 2021, Vol. 42
2021, Vol. 42

2. 中国科学院南京地理与湖泊研究所, 湖泊与环境国家重点实验室, 南京 210008
 , PENG Kai2 , ZHANG Qing-ji2 , CAI Yong-jiu2 , ZHANG Yong-zhi1 , GONG Zhi-jun2
, PENG Kai2 , ZHANG Qing-ji2 , CAI Yong-jiu2 , ZHANG Yong-zhi1 , GONG Zhi-jun2 , XIANG Xian-ling1
, XIANG Xian-ling1
2. State Key Laboratory of Lake Science and Environment, Nanjing Institute of Geography and Limnology, Chinese Academy of Sciences, Nanjing 210008, China
洪泽湖是南水北调工程东线最大的调蓄型湖泊, 也是我国第四大淡水湖泊, 位于淮河中下游, 具有防洪、灌溉、调水、水产、水运和水电等多种功能[1].洪泽湖水质的好坏直接影响着苏北地区的用水质量与安全[2].随着洪泽湖周边经济的快速发展, 人类活动强度加剧, 导致洪泽湖的生态环境不断恶化(如富营养化加剧和水质下降等)[3].洪泽湖健康状态面临的主要问题为:蓄水范围内水域开发利用强度大, 湖泊自由水面率低, 水生植物覆盖度低; 水质较差, 总氮和总磷浓度偏高, 水体处于轻度富营养状态[4].
浮游动物是淡水生态系统中重要的生物类群, 在维持水生态系统结构和功能的完整性(如:是食物网重要的一环, 调节水体的自净能力等)等方面具有重要意义[5].浮游动物对环境变化反应敏感, 是水体中重要的指示类群[6].目前国内外对淡水浮游动物进行了大量研究, 表明不同水域浮游动物群落结构变化的主要影响因素不同, 包括非生物因素(如:水温和富营养化等)和生物因素(如:竞争和捕食等)[7~9].浮游动物作为介于高级消费者和初级生产者之间的中间营养层, 受上行效应和下行效应两种机制的高度影响.因此理化因素以及鱼类和浮游植物生物量均会对浮游动物群落结构产生影响[10~12], 在分析浮游动物群落结构时, 需要综合考虑各种因素对浮游动物的影响程度[13].一般而言, 浮游动物不同类群拥有不同的繁殖策略和扩散方式, 例如枝角类和轮虫的休眠卵可以被不同媒介(如风、鱼和鸟等)携带, 使浮游动物在不同的空间尺度上重新定殖[14].此外, 较小的浮游动物有更高的丰度, 也更容易转移[15, 16].
通常情况下, 受湖泊水温和浮游植物生物量的影响, 浮游动物密度呈现典型的夏高冬低的特征.对于洪泽湖来说, 虽然淮河流域降雨集中于夏季, 但受水位调蓄影响, 多年来洪泽湖水位变化均呈现出夏低冬高的特征, 与江淮流域多数湖泊均有显著不同, 同时洪泽湖也是典型的过水性湖泊, 其水流的变化对洪泽湖浮游动物群落结构影响较大.在洪泽湖这种特殊的湖泊水文条件下, 浮游动物的响应目前鲜有研究.因此洪泽湖可以作为一个很好的研究对象, 来分析水位波动对浮游动物的影响.同时研究洪泽湖浮游动物群落结构的时空变化, 对洪泽湖生态环境和水生资源的管理等也具有重要意义.因此本文通过对洪泽湖为期1 a的浮游动物野外调查, 分析洪泽湖浮游动物群落结构的时空变化, 并结合水环境因子数据, 使用典范对应分析(canonical correspondence analysis, CCA)对洪泽湖浮游动物群落结构与水环境因子的关系进行研究, 探究洪泽湖浮游动物群落结构变化的主要影响因子, 丰富洪泽湖的浮游动物研究, 以期为洪泽湖的生态环境保护与治理提供科学参考.
1 材料与方法 1.1 采样时间及采样点位洪泽湖湖泊面积较大, 平均水位在12.5 m时面积大约为1 576.9 km2.属于淮河中下游段的大型过水性湖泊, 换水周期较快, 出入湖泊的水流在南面蒋坝形成流速较快的过水通道.成子湖区位于北部, 多为中小入湖河流, 水体流动性差, 有水草分布, 溧河洼区位于西部, 该地区设为湿地保护区, 水深较浅, 同时设有大量围网, 中南部湖心区, 水流从西部淮河流入, 从东部入江河道排出, 水流较大, 流动性好, 难以见到水生植物, 水质主要受入湖河流淮河影响.根据洪泽湖具体情况, 考虑水质、围网养殖和水体流动性等情况, 在洪泽湖湖区设置10个采样点, 分成3个不同湖区(图 1), 调查时间设置在2017年3月~2018年2月, 于每月月底进行采样.

|
图 1 洪泽湖采样点分布示意 Fig. 1 Location of sampling sites in Hongze Lake |
水温(WT)、溶解氧(DO)和电导率(COND)等水质指标由YSI6600多参数水质监测仪现场测定; 总氮(TN)用碱性过硫酸钾消解紫外分光光度法测定(GB 11894- 89); 总磷(TP)用钼酸铵分光光度法测定(GB 11893- 89); 高锰酸盐指数用碱性高锰酸钾法测定(GB 11892- 89); 透明度(SD)用塞式透明度盘测定; 水深(WD)用水深仪测定; 叶绿素a(Chla)用热乙醇法测定; 水位(WL)数据来自于蒋坝水文站.
