(a)]. CF3与AWD2处理的nirS基因丰度显著高于其他处理[图 6(b)].在AWD各处理中, 添加生物炭可以显著降低nirK基因丰度[图 6(c)].在相同的水分条件下添加生物炭显著提高土壤nosZ基因丰度, 同时AWD1和AWD2处理的nosZ基因丰度均显著高于CF1和CF2处理[图 6(d)].

|
图 6 不同处理第18 d微生物功能基因丰度 Fig. 6 Microbial functional gene abundance on the 18th day of different treatments |
通过分析(nirK+nirS)/nosZ, 同等生物炭添加量下AWD各处理在第18 d时(nirK+nirS)/nosZ比值显著低于CF各处理, 同等水分条件下, 生物炭添加量与(nirK+nirS)/nosZ成反比(图 7).

|
图 7 不同处理第18 d(nirK+nirS)/nosZ比值 Fig. 7 Value of (nirK+nirS)/nosZ on the 18th day of different treatments |
本试验中N2O排放通量与pH和Eh呈极显著负相关, 但与NO3-呈显著正相关, 与NH4+未表现出显著相关性.同时N2O排放通量和AOB呈极显著正相关, 相关系数达到0.507, 与nirK、nirS和(nirK+nirS)/nosZ呈显著正相关, 其中相关性表现为nirK>(nirK+nirS)/nosZ>nirS, 但与nosZ无显著相关性.AOB与土壤pH呈极显著负相关, 与Eh和NH4+呈显著负相关, 与NO3-呈极显著正相关, 相关性系数达到0.682.nosZ基因与NO3-呈显著负相关, 与nirS基因呈极显著正相关(表 3).
|
|
表 3 N2O排放通量与各因素间的Pearson相关系数1) Table 3 Pearson correlation coefficients between N2O fluxes and various factors |
3 讨论 3.1 水分变化和添加生物炭对N2O排放的影响
一般认为, 水分是调节土壤N2O排放的关键, 本研究假设干湿交替会增加水稻土N2O的排放.在有氧条件下, 硝化过程是产生N2O的主要过程, 厌氧条件下反硝化过程主导N2O排放, 干湿交替条件有利于硝化过程进而促进N2O的排放[31, 32].Liu等[18]的研究发现土壤硝化过程产生的N2O排放速率在50% ~85% WFPS下最高.Ma等[19]的研究发现100% WHC时土壤N2O累积排放量最高, 而40% WHC时微生物活性降低, N2O排放受到抑制.本试验结果表明, 在不添加生物炭的条件下, 相对于持续淹水, 干湿交替条件可以提高18 d时N2O排放通量, 这是由于在干湿交替过程中, 土壤WFPS可以降低至最适宜N2O产生的区间, 这段时期会产生较高的N2O排放, 这与前人研究结果相似(图 1)[18, 19].但前18 d干湿交替处理排放通量低于持续淹水处理, 同时干湿交替处理的N2O排放总量均小于CF各处理, 这与假设不符合.主要由于整个过程中, 干湿交替过程土壤处于最适WFPS的时间较短, WFPS频繁变化过程中过高或过低均会限制微生物硝化与反硝化过程的进行, 从而抑制了N2O的产生[33, 34].持续淹水处理在前中期土壤孔隙中充足的O2导致硝化作用与反硝化作用并存, 中后期由于缺少植物对N的吸收及NO3--N无法淋溶, NO3--N在瓶中大量累积, 反硝化过程主导N2O排放, 导致其N2O减排效果减弱[19].Lin等[35]在淹水状态下培养水稻土也有较高N2O排放.Anderson等[36]对不同WFPS土壤进行培养发现60% WFPS会促进硝化作用的进行, 而100% WFPS下反硝化作用会得到促进, 同时N2O排放通量最大.Setyanto等[37]在印尼水稻土上进行了与本研究类似的干湿交替试验但并未添加生物炭, 结果表明较持续淹水处理相比, 干湿交替处理并不会增加N2O排放.