1.2.2 浮游动物浮游动物采样时, 枝角类、桡足类使用2.5 L有机玻璃采水器采集表层0.5 m下, 底层0.5 m以上水样各5 L, 经25号浮游生物网过滤浓缩, 放入加7%甲醛的小方瓶中固定, 回到实验室后定容到30 mL.分层采集1 L混合水样, 加入10~15 mL鲁哥氏液固定, 回到实验室静置48 h后吸取上清液浓缩定容至30 mL, 用于轮虫样品的鉴定与计数.在鉴定时, 充分摇匀浓缩的样品, 用移液器快速吸取1 mL样品置于1 mL计数板内, 在光学显微镜下全片计数.每个样品计数3片, 求平均值.浮游动物鉴定参照文献[17~19].陈非洲[20]的研究建议在分析淡水浮游动物时一般不包括原生动物.
1.3 数据分析大型浮游动物生物量的计算中, 枝角类的生物量根据体长-体重回归方程换算[21], 桡足类根据外形的几何形状换算为生物量[22]; 轮虫生物量根据其平均湿重进行换算[21].使用浮游动物优势度指数(Y)、Shannon-Wiener多样性指数(H′)、Margalef丰富度指数(D)和Pielou均匀度指数(J)对洪泽湖后生浮游动物群落结构进行分析评价.计算公式如下:
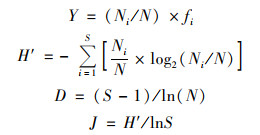
|
式中, Ni为样品中某种生物的总个体数; N为样品中各生物的总个体数; fi为某物种出现的频率; S为所有样品中的种属总数.
数据的前期处理在Excel中完成, 浮游动物年平均密度与生物量的空间分布图在ArcGIS 10.7中绘制, 对物种数据进行Hellinger转换以及环境因子数据标准化之后再进行典范对应分析(CCA), 使用R语言vegan包完成, 在进行CCA分析之前, 先对浮游动物密度数据进行去趋势对应分析(DCA), 得到最大梯度长度>4, 故选用典范对应分析(CCA).
2 结果与分析 2.1 洪泽湖环境因子的变化洪泽湖环境因子逐月变化如图 2所示, 水位逐月变化如图 3所示.各环境因子季节变化显著, 其中高锰酸盐指数、TP、Chla、WT和COND呈现出先上升后下降的变化趋势; TN、DO和WD呈现出先下降后上升的变化趋势; SD呈现波动变化, 并且夏季最低.受湖泊水位调蓄影响, 水位呈现夏季较低, 冬季较高的特征.TN月平均值波动范围为1.67~2.85 mg·L-1, 春冬季TN含量高于夏秋季.TP月平均值波动范围为0.06~0.12 mg·L-1, 夏秋季TP含量高于春冬季.

|
图 2 洪泽湖环境因子的逐月变化 Fig. 2 Monthly changes in environmental factors of Hongze Lake |

|
图 3 2017年3月至2018年2月洪泽湖水位逐月变化 Fig. 3 Monthly changes in water level of Hongze Lake from March 2017 to February 2018 |
洪泽湖的逐月调查的定量样品中, 共鉴定出浮游动物63种.其中轮虫8科12属24种, 枝角类6科13属19种, 桡足类10科14属20种.夏季洪泽湖浮游动物种类最为丰富, 共有44种, 冬季最少, 共有16种.夏季到冬季轮虫与枝角类具有明显的减少趋势.各季节均出现的轮虫种类有萼花臂尾轮虫(Brachuionus calyciflorus)、角突臂尾轮虫(Brachuionus angularis)、螺形龟甲轮虫(Keratella cochlearis)和针簇多肢轮虫(Polyarthra trigla), 枝角类的简弧象鼻溞(Bosmina coregoni), 桡足类的中华窄腹剑水蚤(Limnoithona sinensis)和无节幼体(Nauplii).就不同湖区而言, 过水区平均种类数最少, 成子湖与溧河洼平均种类数相近且较多.全年出现总种类数, 过水区最高, 成子湖次之, 溧河洼最少.
2.3 浮游动物密度与生物量的时空分布洪泽湖浮游动物年均密度为383.87 ind·L-1, 以轮虫为主(占比88.5%), 枝角类与桡足类的密度较低(占比11.5%).洪泽湖浮游动物年均生物量为1.36 mg·L-1, 以枝角类为主(占比72.5%), 轮虫与桡足类生物量较低(占比27.5%).浮游动物生物量与密度的时间分布如图 4所示, 浮游动物生物量最高月份出现在9月, 为7.68 mg·L-1; 生物量较低的为5、1和2月. 5~9月生物量逐渐升高, 9月至翌年2月逐渐降低.浮游动物密度最高月份出现在10月, 为1 736.9 ind·L-1, 较低的为5、1和2月. 3~5月和6~8月密度逐渐降低, 8~10月密度逐渐升高, 10月至翌年1月逐渐降到最低.浮游动物生物量与密度空间分布如图 5所示, 整体上成子湖与过水区东北部浮游动物生物量较高, 溧河洼与过水区生物量较低; 从浮游动物密度来看, 成子湖大于过水区、大于溧河洼.洪泽湖浮游动物生物量各类群中枝角类占据着绝对优势, 而轮虫占据着浮游动物密度的绝对优势, 桡足类的生物量与密度均较低.