前人研究表明, 多数条件下生物炭可以有效减少土壤N2O的排放, 本试验结果与假设一致[38].添加生物炭后, 土壤N2O排放总量显著降低, 可能的原因是生物炭可以抑制土壤硝化过程[39], 同时促进反硝化过程中N2O向N2的还原[40].在干湿交替条件下, 土壤处于好氧-厌氧交替变化过程, N2O主要产生于硝化过程, 其累积排放量与生物炭添加量呈反比(图 2), 这可能是由于生物炭在干湿交替条件下增加了土壤孔隙度, 提升土壤对N2O的截持能力[41, 42], 生物炭较高的CEC提升了土壤对NH4+-N的吸附能力, 降低了硝化过程可利用的底物浓度, 减少硝化过程N2O的排放[43], 同时, 干湿交替处理Eh均处于较高水平(图 4), 限制了反硝化产生N2O过程.相关性分析结果表明N2O排放与AOB丰度呈显著正相关, 添加生物炭可以显著降低AOB-amoA与nirK基因丰度[图 6(a)和6(b)], 限制其硝化过程与反硝化过程, 从而导致N2O排放降低.Wang等[44]和Clough等[45]的研究发现生物炭中的酚类化合物和α-蒎烯化合物抑制了硝化菌的活性, 降低了硝化作用强度.而在持续淹水过程中, 培养前中期土壤中存在充气孔隙, 硝化与反硝化并存, 中后期土壤处于还原条件, 反硝化过程主导了N2O的排放.持续淹水处理添加生物炭后提高了土壤孔隙度, 导致在培养前中期CF2和CF3硝化与反硝化作用并存, 这是导致其减排效果比添加生物炭的AWD2和AWD3弱的原因. CF1处理中AOB-amoA基因丰度处于较高水平, 但显著低于AWD1处理, 而CF2和CF3处理的AOB-amoA基因丰度高于AWD2和AWD3处理, 同时持续淹水处理(nirK+nirS)/nosZ高于干湿交替处理, 这同样说明在前中期持续淹水处理中硝化与反硝化过程并存(图 6和图 7).但生物炭提高了土壤pH和C/N, 促进反硝化过程N2O向N2的还原[46], 因此从累积排放量看, 在持续淹水处理中同样具有减排效果.Harter等[41]和Van Zwieten等[42]分别向砂质土壤和红壤中添加生物炭发现, 生物炭可以增加土壤孔隙度, 这些孔隙对N2O具有截留作用从而减少N2O的排放, 同时, 由于土壤孔隙度的增加产生的有氧条件抑制了厌氧反硝化的进行, 减少了反硝化过程N2O的产生.Tarin等[47]通过在土壤中添加8%木质生物炭同样发现了其有良好的减排效果, 减少了11%的N2O排放.相反的观点认为添加生物炭会促进N2O的排放, Lin等[35]的研究表明添加小麦秸秆生物炭会显著增加土壤N2O排放总量, 这可能是由于土壤类型及生物炭种类差异所导致的.
3.2 水分变化和添加生物炭对土壤性质的影响有研究表明, 添加生物炭可以显著提高土壤pH, Chintala等[48]的研究发现在酸性土壤中添加玉米秸秆和柳枝生物炭均可显著提高土壤pH, 这与本研究的结果相似.培养前期, 土壤pH缓慢上升, 12 d后土壤pH缓慢下降, 最终, 相同水分管理条件下添加生物炭处理的土壤pH显著高于不添加生物炭的处理(图 3), 这可能是由于前期尿素水解和生物炭的共同作用, 导致土壤pH升高, 在后期由于NO3--N浓度的增加, 土壤pH降低[35], 同时, 干湿交替过程削弱了硝化过程, 导致其NO3--N浓度低于CF处理, 产生的NO3--N相对较少, 从而导致AWD各处理在同等生物炭添加量下的pH高于CF各处理[45].