|
图 4 洪泽湖浮游动物密度与生物量时间分布 Fig. 4 Temporal changes in zooplankton density and biomass in Hongze Lake |

|
图 5 洪泽湖浮游动物密度与生物量空间分布 Fig. 5 Spatial distribution of zooplankton density and biomass in Hongze Lake |
洪泽湖浮游动物优势种包括轮虫中的螺形龟甲轮虫与针簇多肢轮虫, 枝角类的简弧象鼻溞, 桡足类的中华窄腹剑水蚤与无节幼体.随着季节的变动, 优势种也发生着变化, 春季优势种有螺形龟甲轮虫、针簇多肢轮虫、梳状疣尾轮虫(Synchaeta pectinate)和简弧象鼻溞; 夏季优势种有矩形龟甲轮虫(Keratella quadrala)、曲腿龟甲轮虫、针簇多肢轮虫、简弧象鼻溞和角突网纹溞(Ceriodaphnia cornuta); 秋季优势种有螺形龟甲轮虫、针簇多肢轮虫和简弧象鼻溞; 冬季优势种有针簇多肢轮虫和无节幼体.优势种主导着浮游动物群落结构, 优势度最高的螺形龟甲轮虫、针簇多肢轮虫和简弧象鼻溞生物量与密度的时空变化, 与浮游动物生物量与密度的时空变化一致.
2.5 多样性指数洪泽湖Shannon-Wiener多样性指数(H′)、Margalef丰富度指数(D)和Pielou均匀度指数(J)时空变化如图 6所示, 时间上H′、D与J存在波动, H′表现出在7、8和9月出现峰值, 9月之后H′逐渐降低.D整体上, 3~7月逐渐上升, 7~12月逐渐降低, 12月至翌年2月逐渐升高. 5月D较为特殊, 最大值达到6.0.J整体上表现出3~5月逐渐升高, 5~6月之间存在下降, 6~10月呈上升趋势, 10月到翌年1月呈降低趋势, 1~2月有所上升.空间上整体表现出, 成子湖H′、D和J相对较高, 溧河洼次之, 过水区最低.

|
图 6 洪泽湖浮游动物多样性指数时空变化 Fig. 6 Spatio-temporal changes in zooplankton diversity indices in Hongze Lake |
CCA分析结果显示, 浮游动物群落结构的时空变化与WL、TN、TP、高锰酸盐指数、Chla、COND、WT、WD以及DO显著相关.前两轴分析结果共解释总方差的23.2%(P < 0.05).第一轴特征根为0.34, 第二轴特征根为0.29, 前两轴的解释率占总解释率的54.2%.根据排序结果, WL、TP和TN与第一轴呈显著负相关, WT和COND与第一轴呈显著正相关, 高锰酸盐指数和第二轴呈显著正相关, 其他环境因子呈显著负相关.环境因子中WL对浮游动物群落结构的影响最大, 其次是TN、WT和TP对浮游动物群落结构的影响也较大, WD、高锰酸盐指数、COND、Chla和DO对浮游动物群落结构影响相对较小.从图 7还可以看出浮游动物群落结构呈现出一定的季节变化, 其中秋、冬季的点位比较密集, 浮游动物群落结构较为稳定, 更多的受到水温、水质等的影响; 春、夏季的点位比较疏散, 浮游动物群落结构不稳定, 受多重因素影响.图 7不同湖区点位集中程度不同, 过水区分布得更为分散, 浮游动物群落结构差异较大, 成子湖和溧河洼分布得相对集中, 浮游动物群落结构差异较小.就物种与环境因子的关系而言, 角突网纹溞、多刺裸腹溞(Moina macrocopa)、广布中剑水蚤(Mesocyclops leuckarti)、矩形龟甲轮虫和蚤状溞(Daphnia pulex)密度的变化与WT、COND和Chla呈正相关, 与其他环境因子呈负相关.

|
P. trig表示针簇多肢轮虫、K. coch表示螺形龟甲轮虫、B. core表示简弧象鼻溞、N. Naup表示无节幼体、K. valg表示曲腿龟甲轮虫、B. caly表示萼花臂尾轮虫、K. quad表示矩形龟甲轮虫、B. angu表示角突臂尾轮虫、F. long表示长三肢轮虫、C. corn表示角突网纹溞、S. vetu表示老年低额溞、L. tetr表示中华窄腹剑水蚤、D. pule表示蚤状溞、F. corn表示角三肢轮虫、S. pect表示梳状疣尾轮虫、M. leuc表示广布中剑水蚤、S. dorr表示汤匙华哲水蚤、E. chan表示兴凯侧突水蚤、M. macr表示多刺裸腹溞、D. hank表示兴凯秀体溞 图 7 洪泽湖浮游动物与环境因子的典范对应分析 Fig. 7 Canonical correspondence analysis of zooplankton and environmental factors in Hongze Lake |
梅海鹏等[23]对洪泽湖近50年的水位变化进行了分析, 根据洪泽湖水位特征值的年际变化规律, 将其分为5个时期, 分别分析各个时期年内变化过程.将洪泽湖水位特征值在年内变化分为3个阶段, 蓄水期(10月至翌年4月)、泄水期(5~6月)和涨水期(7~9月).这与本次调查期间洪泽湖水位变化较为相似.在7月初至8月初涨水期存在较大的波动.湖泊透明度主要受水体中悬浮颗粒物和藻类浓度的影响, 但洪泽湖作为大型浅水过水性湖泊, 水体中悬浮颗粒物浓度较大, 是影响透明度的主要因子.尤其在水位调控期间, 水位较低, 受风浪扰动作用更为强烈, 水体透明度会显著下降.洪泽湖水体营养盐以及叶绿素呈现显著的季节变化, 例如总氮夏低冬高、总磷夏高冬低和叶绿素夏高冬低等特征, 这与多数长江中下游地区湖泊较为相似, 季节效应明显.前人也对出现这种变化的原因开展了诸多研究, 例如总氮夏季低可能是由于夏季温度较高, 反硝化作用强烈导致水体氮的去除[24], 总磷夏季高可能是由于藻类的奢侈消费吸收过量的磷死亡后向水体中释放[25, 26]以及夏季浮游植物光合作用导致pH增加促进磷的释放[27, 28], 此外温度也是促进夏季磷内源释放加快的主要因子之一, 同时雨季的充沛降雨也会导致流域面源磷的输入[29].对于洪泽湖而言, 虽然其营养盐指标大体上也呈现这种变化特征, 但与之不同的是洪泽湖营养盐及叶绿素各指标在5~8月常出现波动.由于5~8月是重点水位调控阶段, 结合洪泽湖过水性湖泊的特征, 这可能是由于上游来水水质不稳定, 同时水位调控期间湖泊水容量较低, 湖泊对上游污染物输入的缓冲能力下降, 水质更容易受上游来水影响.因此洪泽湖水质的年内变化受季节变化和水位调控共同影响, 其中水位调控容易造成湖泊水质波动变化.
3.2 洪泽湖浮游动物的演变据文献[30], 1974~1975年间洪泽湖有浮游动物70种, 其中轮虫37种, 枝角类19种, 桡足类14种.优势种有轮虫中的萼花臂尾轮虫、台式合甲轮虫(Diplois daviesiae)、螺形龟甲轮虫、矩形龟甲轮虫、曲腿龟甲轮虫、晶囊轮属(Asplanchna sp.)、盖刺异尾轮虫(Trichocerca capucina)、奇异巨腕轮虫(Pedalia mira)和独角聚花轮虫(Conochilus unicornis); 枝角类的短尾秀体溞(Diaphanosoma brachyurum)、僧帽溞(Daphnia cucullate)、长刺溞(Daphnia longispina)、角突网纹溞、棘体网纹溞(Ceriodaphnia setosa)、微型裸腹溞(Moina micrura)、长额象鼻溞(Bosmina longirostris)、简弧象鼻溞、颈沟基合溞(Bosminopsis deitersi)和圆形盘肠溞(Chydorus sphaericus); 桡足类的汤匙华哲水蚤(Sinocalanus dorrii)、指状许水蚤(Schmackeria inopinus)、球状许水蚤(Schmackeria forbesi)、中华窄腹剑水蚤、广布中剑水蚤和中国剑水蚤(Thermocyclops taihokuensis)等.轮虫密度为166.4 ind·L-1, 生物量为0.199 4 mg·L-1; 枝角类密度为16.6 ind·L-1, 生物量为0.401 5 mg·L-1; 桡足类密度为36.6 ind·L-1, 生物量为0.581 4 mg·L-1.都雪等[31]在2010年5月至2011年2月对洪泽湖浮游动物进行季度采样, 共观察到浮游动物53种, 其中轮虫34种, 枝角类9种, 桡足类10种, 主要优势种为螺形龟甲轮虫、前节晶囊轮虫、长肢多肢轮虫、萼花臂尾轮虫和曲腿龟甲轮虫; 枝角类中的长额象鼻溞、透明溞(Daphnia hyaline); 桡足类中的汤匙华哲水蚤和中华窄腹剑水蚤.轮虫年平均密度和生物量分别为323.6 ind·L-1和0.388 mg·L-1.刘孝珍[32]在2014年的5月和8月的两次调查中共检出浮游动物28种, 常见种类有:枝角类的长额象鼻溞, 桡足类的汤匙华哲水蚤等.浮游动物密度在293~637 ind·L-1, 生物量在1.30~4.51 mg·L-1.临淮、新开河和剪草沟的密度与生物量较大.本次调查共鉴定出浮游动物63种, 其中轮虫24种, 枝角类19种, 桡足类20种.优势种为螺形龟甲轮虫、针簇多肢轮虫和简弧象鼻溞.年均密度为383.87 ind·L-1, 年均生物量为1.36 mg·L-1.综上可知多年来洪泽湖浮游动物密度与生物量变化不大, 这可能是由于洪泽湖是过水性湖泊, 流速较大, 导致浮游动物密度与生物量较低.多年来优势种变化明显, 优势种种类数大幅度减少, 清洁物种减少耐污种占据主导地位, 如独角聚花轮虫、异尾轮虫等是贫营养水体的指示物种[33], 螺形龟甲轮虫、针簇多肢轮虫和简弧象鼻溞等为富营养水体的指示物种[34].因此富营养化导致的浮游动物群落结构趋于简单化是洪泽湖浮游动物群落结构演变的主要驱动因子.