土壤Eh的变化主要与土壤充水孔隙有关, 一般在淹水条件下, 土壤呈还原条件[49, 50].本试验结果与前人的研究一致, 同时笔者发现, 在培养30 d后同等水分管理条件下, 添加生物炭可以显著降低土壤Eh(图 4), 这可能是因为生物炭增加了土壤还原过程中微生物对电子的利用效率, 从而导致土壤Eh降低[51].
NH4+-N和NO3--N在培养过程中呈此消彼长的关系(图 5).在同等水分条件下添加生物炭处理在培养结束后NH4+-N浓度显著高于不添加生物炭的处理, 这与Yang等[52]的研究结果一致, 生物炭可以有效吸附土壤中的NH4+-N, 使土壤硝化过程底物浓度降低.同时AWD各处理NH4+-N浓度均高于同等生物炭添加量下的CF各处理, 这主要是由于干湿交替削弱了硝化过程, 同时生物炭对NH4+-N吸附和抑制AOB的硝化过程共同作用的结果.Zheng等[53]的研究发现添加生物炭可以有效抑制土壤NH4+-N向NO3--N的转变和淋失.Wang等[44]的研究表明生物炭中的酚类化合物可以抑制AOB的活性从而抑制硝化过程.而NO3--N在培养前期逐渐升高则是由于硝化作用导致NO3--N的积累, 后期硝化作用减弱, 反硝化作用消耗NO3--N导致其浓度下降.
3.3 水分变化和添加生物炭对微生物功能基因的影响有研究表明, AOB主导土壤硝化过程中N2O的排放.Wang等[44]的研究表明生物炭中的酚类化合物可以抑制AOB的活性.陈晨等[54]通过在蔬菜地添加尿素和生物炭发现相比只加尿素处理, AOB丰度减少了105.8%.通过第18 d的微生物功能基因丰度发现, 生物炭可以显著降低酸性土壤AOB丰度, 在AWD条件下, AWD3处理相比AWD1处理AOB丰度降低了92.5%, CF3处理与CF1处理相比AOB丰度降低了79.0%[图 6(a)].AWD3处理AOB丰度最低, 可能的原因是较持续淹水处理相比, 干湿交替过程中微生物群落结构变化较大.Wu等[55]的研究发现土壤干湿交替过程中微生物量减少了约44%.Lin等[35]的研究认为生物炭可以提高AOB丰度, 促进N2O排放, 这与本研究结果存在差异, 导致差异存在可能的原因是添加生物炭种类的不同和土壤类型的不同导致施入氮源后土壤pH的差异.
反硝化过程同样是N2O产生的重要途径, 主导反硝化过程的基因包括nirK、nirS和nosZ[56, 57].相关性分析结果表明nirK和nirS基因丰度与N2O排放通量呈显著正相关, 这与前人的研究结果一致(表 3).Yanai等[58]的研究表明添加生物炭可以利用其石灰效应提升土壤pH, 提高土壤N2O还原酶活性, 本研究结果与之相符, 在18 d时, 干湿交替条件下生物炭可以显著降低反硝化过程底物浓度, 进而导致nirK基因丰度的降低, 同等水分条件下添加生物炭处理nosZ基因丰度显著高于未添加生物炭处理, 而nirS基因丰度未表现出明显的规律, 但添加生物炭可以显著降低(nirK+nirS)/nosZ(图 7).Li等[59]的研究发现, 由于生物炭较大的比表面积和较多的纳米孔隙, 可以增加土壤孔隙度抑制反硝化过程, 进而显著减少反硝化过程中nirK的基因丰度.Wu等[60]的研究表明在水稻田中添加生物炭可以显著降低反硝化酶活性, 同时增加了N2O还原酶活性, 使nosZ基因丰度增加, 进而显著降低(nirK+nirS)/nosZ, 减少反硝化过程N2O的排放.