3.3 与临近的大型淡水湖泊浮游动物群落结构的比较东部平原大型淡水湖泊有巢湖、太湖和南四湖.南四湖与洪泽湖同属于南水北调东线工程的调蓄型湖泊, 巢湖、洪泽湖和太湖均属于江淮流域大型浅水湖泊, 其浮游动物群落结构如表 1所示[35~38].优势种属方面, 4个湖泊均以小型浮游动物为主; 浮游动物种类数、多样性指数、密度和生物量方面南四湖最高, 洪泽湖最低.洪泽湖浮游动物群落结构不如其他3个湖泊丰富.与南四湖相比, 总氮、水温、pH和电导率是影响南四湖浮游动物群落结构的主要因素, 南四湖水生植被分布较多, 生境多样性较高, 能够容许更多的浮游动物生存[37].与太湖相比, 水温、营养盐和浮游植物等直接影响太湖浮游动物群落结构, 透明度和风力等间接影响着浮游动物群落结构[39, 40].一方面太湖水体处于富营养水平, 能为浮游动物提供充足的食物, 另一方面太湖换水周期长, 水体流动性差适宜浮游动物的生存.与巢湖相比, 水温和浮游植物等是影响巢湖浮游动物群落结构的主要因素[36], 一方面巢湖水体处于富营养水平, 蓝藻的暴发导致小型枝角类与轮虫成为优势种[41], 另一方面巢湖换水周期长, 水体流动性差适宜浮游动物的生存.洪泽湖是换水周期短的中度富营养化的湖泊, 对于洪泽湖来说, 富营养化水平和换水周期共同主导着洪泽湖的浮游动物群落结构和多样性水平.
|
|
表 1 洪泽湖及周边大型淡水湖泊的浮游动物群落结构 Table 1 Zooplankton community structure in Hongze Lake and other large freshwater lakes |
3.4 洪泽湖浮游动物群落动态与环境因子的关系
水温被认为是影响浮游动物生长、发育和物种组成的重要因子, 也是影响浮游动物种类季节演替的主要驱动因子[42].浮游动物群落结构呈现显著的季节变化, 如多样性和种类数呈现明显的夏高冬低, 密度与生物量呈现出秋高冬低的特征.洪泽湖的浮游动物多为广温性种, 如臂尾轮属和龟甲轮属等[43], 较少地出现暖温性种, 如异尾轮属和角突网纹溞等[43].洪泽湖浮游动物年内变化显著, 在春季与冬季浮游动物种类最少, 群落结构最为简单. CCA结果表明, 春季与冬季浮游动物点位较为密集, 且主要受到水温、水位、总氮、叶绿素和总磷等的主导.因此在冬季与春季浮游动物群落结构的变化, 可能是因为水温较低, 营养物质较少, 鱼类的捕食, 以及水流扰动等因素对浮游动物的生长、发育和繁殖不利造成的.
浮游动物不同类群对栖息地环境的变化表现出不同的响应.由于体型小, 浮游动物往往受到栖息地的水文条件的影响[44].其中影响最为显著的是水流.刘翊竣等[45]的研究指出, 洪泽湖的溧河洼区和成子湖区流场结构主要由风场决定, 而南部湖区和东部湖区流场结构由风场和吞吐流共同决定, 且以吞吐流为主.因此, 洪泽湖浮游动物空间分布不均匀的原因是浮游动物运动能力较弱, 主要是被动转移.在过水区洪泽湖湖体流速较大不易于浮游动物的生长繁殖, 密度与生物量较低, 但是能带来更多的浮游动物类群.在相对静止的成子湖区与溧河洼湖区, 浮游动物生物量与密度较高. CCA结果显示浮游动物的季节变化显著, 相对于空间变化来说, 季节变化更大, 导致空间变化不显著.
CCA结果显示夏季点位最为分散, 浮游动物群落结构变化最明显, 并且这种差异主要受温度与水位驱动.一方面温度驱使的浮游动物季节变化导致夏季洪泽湖浮游动物群落发生演替, 从而造成夏季点位与其他季节产生差异.另一方面, 水位的波动意味着流速的不稳定, 时快时慢, 极不利于浮游动物生长繁殖, 加上水质波动变化, 因此造成夏季浮游动物群落结构变化较大, 体现在CCA排序图上的就是点位分散.与非参数分析不同的是, CCA分析属于参数分析, 其浮游动物密度的数量差异就会体现在排序结果中.
秋冬季点位较为密集, 水位相对稳定, 调水对浮游动物的影响较小, 此时水温, 营养盐及叶绿素各指标对浮游动物的影响较大, 这与其他湖泊较为相似[35, 46, 47].笔者注意到3~5月浮游动物密度在逐渐降低, 主要表现为轮虫的减少, 这与很多湖泊不同[7], 而洪泽湖3~5月Chla浓度降低, 此期间浮游动物密度降低可能主要是因为食物的匮乏.
4 结论(1) 本次调查共鉴定出浮游动物63种, 其中轮虫有24种, 枝角类19种和桡足类20种.年均密度为383.87 ind·L-1, 年均生物量为1.36 mg·L-1.洪泽湖浮游动物群落结构时空变化较大, 时间上夏季浮游动物波动较大, 秋季浮游动物密度与生物量均达到最大值, 冬季浮游动物群落结构最为简单; 空间上成子湖和溧河洼浮游动物较为丰富, 差异较小, 过水区浮游动物较少且变化较大.
(2) 洪泽湖浮游动物群落结构的时空变化主要由水位、总磷、水温和总氮等因素共同决定.不同时间浮游动物群落结构变化的主导因素不同, 夏季主导因素是水位与水温; 秋冬季主导因素是水温、营养盐及叶绿素等.空间上不同湖区点位集中程度不同, 过水区分布得更为分散, 浮游动物群落结构差异较大主要由水位变化主导, 成子湖和溧河洼分布得相对集中, 浮游动物群落结构差异较小.
(3) 水位波动对浮游动物群落结构的变化影响最大, 表现在水位波动会直接影响浮游动物群落结构, 也可能通过影响水质, 进而间接影响浮游动物群落结构.
| [1] |
杨广利, 韩爱民, 刘轶琨, 等. 洪泽湖富营养化与环境理化因子间的关系[J]. 环境监测管理与技术, 2003, 15(2): 17-20. Yang G L, Han A M, Liu Y K, et al. The relationship of eutrophication of the lake of Hongze and environmental physical-chemical factors[J]. Administration and Technique of Environmental Monitoring, 2003, 15(2): 17-20. DOI:10.3969/j.issn.1006-2009.2003.02.008 |
| [2] |
李为, 都雪, 林明利, 等. 基于PCA和SOM网络的洪泽湖水质时空变化特征分析[J]. 长江流域资源与环境, 2013, 22(12): 1593-1601. Li W, Du X, Lin M L, et al. Spatial-temporal variation characteristics of water quality in Hongze Lake based on PCA and SOM[J]. Resources and Environment in the Yangtze Basin, 2013, 22(12): 1593-1601. |
| [3] |
张超文, 张堂林, 朱挺兵, 等. 洪泽湖大型底栖动物群落结构及其与环境因子的关系[J]. 水生态学杂志, 2012, 33(3): 27-33. Zhang C W, Zhang T L, Zhu T B, et al. Community structure of macrozoobenthos and its relationship with environmental factors in Lake Hongze[J]. Journal of Hydroecology, 2012, 33(3): 27-33. |
| [4] |
蔡永久, 张祯, 唐荣桂, 等. 洪泽湖生态系统健康状况评价和保护[J]. 江苏水利, 2020(7): 1-7, 13. Cai Y J, Zhang Z, Tang R G, et al. Evaluation and protection of ecosystem health in Hongze Lake[J]. Jiangsu Water Resources, 2020(7): 1-7, 13. |
| [5] |
胡梦红, 杨丽丽, 刘其根. 竞争捕食作用对千岛湖浮游动物群落结构的影响[J]. 湖泊科学, 2014, 26(5): 751-758. Hu M H, Yang L L, Liu Q G. Effects of predation and competition on zooplankton community structure in Lake Qiandao[J]. Journal of Lake Sciences, 2014, 26(5): 751-758. |
| [6] | Duggan I C, Green J D, Shiel R J. Distribution of rotifers in North Island, New Zealand, and their potential use as bioindicators of lake trophic state[J]. Hydrobiologia, 2001, 446-447(1): 155-164. DOI:10.1023%2FA%3A1017503407240 |
| [7] |
杨佳, 周健, 秦伯强, 等. 太湖梅梁湾浮游动物群落结构长期变化特征(1997~2017年)[J]. 环境科学, 2020, 41(3): 1246-1255. Yang J, Zhou J, Qin B Q, et al. Long-term variation characteristics of zooplankton community structure in Meiliang Bay, Lake Taihu[J]. Environmental Science, 2020, 41(3): 1246-1255. DOI:10.3969/j.issn.1000-6923.2020.03.036 |
| [8] | Ermolaeva N I, Zarubina E Y, Bazhenova O P, et al. Influence of abiotic and trophic factors on the daily horizontal migration of zooplankton in the littoral zone of the Novosibirsk Reservoir[J]. Inland Water Biology, 2019, 12(4): 418-427. DOI:10.1134/S1995082919030052 |
| [9] |
温超男, 黄蔚, 陈开宁, 等. 太湖滨岸带浮游动物群落结构特征与环境因子的典范对应分析[J]. 水生态学杂志, 2020, 41(2): 36-44. Wen C N, Huang W, Chen K N, et al. Canonical correspondence analysis between zooplankton community structure and environmental factors in the littoral zone of Taihu Lake[J]. Journal of Hydroecology, 2020, 41(2): 36-44. |
| [10] | Havel J E, Shurin J B. Mechanisms, effects, and scales of dispersal in freshwater zooplankton[J]. Limnology and Oceanography, 2004, 49(S4): 1229-1238. |
| [11] | Schamp B S, Arnott S E, Joslin K L. Dispersal strength influences zooplankton co-occurrence patterns in experimental mesocosms[J]. Ecology, 2015, 96(4): 1074-1083. DOI:10.1890/14-1128.1 |
| [12] | Xiong W, Ni P, Chen Y Y, et al. Zooplankton community structure along a pollution gradient at fine geographical scales in river ecosystems: the importance of species sorting over dispersal[J]. Molecular Ecology, 2017, 26(16): 4351-4360. DOI:10.1111/mec.14199 |
| [13] | Zhang J, Xie P, Tao M, et al. The impact of fish predation and cyanobacteria on zooplankton size structure in 96 subtropical lakes[J]. PLoS One, 2018, 8(10). DOI:10.1371/journal.pone.0076378 |