4 结论(1) 不同水分条件下添加生物炭均可减少N2O的排放, 在干湿交替条件下生物炭对N2O的减排效果更佳, 说明生物炭在琼北地区水稻田N2O减排方面具有较大的利用价值.
(2) 在琼北地区酸性水稻土中添加玉米秸秆生物炭可以有效提高土壤pH, 改良土壤酸化状况, 同时吸附土壤NH4+-N, 抑制土壤硝化作用.
(3) 添加生物炭可以显著降低土壤AOB丰度, 抑制硝化作用下N2O的产生, 同时增加nosZ基因丰度, 降低(nirK+nirS)/nosZ, 促进反硝化过程N2O的还原, 进而减少N2O排放.
| [1] | Agliardi E, Alexopoulos T, Cech C. On the relationship between GHGs and global temperature anomalies: Multi-level rolling analysis and copula calibration[J]. Environmental and Resource Economics, 2019, 72(1): 109-133. DOI:10.1007/s10640-018-0259-3 |
| [2] | IPCC. Climate Change 2014:Mitigation of climate change. Contribution of working group Ⅲ to the fifth assessment report of the intergovernmental panel on climate change[M]. Cambridge: Cambridge University Press, 2014. |
| [3] |
聂皇华, 张家升, 和周明, 等. 长期定位施用牛粪对夏玉米-冬小麦体系农田N2O和NO排放的影响[J]. 环境科学, 2019, 40(2): 885-892. Nie H H, Zhang J S, He Z M, et al. Effect of long-term dairy manure amendment on N2O and NO emissions from summer maize-winter wheat cropping systems[J]. Environmental Science, 2019, 40(2): 885-892. |
| [4] | Carlson K M, Gerber J S, Mueller N D, et al. Greenhouse gas emissions intensity of global croplands[J]. Nature Climate Change, 2017, 7(1): 63-68. DOI:10.1038/nclimate3158 |
| [5] | 蔡祖聪, 徐华, 马静. 稻田生态系统CH4和N2O排放[M]. 合肥: 中国科学技术大学出版社, 2009. |
| [6] |
刘若萱, 贺纪正, 张丽梅. 稻田土壤不同水分条件下硝化/反硝化作用及其功能微生物的变化特征[J]. 环境科学, 2014, 35(11): 4275-4283. Liu R X, He J Z, Zhang L M. Response of nitrification/denitrification and their associated microbes to soil moisture change in paddy soil[J]. Environmental Science, 2014, 35(11): 4275-4283. |
| [7] |
伍延正, 张苗苗, 秦红灵, 等. 不同土地利用方式下冬季N2O排放及其影响因素[J]. 环境科学, 2013, 34(8): 2968-2974. Wu Y Z, Zhang M M, Qin H L, et al. N2O flux in winter and its affecting factors under different land use patterns[J]. Environmental Science, 2013, 34(8): 2968-2974. |
| [8] |
董艳芳, 黄景, 李伏生, 等. 不同灌溉模式和施氮处理下稻田CH4和N2O排放[J]. 植物营养与肥料学报, 2017, 23(3): 578-588. Dong Y F, Huang J, Li F S, et al. Emissions of CH4 and N2O under different irrigation methods and nitrogen treatments[J]. Journal of Plant Nutrition and Fertilizer, 2017, 23(3): 578-588. |
| [9] | Li L B, Li F S, Dong Y F. Greenhouse gas emissions and global warming potential in double-cropping rice fields as influenced by two water-saving irrigation modes in South China[J]. Journal of Soil Science and Plant Nutrition, 2020, 20(4): 2617-2630. DOI:10.1007/s42729-020-00328-5 |
| [10] | Du Y G, Xin Y, Shu K, et al. Effect of nitrification inhibitor on plant biomass and N2O emission rates in alpine meadows on the Tibetan Plateau[J]. Chemistry and Ecology, 2020, 36(5): 410-418. DOI:10.1080/02757540.2020.1745782 |
| [11] |
陈友德, 赵杨, 高杜娟, 等. 稻油不同轮作模式对农田甲烷和氧化亚氮排放的影响[J]. 环境科学, 2020, 41(10): 4701-4710. Chen Y D, Zhao Y, Gao D J, et al. Effects of different rotation patterns of oil-rice on methane and nitrous oxide emissions in rice fields[J]. Environmental Science, 2020, 41(10): 4701-4710. |
| [12] |