| [14] | Pinceel T, Vanschoenwinkel B, Brendonck L. Flexible dispersal dimorphism in zooplankton resting eggs: an example of repeated phenotypic coin flipping?[J]. Biological Journal of the Linnean Society, 2013, 110(4): 749-756. DOI:10.1111/bij.12161 |
| [15] | Jungblut A. Biogeography of microscopic organisms: is everything small everywhere? Edited by Diego Fontaneto[J]. Cambridge: Cambridge University Press, 2011, 176(4): 582-583. |
| [16] | Dias J D, Simöes N R, Meerhoff M, et al. Hydrological dynamics drives zooplankton metacommunity structure in a Neotropical floodplain[J]. Hydrobiologia, 2016, 781(1): 109-125. DOI:10.1007/s10750-016-2827-2 |
| [17] | 王家楫. 中国淡水轮虫志[M]. 北京: 科学出版社, 1961. |
| [18] | 蒋燮治, 堵南山. 中国动物志—节肢动物门甲壳纲淡水枝角类[M]. 北京: 科学出版社, 1979. |
| [19] | 中国科学院动物研究所甲壳动物研究组. 中国动物志—节肢动物门甲壳纲淡水桡足类[M]. 北京: 科学出版社, 1979. |
| [20] |
陈非洲. [读者来信]关于淡水浮游动物和原生动物分开研究的建议[J]. 湖泊科学, 2016, 28(3): C3. Chen F Z. Suggestion on compositions of freshwater zooplankton vs photozoa[J]. Journal of Lake Sciences, 2016, 28(3). |
| [21] | 赵文. 水生生物学[M]. 北京: 中国农业出版社, 2005. |
| [22] |
陈雪梅. 淡水桡足类生物量的测算[J]. 水生生物学集刊, 1981, 7(3): 397-408. Chen X M. Biomass calculation of freshwater copepoda[J]. Acta Hydrobiologica Sinica, 1981, 7(3): 397-408. |
| [23] |
梅海鹏, 王振龙, 刘猛, 等. 洪泽湖近50 a特征水位变化规律及影响因素[J]. 长江科学院院报, 2021, 38(1): 35-40. Mei H P, Wang Z L, Liu M, et al. Characteristic water levels of Hongze Lake in the Past Five Decades: variation rules and influencing factors[J]. Journal of Yangtze River Scientific Research Institute, 2021, 38(1): 35-40. |
| [24] | Paerl H W, Xu H, McCarthy M J, et al. Controlling harmful cyanobacterial blooms in a hyper-eutrophic lake (Lake Taihu, China): The need for a dual nutrient (N & P) management strategy[J]. Water Research, 2011, 45(5): 1973-1983. DOI:10.1016/j.watres.2010.09.018 |
| [25] | Elrifi I R, Turpin D H. Steady-state luxury consumption and the concept of optimum nutrient ratios: a study with phosphate and nitrate limited Selenastrum minutum (Chlorophyta)[J]. Journal of Phycology, 1985, 21(4): 592-602. |
| [26] | Ahn C Y, Chung A S, Oh H M. Rainfall, phycocyanin, and N: P ratios related to cyanobacterial blooms in a Korean large reservoir[J]. Hydrobiologia, 2002, 474(1-3): 117-124. |
| [27] | Boers P C M. The influence of pH on phosphate release from lake sediments[J]. Water Research, 1991, 25(3): 309-311. DOI:10.1016/0043-1354(91)90010-N |
| [28] | Xie L Q, Xie P, Tang H J. Enhancement of dissolved phosphorus release from sediment to lake water by Microcystis blooms-an enclosure experiment in a hyper-eutrophic, subtropical Chinese lake[J]. Environmental Pollution, 2003, 122(3): 391-399. DOI:10.1016/S0269-7491(02)00305-6 |
| [29] | Yang Z, Zhang M, Shi X L, et al. Nutrient reduction magnifies the impact of extreme weather on cyanobacterial bloom formation in large shallow Lake Taihu (China)[J]. Water Research, 2016, 103: 302-310. DOI:10.1016/j.watres.2016.07.047 |
| [30] | 王苏民, 窦鸿身. 中国湖泊志[M]. 北京: 科学出版社, 1998: 389-390. |
| [31] |
都雪, 王齐东, 张超文, 等. 洪泽湖轮虫群落结构及其与环境因子的关系[J]. 湖泊科学, 2014, 26(2): 269-276. Du X, Wang Q D, Zhang C W, et al. Community structure of rotifers in relation to environmental factors in Lake Hongze[J]. Journal of Lake Sciences, 2014, 26(2): 269-276. |
| [32] |
刘孝珍. 洪泽湖渔业资源现状、问题及对策[D]. 南京: 南京农业大学, 2015. Liu X Z. Current situation, problems and countermeasures of fishery resources in Hongze Lake[D]. Nanjing: Nanjing Agricultural University, 2015. |
| [33] |
吴利, 李源玲, 陈延松. 淮河干流浮游动物群落结构特征[J]. 湖泊科学, 2015, 27(5): 932-940. Wu L, Li Y L, Chen Y S. Characteristics of community structures of zooplankton in the mainstream of Huaihe River[J]. Journal of Lake Sciences, 2015, 27(5): 932-940. |
| [34] |