郝耀旭, 刘继璇, 袁梦轩, 等. 长期定位有机物料还田对关中平原冬小麦-玉米轮作土壤N2O排放的影响[J]. 环境科学, 2017, 38(6): 2586-2593. Hao Y X, Liu J X, Yuan M X, et al. Effects of long-term organic amendments on soil N2O emissions from winter wheat-maize cropping systems in the Guanzhong Plain[J]. Environmental Science, 2017, 38(6): 2586-2593. |
| [13] | Wei W L, Yang H Q, Fan M S, et al. Biochar effects on crop yields and nitrogen loss depending on fertilization[J]. Science of the Total Environment, 2020, 702. DOI:10.1016/j.scitotenv.2019.134423 |
| [14] | Uchimiya M, Lima I M, Klasson K T, et al. Contaminant immobilization and nutrient release by biochar soil amendment: Roles of natural organic matter[J]. Chemosphere, 2010, 80(8): 935-940. DOI:10.1016/j.chemosphere.2010.05.020 |
| [15] | Cayuela M L, Van Zwieten L, Singh B P, et al. Biochar's role in mitigating soil nitrous oxide emissions: a review and meta-analysis[J]. Agriculture, Ecosystems & Environment, 2014, 191: 5-16. |
| [16] | Feng Z J, Zhu L Z. Impact of biochar on soil N2O emissions under different biochar-carbon/fertilizer-nitrogen ratios at a constant moisture condition on a silt loam soil[J]. Science of the Total Environment, 2017, 584-585: 776-782. DOI:10.1016/j.scitotenv.2017.01.115 |
| [17] | Xiang J, Liu D Y, Ding W X, et al. Effects of biochar on nitrous oxide and nitric oxide emissions from paddy field during the wheat growth season[J]. Journal of Cleaner Production, 2015, 104: 52-58. DOI:10.1016/j.jclepro.2014.12.038 |
| [18] | Liu R, Hayden H L, Suter H, et al. The effect of temperature and moisture on the source of N2O and contributions from ammonia oxidizers in an agricultural soil[J]. Biology and Fertility of Soils, 2017, 53(1): 141-152. DOI:10.1007/s00374-016-1167-8 |
| [19] | Ma L, Cheng Y, Wang J Y, et al. Mechanical insights into the effect of fluctuation in soil moisture on nitrous oxide emissions from paddy soil[J]. Paddy and Water Environment, 2017, 15(2): 359-369. DOI:10.1007/s10333-016-0554-y |
| [20] | Singla A, Inubushi K. Effect of biochar on CH4 and N2O emission from soils vegetated with paddy[J]. Paddy and Water Environment, 2014, 12(1): 239-243. DOI:10.1007/s10333-013-0357-3 |
| [21] | Wu D, Feng Y F, Xue L H, et al. Biochar combined with vermicompost increases crop production while reducing ammonia and nitrous oxide emissions from a paddy soil[J]. Pedosphere, 2019, 29(1): 82-94. DOI:10.1016/S1002-0160(18)60050-5 |
| [22] | Chen J J, Kim H, Yoo G. Effects of biochar addition on CO2 and N2O emissions following fertilizer application to a cultivated grassland soil[J]. PLoS One, 2015, 10(5). DOI:10.1371/journal.pone.0126841 |
| [23] |
田伟, 伍延正, 汤水荣, 等. 不同施肥模式对热区晚稻水田CH4和N2O排放的影响[J]. 环境科学, 2019, 40(5): 2426-2434. Tian W, Wu Y Z, Tang S R, et al. Effects of different fertilization modes on greenhouse gas emission characteristics of paddy fields in Hot Areas[J]. Environmental Science, 2019, 40(5): 2426-2434. |
| [24] |