李强, 田华, 姜民, 等. 淀山湖浮游动物群落结构特征及其影响因子[J]. 水生态学杂志, 2015, 36(4): 69-77. Li Q, Tian H, Jiang M, et al. Community structure of zooplankton and influencing factors in Dianshan Lake of Shanghai[J]. Journal of Hydroecology, 2015, 36(4): 69-77. |
| [35] |
周义道. 太湖浮游动物群落结构及环境相关性研究[D]. 上海: 上海师范大学, 2019. Zhou Y D. Zooplankton community structure and its relation to environmental factors in Taihu Lake[D]. Shanghai: Shanghai Normal University, 2019. |
| [36] |
李静, 崔凯, 卢文轩, 等. 春季和夏季巢湖浮游生物群落组成及其动态分析[J]. 水生生物学报, 2015, 39(1): 185-192. Li J, Cui K, Lu W X, et al. Community dynamics of spring-summer plankton in Lake Chaohu[J]. Acta Hydrobiologica Sinica, 2015, 39(1): 185-192. |
| [37] |
李怀国, 杨长明, 王育来. 巢湖水质现状及浮游生物群落结构特征[J]. 安徽农业科学, 2017, 45(22): 13-16. Li H G, Yang C M, Wang Y L. Water quality and community structure characteristics of plankton in Chaohu Lake[J]. Journal of Anhui Agricultural Sciences, 2017, 45(22): 13-16. DOI:10.3969/j.issn.0517-6611.2017.22.004 |
| [38] |
陈磊, 高东泉, 舒凤月, 等. 南四湖浮游动物群落结构特征及其与环境因子的关系[J]. 动物学杂志, 2016, 51(1): 113-120. Chen L, Gao D Q, Shu F Y, et al. Zooplankton community structure and its relationship with environmental factors in Nasi Lake[J]. Chinese Journal of Zoology, 2016, 51(1): 113-120. |
| [39] | Jeppesen E, Jensen J P, Søndergaard M, et al. Trophic structure, species richness and biodiversity in Danish lakes: changes along a phosphorus gradient[J]. Freshwater Biology, 2000, 45(2): 201-218. DOI:10.1046/j.1365-2427.2000.00675.x |
| [40] |
李娟, 温周瑞, 李庚辰, 等. 太湖梅梁湾和贡湖湾浮游甲壳动物群落结构及其与环境因子的相互关系[J]. 长江流域资源与环境, 2014, 23(S1): 81-90. Li J, Wen Z R, Li G C, et al. Relationship between crustacean zooplankton community structure and aquatic environmental factors of Meiliang and Gonghu bay in Lake Taihu[J]. Resources and Environment in the Yangtze Basin, 2014, 23(S1): 81-90. |
| [41] | Geng H, Xie P, Deng D G. The rotifer assemblage in a shallow, eutrophic Chinese lake and its relationships with cyanobacterial blooms and crustacean zooplankton[J]. Journal of Freshwater Ecology, 2005, 20(1): 93-100. DOI:10.1080/02705060.2005.9664941 |
| [42] | Marques S C, Azeiteiro U M, Marques J C, et al. Zooplankton and ichthyoplankton communities in a temperate estuary: spatial and temporal patterns[J]. Journal of Plankton Research, 2006, 28(3): 297-312. DOI:10.1093/plankt/fbi126 |
| [43] |
陈立婧, 顾静, 彭自然, 等. 上海崇明岛明珠湖轮虫群落结构[J]. 应用生态学报, 2009, 20(12): 3057-3062. Chen L J, Gu J, Peng Z R, et al. Community structure of rotifer in Mingzhu Lake of Chongming Island, Shanghai[J]. Chinese Journal of Applied Ecology, 2009, 20(12): 3057-3062. |
| [44] | Brucet S, Boix D, Quintana X D, et al. Factors influencing zooplankton size structure at contrasting temperatures in coastal shallow lakes: Implications for effects of climate change[J]. Limnology and Oceanography, 2010, 55(4): 1697-1711. DOI:10.4319/lo.2010.55.4.1697 |
| [45] |
刘翊竣, 徐国宾. 风场和吞吐流对洪泽湖流场的影响分析[J]. 水资源与水工程学报, 2020, 31(2): 174-178, 184. Liu Y J, Xu G B. Effects of wind field and throughput flow on the flow field of Hongze Lake[J]. Journal of Water Resources and Water Engineering, 2020, 31(2): 174-178, 184. |
| [46] |
杜彩丽, 杨丽, 赵诣, 等. 淀山湖浮游动物群落时空分布特征及其与环境因子的关系[J]. 环境科学, 2019, 40(10): 4513-4522. Du C L, Yang L, Zhao Y, et al. Temporal and spatial variation of zooplankton community structure and its relationship with environmental factors in Dianshan Lake, Shanghai[J]. Environmental Science, 2019, 40(10): 4513-4522. |
| [47] |
陈佳琪, 赵坤, 曹玥, 等. 鄱阳湖浮游动物群落结构及其与环境因子的关系[J]. 生态学报, 2020, 40(18): 6644-6658. Chen J Q, Zhao K, Cao Y, et al. Zooplankton community structure and its relationship with environmental factors in Poyang Lake[J]. Acta Ecologica Sinica, 2020, 40(18): 6644-6658. |