胡玉麟, 汤水荣, 陶凯, 等. 优化施肥模式对我国热带地区水稻-豇豆轮作系统N2O和CH4排放的影响[J]. 环境科学, 2019, 40(11): 5182-5190. Hu Y L, Tang S R, Tao K, et al. Effects of optimizing fertilization on N2O and CH4 emissions in a paddy-cowpea rotation system in the tropical region of China[J]. Environmental Science, 2019, 40(11): 5182-5190. |
| [25] |
周余乐. 稻菜轮作对荣县蔬菜常见病虫害的影响[D]. 北京: 中国农业科学院, 2013. Zhou Y L. Effects of rice-vegetable rotation on common vegetable diseases and insect pest in Rong County[D]. Beijing: Chinese Academy of Agricultural Sciences, 2013. |
| [26] | 国家统计局. 中国统计年鉴-2019[M]. 北京: 中国统计出版社, 2019. |
| [27] |
唐清杰, 严小微, 孟卫东. 海南水稻生产现状分析及发展对策[J]. 杂交水稻, 2015, 30(1): 1-5. Tang Q J, Yan X W, Meng W D. Current status and developing strategy of rice production in Hainan[J]. Hybrid Rice, 2015, 30(1): 1-5. |
| [28] | Borchard N, Schirrmann M, Cayuela M L, et al. Biochar, soil and land-use interactions that reduce nitrate leaching and N2O emissions: a meta-analysis[J]. Science of the Total Environment, 2019, 651: 2354-2364. DOI:10.1016/j.scitotenv.2018.10.060 |
| [29] | Dong W J, Guo J, Xu L J, et al. Water regime-nitrogen fertilizer incorporation interaction: field study on methane and nitrous oxide emissions from a rice agroecosystem in Harbin, China[J]. Journal of Environmental Sciences, 2018, 64: 289-297. DOI:10.1016/j.jes.2017.06.007 |
| [30] |
刘杏认, 赵光昕, 张晴雯, 等. 生物炭对华北农田土壤N2O通量及相关功能基因丰度的影响[J]. 环境科学, 2018, 39(8): 3816-3825. Liu X R, Zhao G X, Zhang Q W, et al. Effects of biochar on nitrous oxide fluxes and the abundance of related functional genes from agriculture soil in the North China Plain[J]. Environmental Science, 2018, 39(8): 3816-3825. |
| [31] |
李香兰, 徐华, 蔡祖聪. 水分管理影响稻田氧化亚氮排放研究进展[J]. 土壤, 2009, 41(1): 1-7. Li X L, Xu H, Cai Z C. Effect of water management on nitrous oxide emission from rice paddy field: a review[J]. Soils, 2009, 41(1): 1-7. |
| [32] | Kuang W N, Gao X P, Tenuta M, et al. Relationship between soil profile accumulation and surface emission of N2O: effects of soil moisture and fertilizer nitrogen[J]. Biology and Fertility of Soils, 2019, 55(2): 97-107. DOI:10.1007/s00374-018-01337-4 |
| [33] | Lan T, Han Y, Roelcke M, et al. Processes leading to N2O and NO emissions from two different Chinese soils under different soil moisture contents[J]. Plant and Soil, 2013, 371(1-2): 611-627. DOI:10.1007/s11104-013-1721-1 |
| [34] | Di H J, Cameron K C, Podolyan A, et al. Effect of soil moisture status and a nitrification inhibitor, dicyandiamide, on ammonia oxidizer and denitrifier growth and nitrous oxide emissions in a grassland soil[J]. Soil Biology and Biochemistry, 2014, 73: 59-68. DOI:10.1016/j.soilbio.2014.02.011 |
| [35] | Lin Y X, Ding W X, Liu D Y, et al. Wheat straw-derived biochar amendment stimulated N2O emissions from rice paddy soils by regulating the amoA genes of ammonia-oxidizing bacteria[J]. Soil Biology and Biochemistry, 2017, 113: 89-98. DOI:10.1016/j.soilbio.2017.06.001 |
| [36] | Anderson F L, Cooper J A, Amador J A. Laboratory-scale evaluation of the effects of water-filled pore space on emissions of CO2, CH4, N2O, and N2 from soil-based wastewater treatment[J]. Water, Air, & Soil Pollution, 2019, 230. DOI:10.1007/s11270-019-4294-7 |
| [37] | Setyanto P, Pramono A, Adriany T A, et al. Alternate wetting and drying reduces methane emission from a rice paddy in Central Java, Indonesia without yield loss[J]. Soil Science and Plant Nutrition, 2018, 64(1): 23-30. DOI:10.1080/00380768.2017.1409600 |
| [38] | Zhang Q, Xiao J, Xue J H, et al. Quantifying the effects of biochar application on greenhouse gas emissions from agricultural soils: A global meta-analysis[J]. Sustainability, 2020, 12(8). DOI:10.3390/su12083436 |
| [39] | Wang J Y, Zhang M, Xiong Z Q, et al. Effects of biochar addition on N2O and CO2 emissions from two paddy soils[J]. Biology and Fertility of Soils, 2011, 47(8): 887-896. DOI:10.1007/s00374-011-0595-8 |
| [40] | Cayuela M L, Sánchez-Monedero M A, Roig A, et al. Biochar and denitrification in soils: when, how much and why does biochar reduce N2O emissions?[J]. Scientific Reports, 2013, 3. DOI:10.1038/srep01732 |
| [41] | Harter J, Guzman-Bustamante I, Kuehfuss S, et al. Gas entrapment and microbial N2O reduction reduce N2O emissions from a biochar-amended sandy clay loam soil[J]. Scientific Reports, 2016, 6. DOI:10.1038/srep39574 |
| [42] | Van Zwieten L, Kimber S, Morris S, et al. Influence of biochars on flux of N2O and CO2 from Ferrosol[J]. Soil Research, 2010, 48(7): 555-568. DOI:10.1071/SR10004 |
| [43] | Kuo Y L, Lee C H, Jien S H. Reduction of nutrient leaching potential in coarse-textured soil by using biochar[J]. Water, 2020, 12(7). DOI:10.3390/w12072012 |
| [44] | Wang Z Y, Zong H Y, Zheng H, et al. Reduced nitrification and abundance of ammonia-oxidizing bacteria in acidic soil amended with biochar[J]. Chemosphere, 2015, 138: 576-583. DOI:10.1016/j.chemosphere.2015.06.084 |
| [45] | Clough T J, Bertram J E, Ray J L, et al. Unweathered wood biochar impact on nitrous oxide emissions from a bovine-urine-amended pasture soil[J]. Soil Science Society of America Journal, 2010, 74(3): 852-860. DOI:10.2136/sssaj2009.0185 |
| [46] | Huang Y, Zou J W, Zheng X H, et al. Nitrous oxide emissions as influenced by amendment of plant residues with different C: N ratios[J]. Soil Biology and Biochemistry, 2004, 36(6): 973-981. DOI:10.1016/j.soilbio.2004.02.009 |
| [47] | Tarin M W K, Khaliq M A, Fan L L, et al. Divergent consequences of different biochar amendments on carbon dioxide (CO2) and nitrous oxide (N2O) emissions from the red soil[J]. Science of the Total Environment, 2021, 754. DOI:10.1016/j.scitotenv.2020.141935 |
| [48] | Chintala R, Mollinedo J, Schumacher T E, et al. Effect of biochar on chemical properties of acidic soil[J]. Archives of Agronomy and Soil Science, 2014, 60(3): 393-404. DOI:10.1080/03650340.2013.789870 |
| [49] |
王桂良, 崔振岭, 陈新平, 等. 南方稻田活性氮损失途径及其影响因素[J]. 应用生态学报, 2015, 26(8): 2337-2345. Wang G L, Cui Z L, Chen X P, et al. Reactive nitrogen loss pathways and their effective factors in paddy field in southern China[J]. Chinese Journal of Applied Ecology, 2015, 26(8): 2337-2345. |
| [50] |
田桃, 曾敏, 周航, 等. 水分管理模式与土壤Eh值对水稻Cd迁移与累积的影响[J]. 环境科学, 2017, 38(1): 343-351. Tian T, Zeng M, Zhou H, et al. Effects of different water managements and soil Eh on migration and accumulation of Cd in rice[J]. Environmental Science, 2017, 38(1): 343-351. |
| [51] | Joseph S D, Camps-Arbestain M, Lin Y, et al. An investigation into the reactions of biochar in soil[J]. Soil Research, 2010, 48(7): 501-515. DOI:10.1071/SR10009 |
| [52] | Yang F, Cao X D, Gao B, et al. Short-term effects of rice straw biochar on sorption, emission, and transformation of soil NH4+-N[J]. Environmental Science and Pollution Research, 2015, 22(12): 9184-9192. DOI:10.1007/s11356-014-4067-1 |
| [53] | Zheng H, Wang Z Y, Deng X, et al. Impacts of adding biochar on nitrogen retention and bioavailability in agricultural soil[J]. Geoderma, 2013, 206: 32-39. DOI:10.1016/j.geoderma.2013.04.018 |
| [54] |
陈晨, 许欣, 毕智超, 等. 生物炭和有机肥对菜地土壤N2O排放及硝化、反硝化微生物功能基因丰度的影响[J]. 环境科学学报, 2017, 37(5): 1912-1920. Chen C, Xu X, Bi Z C, et al. Effects of biochar and organic manure on N2O emissions and the functional gene abundance of nitrification and denitrification microbes under intensive vegetable production[J]. Acta Scientiae Circumstantiae, 2017, 37(5): 1912-1920. |
| [55] | Wu J, Brookes P C. The proportional mineralisation of microbial biomass and organic matter caused by air-drying and rewetting of a grassland soil[J]. Soil Biology and Biochemistry, 2005, 37(3): 507-515. DOI:10.1016/j.soilbio.2004.07.043 |
| [56] | Butterbach-Bahl K, Baggs E M, Dannenmann M, et al. Nitrous oxide emissions from soils: how well do we understand the processes and their controls?[J]. Philosophical Transactions of the Royal Society B: Biological Sciences, 2013, 368(1621). DOI:10.1098/rstb.2013.0122 |
| [57] | Thomson A J, Giannopoulos G, Pretty J, et al. Biological sources and sinks of nitrous oxide and strategies to mitigate emissions[J]. Philosophical Transactions of the Royal Society B: Biological Sciences, 2012, 367(1593): 1157-1168. DOI:10.1098/rstb.2011.0415 |
| [58] | Yanai Y, Toyota K, Okazaki M, et al. Effects of charcoal addition on N2O emissions from soil resulting from rewetting air-dried soil in short-term laboratory experiments[J]. Soil Science and Plant Nutrition, 2007, 53(2): 181-188. DOI:10.1111/j.1747-0765.2007.00123.x |
| [59] | Li S Q, Song L N, Jin Y G, et al. Linking N2O emission from biochar-amended composting process to the abundance of denitrify (nirK and nosZ) bacteria community[J]. AMB Express, 2016, 6. DOI:10.1186/s13568-016-0208-x |
| [60] | Wu Z, Zhang X, Dong Y B, et al. Microbial explanations for field-aged biochar mitigating greenhouse gas emissions during a rice-growing season[J]. Environmental Science and Pollution Research, 2018, 25(31): 31307-31317. DOI:10.1007/s11356-018-3112-x |